1Department of Crop Science, Chungbuk National University, Cheongju 361-763, Republic of Korea
2Department of Horticulture, Hankyong National University, Ansung 456-749, Republic of Korea
3Department of Horticulture, Sunchon National University, Sunchon 540-742, Republic of Korea
4Chuncheon Substation, National Institute of Crop Science, Chuncheon 200-940, Republic of Korea
Copyright © 2013 The Korean Society of Breeding Science
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
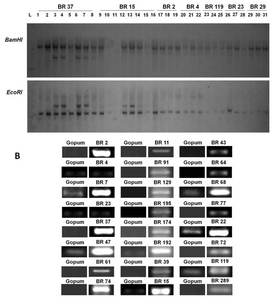

| Generation* | Confirmed no. of gene inserts | No. of lines |
|---|---|---|
| T1 | 137 | 470 |
| T2 | 88 | 408 |
| T3 | 14 | 46 |
| T4 | 11 | 226 |
|
|
||
| Total | 250 | 1,150 |
| Code | Gene Function | Cold stress scorea | Salinity stress scoreb | Observed phenotypec |
|---|---|---|---|---|
| BR 2 | cysteine protease 1 | 5.7 | 9.0 | more green leaves |
| BR 4 | ADF2 (Actin depolymerizing factor 2) | 5.0 | 9.0 | more number of tiller |
| BR 5 | PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 | 3.0 | 9.0 | more green leaves |
| BR 6 | Zea mays clone 93807 mRNA sequence | 5.0 | 9.0 | more green leaves |
| BR 7 | Dead box RNA helicase RH15 - like protein | 3.0 | 3.0 | same phenotype |
| BR 9 | PEC-1 mRNA for Lipid transfer protein | 5.0 | 9.0 | more green leaves |
| BR 12 | Rac GTPase activator | 5.0 | 9.0 | same phenotype |
| BR 14 | CBL-interacting protein kinase 3 | 5.2 | 5.5 | less green leaves |
| BR 17 | mRNA for aluminum-induced | 5.0 | 9.0 | less green leaves |
| BR 23 | endoplasmic reticulum auxin-binding protein 2 | 9.0 | 7.0 | less green leaves |
| BR 26 | Brassica rapa subsp. pekinensis clone KBrH004A06, complete sequence | 5.3 | 9.0 | same phenotype |
| BR 29 | Phospholipase C | 5.2 | 7.0 | less green leaves |
| BR 33 | ABC transporter-like protein | 9.0 | 9.0 | less green leaves |
| BR 37 | CBL-interacting protein kinase 1 | 5.4 | 3.0 | less green leaves, less plant height |
| BR 39 | carbohydrate binding | 4.3 | 9.0 | less green leaves |
| BR 40 | protein phosphatase 2C, putative/PP2C, | 4.7 | 3.0 | less green leaves, shorter culm length |
| BR 46 | plasma membrane intrinsic protein 2 (PIP2) | 5.0 | 9.0 | less green leaves, more taller |
| BR 48 | senescence-associated cysteine protease (CP1) | 4.8 | 5.0 | more number of tiller |
| BR 49 | polypeptide of photosystem II reaction center | 9.0 | 9.0 | less green leaves |
| BR 50 | chlorophyll a/b binding protein | 9.0 | less green leaves | |
| BR 51 | methyl-CpG-binding domain 4 (MBD4) | 9.0 | 9.0 | less green leaves, longer panicle length |
| BR 57 | putative protein transport protein SEC61 alpha subunit | 5.0 | 7.0 | less green leaves |
| BR 60 | Brassica rapa subsp. pekinensis clone KBrB042D24, complete sequence | 5.0 | 9.0 | less green leaves |
| BR 61 | rubredoxin family protein | 6.0 | 9.0 | less green leaves |
| BR 64 | GDSL esterase/lipase APG | 5.0 | 9.0 | less green leaves, longer panicle length, Shorter culm length |
| BR 72 | WSCP1 mRNA for water-soluble chlorophyll protein precursor | 5.0 | 9.0 | same phenotype |
| BR 73 | unknown protein | 6.0 | 9.0 | less green leaves |
| BR 74 | haloacid dehalogenase-like hydrolase family protein | 5.0 | 3.0 | less green leaves, more number of tiller |
| BR81 | regulatory subunit of protein kinase CK2 | 5.0 | 7.0 | less green leaves, shorter culm length |
| BR 84 | ATCTIMC (Cytosolic triose phosphate isomerase) | 5.0 | 9.0 | shorter culm length |
| BR 89 | Brassica rapa subsp. pekinensis clone KBrH004P05, complete sequence | 5.7 | 9.0 | same phenotype |
| BR 91 | unknown protein | 5.2 | 3.0 | less green leaves, longer culm length |
| BR 92 | 5-enolpyruvylshikimate-3-phosphate synthase (EPSP) | 5.0 | 9.0 | same phenotype |
| BR 97 | G6PD1 (glucose-6-phosphate dehydrogenase 1) | 5.1 | 9.0 | same phenotype |
| BR 99 | 40S ribosomal protein S7 (RPS7C) | 5.0 | 9.0 | same phenotype |
| BR 104 | protein binding/zinc ion binding | 5.0 | 9.0 | same phenotype |
| BR 105 | Lhcb6 protein mRNA | 5.0 | 9.0 | more number of tiller |
| BR 111 | protein kinase family protein | 5.0 | 9.0 | same phenotype |
| BR 112 | Arabidopsis thaliana AT5g47540/MNJ7_13 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 113 | Brassica rapa subsp. pekinensis clone KBrB052L10, complete sequence | 5.8 | 9.0 | same phenotype |
| BR 114 | Zea mays clone 13119 mRNA sequence | 5.0 | 9.0 | less plant height |
| BR 115 | Arabidopsis thaliana At5g25810 mRNA, complete cds | 3.0 | 9.0 | same phenotype |
| BR 116 | Unknown | 5.0 | 9.0 | same phenotype |
| BR 118 | Vitis vinifera, whole genome shotgun sequence, contig VV78X164547.5, clone ENTAV 115 | 6.0 | 9.0 | same phenotype |
| BR 119 | unknown protein | 3.0 | 3.0 | shorter plant height |
| BR 120 | Zea mays clone 14950 mRNA sequence | 5.0 | 6.2 | same phenotype |
| BR 122 | DNAJ heat shock N-terminal domain-containing protein | 5.0 | 9.0 | same phenotype |
| BR 124 | putative thioredoxin reductase | 4.8 | 9.0 | same phenotype |
| BR 125 | Brassica rapa subsp. pekinensis clone KBrS004A14, complete sequence | 5.0 | 9.0 | less plant height |
| BR 128 | Zea mays clone 9139 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 129 | dimethylmenaquinone methyltransferase family protein | 5.0 | 9.0 | same phenotype |
| BR133 | prephenate dehydratase family protein | 5.0 | 9.0 | same phenotype |
| BR139 | 231 CLA1-like protein mRNA | 5.1 | 9.0 | same phenotype |
| BR147 | 231 CLA1-like protein mRNA | 5.0 | 9.0 | same phenotype |
| BR148 | protein kinase family protein/C-type lectin domain-containing protein | 5.0 | 5.2 | same phenotype |
| BR 149 | XTR4 (xyloglucan endotransglycolase 4) | 5.0 | 9.0 | same phenotype |
| BR 151 | clathrin adaptor complexes medium subunit-related | 5.0 | 9.0 | same phenotype |
| BR 152 | unknown mRNA | 5.0 | 9.0 | same phenotype |
| BR 158 | unknown protein (AT1G34010) mRNA | 5.0 | 9.0 | same phenotype |
| BR 160 | SCL5; transcription factor (SCL5) | 5.0 | 5.8 | same phenotype |
| BR162 | Brassica rapa subsp. pekinensis clone KBrB005N03, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 165 | Arabidopsis thaliana genomic DNA, chromosome 5, P1 clone:MZN1 | 5.0 | 9.0 | more number of tiller |
| BR 171 | Brassica rapa subsp. pekinensis clone KBrH004A18, complete sequence | 5.5 | 9.0 | more number of tiller |
| BR 173 | BSD domain-containing protein | 4.7 | 9.0 | same phenotype |
| BR 174 | SEC14 cytosolic factor family protein/phosphoglyceride transfer family protein | 5.5 | 9.0 | same phenotype |
| BR 178 | unknown protein | 4.0 | 9.0 | same phenotype |
| BR 179 | Zea mays clone 8974 mRNA sequence | 5.5 | 5.6 | same phenotype |
| BR 192 | glycosyltransferase family protein | 5.0 | 9.0 | same phenotype |
| BR 195 | ASP2 (aspartate aminotransferase 2 (ASP2) | 5.7 | 9.0 | same phenotype |
| BR 225 | unknown | 5.0 | 9.0 | same phenotype |
| BR 240 | Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 246 | CTR1 (constitutive triple response 1 | 5.0 | 9.0 | same phenotype |
| BR 250 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 255 | Indole-3-glycerol phosphate synthase | 5.5 | 9.0 | same phenotype |
| BR 256 | leucine-rich repeat transmembrane protein kinase | 5.0 | 9.0 | same phenotype |
| BR 257 | Unknown | 5.0 | 9.0 | same phenotype |
| BR 261 | Arabidopsis thaliana Full-length cDNA Complete sequence from clone GSLTSIL12ZB10 of Silique of strain col-0 of Arabidopsis thaliana (thale cress) | 5.0 | 9.0 | same phenotype |
| BR 262 | Glycosyl hydrolase family 17 protein | 4.8 | 9.0 | same phenotype |
| BR 263 | CA2 (beta carbonic anhydrase 2); carbonate dehydratase/zinc ion binding (CA2) | 5.0 | 9.0 | longer panicle length |
| BR 264 | Eukaryotic translation initiation factor 1A | 5.0 | 9.0 | same phenotype |
| BR 265 | Zea mays clone 11869 mRNA sequence | 5.0 | 9.0 | shorter plant height |
| BR 271 | rhodanese-like domain-containing protein | 5.0 | 9.0 | same phenotype |
| BR 273 | WD-40 repeat family protein | 5.0 | 9.0 | same phenotype |
| BR 274 | Brassica rapa subsp. pekinensis clone KBrH006E24, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 275 | pentatricopeptide (PPR) repeat-containing protein | 5.0 | 6.0 | same phenotype |
| BR 293 | Zea mays clone 11032 mRNA sequence | 4.0 | 9.0 | same phenotype |
| BR 303 | nodulin family protein | 5.0 | 9.0 | same phenotype |
| BR 305 | Unknown protein | 5.0 | 9.0 | same phenotype |
| BR 308 | Zea mays clone 15053 mRNA sequence | 4.9 | 9.0 | same phenotype |
| BR 311 | Actin (Act1) mRNA | 5.0 | 9.0 | same phenotype |
| BR 318 | unknown protein | 5.4 | 9.0 | same phenotype |
| BR 319 | aldehyde dehydrogenase | 5.0 | 9.0 | same phenotype |
| BR 322 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 336 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 323 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 330 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 336 | unknown protein | 5.0 | 9.0 | shorter plant height |
| BR 339 | HDA6 (histone deacetylase 6) | 5.0 | 9.0 | shorter plant height |
| BR 343 | ATNUDT2 (Arabidopsis thaliana Nudix hydrolase homolog 2); ADP-ribose diphosphatase/ NAD binding/hydrolase (ATNUDT2) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 348 | ELO1 (elongate 1) mRNA | 5.0 | 9.0 | more green leaves, shorter plant height |
| BR 349 | unknown protein | 5.0 | 6.0 | same phenotype |
| BR 367 | unknown gene function | 4.8 | 9.0 | same phenotype |
| BR 368 | ribulose 1,5-bisphosphate carboxylase/oxygenase small subunit | 5.0 | 9.0 | more green leaves |
| BR 369 | 60S ribosomal protein L10A (RPL10aC) | 5.0 | 9.0 | same phenotype |
| BR 370 | tubulin-specific chaperone C-related | 5.0 | 9.0 | same phenotype |
| BR 374 | ABC1 family protein | 5.0 | 9.0 | same phenotype |
| BR 375 | Zea mays clone 13431 mRNA sequence | 5.5 | 9.0 | same phenotype |
| BR 376 | transcriptional repressor | 5.0 | 9.0 | same phenotype |
| BR 377 | Arabidopsis thaliana clone 37944 mRNA, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 383 | CLA1-like protein mRNA | 5.0 | 9.0 | same phenotype |
| BR 384 | unknown protein | 5.0 | 9.0 | shorter plant height |
| BR 386 | 60S ribosomal protein L28 (RPL28C) | 5.0 | 9.0 | less green leaves, shorter plant height |
| BR 393 | unknown protein | 5.5 | 9.0 | same phenotype |
| BR 418 | WNK1 (WITH NO LYSINE (K) 1); kinase (WNK1) mRNA | 5.2 | 9.0 | same phenotype |
| BR 425 | Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence | 5.5 | 9.0 | shorter plant height, longer panicle length |
| BR 436 | putative nuclear transport factor 2 mRNA | 5.0 | 9.0 | same phenotype |
| BR 438 | 60S acidic ribosomal protein P0 (RPP0B) | 5.0 | 9.0 | same phenotype |
| BR 445 | transporter-related | 5.0 | 9.0 | more green leaves |
| BR 449 | FTSH8 (FtsH protease 8); ATP-dependent peptidase/ATPase/ metallopeptidase/ zinc ion binding (FTSH8) mRNA | 5.0 | 6.9 | same phenotype |
| BR 450 | Brassica rapa subsp. pekinensis clone KBrS015K01, complete sequence | 5.0 | 9.0 | more green leaves |
| BR 452 | nuclear transport factor 2 mRNA | 5.2 | 9.0 | more green leaves |
| BR 499 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 7.0 | same phenotypE |
| BR467 | Arabidopsis thaliana PTAC16 (PLASTID TRANSCRIPTIONALLY ACTIVE 16); binding/catalytic (PTAC16) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 468 | Brassica rapa subsp. pekinensis clone KBrB034N10, complete sequence | 5.0 | 9.0 | more green leaves |
| BR 471 | Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 470 | B-box type zinc finger protein (Bbox1) mRNA | 9.0 | 9.0 | more number of tiller |
| BR 472 | Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence | 5.0 | 6.4 | same phenotype |
| BR 473 | Zea mays clone 17728 mRNA sequence | 5.0 | 9.0 | shorter culm length |
| BR 474 | ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 475 | Zea mays clone 17728 mRNA sequence | 5.0 | 9.0 | more taller plant height |
| BR 478 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 479 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 480 | Arabidopsis thaliana unknown protein (AT3G18050) mRNA | 5.0 | 9.0 | same phenotype |
| BR 481 | AL5 (ALFIN-LIKE 5); DNA binding/methylated histone residue binding (AL5) | 5.0 | 9.0 | same phenotype |
| BR 482 | Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds | 5.0 | 9.0 | more green leaves, longer panicle length |
| BR 482 | unknown protein | 5.0 | 9.0 | more green leaves |
| BR 484 | Zea mays clone 12745 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 486 | Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 487 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 488 | unknown gene function | 5.7 | 9.0 | more green leaves |
| BR 489 | Arabidopsis thaliana 60S ribosomal protein L13 (RPL13D) (AT5G23900) mRNA, complete cds | 5.0 | 9.0 | more green leaves |
| BR 490 | short-chain dehydrogenase/reductase (SDR) family protein | 5.0 | 9.0 | more green leaves |
| BR 491 | transducin family protein/WD-40 repeat family protein (AT2G16405) mRNA | 5.0 | 9.0 | same phenotype |
| BR 492 | Phagocytosis and cell motility protein ELMO1-related (AT3G03610) mRNA | 5.0 | 9.0 | same phenotype |
| BR 494 | HD2C (HISTONE DEACETYLASE 2C) | 5.0 | 9.0 | same phenotype |
| BR 495 | unknown protein | 5.1 | 9.0 | same phenotype |
| BR 496 | unknown gene function | 5.0 | 9.0 | shorter culm length |
| BR 497 | unknown gene function | 5.0 | 9.0 | same phenotype |
| BR 498 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 499 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 485 | Arabidopsis thaliana unknown protein (At1g15180) mRNA, complete cds/MATE efflux family protein [Arabidopsis thaliana] | 5.0 | 9.0 | shorter culm length |
| BR 500 | Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA | 5.1 | 9.0 | same phenotype |
| BR 501 | Brassica rapa subsp. pekinensis clone KBrB030F12, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 502 | Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 503 | metal tolerance protein 5 (MTP5) | 5.0 | 9.0 | same phenotype |
| BR 504 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 505 | Brassica napus 1433-3 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 506 | Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds | 5.0 | 9.0 | shorter culm length |
| BR 507 | Arabidopsis thaliana 60S ribosomal protein L21 (RPL21C) (AT1G09690) mRNA, complete cds | 5.0 | 9.0 | shorter culm length |
| BR 508 | Arabidopsis thaliana BOU (A BOUT DE SOUFFLE); binding/transporter (BOU) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 509 | Arabidopsis thaliana ING1 (INHIBITOR OF GROWTH 1); DNA binding/methylated histone residue binding (ING1) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 510 | hypothetical protein | 5.0 | 9.0 | shorter culm length |
| BR 511 | VHS domain-containing protein/GAT domain-containing protein | 5.3 | 9.0 | same phenotype |
| BR 512 | RsGGT2 mRNA for gamma-glutamyl transferase | 5.0 | 9.0 | same phenotype |
| BR 513 | hypothetical protein | 4.5 | 5.0 | same phenotype |
| BR 514 | Arabidopsis thaliana clone 25991 mRNA, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 515 | Prephenate dehydrogenase family protein | 5.0 | 9.0 | same phenotype |
| BR 516 | Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 517 | ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) | 5.0 | 9.0 | same phenotype |
| BR 518 | Arabidopsis thaliana ATS1; glycerol-3-phosphate O-acyltransferase (ATS1) mRNA, complete cds | 3.0 | 9.0 | same phenotype |
| BR 519 | Arabidopsis thaliana HVA22J (HVA22-LIKE PROTEIN J) | 5.0 | 9.0 | same phenotype |
| BR 520 | Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 521 | Zea mays clone 10882 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 522 | hypothetical protein, mRNA | 3.0 | 9.0 | same phenotype |
| BR 523 | Arabidopsis thaliana AtMAPR2 (Arabidopsis thaliana membrane-associated progesterone binding protein 2); heme binding (AtMAPR2) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 524 | inorganic pyrophosphatase family protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 525 | unknown gene function | 5.5 | 9.0 | same phenotype |
| BR 526 | PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 | 5.0 | 9.0 | same phenotype |
| BR 527 | Brassica rapa clone 231 CLA1-like protein mRNA, complete cd | 5.0 | 9.0 | same phenotype |
| BR 528 | NAK; ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) mRNA | 5.0 | 9.0 | same phenotype |
| BR 529 | unknown gene function | 6.0 | 9.0 | same phenotype |
| BR 530 | Myb family transcription factor | 5.2 | 9.0 | same phenotype |
| BR 531 | Zea mays clone 14041 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 532 | Sterol 4-alpha-methyl-oxidase 2 (SMO2-2) | 5.0 | 9.0 | same phenotype |
| BR 533 | hypothetical protein | 5.0 | 9.0 | more taller plant height |
| BR 534 | hypothetical protein | 4.5 | 9.0 | same phenotype |
| BR 535 | mRNA for ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 536 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 537 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 538 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 539 | Raphanus sativus SAUR gene | 5.0 | 9.0 | same phenotype |
| BR 541 | LHCB4.2 (light harvesting complex PSII) | 3.0 | 9.0 | more taller plant height, longer panicle length |
| BR 542 | Brassica rapa subsp. pekinensis clone KBrB048F07, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 544 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | more taller plant height |
| BR 545 | Arabidopsis thaliana mRNA for 50S ribosomal protein L3, complete cds, clone: RAFL04-10-N10 | 5.0 | 9.0 | more taller plant height |
| BR 546 | Arabidopsis thaliana ATNUDX19 (ARABIDOPSIS THALIANA NUDIX HYDROLASE HOMOLOG 19); hydrolase/ metal ion binding (ATNUDX19) mRNA, complete cds | 4.0 | 9.0 | more taller plant height |
| BR 547 | UDP-galactose transporter 3 (ATUTR3) | 3.0 | 5.0 | same phenotype |
| BR 548 | A. lyrata subsp. lyrata inorganic pyrophosphatase family protein, mRNA | 5.0 | 9.0 | more taller plant height |
| BR 549 | BcRUBP mRNA for ribulose 1,5-bisphosphate carboxylase/oxygenase | 5.0 | 9.0 | more taller plant height |
| BR 550 | Arabidopsis thaliana ATRDH2 (ARABIDOPSIS THALIANA RHODANESE HOMOLOGUE 2); 3-mercaptopyruvate sulfurtransferase/ thiosulfate sulfurtransferase (ATRDH2) mRNA, complete cds | 5.0 | 9.0 | shorter plant height |
| BR 551 | Arabidopsis thaliana RNA binding (AT4G39040) mRNA, complete cds | 5.0 | 5.0 | same phenotype |
| BR 552 | ribosomal protein S6 family protein | 5.0 | 9.0 | same phenotype |
| BR 553 | Brassica rapa PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2, complete cds | 5.0 | 9.0 | same phenotype |
| BR 554 | transducin family protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 555 | Brassica rapa subsp. pekinensis clone KBrB034A02, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 556 | Hypothetical protein, mRNA | 5.0 | 9.0 | more taller plant height |
| BR 557 | BnD21 mRNA sequence | 5.0 | 5.0 | more taller plant height |
| BR 558 | unknown gene function | 5.0 | 9.0 | more taller plant height |
| BR 560 | B.oleracea mRNA for ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 561 | Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds | 5.0 | 5.8 | longer culm length |
| BR 562 | Arabidopsis thaliana protein binding/zinc ion binding (AT5G45100) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 563 | Brassica rapa subsp. pekinensis clone KBrB036G18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 566 | Arabidopsis thaliana ALB3 (ALBINO 3); P-P-bond-hydrolysis-driven protein transmembrane transporter (ALB3) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 567 | Zea mays clone 12993 mRNA sequence | 9.0 | 9.0 | same phenotype |
| BR 569 | Zea mays clone 14041 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 571 | Arabidopsis thaliana mRNA for putative mrp protein, complete cds, clone: RAFL05-19-J04 | 5.0 | 9.0 | same phenotype |
| BR 572 | Brassica rapa clone 231 CLA1-like protein mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 579 | Zea mays clone 20053 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 734 | Arabidopsis thaliana PDCB1 (PLASMODESMATA CALLOSE-BINDING PROTEIN 1); callose binding/polysaccharide binding (PDCB1) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 582 | isoflavone reductase family protein | 5.0 | 9.0 | same phenotype |
| BR 583 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 525 | disease resistance response protein | 5.0 | 6.8 | same phenotype |
| BR 633 | ribosomal protein L9 family protein | 5.0 | 9.0 | same phenotype |
| BR 640 | TMS membrane family protein/tumour differentially expressed (TDE) family protein | 5.0 | 9.0 | same phenotype |
| BR 660 | translational inhibitor protein like | 5.0 | 9.0 | same phenotype |
| BR 670 | RPL9 (RIBOSOMAL PROTEIN L9); structural constituent of ribosome (RPL9) | 5.0 | 9.0 | same phenotype |
| BR 700 | tonoplast intrinsic protein bobTIP26-1 | 5.0 | 9.0 | same phenotype |
| BR 708 | ATPHB2 (PROHIBITIN 2) (ATPHB2) | 5.0 | 9.0 | same phenotype |
| BR 721 | unknown protein | 5.0 | 5.3 | same phenotype |
| BR 725 | 40S ribosomal protein S4 (RPS4A) | 5.0 | 9.0 | same phenotype |
| BR 735 | SPP2 (SUCROSE-6F-PHOSPHATE PHOSPHOHYDROLASE 2); catalytic/ magnesium ion binding/phosphatase/ sucrose-phosphatase (SPP2) | 5.0 | 9.0 | same phenotype |
| BR 743 | mitochondrial pyruvate dehydrogenase kinase (PDK) | 5.0 | 7.0 | same phenotype |
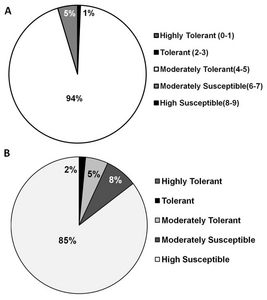
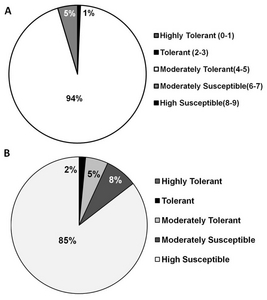
Acold stress score: 1= seedling dark green, 3=seedling light green, 5=seedling yellow, 7=seedling brown, 9=seedling dead. The cold screening was conducted at Rural Development Administration, Chuncheon, Korea
BSalinity stress score was based on the following at seedling stage: 1=Growth nearly normal, 3= Growth nearly normal but some leaves rolled sal), 5= Growth reduced and most leaves rolled(salt; only a few elongating, 7= Growth completely ceases; most leaves dry; some plants dying, 9= Almost all plants dead or dying
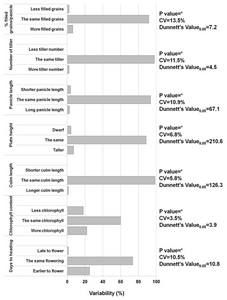
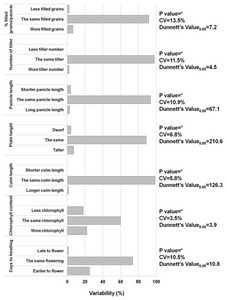
CObserved phenotype was based on significant differences between the wilt type Gopum using pair-wise Dunnetts multiple comparison. The same phenotype means no significant differences in the agro-morphological character such as plant height, culm length, chlorophyll contents, panicle length, number of tillers.
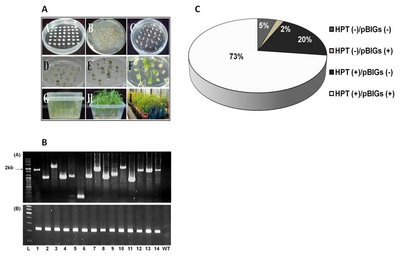
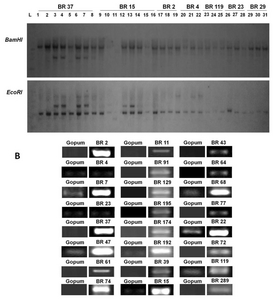

Identified transgenic rice with cold, salinity and drought tolerance.
| Clone | Tolerance to NaCl stresses (130 mM) | Tolerance to Cold | Drought (20% PEG) | Gene function (based on BLAST) |
|---|---|---|---|---|
| BR5 | − | ++ | − | Unknown function |
| BR7 | + | ++ | + | DEAD BOX RNA helicase RH15 |
| BR15 | − | + | + | UDP-glucose 4-epimerase |
| BR37 | + | ++ | + | CBL-interacting protein kinase 1 |
| BR39 | − | ++ | − | Carbon-monoxide oxygenase (ACYB-1) |
| BR40 | + | + | − | Phosphatase 2C, putative/PP2C |
| BR74 | + | + | − | Haloacid dehydrogenase-like hydrolase |
| BR91 | + | + | − | Unknown protein (AT3G12210) |
| BR115 | + | ++ | + | At5g25810 mRNA |
| BR119 | + | ++ | + | Unknown function |
| BR503 | − | ++ | − | Unknown function |
| BR522 | − | ++ | − | Hypothetical protein |
| BR541 | − | ++ | − | Light harvesting complex PSII |
| BR547 | − | ++ | − | UDP galactose transporter 3 |
+, tolerant; ++, highly tolerant; −, susceptible. Wild type, Gopum is moderately tolerant to cold but susceptible to either salinity and drought. Evaluation was based on SES for Rice (IRRI-SES, 2002)
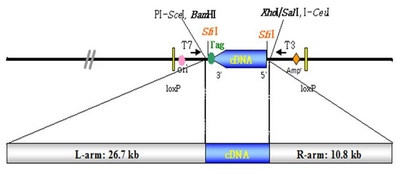
Composition of media used for tissue culture and transformation of rice.
| Medium | Composition |
|---|---|
| 2N6 (Embryogenic callus induction) | 4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 2 mg/L 2.4-D, 2.5 g/L Gelrite (pH 5.8) |
| AB (Agrobacterium culture) | 5 g/L Glucose, 15 g/L Bacto agar, 3 g/L K2HPO4, 1 g/L NaH2PO4, 1 g/L NH4Cl, 0.3 g/L MgSO4 · 7H2O, 0.15 g/L KCl, 13.25 mg/L CaCl2 · 2H2O, 2.5 mg/L FeSO4 · 7H2O (pH 7.2) |
| AAM-AS (Agrobacterium suspension and infection) | 0.12 g/L MgSO4, 0.15 g/L CaCl2 · 2H2O, 0.15 g/L NaH2PO4 · 2H2O, 3 g/L KCl, 0.5 g/L Casamino acid, 0.177 g/L L-Arginine, 0.04 g/L EDTA Ferric sodium, 7.6 mg/L MnSO4 · H2O, 2 mg/L ZnSO4· 7H2O, 0.025 mg/L CuSO4 · 5H2O, 0.025 mg/L CoCl2 · 6H2O, 0.75 mg/L KI, 3 mg/L H3BO3, 0.25 mg/L Na2MoO4 · 2H2O, 0.75 mg/L Glycine, 0.09 g/L L-Glutamine, 0.03 g/L L-Aspartic acid, 0.01 g/L Myo-Inositol, 0.1 mg/L Nicotinic acid, 0.1 mg/L Pyridoxine HCl, 1 mg/L Thiamine HCl, 68.5 g/L Sucrose, 36 g/L D-Glucose, 20 mg/L Acetosyringone (pH 5.2) |
| 2N6-AS (Co-culture) | 4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 10 g/L Glucose, 2 mg/L 2.4-D, 20 mg/L Acetosyringone, 0.15 g/L Dithiothreitol, 0.25 g/L Sodium thiosulfate, 5 mg/L Silver nitrate (pH 5.2) |
| N6D (Transgenic callus selection) | 4 g/L Chu (N6) medium (Micro and macro elements including vitamins) (Cat. No. C0204, Duchefa Biochemie B.V.), 0.3 g/L Casein hydrolysate, 0.5 g/L Glutamine, 0.5 g/L Proline, 30 g/L Sucrose, 2 mg/L 2.4-D, 2.5 g/L Gelrite, 250 mg/L Cefotaxime, 50 mg/L Hygromycin (pH 5.8) |
| REIII (Shoot formation) | 4.4 g/L MS medium including vitamins (Cat. No. M0222, Duchefa Biochemie B.V.), 0.5 mg/L NAA, 2 mg/L 6-BA, 20 g/L Sucrose, 30 g/L Sorbitol, 2 g/L Casamino acid, 250 mg/L Cefotaxime, 80 mg/L Hygromycin, 4 g/L Gelrite (pH 5.8) |
| HF (Root formation) | 4.4 g/L MS medium including vitamins (Cat. No. M0222, Duchefa Biochemie B.V.), 30 g/L Sucrose, 7.5 g/L Plant agar, 250 mg/L Cefotaxime, 80 mg/L Hygromycin (pH 5.8) |
Primers sequences used in the RT-PCR analysis and annotation of gene function by the NCBI database.
| Gene | Forward Primer | Reverse Primer | Gene Function |
|---|---|---|---|
| BR 2 | 5′-GTGTTGACGGACGTTGTGAC-3′ | 5′-CCGGGTCTAGATCTGTTCCA-3′ | Cysteine protease 1 |
| BR 4 | 5′-TGCCGTTATGCCATTTATGA-3′ | 5′-ATCAAGACCCATCTCGGTTG-3′ | Actin binding/depolymerizing factor 2 |
| BR 7 | 5′-GCATTTCCTAAAGCCACCAA-3′ | 5′-CCAGGAGTAGGCTCGATCTG-3′ | DEAD BOX RNA helicase RH15 - like protein |
| BR 11 | 5′-AACGAGAGAGGAGGGATGAT-3′ | 5′-CCAGAGGAGCTTCAGGGTTG-3′ | Alcohol dehydrogenase 1 |
| BR 15 | 5′-GCTTTGCGTGCCTTCTTATC-3′ | 5′-GAAGAACCCACTTGGCAAAA-3′ | UDP-glucose 4-epimerase 1 |
| BR 22 | 5′-ACTTGGCGAGCTTGACTCAT-3′ | 5′-GAGCATTTCTCCTGCGTTTC-3′ | Hydrogen transporting ATPase |
| BR 23 | 5′-AATGCACGGAATAGAGG-3′ | 5′-CAGGTCCTCATTACCGGTGT-3′ | Endoplasmic reticulum auxin-binding protein 2 mRNA |
| BR 37 | 5′-ATCGGAGGCAGCAAGTCAAGCG-3′ | 5′-CTCCATCGTAGCCTCCGTCAT-3′ | CBL-interacting protein kinase 1 |
| BR 39 | 5′-CAGGAAAAGAAGCGGATGAG-3′ | 5′-AGTGTGCCTGTCATCGTGAG-3′ | Carbon-monoxide oxygenase (ACYB-1) |
| BR 43 | 5′-AACTCCAAGCGTTGGATATG-3′ | 5′-TCTCCAGGTTCGTTTGCTCT-3′ | Hypothetical protein |
| BR 47 | 5′-CTAATGCTGACGGTGGGTTT-3′ | 5′-ACGAAAAGGTCGAAAGCTGA-3′ | Unknown gene function |
| BR 61 | 5′-CCTCTGATCCCACCACACTT-3′ | 5′-TTAGGAGCAGCACACACTGG-3′ | Rubredoxin family protein |
| BR 64 | 5′-AGAAGCTGATGGCTCGGTTA-3′ | 5′-CCTCTTCTCGGCTGAACAAC-3′ | Beta-lg-H3 domain-containing protein (FLA18) |
| BR 68 | 5′-CCTTACCGCCGCCACCATGTTT-3′ | 5′-AGGTGTCAAAAGTCGGGTTG-3′ | Unknown gene function |
| BR 72 | 5′-ACCAAACACTCACCGGAGTC-3′ | 5′-ATCACCACCCGCAAGTAAAG-3′ | Unknown protein function |
| BR 74 | 5′-CGATGTTCGATGTCCTCCTT-3′ | 5′-GGCTCGATCATCGTCATTCT-3′ | Haloacid dehalogenase-like hydrolase family protein |
| BR 77 | 5′-CATCTACCGAGCAACAGCAA-3′ | 5′-AATGCCCAATCCATACCAAA-3′ | Unknown (Zeamays clone 92871) |
| BR 91 | 5′-TGGAAGCTCTCTCGTGGAA-3′ | 5′-CTCGTCGGACATAAATGGT-3′ | Unknown protein |
| BR 119 | 5′-GTATCACTCTCGGGGTTCCA-3′ | 5′-CCAAACAAATGGGACAATCC-3′ | Unknown gene function |
| BR 129 | 5′-GTGGAGGGAGTCAACGATGT-3′ | 5′-CCGAGTCCCTGCGATATTTA-3′ | Dimethylmenaquinone methyltransferase family protein |
| BR 192 | 5′-CATAAGGTACGCGACGGTTT-3′ | 5′-ACTAAAGCAGCTTCGGTCCA-3′ | Glycosyltransferase family protein |
| BR 195 | 5′-CACTGGAGTTGACCCAAC-3′ | 5′-AATGCTAAGGGCACCAACAC-3′ | Aspartate aminotransferase 2 |
| BR 289 | 5′-TGATCAGATGGAGTGGTGGA-3′ | 5′-CTCCTCGCTTGTTGGAAGAC-3′ | Plastid-lipid associated protein PAP/fibrillin family protein |
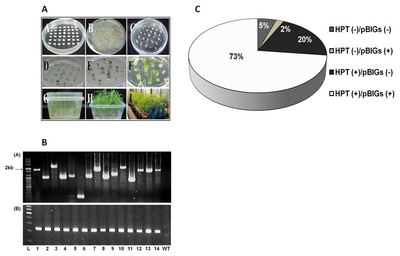
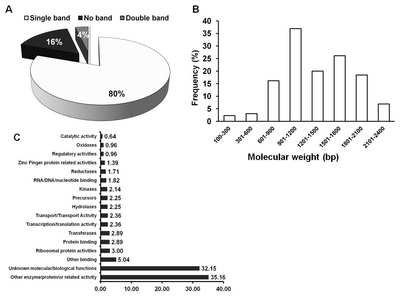
Number of transgenic rice lines produced, confirmed gene inserts and its respective generation.
| Generation* | Confirmed no. of gene inserts | No. of lines |
|---|---|---|
| T1 | 137 | 470 |
| T2 | 88 | 408 |
| T3 | 14 | 46 |
| T4 | 11 | 226 |
|
| ||
| Total | 250 | 1,150 |
List of transgenic rice with full-length cDNA from B. rapa, its annotation function and their morpho-agronomic traits.
| Code | Gene Function | Cold stress scorea | Salinity stress scoreb | Observed phenotypec |
|---|---|---|---|---|
| BR 2 | cysteine protease 1 | 5.7 | 9.0 | more green leaves |
| BR 4 | ADF2 (Actin depolymerizing factor 2) | 5.0 | 9.0 | more number of tiller |
| BR 5 | PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 | 3.0 | 9.0 | more green leaves |
| BR 6 | Zea mays clone 93807 mRNA sequence | 5.0 | 9.0 | more green leaves |
| BR 7 | Dead box RNA helicase RH15 - like protein | 3.0 | 3.0 | same phenotype |
| BR 9 | PEC-1 mRNA for Lipid transfer protein | 5.0 | 9.0 | more green leaves |
| BR 12 | Rac GTPase activator | 5.0 | 9.0 | same phenotype |
| BR 14 | CBL-interacting protein kinase 3 | 5.2 | 5.5 | less green leaves |
| BR 17 | mRNA for aluminum-induced | 5.0 | 9.0 | less green leaves |
| BR 23 | endoplasmic reticulum auxin-binding protein 2 | 9.0 | 7.0 | less green leaves |
| BR 26 | Brassica rapa subsp. pekinensis clone KBrH004A06, complete sequence | 5.3 | 9.0 | same phenotype |
| BR 29 | Phospholipase C | 5.2 | 7.0 | less green leaves |
| BR 33 | ABC transporter-like protein | 9.0 | 9.0 | less green leaves |
| BR 37 | CBL-interacting protein kinase 1 | 5.4 | 3.0 | less green leaves, less plant height |
| BR 39 | carbohydrate binding | 4.3 | 9.0 | less green leaves |
| BR 40 | protein phosphatase 2C, putative/PP2C, | 4.7 | 3.0 | less green leaves, shorter culm length |
| BR 46 | plasma membrane intrinsic protein 2 (PIP2) | 5.0 | 9.0 | less green leaves, more taller |
| BR 48 | senescence-associated cysteine protease (CP1) | 4.8 | 5.0 | more number of tiller |
| BR 49 | polypeptide of photosystem II reaction center | 9.0 | 9.0 | less green leaves |
| BR 50 | chlorophyll a/b binding protein | 9.0 | less green leaves | |
| BR 51 | methyl-CpG-binding domain 4 (MBD4) | 9.0 | 9.0 | less green leaves, longer panicle length |
| BR 57 | putative protein transport protein SEC61 alpha subunit | 5.0 | 7.0 | less green leaves |
| BR 60 | Brassica rapa subsp. pekinensis clone KBrB042D24, complete sequence | 5.0 | 9.0 | less green leaves |
| BR 61 | rubredoxin family protein | 6.0 | 9.0 | less green leaves |
| BR 64 | GDSL esterase/lipase APG | 5.0 | 9.0 | less green leaves, longer panicle length, Shorter culm length |
| BR 72 | WSCP1 mRNA for water-soluble chlorophyll protein precursor | 5.0 | 9.0 | same phenotype |
| BR 73 | unknown protein | 6.0 | 9.0 | less green leaves |
| BR 74 | haloacid dehalogenase-like hydrolase family protein | 5.0 | 3.0 | less green leaves, more number of tiller |
| BR81 | regulatory subunit of protein kinase CK2 | 5.0 | 7.0 | less green leaves, shorter culm length |
| BR 84 | ATCTIMC (Cytosolic triose phosphate isomerase) | 5.0 | 9.0 | shorter culm length |
| BR 89 | Brassica rapa subsp. pekinensis clone KBrH004P05, complete sequence | 5.7 | 9.0 | same phenotype |
| BR 91 | unknown protein | 5.2 | 3.0 | less green leaves, longer culm length |
| BR 92 | 5-enolpyruvylshikimate-3-phosphate synthase (EPSP) | 5.0 | 9.0 | same phenotype |
| BR 97 | G6PD1 (glucose-6-phosphate dehydrogenase 1) | 5.1 | 9.0 | same phenotype |
| BR 99 | 40S ribosomal protein S7 (RPS7C) | 5.0 | 9.0 | same phenotype |
| BR 104 | protein binding/zinc ion binding | 5.0 | 9.0 | same phenotype |
| BR 105 | Lhcb6 protein mRNA | 5.0 | 9.0 | more number of tiller |
| BR 111 | protein kinase family protein | 5.0 | 9.0 | same phenotype |
| BR 112 | Arabidopsis thaliana AT5g47540/MNJ7_13 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 113 | Brassica rapa subsp. pekinensis clone KBrB052L10, complete sequence | 5.8 | 9.0 | same phenotype |
| BR 114 | Zea mays clone 13119 mRNA sequence | 5.0 | 9.0 | less plant height |
| BR 115 | Arabidopsis thaliana At5g25810 mRNA, complete cds | 3.0 | 9.0 | same phenotype |
| BR 116 | Unknown | 5.0 | 9.0 | same phenotype |
| BR 118 | Vitis vinifera, whole genome shotgun sequence, contig VV78X164547.5, clone ENTAV 115 | 6.0 | 9.0 | same phenotype |
| BR 119 | unknown protein | 3.0 | 3.0 | shorter plant height |
| BR 120 | Zea mays clone 14950 mRNA sequence | 5.0 | 6.2 | same phenotype |
| BR 122 | DNAJ heat shock N-terminal domain-containing protein | 5.0 | 9.0 | same phenotype |
| BR 124 | putative thioredoxin reductase | 4.8 | 9.0 | same phenotype |
| BR 125 | Brassica rapa subsp. pekinensis clone KBrS004A14, complete sequence | 5.0 | 9.0 | less plant height |
| BR 128 | Zea mays clone 9139 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 129 | dimethylmenaquinone methyltransferase family protein | 5.0 | 9.0 | same phenotype |
| BR133 | prephenate dehydratase family protein | 5.0 | 9.0 | same phenotype |
| BR139 | 231 CLA1-like protein mRNA | 5.1 | 9.0 | same phenotype |
| BR147 | 231 CLA1-like protein mRNA | 5.0 | 9.0 | same phenotype |
| BR148 | protein kinase family protein/C-type lectin domain-containing protein | 5.0 | 5.2 | same phenotype |
| BR 149 | XTR4 (xyloglucan endotransglycolase 4) | 5.0 | 9.0 | same phenotype |
| BR 151 | clathrin adaptor complexes medium subunit-related | 5.0 | 9.0 | same phenotype |
| BR 152 | unknown mRNA | 5.0 | 9.0 | same phenotype |
| BR 158 | unknown protein (AT1G34010) mRNA | 5.0 | 9.0 | same phenotype |
| BR 160 | SCL5; transcription factor (SCL5) | 5.0 | 5.8 | same phenotype |
| BR162 | Brassica rapa subsp. pekinensis clone KBrB005N03, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 165 | Arabidopsis thaliana genomic DNA, chromosome 5, P1 clone:MZN1 | 5.0 | 9.0 | more number of tiller |
| BR 171 | Brassica rapa subsp. pekinensis clone KBrH004A18, complete sequence | 5.5 | 9.0 | more number of tiller |
| BR 173 | BSD domain-containing protein | 4.7 | 9.0 | same phenotype |
| BR 174 | SEC14 cytosolic factor family protein/phosphoglyceride transfer family protein | 5.5 | 9.0 | same phenotype |
| BR 178 | unknown protein | 4.0 | 9.0 | same phenotype |
| BR 179 | Zea mays clone 8974 mRNA sequence | 5.5 | 5.6 | same phenotype |
| BR 192 | glycosyltransferase family protein | 5.0 | 9.0 | same phenotype |
| BR 195 | ASP2 (aspartate aminotransferase 2 (ASP2) | 5.7 | 9.0 | same phenotype |
| BR 225 | unknown | 5.0 | 9.0 | same phenotype |
| BR 240 | Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 246 | CTR1 (constitutive triple response 1 | 5.0 | 9.0 | same phenotype |
| BR 250 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 255 | Indole-3-glycerol phosphate synthase | 5.5 | 9.0 | same phenotype |
| BR 256 | leucine-rich repeat transmembrane protein kinase | 5.0 | 9.0 | same phenotype |
| BR 257 | Unknown | 5.0 | 9.0 | same phenotype |
| BR 261 | Arabidopsis thaliana Full-length cDNA Complete sequence from clone GSLTSIL12ZB10 of Silique of strain col-0 of Arabidopsis thaliana (thale cress) | 5.0 | 9.0 | same phenotype |
| BR 262 | Glycosyl hydrolase family 17 protein | 4.8 | 9.0 | same phenotype |
| BR 263 | CA2 (beta carbonic anhydrase 2); carbonate dehydratase/zinc ion binding (CA2) | 5.0 | 9.0 | longer panicle length |
| BR 264 | Eukaryotic translation initiation factor 1A | 5.0 | 9.0 | same phenotype |
| BR 265 | Zea mays clone 11869 mRNA sequence | 5.0 | 9.0 | shorter plant height |
| BR 271 | rhodanese-like domain-containing protein | 5.0 | 9.0 | same phenotype |
| BR 273 | WD-40 repeat family protein | 5.0 | 9.0 | same phenotype |
| BR 274 | Brassica rapa subsp. pekinensis clone KBrH006E24, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 275 | pentatricopeptide (PPR) repeat-containing protein | 5.0 | 6.0 | same phenotype |
| BR 293 | Zea mays clone 11032 mRNA sequence | 4.0 | 9.0 | same phenotype |
| BR 303 | nodulin family protein | 5.0 | 9.0 | same phenotype |
| BR 305 | Unknown protein | 5.0 | 9.0 | same phenotype |
| BR 308 | Zea mays clone 15053 mRNA sequence | 4.9 | 9.0 | same phenotype |
| BR 311 | Actin (Act1) mRNA | 5.0 | 9.0 | same phenotype |
| BR 318 | unknown protein | 5.4 | 9.0 | same phenotype |
| BR 319 | aldehyde dehydrogenase | 5.0 | 9.0 | same phenotype |
| BR 322 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 336 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 323 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 330 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 336 | unknown protein | 5.0 | 9.0 | shorter plant height |
| BR 339 | HDA6 (histone deacetylase 6) | 5.0 | 9.0 | shorter plant height |
| BR 343 | ATNUDT2 (Arabidopsis thaliana Nudix hydrolase homolog 2); ADP-ribose diphosphatase/ NAD binding/hydrolase (ATNUDT2) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 348 | ELO1 (elongate 1) mRNA | 5.0 | 9.0 | more green leaves, shorter plant height |
| BR 349 | unknown protein | 5.0 | 6.0 | same phenotype |
| BR 367 | unknown gene function | 4.8 | 9.0 | same phenotype |
| BR 368 | ribulose 1,5-bisphosphate carboxylase/oxygenase small subunit | 5.0 | 9.0 | more green leaves |
| BR 369 | 60S ribosomal protein L10A (RPL10aC) | 5.0 | 9.0 | same phenotype |
| BR 370 | tubulin-specific chaperone C-related | 5.0 | 9.0 | same phenotype |
| BR 374 | ABC1 family protein | 5.0 | 9.0 | same phenotype |
| BR 375 | Zea mays clone 13431 mRNA sequence | 5.5 | 9.0 | same phenotype |
| BR 376 | transcriptional repressor | 5.0 | 9.0 | same phenotype |
| BR 377 | Arabidopsis thaliana clone 37944 mRNA, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 383 | CLA1-like protein mRNA | 5.0 | 9.0 | same phenotype |
| BR 384 | unknown protein | 5.0 | 9.0 | shorter plant height |
| BR 386 | 60S ribosomal protein L28 (RPL28C) | 5.0 | 9.0 | less green leaves, shorter plant height |
| BR 393 | unknown protein | 5.5 | 9.0 | same phenotype |
| BR 418 | WNK1 (WITH NO LYSINE (K) 1); kinase (WNK1) mRNA | 5.2 | 9.0 | same phenotype |
| BR 425 | Brassica rapa subsp. pekinensis clone KBrH080C09, complete sequence | 5.5 | 9.0 | shorter plant height, longer panicle length |
| BR 436 | putative nuclear transport factor 2 mRNA | 5.0 | 9.0 | same phenotype |
| BR 438 | 60S acidic ribosomal protein P0 (RPP0B) | 5.0 | 9.0 | same phenotype |
| BR 445 | transporter-related | 5.0 | 9.0 | more green leaves |
| BR 449 | FTSH8 (FtsH protease 8); ATP-dependent peptidase/ATPase/ metallopeptidase/ zinc ion binding (FTSH8) mRNA | 5.0 | 6.9 | same phenotype |
| BR 450 | Brassica rapa subsp. pekinensis clone KBrS015K01, complete sequence | 5.0 | 9.0 | more green leaves |
| BR 452 | nuclear transport factor 2 mRNA | 5.2 | 9.0 | more green leaves |
| BR 499 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 7.0 | same phenotypE |
| BR467 | Arabidopsis thaliana PTAC16 (PLASTID TRANSCRIPTIONALLY ACTIVE 16); binding/catalytic (PTAC16) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 468 | Brassica rapa subsp. pekinensis clone KBrB034N10, complete sequence | 5.0 | 9.0 | more green leaves |
| BR 471 | Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 470 | B-box type zinc finger protein (Bbox1) mRNA | 9.0 | 9.0 | more number of tiller |
| BR 472 | Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence | 5.0 | 6.4 | same phenotype |
| BR 473 | Zea mays clone 17728 mRNA sequence | 5.0 | 9.0 | shorter culm length |
| BR 474 | ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 475 | Zea mays clone 17728 mRNA sequence | 5.0 | 9.0 | more taller plant height |
| BR 478 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 479 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 480 | Arabidopsis thaliana unknown protein (AT3G18050) mRNA | 5.0 | 9.0 | same phenotype |
| BR 481 | AL5 (ALFIN-LIKE 5); DNA binding/methylated histone residue binding (AL5) | 5.0 | 9.0 | same phenotype |
| BR 482 | Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds | 5.0 | 9.0 | more green leaves, longer panicle length |
| BR 482 | unknown protein | 5.0 | 9.0 | more green leaves |
| BR 484 | Zea mays clone 12745 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 486 | Arabidopsis thaliana LHCB3 (LIGHT-HARVESTING CHLOROPHYLL B-BINDING PROTEIN 3); structural molecule (LHCB3) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 487 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 488 | unknown gene function | 5.7 | 9.0 | more green leaves |
| BR 489 | Arabidopsis thaliana 60S ribosomal protein L13 (RPL13D) (AT5G23900) mRNA, complete cds | 5.0 | 9.0 | more green leaves |
| BR 490 | short-chain dehydrogenase/reductase (SDR) family protein | 5.0 | 9.0 | more green leaves |
| BR 491 | transducin family protein/WD-40 repeat family protein (AT2G16405) mRNA | 5.0 | 9.0 | same phenotype |
| BR 492 | Phagocytosis and cell motility protein ELMO1-related (AT3G03610) mRNA | 5.0 | 9.0 | same phenotype |
| BR 494 | HD2C (HISTONE DEACETYLASE 2C) | 5.0 | 9.0 | same phenotype |
| BR 495 | unknown protein | 5.1 | 9.0 | same phenotype |
| BR 496 | unknown gene function | 5.0 | 9.0 | shorter culm length |
| BR 497 | unknown gene function | 5.0 | 9.0 | same phenotype |
| BR 498 | unknown protein | 5.0 | 9.0 | same phenotype |
| BR 499 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 485 | Arabidopsis thaliana unknown protein (At1g15180) mRNA, complete cds/MATE efflux family protein [Arabidopsis thaliana] | 5.0 | 9.0 | shorter culm length |
| BR 500 | Arabidopsis lyrata subsp. lyrata hypothetical protein, mRNA | 5.1 | 9.0 | same phenotype |
| BR 501 | Brassica rapa subsp. pekinensis clone KBrB030F12, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 502 | Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 503 | metal tolerance protein 5 (MTP5) | 5.0 | 9.0 | same phenotype |
| BR 504 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 505 | Brassica napus 1433-3 mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 506 | Brassica oleracea var. botrytis tonoplast intrinsic protein bobTIP26-1 mRNA, complete cds | 5.0 | 9.0 | shorter culm length |
| BR 507 | Arabidopsis thaliana 60S ribosomal protein L21 (RPL21C) (AT1G09690) mRNA, complete cds | 5.0 | 9.0 | shorter culm length |
| BR 508 | Arabidopsis thaliana BOU (A BOUT DE SOUFFLE); binding/transporter (BOU) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 509 | Arabidopsis thaliana ING1 (INHIBITOR OF GROWTH 1); DNA binding/methylated histone residue binding (ING1) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 510 | hypothetical protein | 5.0 | 9.0 | shorter culm length |
| BR 511 | VHS domain-containing protein/GAT domain-containing protein | 5.3 | 9.0 | same phenotype |
| BR 512 | RsGGT2 mRNA for gamma-glutamyl transferase | 5.0 | 9.0 | same phenotype |
| BR 513 | hypothetical protein | 4.5 | 5.0 | same phenotype |
| BR 514 | Arabidopsis thaliana clone 25991 mRNA, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 515 | Prephenate dehydrogenase family protein | 5.0 | 9.0 | same phenotype |
| BR 516 | Brassica rapa subsp. pekinensis clone KBrB028I01, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 517 | ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) | 5.0 | 9.0 | same phenotype |
| BR 518 | Arabidopsis thaliana ATS1; glycerol-3-phosphate O-acyltransferase (ATS1) mRNA, complete cds | 3.0 | 9.0 | same phenotype |
| BR 519 | Arabidopsis thaliana HVA22J (HVA22-LIKE PROTEIN J) | 5.0 | 9.0 | same phenotype |
| BR 520 | Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 521 | Zea mays clone 10882 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 522 | hypothetical protein, mRNA | 3.0 | 9.0 | same phenotype |
| BR 523 | Arabidopsis thaliana AtMAPR2 (Arabidopsis thaliana membrane-associated progesterone binding protein 2); heme binding (AtMAPR2) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 524 | inorganic pyrophosphatase family protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 525 | unknown gene function | 5.5 | 9.0 | same phenotype |
| BR 526 | PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2 | 5.0 | 9.0 | same phenotype |
| BR 527 | Brassica rapa clone 231 CLA1-like protein mRNA, complete cd | 5.0 | 9.0 | same phenotype |
| BR 528 | NAK; ATP binding/kinase/ protein kinase/ protein serine/threonine kinase/ protein tyrosine kinase (NAK) mRNA | 5.0 | 9.0 | same phenotype |
| BR 529 | unknown gene function | 6.0 | 9.0 | same phenotype |
| BR 530 | Myb family transcription factor | 5.2 | 9.0 | same phenotype |
| BR 531 | Zea mays clone 14041 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 532 | Sterol 4-alpha-methyl-oxidase 2 (SMO2-2) | 5.0 | 9.0 | same phenotype |
| BR 533 | hypothetical protein | 5.0 | 9.0 | more taller plant height |
| BR 534 | hypothetical protein | 4.5 | 9.0 | same phenotype |
| BR 535 | mRNA for ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 536 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 537 | Brassica rapa subsp. pekinensis clone KBrB021M22, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 538 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 539 | Raphanus sativus SAUR gene | 5.0 | 9.0 | same phenotype |
| BR 541 | LHCB4.2 (light harvesting complex PSII) | 3.0 | 9.0 | more taller plant height, longer panicle length |
| BR 542 | Brassica rapa subsp. pekinensis clone KBrB048F07, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 544 | Brassica rapa subsp. pekinensis clone KBrH003N18, complete sequence | 5.0 | 9.0 | more taller plant height |
| BR 545 | Arabidopsis thaliana mRNA for 50S ribosomal protein L3, complete cds, clone: RAFL04-10-N10 | 5.0 | 9.0 | more taller plant height |
| BR 546 | Arabidopsis thaliana ATNUDX19 (ARABIDOPSIS THALIANA NUDIX HYDROLASE HOMOLOG 19); hydrolase/ metal ion binding (ATNUDX19) mRNA, complete cds | 4.0 | 9.0 | more taller plant height |
| BR 547 | UDP-galactose transporter 3 (ATUTR3) | 3.0 | 5.0 | same phenotype |
| BR 548 | A. lyrata subsp. lyrata inorganic pyrophosphatase family protein, mRNA | 5.0 | 9.0 | more taller plant height |
| BR 549 | BcRUBP mRNA for ribulose 1,5-bisphosphate carboxylase/oxygenase | 5.0 | 9.0 | more taller plant height |
| BR 550 | Arabidopsis thaliana ATRDH2 (ARABIDOPSIS THALIANA RHODANESE HOMOLOGUE 2); 3-mercaptopyruvate sulfurtransferase/ thiosulfate sulfurtransferase (ATRDH2) mRNA, complete cds | 5.0 | 9.0 | shorter plant height |
| BR 551 | Arabidopsis thaliana RNA binding (AT4G39040) mRNA, complete cds | 5.0 | 5.0 | same phenotype |
| BR 552 | ribosomal protein S6 family protein | 5.0 | 9.0 | same phenotype |
| BR 553 | Brassica rapa PsbQ2 mRNA for oxygen-evolving enhancer protein 3-2, complete cds | 5.0 | 9.0 | same phenotype |
| BR 554 | transducin family protein, mRNA | 5.0 | 9.0 | same phenotype |
| BR 555 | Brassica rapa subsp. pekinensis clone KBrB034A02, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 556 | Hypothetical protein, mRNA | 5.0 | 9.0 | more taller plant height |
| BR 557 | BnD21 mRNA sequence | 5.0 | 5.0 | more taller plant height |
| BR 558 | unknown gene function | 5.0 | 9.0 | more taller plant height |
| BR 560 | B.oleracea mRNA for ACC oxidase (ACC0x2) | 5.0 | 9.0 | same phenotype |
| BR 561 | Arabidopsis thaliana COX6B (CYTOCHROME C OXIDASE 6B); cytochrome-c oxidase (COX6B) mRNA, complete cds | 5.0 | 5.8 | longer culm length |
| BR 562 | Arabidopsis thaliana protein binding/zinc ion binding (AT5G45100) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 563 | Brassica rapa subsp. pekinensis clone KBrB036G18, complete sequence | 5.0 | 9.0 | same phenotype |
| BR 566 | Arabidopsis thaliana ALB3 (ALBINO 3); P-P-bond-hydrolysis-driven protein transmembrane transporter (ALB3) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 567 | Zea mays clone 12993 mRNA sequence | 9.0 | 9.0 | same phenotype |
| BR 569 | Zea mays clone 14041 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 571 | Arabidopsis thaliana mRNA for putative mrp protein, complete cds, clone: RAFL05-19-J04 | 5.0 | 9.0 | same phenotype |
| BR 572 | Brassica rapa clone 231 CLA1-like protein mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 579 | Zea mays clone 20053 mRNA sequence | 5.0 | 9.0 | same phenotype |
| BR 734 | Arabidopsis thaliana PDCB1 (PLASMODESMATA CALLOSE-BINDING PROTEIN 1); callose binding/polysaccharide binding (PDCB1) mRNA, complete cds | 5.0 | 9.0 | same phenotype |
| BR 582 | isoflavone reductase family protein | 5.0 | 9.0 | same phenotype |
| BR 583 | hypothetical protein | 5.0 | 9.0 | same phenotype |
| BR 525 | disease resistance response protein | 5.0 | 6.8 | same phenotype |
| BR 633 | ribosomal protein L9 family protein | 5.0 | 9.0 | same phenotype |
| BR 640 | TMS membrane family protein/tumour differentially expressed (TDE) family protein | 5.0 | 9.0 | same phenotype |
| BR 660 | translational inhibitor protein like | 5.0 | 9.0 | same phenotype |
| BR 670 | RPL9 (RIBOSOMAL PROTEIN L9); structural constituent of ribosome (RPL9) | 5.0 | 9.0 | same phenotype |
| BR 700 | tonoplast intrinsic protein bobTIP26-1 | 5.0 | 9.0 | same phenotype |
| BR 708 | ATPHB2 (PROHIBITIN 2) (ATPHB2) | 5.0 | 9.0 | same phenotype |
| BR 721 | unknown protein | 5.0 | 5.3 | same phenotype |
| BR 725 | 40S ribosomal protein S4 (RPS4A) | 5.0 | 9.0 | same phenotype |
| BR 735 | SPP2 (SUCROSE-6F-PHOSPHATE PHOSPHOHYDROLASE 2); catalytic/ magnesium ion binding/phosphatase/ sucrose-phosphatase (SPP2) | 5.0 | 9.0 | same phenotype |
| BR 743 | mitochondrial pyruvate dehydrogenase kinase (PDK) | 5.0 | 7.0 | same phenotype |
Acold stress score: 1= seedling dark green, 3=seedling light green, 5=seedling yellow, 7=seedling brown, 9=seedling dead. The cold screening was conducted at Rural Development Administration, Chuncheon, Korea
BSalinity stress score was based on the following at seedling stage: 1=Growth nearly normal, 3= Growth nearly normal but some leaves rolled sal), 5= Growth reduced and most leaves rolled(salt; only a few elongating, 7= Growth completely ceases; most leaves dry; some plants dying, 9= Almost all plants dead or dying
CObserved phenotype was based on significant differences between the wilt type Gopum using pair-wise Dunnetts multiple comparison. The same phenotype means no significant differences in the agro-morphological character such as plant height, culm length, chlorophyll contents, panicle length, number of tillers.
+, tolerant; ++, highly tolerant; −, susceptible. Wild type, Gopum is moderately tolerant to cold but susceptible to either salinity and drought. Evaluation was based on SES for Rice (IRRI-SES, 2002)
cold stress score: 1= seedling dark green, 3=seedling light green, 5=seedling yellow, 7=seedling brown, 9=seedling dead. The cold screening was conducted at Rural Development Administration, Chuncheon, Korea
Salinity stress score was based on the following at seedling stage: 1=Growth nearly normal, 3= Growth nearly normal but some leaves rolled sal), 5= Growth reduced and most leaves rolled(salt; only a few elongating, 7= Growth completely ceases; most leaves dry; some plants dying, 9= Almost all plants dead or dying
Observed phenotype was based on significant differences between the wilt type Gopum using pair-wise Dunnetts multiple comparison. The same phenotype means no significant differences in the agro-morphological character such as plant height, culm length, chlorophyll contents, panicle length, number of tillers.