Abstract
Tomato (Solanum lycopersicum) is a premier global crop, celebrated not only for its economic importance but also as a fundamental “well-being food” that provides essential vitamins, minerals, and potent antioxidants. This review highlights the transformative impact of CRISPR/Cas9 and next-generation precision tools, such as base and prime editing, on modern tomato breeding. Beyond functional genomics, we focus on the strategic engineering of tomato to maximize productivity through fruit size optimization and the biofortification of high-value metabolites (e.g., GABA, lycopene, and vitamins). Furthermore, we examine recent advances in developing multi-stress resilience against both biotic and abiotic challenges. A significant emphasis is placed on re-designing plant architecture and developmental traits to tailor tomato varieties for future agricultural systems, particularly indoor and vertical farming. By advanced delivery methods, CRISPR technologies offer unprecedented potential to accelerate the development of “smart” tomato, ensuring sustainable agriculture and global food security in a changing climate.
-
Key words: Tomato, CRISPR/Cas9, Stress Tolerance, Genome editing, Productivity, Well-being
Introduction
Tomato (
Solanum lycopersicum) is a globally significant economic crop and a fundamental “well-being food” providing essential vitamins, minerals, and potent antioxidants like lycopene (
Wang et al. 2023). However, escalating biotic and abiotic stresses driven by climate change, coupled with shrinking arable land and surging consumer demand for highly functional foods, have exposed the limitations of conventional crossbreeding, necessitating innovative agricultural solutions.
In this context, next-generation precision genome editing technologies “prominently the Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-associated protein 9 (CRISPR/Cas9) system, alongside base and prime editing” are fundamentally revolutionizing modern tomato breeding. Beyond traditional functional genomics, these advanced tools enable the strategic engineering of tomato to maximize agricultural productivity through optimization of plant development and abiotic and biotic stress resistance, and to achieve rapid bio fortification of high-value metabolites, such as GABA and lycopene (
Zhang et al. 2024;
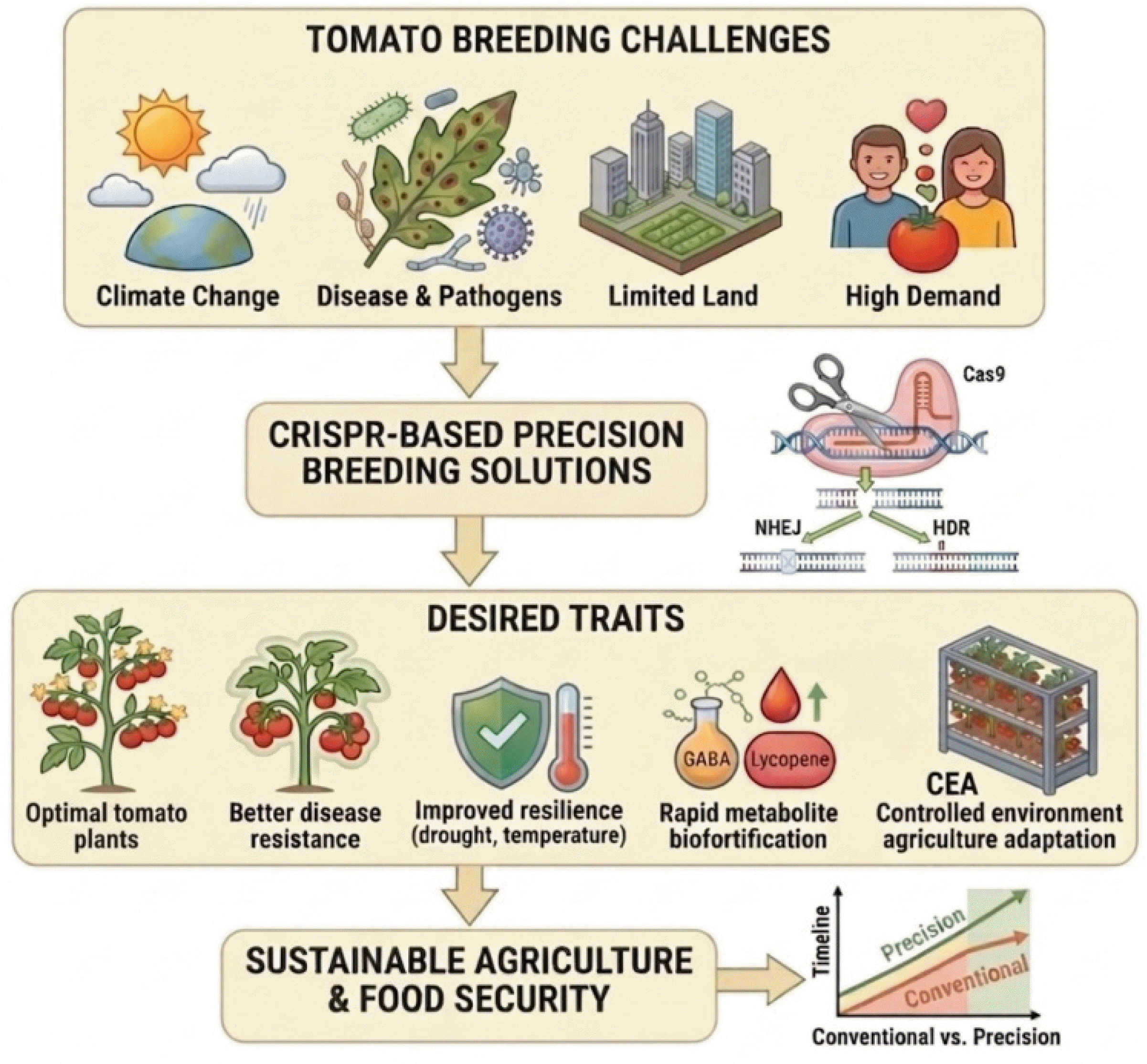
Fig. 1).
Furthermore, recent breakthroughs have expanded beyond simple trait improvement, focusing on the sophisticated re-designing of plant architecture and developmental processes to tailor varieties specifically for controlled-environment agriculture (CEA) and urban vertical farming, while simultaneously conferring multi-stress resilience (
Kwon et al. 2020). Consequently, this review comprehensively examines the transformative impact of advanced CRISPR technologies on tomato breeding, providing a roadmap for the development of tomato to ensure sustainable agriculture and global food security in a changing climate (
Fig. 1).
CRISPR/Cas9 and Advanced Precise Editing Systems in Tomato
Fundamental Mechanisms: NHEJ and HDR
CRISPR/Cas9 remains the most widely used genome editing technology in tomato research and breeding programs because of its simplicity, versatility, and high editing efficiency (
Wada et al. 2020). The conventional CRISPR/Cas9 system functions by generating a DNA double-strand break (DSB) at a target genomic locus through Cas9 nuclease activity guided by single-guide RNA (sgRNA) (
Jinek et al. 2012;
Pacesa et al. 2024). The DSB is subsequently repaired through endogenous cellular pathways, primarily non-homologous end joining (NHEJ) or homology-directed repair (HDR) (
Pacesa et al. 2024;
Vu et al. 2020). NHEJ frequently introduces insertion/deletion mutations (indels), which can disrupt gene function and create knockout mutants (
Larriba et al. 2024;
Malzahn et al. 2017;
Xia et al. 2021). Because of its robustness and ease of design, DSB-mediated CRISPR/Cas9 technology has become the foundation for most tomato genome editing studies reported (
Fig. 1).
Tomato is considered one of the best model crops for CRISPR optimization because of its relatively efficient
Agrobacterium-mediated transformation system and short generation cycle. CRISPR/Cas9-mediated DSB editing has been extensively used for improving fruit size, yield, shelf life, disease resistance, stress tolerance, and nutritional quality in tomato (
Nie et al. 2022;
Tran et al. 2023;
Yang et al. 2024).
Recent studies particularly focused on multiplex genome editing in which several genes are simultaneously targeted using multiple sgRNAs. Multiplex editing allows rapid modification of complex quantitative traits regulated by several loci. Researchers successfully edited genes associated with flowering, carotenoid biosynthesis, fruit ripening, and plant architecture in tomato (
Munir et al. 2024;
Uranga et al. 2024;
Yang et al. 2023). In addition, HDR-mediated precise insertion strategies have been optimized in tomato although HDR efficiency remains relatively low compared with NHEJ (
Van Vu et al. 2026;
Vu et al. 2020;
Vu et al. 2024). New geminiviral replicon systems and engineered repair templates significantly improved HDR frequencies in tomato tissues (
Dahan-Meir et al. 2018;
Larriba et al. 2024;
Tiwari et al. 2023).
Another major achievement in tomato is the development of high-fidelity Cas9 variants that minimize off-target mutations (
Movahedi et al. 2022). Off-target editing has been a significant limitation of DSB-mediated systems because unintended cleavage may alter unrelated genomic regions. High-fidelity Cas9 nucleases and optimized sgRNA design algorithms substantially reduced off-target activity in tomato. Furthermore, researchers developed transgene-free edited tomato plants through transient Cas9 expression systems, enabling more acceptable regulatory frameworks for commercial breeding. These advances demonstrate that DSB-based CRISPR/Cas9 remains the central platform for tomato functional genomics and precision breeding (
Danilo et al. 2019;
Zhang et al. 2020).
Although DSB-mediated CRISPR/Cas9 systems are highly efficient, the generation of double-strand breaks may induce unintended insertions, chromosomal rearrangements, or large deletions. To overcome these limitations, base editing and prime editing technologies were developed as next-generation precision genome editing platforms (
Kosicki et al. 2018;
Leibowitz et al. 2021). Base editors enable direct nucleotide conversion without generating DSBs. Cytosine base editors (CBEs) convert C⋅G base pairs into T⋅A, whereas adenine base editors (ABEs) convert A⋅T into G⋅C. These systems typically consist of catalytically impaired Cas proteins fused with deaminase enzymes (
Komor et al. 2016).
Crop species such as rice and tomato have become one of the most important model crops for optimizing plant base editing systems. Researchers successfully used CBEs and ABEs to modify genes associated with fruit ripening, carotenoid accumulation, disease resistance, and plant morphology (
Shimatani et al. 2017). Compared with DSB-mediated knockout approaches, base editing allows subtle modification of target genes while maintaining overall genome stability (
Gaudelli et al. 2017). This advantage is particularly important for quantitative trait improvement where precise amino acid substitutions are preferred over complete gene disruption.
Prime editing represents an even more advanced genome editing technology because it can induce all twelve possible base substitutions as well as small insertions and deletions without donor DNA or DSB formation. Prime editors consist of Cas9 nickase fused to reverse transcriptase and utilize prime editing guide RNA (pegRNA). Tomato studies demonstrated that prime editing efficiency can be substantially improved through pegRNA optimization, codon optimization, and engineered reverse transcriptase variants (
Anzalone et al. 2019;
Lin et al. 2021).
Recent tomato research particularly emphasized optimization of editing efficiency because prime editing initially showed relatively low activity in dicot plants (
Vu et al. 2024). New PE5- and twin prime editing-derived systems significantly enhanced editing frequencies in tomato callus and regenerated plants (
Lou et al. 2025;
Van Vu et al. 2026). Researchers also reported successful multiplex prime editing in tomato, suggesting that complex trait engineering may become feasible in future breeding programs. Moreover, prime editing minimized unintended indels and off-target effects compared with DSB-based editing systems. These advantages make prime editing one of the most promising technologies for future precision tomato breeding (
Arabi et al. 2026;
Van Vu et al. 2026).
Applications in Tomato Engineering
Enhancing Productivity
Recent advances in CRISPR/Cas-based genome editing have significantly transformed tomato productivity and breeding strategies by enabling precise modification of genes controlling yield, flowering, plant architecture, and adaptation to modern cultivation systems. Unlike conventional breeding, targeted genome editing allows rapid optimization of agronomic traits while minimizing linkage drag (
Huang et al. 2025). Current approaches focus on improving yield components, engineering compact growth habits for indoor agriculture, and redesigning plant architecture for high-density planting and mechanized harvesting. The advancement of tomato productivity relies heavily on the precise regulation of meristem activity and fruit development. A central target for genetic improvement is the CLAVATA3 (CLV3)-WUSCHEL (WUS) feedback loop, which maintains stem cell homeostasis. Research has shown that mutations in tomato orthologs such as
lc (
locule number/
WUS) and
fas (
fasciated/
CLV3) enlarge floral meristems, significantly increasing locule number and overall fruit size (
Chu et al. 2019;
Muños et al. 2011). Beyond natural mutations, CRISPR/Cas9 technology has been employed to edit the cis-regulatory (promoter) regions of
CLV3 (
Lou et al. 2025). This approach demonstrates that fine-tuning the expression of these genes can optimize yield-related traits by adjusting meristem size without causing the severe developmental defects often associated with protein-coding mutations (
Ciren et al. 2024;
Rodríguez-Leal et al. 2017).
Inflorescence architecture also plays a vital role in determining fruit output. The MADS-box transcription factor
STM3 (a
SOC1 homolog) positively regulates branching and flowering in tomato; elevating its expression balances vegetative and reproductive growth, thereby enhancing total reproductive output (
Wang et al. 2021). Similarly, the manipulation of flowering regulators—including
SELF PRUNING (
SP),
SELF PRUNING 5G (
SP5G), and
SINGLE FLOWER TRUSS (
SFT)—has enabled the development of varieties tailored for modern industrial agriculture (
Lim et al. 2025). Specifically, the
sp mutation converts indeterminate growth into a determinate stem growth, which synchronizes flowering and fruit ripening to facilitate mechanized harvesting (
Kang et al. 2022;
Lee et al. 2023;
Lifschitz et al. 2006;
Park et al. 2014). Furthermore, editing the floral repressor
SP5G has allowed for the creation of day-neutral cultivars that remain productive regardless of day length (
Soyk et al. 2017).
Genome editing is also a powerful tool for designing tomato optimized for urban and indoor farming systems. By simultaneously editing
SP,
SP5G, and
SlER, researchers have generated compact, early-yielding plants suitable for high-density vertical farming without significant yield penalties (
Kwon et al. 2020). This multiplex CRISPR strategy has even been applied to wild tomato relatives; by targeting
SP,
OVATE, and
FW2.2, scientists have successfully improved fruit size, number, and plant morphology in a single generation. This “de novo domestication” preserves the resilience of wild species while achieving the productivity of commercial varieties (Zsögön et al. 2017). Additionally, fruit shape and elongation are precisely managed through the interaction of
OVATE,
SUN, and
SlOFP20, allowing for the customization of fruit morphology (
Xiao et al. 2008;
Zhang et al. 2023).
Finally, enhancing harvest efficiency and resource allocation further boosts the commercial value of tomato crops. Manipulating
SPL13, which functions downstream of strigolactone signaling, helps reduce excessive lateral branching and improves the allocation of nutrients toward fruit production (
Chen et al. 2023;
Jeon et al. 2025). The strategic combination of
jointless2 (
j2) and
enhancer of j2 (
ej2) mutations maximizes harvestable yield by eliminating the pedicel abscission zone for mechanical harvesting while suppressing unproductive excessive branching (
Alonge et al. 2020;
Soyk et al. 2017). Productivity is further enhanced by re-engineering canopy architecture; for instance, regulating the
narrow leaf angle (
nla) mutant produces plants with upright leaves. This modification improves light interception and allows for higher planting density, ultimately maximizing the yield potential per unit area (
Cui et al. 2026;
Fig. 1).
Abiotic stresses, such as drought, salinity, high temperatures, and low temperatures, which are becoming more frequent due to climate change, disrupt plant physiological processes and pose a serious threat to the productivity and yield stability of tomato. In this context, CRISPR/Cas9 genome editing technology has established itself as the most effective and innovative alternative for maximizing the environmental adaptability of tomato by overcoming the limitations of conventional crossbreeding, such as long breeding periods and gene linkages, and by precisely correcting genes involved in stress perception, signal transduction, and protective responses (
Keerthana et al. 2025).
Heat stress causes intracellular oxidative damage and disrupts cellular homeostasis in tomato plants. CRISPR/ Cas9-mediated studies revealed that SlMAPK3 plays an important role in regulating reactive oxygen species (ROS) homeostasis during heat stress, and its disruption significantly alters tomato thermotolerance (
Yu et al. 2019). Additionally, studies in which the
SlNPR1, involved in maintaining cell membrane stability and homeostasis, was inactivated confirmed stomatal enlargement and water loss, thereby elucidating their defensive roles (
Li et al. 2019). Conversely, there have been reported cases where drought tolerance was successfully enhanced to positively strengthen crop tolerance by increasing water use efficiency without reducing yield through the inhibition of the gibberellin receptor
GID1a (
Illouz-Eliaz et al. 2020), or by inducing a decrease in stomatal density and epidermal cell expansion through the regulation of the
SlTLFP8 (
Li et al. 2020). Problems regarding osmotic imbalance and toxicity caused by salt accumulation in the soil are also being overcome through strategies to block negative regulatory factors using CRISPR/Cas9. When
SlHyPRP1, which inhibits abscisic acid (ABA) signaling, or
ARF4, an auxin-responsive factor, are knocked out, transpiration decreases and root development is promoted, leading to a dramatic increase in the plant's ability to regulate osmotic pressure. Conversely, the mechanism of ion homeostasis has been demonstrated by showing that the removal of the
SlSOS1, which contributes to sodium excretion, results in a fatal response to salt (
Wang et al. 2020). Recently, groundbreaking breeding strategies utilizing multiplexing techniques to simultaneously improve various stress-related traits have also been proposed (
Tran et al. 2023).
Furthermore, high-temperature stress caused by global warming drastically reduces pollen survival rates and fruit set rates in tomato; however, recent genome editing techniques protect reproductive capacity by targeting heat shock transcription factors (HSFs) or correcting promoters to induce the expression of heat shock proteins (HSPs) (
Yu et al. 2019). In particular, research that enhanced transcriptional responsiveness by directly inserting heat shock elements (HSE) into endogenous promoters using state-of-the-art prime editing (
Lou et al. 2025), and the achievement of enabling seedless fruit production through parthenocarpy-fruiting even at extreme high temperatures of 38℃ by editing the
SlAGL6 (
Klap et al. 2017) have very high agricultural utility value. In addition, the importance of calcium and brassinosteroid signaling pathways in response to high temperatures has been confirmed through loss-of-function models of
cpk28 kinase and
BZR1 transcription factor. Furthermore, editing of the
HyPRP1, which previously contributed to improved salt tolerance, was shown to simultaneously exhibit excellent efficacy in defending against heat shock at 42-45℃ (
Tran et al. 2021;
Tran et al. 2023). Including studies on defense mechanisms against cold stress in the 0-15℃ range (
Weiss et al. 2009), CRISPR/Cas9-mediated genome editing technology will become a core platform for sustainable future agriculture that preserves tomato yields and quality in the face of complex climate disasters.
Bacterial, fungal, and viral diseases are major constraints limiting tomato productivity, fruit quality, and postharvest stability worldwide (
Mansfield et al. 2012). Bacterial pathogens such as
Xanthomonas spp.,
Pseudomonas syringae pv.
tomato,
Ralstonia solanacearum, and
Clavibacter michiganensis subsp.
michiganensis cause chlorosis, necrosis, wilting, and fruit lesions that significantly reduce marketability and storage quality (
Fan et al. 2022;
Wang et al. 2025). CRISPR/Cas-mediated genome editing has emerged as an effective strategy for improving disease resistance while maintaining agronomic performance and fruit quality. One of the best-characterized targets is the susceptibility gene
SlDMR6-1, whose disruption enhances resistance to bacterial spot, bacterial speck, and late blight through activation of salicylic acid-mediated defense pathways without severe growth penalties (
Thomazella et al. 2021). Edited plants exhibit reduced disease symptoms, improved fruit integrity, delayed tissue deterioration, and enhanced shelf life.
Additional studies demonstrated that editing susceptibility-associated genes such as
SlBSs and
SlBSsL, orthologs of pepper
CaBSs and
CaBSsL, increases resistance against
Xanthomonas infection in tomato (
Ortega et al. 2024). Unlike conventional resistance breeding, which often depends on single resistance genes vulnerable to pathogen evolution, CRISPR-mediated susceptibility-gene editing may provide broader and more durable resistance. Recent studies also highlighted that promoter and cis-regulatory editing can reduce the growth-defense tradeoff associated with constitutive immune activation (
Wang et al. 2025). Furthermore, CRISPR activation (CRISPRa) systems have recently emerged as promising tools for inducing endogenous defense responses without permanently altering coding sequences (
McLaughlin et al. 2025).
Fungal diseases including powdery mildew, Fusarium wilt, gray mold, and late blight remain serious threats to tomato production, particularly under greenhouse conditions and climate change-associated humidity stress (
Elad et al. 2014). Powdery mildew reduces photosynthetic efficiency through chlorosis and premature senescence, whereas
Fusarium oxysporum disrupts vascular transport, causing severe wilting and plant death (
Michielse et al. 2009). Genome editing targeting fungal susceptibility pathways improves canopy health, photosynthetic activity, fruit set, and yield stability while reducing postharvest losses caused by tissue decay and secondary microbial infection.
Viral pathogens such as Tomato yellow leaf curl virus (TYLCV), Tomato mosaic virus (ToMV), and Tomato spotted wilt virus (TSWV) also severely impair tomato productivity and fruit quality. Among these, TYLCV is particularly destructive because it induces leaf curling, growth retardation, flower abortion, and major yield reduction. The recessive resistance gene
ty-5, which encodes the pelota protein associated with viral susceptibility, is one of the most important resistance loci against TYLCV (
Lapidot et al. 2015). CRISPR/Cas systems targeting
Ty-5-associated pathways and viral replication regions have shown strong potential for engineering durable viral resistance and suppressing systemic viral accumulation (
Tashkandi et al. 2018). Enhanced viral resistance improves fruit uniformity, sugar accumulation, color development, and overall market quality by preventing virus-induced developmental abnormalities.
Collectively, recent advances in CRISPR/Cas-based genome editing demonstrate strong potential for simultaneously improving disease resistance, productivity, fruit quality, and postharvest performance in tomato. Multiplex genome editing and cis-regulatory engineering are expected to further accelerate the development of climate-resilient and high-value tomato cultivars suitable for sustainable agriculture and next-generation farming systems (
Fig. 1).
Modern genetic improvement of tomato has expanded beyond increasing yield to restoring flavor and enhancing health-promoting phytochemicals. Decades of intensive breeding for productivity inadvertently led to the loss of complex flavor profiles found in heirloom varieties. To address this, researchers have utilized whole-genome sequencing along with transcriptomic and metabolomic analyses to understand the metabolic pathways of sugars, organic acids, and over 400 volatile organic compounds. These comprehensive studies have established a robust foundation for precision engineering aimed at elevating tomato flavor and functional nutrients (
Du et al. 2025;
Pereira et al. 2021).
Leveraging this understanding, the CRISPR/Cas9 system is being extensively used to upregulate the biosynthesis of high-value compounds, particularly antioxidants and vitamins. A primary focus is the enhancement of lycopene, a potent natural antioxidant. Historically, breeders sought to increase lycopene levels by inducing mutations in the
CYC-B, which is responsible for lycopene catabolism. However, a major bottleneck existed due to the tight genetic linkage between the
CYC-B locus and
SP locus. To circumvent this, researchers employed CRISPR/Cas9 to directly knock out
CYC-B in elite indeterminate genotypes (LT16 and LT46), successfully producing high-lycopene tomato without compromising plant architecture (
Arruabarrena et al. 2023).
Recent breakthroughs have further extended to the biofortification of essential vitamins, notably Vitamin A and Vitamin D. By utilizing the CRISPR/Cas system, scientists targeted the
SlLCYe and
SlBCH to prevent the degradation of beta-carotene, a vital provitamin A precursor, resulting in a significant accumulation of this nutrient (
Liu et al. 2025).
More remarkably, researchers have successfully engineered tomato to serve as a plant-based source of Vitamin D3. By knocking out the
Sl7DR2 (
7-dehydrocholesterol reductase), which normally converts the Vitamin D3 precursor provitamin D3 (7-DHC) into cholesterol, scientists allowed 7-DHC to accumulate in the leaves and fruit. Upon exposure to UVB light, this precursor is converted into Vitamin D3, providing a sustainable dietary solution for global Vitamin D deficiency (
Li et al. 2022).
Beyond vitamins, the production of specialized metabolites like saponins and GABA has seen significant progress. In certain tomato species, metabolic engineering of the
GAME (
Glycoalkaloid Metabolic Engineering) genes—specifically targeting the cytochrome P450 enzymes and glycosyltransferases—has allowed for the precise modulation of steroidal saponins and alkaloids (
Lim et al. 2026). These compounds are highly valued for their anti-inflammatory, antimicrobial, and anticancer properties (
Cárdenas et al. 2019;
Itkin et al. 2013). Furthermore, genetic editing has successfully increased the levels of gamma-aminobutyric acid (GABA), an inhibitory neurotransmitter that helps lower blood pressure. By using CRISPR/Cas9 to delete the autoinhibitory domain of glutamate decarboxylase (GAD), researchers achieved a massive increase in GABA content in the fruit (
Nonaka et al. 2017). These GABA-enriched tomato represent a major milestone, as they have already successfully transitioned from the laboratory to the commercial market as the world's first CRISPR-edited food (
Waltz 2022).
Conclusion and Future Perspectives
Recent advances in CRISPR/Cas-based genome editing technologies have fundamentally transformed tomato breeding by enabling precise, rapid, and efficient modification of genes associated with productivity, stress resilience, nutritional quality, and plant architecture. Compared with conventional breeding approaches, genome editing provides unprecedented flexibility for introducing desirable traits while minimizing linkage drag and reducing breeding time. In particular, the integration of CRISPR/Cas9, base editing, and prime editing technologies has expanded the scope of tomato improvement beyond simple gene knockouts toward highly precise nucleotide substitutions, promoter engineering, multiplex editing, and transgene-free breeding strategies. These technological developments have accelerated both functional genomics research and practical cultivar development.
This review highlights the broad applications of genome editing in tomato, including enhancement of abiotic and biotic stress tolerance, optimization of flowering and yield-related traits, redesign of plant architecture for indoor and vertical farming systems, and metabolic engineering for value-added compounds such as lycopene, β-carotene, and GABA. Recent studies further demonstrate that editing cis-regulatory elements and developmental pathways can fine-tune quantitative traits without severe pleiotropic effects, providing new opportunities for sustainable crop improvement. Moreover, advances in compact growth habits, mechanized harvesting traits, and resource-efficient architectures support the development of “smart tomato” cultivars tailored for next-generation agricultural systems (
Fig. 1).
Despite these achievements, several challenges remain before genome-edited tomato can be fully integrated into global agricultural production systems. Editing efficiency, particularly for HDR and prime editing-based approaches, still requires optimization in tomato tissues. Off-target mutations, genotype-dependent transformation efficiency, and regulatory uncertainty in different countries continue to limit commercialization. Furthermore, the long-term durability of disease resistance and stress tolerance under field conditions requires additional evaluation across diverse environmental backgrounds.
Future research should therefore focus on improving delivery systems, expanding transgene-free editing platforms, and integrating artificial intelligence, multi-omics analyses, and speed breeding technologies to accelerate precision breeding. In addition, multiplex genome editing combined with cis-regulatory engineering will likely become a central strategy for simultaneously improving yield, nutritional quality, climate resilience, and adaptability to controlled-environment agriculture. Ultimately, CRISPR-based precision breeding offers enormous potential to develop sustainable, high-value tomato cultivars capable of supporting future food security and climate-resilient agriculture.
Acknowledgments
This work was supported by the Research Sabbatical Grant for Research Professors from Gyeongsang National University (2025; No. GNU-SGRP-0009) and by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (No. RS-2025-16070781).
Fig. 1Systematic workflow of precision tomato breeding and sustainability using CRISPR-Cas9 technology. The schematic illustrates a systematic workflow starting with global tomato breeding challenges (top brown color box) that drive innovation. Next, CRISPR-based precision breeding solutions (middle), utilizing the Cas9 protein and cellular repair pathways like NHEJ or HDR, are applied. These advanced technologies enable the direct engineering of desired traits (lower brown color box), including selection of optimal plant growth and harvest, and adaptation for CEA. Ultimately, this approach leads to sustainable agriculture & food security, demonstrated by the superior developmental timeline of precision breeding compared to conventional methods (bottom right prediction).
References
- Alonge M, Wang X, Benoit M, Soyk S, Pereira L, Zhang L, et al. 2020. Major impacts of widespread structural variation on gene expression and crop improvement in tomato. Cell. 182(1): 145-161.
- Anzalone AV, Randolph PB, Davis JR, Sousa AA, Koblan LW, Levy JM, et al. 2019. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature. 576(7785): 149-157.
- Arabi M, Alizadeh F, Yousefi Y, Afarandeh H, Jovin SM, Eslahi A, et al. 2026. Enhancement of prime editing by recruiting engineered or evolved components and implementing novel strategies. Biochem. Biophys. Rep. 45: 102495
- Arruabarrena A, Lado J, González-Arcos M, Vidal S. 2023. Targeted disruption of tomato chromoplast-specific lycopene β-cyclase (CYC-B) gene promotes early accumulation of lycopene in fruits and enhanced postharvest cold tolerance. Plant Biotechnol. J. 21(12): 2420
- Cárdenas PD, Sonawane PD, Heinig U, Jozwiak A, Panda S, Abebie B, et al. 2019. Pathways to defense metabolites and evading fruit bitterness in genus Solanum evolved through 2-oxoglutarate-dependent dioxygenases. Nat. Commun. 10(1): 5169
- Chen S, Song X, Zheng Q, Liu Y, Yu J, Zhou Y, et al. 2023. The transcription factor SPL13 mediates strigolactone suppression of shoot branching by inhibiting cytokinin synthesis in Solanum lycopersicum. J. Exp. Bot. 74(18): 5722-5735.
- Chu YH, Jang JC, Huang Z, Van der Knaap E. 2019. Tomato locule number and fruit size are controlled by natural alleles of lc and fas. Plant Direct. 3(7): e00142
- Ciren D, Zebell S, Lippman ZB. 2024. Extreme restructuring of cis-regulatory regions controlling a deeply conserved plant stem cell regulator. PLoS Genet. 20(3): e1011174
- Cui L, Zheng F, Jia L, Hu F, Ai G, Ye J, et al. 2026. SPL13 controls tomato lateral branch outgrowth by regulating brassinosteroid biosynthesis and signal transduction. Hortic. Res. 13(4): uhag007
- Dahan-Meir T, Filler-Hayut S, Melamed-Bessudo C, Bocobza S, Czosnek H, Aharoni A, et al. 2018. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J. 95(1): 5-16.
- Danilo B, Perrot L, Mara K, Botton E, Nogué F, Mazier M. 2019. Efficient and transgene-free gene targeting using Agrobacterium-mediated delivery of the CRISPR/Cas9 system in tomato. Plant Cell Rep. 38(4): 459-462.
- Du M, Sun C, Deng L, Zhou M, Li J, Du Y, et al. 2025. Molecular breeding of tomato: Advances and challenges. J. Integr. Plant Biol. 67(3): 669-721.
- Elad Y, Pertot I. 2014. Climate change impacts on plant pathogens and plant diseases. J. Crop Improv. 28(1): 99-139.
- Fan X, Saleem T, Zou H. 2022. Copper resistance mechanisms in plant pathogenic bacteria. Phytopathol. Mediterr. 61: 129-138.
- Gaudelli NM, Komor AC, Rees HA, Packer MS, Badran AH, Bryson DI, et al. 2017. Programmable base editing of A⋅T to G⋅C in genomic DNA without DNA cleavage. Nature. 551(7681): 464-471.
- Huang X, Xiao N, Xie Y, Xu C. 2025. ROS burst prolongs transcriptional condensation to slow shoot apical meristem maturation and achieve heat-stress resilience in tomato. Dev. Cell. 60(15): 2032-2045.
- Illouz-Eliaz N, Nissan I, Nir I, Ramon U, Shohat H, Weiss D. 2020. Mutations in the tomato gibberellin receptors suppress xylem proliferation and reduce water loss under water-deficit conditions. J. Exp. Bot. 71(12): 3603-3612.
- Itkin M, Heinig U, Tzfadia O, Bhide A, Shinde B, Cardenas P, et al. 2013. Biosynthesis of antinutritional alkaloids in solanaceous crops is mediated by clustered genes. Science. 341(6142): 175-179.
- Jeon Y-J, Hong S, Lee TS, Park SH, Song G, Seo M-G, et al. 2025. Volumetric deep learning-based precision phenotyping of gene-edited tomato for vertical farming. Plant Phenomics. 7(3): 100095
- Jeong YY, Hong C, Han JH, Bae S, Seo PJ. 2025. Development of a prime editor with improved editing efficiency in Arabidopsis. BMB Rep. 58(2): 70-74.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. 2012. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 337(6096): 816-821.
- Kang M-S, Kim YJ, Heo J, Rajendran S, Wang X, Bae JH, et al. 2022. Newly discovered alleles of the tomato antiflorigen gene SELF PRUNING provide a range of plant compactness and yield. Int. J. Mol. Sci. 23(13): 7149
- Keerthana R, Rakshana P, Salunkhe SR, Sakthi AR, Kokiladevi E, Saraswathi T, et al. 2025. CRISPR-Cas9 mediated enhancement of abiotic stress resilience in tomato: A comprehensive review of target genes. Mol. Biol. Rep. 52(1): 538
- Klap C, Yeshayahou E, Bolger AM, Arazi T, Gupta SK, Shabtai S, et al. 2017. Tomato facultative parthenocarpy results from Sl AGAMOUS-LIKE 6 loss of function. Plant Biotechnol. J. 15(5): 634-647.
- Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR. 2016. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature. 533(7603): 420-424.
- Kosicki M, Tomberg K, Bradley A. 2018. Repair of double-strand breaks induced by CRISPR-Cas9 leads to large deletions and complex rearrangements. Nat. Biotechnol. 36(8): 765-771.
- Kwon C-T, Heo J, Lemmon ZH, Capua Y, Hutton SF, Van Eck J, et al. 2020. Rapid customization of Solanaceae fruit crops for urban agriculture. Nat. Biotechnol. 38(2): 182-188.
- Lapidot M, Karniel U, Gelbart D, Fogel D, Evenor D, Kutsher Y, et al. 2015. A novel route controlling begomovirus resistance by the messenger RNA surveillance factor pelota. PLoS Genet. 11(10): e1005538
- Larriba E, Yaroshko O, Pérez-Pérez JM. 2024. Recent advances in tomato gene editing. Int. J. Mol. Sci. 25(5): 2606
- Lee ES, Heo J, Bang WY, Chougule KM, Waminal NE, Hong NT, et al. 2023. Engineering homoeologs provide a fine scale for quantitative traits in polyploid. Plant Biotechnol. J. 21(12): 2458-2472.
- Leibowitz ML, Papathanasiou S, Doerfler PA, Blaine LJ, Sun L, Yao Y, et al. 2021. Chromothripsis as an on-target consequence of CRISPR-Cas9 genome editing. Nat. Genet. 53(6): 895-905.
- Li J, Scarano A, Gonzalez NM, D'Orso F, Yue Y, Nemeth K, et al. 2022. Biofortified tomatoes provide a new route to vitamin D sufficiency. Nat. Plants. 8(6): 611-616.
- Li R, Liu C, Zhao R, Wang L, Chen L, Yu W, et al. 2019. CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 19(1): 38
- Li S, Zhang J, Liu L, Wang Z, Li Y, Guo L, et al. 2020. SlTLFP8 reduces water loss to improve water-use efficiency by modulating cell size and stomatal density via endoreduplication. Plant Cell Environ. 43(11): 2666-2679.
- Lifschitz E, Eviatar T, Rozman A, Shalit A, Goldshmidt A, Amsellem Z, et al. 2006. The tomato FT ortholog triggers systemic signals that regulate growth and flowering and substitute for diverse environmental stimuli. Proc. Natl. Acad. Sci. U. S. A. 103(16): 6398-6403.
- Lim J, Kim K, Heo J, Seo SM, Choung S, Kim H, et al. 2026. Rewiring steroidal metabolic pathways for diosgenin production in solanum nigrum. Plant Biotechnol. J. 24(5): 3032-3047.
- Lim Y, Seo M-G, Lee J, Hong S, An J-T, Jeong H-Y, et al. 2025. Optimizing plant size for vertical farming by editing stem length regulators. Plant Biotechnol. J. 23(8): 3041-3053.
- Lin Q, Jin S, Zong Y, Yu H, Zhu Z, Liu G, et al. 2021. High-efficiency prime editing with optimized, paired pegRNAs in plants. Nat. Biotechnol. 39(8): 923-927.
- Liu X, Fu Y, Li M, Xiong S, Huang L, Zhang S, et al. 2025. Biofortification of tomatoes with beta-carotene through targeted gene editing. Int. J. Biol. Macromol. 327(2): 147396
- Lou H, Li S, Shi Z, Zou Y, Zhang Y, Huang X, et al. 2025. Engineering source-sink relations by prime editing confers heat-stress resilience in tomato and rice. Cell. 188(2): 530-549.
- Malzahn A, Lowder L, Qi Y. 2017. Plant genome editing with TALEN and CRISPR. Cell Biosci. 7(1): 21
- Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, Ronald P, et al. 2012. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 13(6): 614-629.
- McLaughlin JE, Kue Foka IC, Lawton MA, Di R. 2025. CRISPR activation: Identifying and using novel genes for plant disease resistance breeding. Front. Genome Ed. 7: 1596600
- Michielse CB, Rep M. 2009. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 10(3): 311-324.
- Movahedi A, Hajiahmadi Z, Wei H, Yang L, Ruan H, Zhuge Q. Mysore KS, Senthil-Kumar M, 2022. A method to reduce off-targets in CRISPR/Cas9 system in plants. editors. Plant Gene Silencing: Methods and Protocols. Springer, Humana. New York, NY: pp. 317-324.
- Munir A, Amin I, Zahoor MK, Majeed HN, Almoammar H, Ghaffar A, et al. Abd-Elsalam KA, Ahmad A, Zhang B, 2024. Multiplex genome editing in plants through CRISPR-Cas. editors. Plant Gene Silencing: Methods and Protocols. Elsevier. Amsterdam, The Netherlands: pp. 127-142.
- Muños S, Ranc N, Botton E, Bérard A, Rolland S, Duffé P, et al. 2011. Increase in tomato locule number is controlled by two single-nucleotide polymorphisms located near WUSCHEL. Plant Physiol. 156(4): 2244-2254.
- Nie H, Shi Y, Geng X, Xing G. 2022. CRISRP/Cas9-mediated targeted mutagenesis of tomato polygalacturonase gene (SlPG) delays fruit softening. Front. Plant Sci. 13: 729128
- Nonaka S, Arai C, Takayama M, Matsukura C, Ezura H. 2017. Efficient increase of ɣ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 7(1): 7057
- Ortega A, Seong K, Schultink A, de Toledo Thomazella DP, Seo E, Zhang E, et al. 2024. CRISPR/Cas9-mediated editing of Bs5 and Bs5L in tomato leads to resistance against Xanthomonas. Plant Biotechnol. J. 22(10): 2785-2787.
- Pacesa M, Pelea O, Jinek M. 2024. Past, present, and future of CRISPR genome editing technologies. Cell. 187(5): 1076-1100.
- Park SJ, Jiang K, Tal L, Yichie Y, Gar O, Zamir D, et al. 2014. Optimization of crop productivity in tomato using induced mutations in the florigen pathway. Nat. Genet. 46(12): 1337-1342.
- Pereira L, Sapkota M, Alonge M, Zheng Y, Zhang Y, Razifard H, et al. 2021. Natural genetic diversity in tomato flavor genes. Front. Plant Sci. 12: 642828
- Rodríguez-Leal D, Lemmon ZH, Man J, Bartlett ME, Lippman ZB. 2017. Engineering quantitative trait variation for crop improvement by genome editing. Cell. 171(2): 470-480.
- Shimatani Z, Kashojiya S, Takayama M, Terada R, Arazoe T, Ishii H, et al. 2017. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 35(5): 441-443.
- Soyk S, Müller NA, Park SJ, Schmalenbach I, Jiang K, Hayama R, et al. 2017. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 49(1): 162-168.
- Tashkandi M, Ali Z, Aljedaani F, Shami A, Mahfouz MM. 2018. Engineering resistance against Tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato. Plant Signal Behav. 13(10): e1525996
- Thomazella DPdT, Seong K, Mackelprang R, Dahlbeck D, Geng Y, Gill US, et al. 2021. Loss of function of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. Proc. Natl. Acad. Sci. U. S. A. 118(27): e2026152118
- Tiwari JK, Singh AK, Behera TK. 2023. CRISPR/Cas genome editing in tomato improvement: Advances and applications. Front. Plant Sci. 14: 1121209
- Tran MT, Doan DTH, Kim J, Song YJ, Sung YW, Das S, et al. 2021. CRISPR/Cas9-based precise excision of SlHyPRP1 domain (s) to obtain salt stress-tolerant tomato. Plant Cell Rep. 40(6): 999-1011.
- Tran MT, Son GH, Song YJ, Nguyen NT, Park S, Thach TV, et al. 2023. CRISPR-Cas9-based precise engineering of SlHyPRP1 protein towards multi-stress tolerance in tomato. Front. Plant Sci. 14: 1186932
- Uranga M, Aragonés V, García A, Mirabel S, Gianoglio S, Presa S, et al. 2024. RNA virus-mediated gene editing for tomato trait breeding. Hortic. Res. 11(1): uhad279
- Van Vu T, Thi Nguyen N, Kim J, Hoai Nguyen T, Kim J-Y. 2026. Development of an ultra-efficient prime editing system in tomato. Nat. Commun. 17(1): 95
- Vu TV, Nguyen NT, Kim J, Song YJ, Nguyen TH, Kim J-Y. 2024. Optimized dicot prime editing enables heritable desired edits in tomato and Arabidopsis. Nat. Plants. 10(10): 1502-1513.
- Vu TV, Sivankalyani V, Kim EJ, Doan DTH, Tran MT, Kim J, et al. 2020. Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato. Plant Biotechnol. J. 18(10): 2133-2143.
- Wada N, Ueta R, Osakabe Y, Osakabe K. 2020. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 20(1): 234
- Waltz E. 2022. GABA-enriched tomato is first CRISPR-edited food to enter market. Nat. Biotechnol. 40(1): 9-11.
- Wang D, Mandal P, Rahman MS, Yang L. 2025. Engineering tomato disease resistance by manipulating susceptibility genes. Front. Genome Ed. 7: 1537148
- Wang S, Qiang Q, Xiang L, Fernie AR, Yang J. 2023. Targeted approaches to improve tomato fruit taste. Hortic. Res. 10(1): uhac229
- Wang X, Liu Z, Sun S, Wu J, Li R, Wang H, et al. 2021. SISTER OF TM3 activates FRUITFULL1 to regulate inflorescence branching in tomato. Hortic. Res. 8: 251
- Wang Z, Hong Y, Li Y, Shi H, Yao J, Liu X, et al. 2020. Natural variations in SlSOS1 contribute to the loss of salt tolerance during tomato domestication. Plant Biotechnol. J. 19(1): 20
- Weiss J, Egea-Cortines M. 2009. Transcriptomic analysis of cold response in tomato fruits identifies dehydrin as a marker of cold stress. J. Appl. Genet. 50(4): 311-319.
- Xia X, Cheng X, Li R, Yao J, Li Z, Cheng Y. 2021. Advances in application of genome editing in tomato and recent development of genome editing technology. Theor. Appl. Genet. 134(9): 2727-2747.
- Xiao H, Jiang N, Schaffner E, Stockinger EJ, Van der Knaap E. 2008. A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit. Science. 319(5869): 1527-1530.
- Yang Q, Cai L, Wang M, Gan G, Li W, Li W, et al. 2024. CRISPR/cas9 allows for the quick improvement of tomato firmness breeding. Curr. Issues Mol. Biol. 47(1): 9
- Yang T, Ali M, Lin L, Li P, He H, Zhu Q, et al. 2023. Recoloring tomato fruit by CRISPR/Cas9-mediated multiplex gene editing. Hortic. Res. 10(1): uhac214
- Yu W, Wang L, Zhao R, Sheng J, Zhang S, Li R, et al. 2019. Knockout of SlMAPK3 enhances tolerance to heat stress involving ROS homeostasis in tomato plants. BMC Plant Biol. 19(1): 354
- Zhang BY, Li Q, Keyhaninejad N, Taitano N, Sapkota M, Snouffer A, et al. 2023. A combinatorial TRM-OFP module bilaterally fine-tunes tomato fruit shape. New Phytol. 238(6): 2393-2409.
- Zhang N, Roberts HM, Van Eck J, Martin GB. 2020. Generation and molecular characterization of CRISPR/Cas9-induced mutations in 63 immunity-associated genes in tomato reveals specificity and a range of gene modifications. Front. Plant Sci. 11: 10
- Zhang Q, Zhu L, Li H, Chen Q, Li N, Li J, et al. 2024. Insights and progress on the biosynthesis, metabolism, and physiological functions of gamma-aminobutyric acid (GABA): A review. PeerJ. 12: e18712