Abstract
Carbohydrates and genetic factors influence cell division in citrus fruits from ovule cultures. Here, we investigated the effects of various carbohydrate sources (sorbitol, maltose, galactose, and lactose) on cell proliferation, somatic embryogenesis, and plant regeneration in three Citrus unshiu cultivars (‘Miyagawa wase’ [MW], ‘Haryejosaeng’ [HW], and ‘Nankan 20 gou’ [NW]), and one C. reticulata cultivar (‘Ootaponkan’ [PM]). In PM, carbohydrate sources did not affect proliferation; MW showed a significant but marginal increase in media containing 300 mM mannitol. It also proliferated well in media supplemented with 300 mM sorbitol and a mixture of galactose and lactose. NW achieved the best rate with 100 mM mannitol and 146 mM lactose, whereas HW proliferated well with 100 mM mannitol alone. Phylogenetic analysis using 18 citrus varieties showed distinct genetic differences among the cultivars, suggesting that the differences in regeneration capacities in different carbohydrate sources could be attributed to their genetic differences. This study provides valuable insights for optimizing tissue culture protocols for mandarin varieties and may enhance the efficiency of citrus breeding and tissue culture applications.
-
Key words: ovule, carbohydrate sources, cell proliferation, somatic embryogenesis, plant regeneration, phylogenetics
Introduction
Citrus, a globally cultivated tropical fruit crop, belongs to the family Rutaceae, which comprises
Citrus and other genera, including
Fortunella and
Poncirus (
Kim et al. 2020;
Praveen et al. 2003).
Citrus spp. are divided into oranges, mandarins, lemons, lime, grapefruit, and pomelo (
Richa et al. 2023). Among these, mandarins (
C. reticulata) are popular for their flavor and nutritional benefits. Mandarins comprise several varieties, including satsuma mandarin (
C. unshiu Marc.;
Swingle 1943), one of the most popular types of mandarin globally. Satsuma mandarins are predominantly cultivated in Japan and Korea (
Jin et al. 2007;
Ling et al. 1990) and exhibit polyploidy and male sterility, which hinders their improvement through crossbreeding (
An et al. 2008;
Jin et al. 2020). Consequently, the enhancement of this cultivar relies on selecting bud-sport mutants (
Park et al. 2013) and seed reproduction via embryos from nucellar cell tissues around the embryo sac (
Jin et al. 2020;
Kepiro et al. 2007;
Kim et al. 2016; Zhang et al. 2018). Furthermore, variants with low citric acid and high sugar contents show genetic similarity to the maternal plants (
Liu et al. 2009;
Zhang et al. 2006), thereby complicating mutant identification based solely on morphological characteristics (
Jin et al. 2020), exhibit lower efficiency in callus formation and plant regeneration than other citrus varieties (
Han et al. 2002;
Ling et al. 1990;
Yun et al. 2006). Despite the recent advancements in shoot-tip culture (
Jin et al. 2007;
Jin et al. 2017), selecting high somatic embryo-forming satsuma mandarin lines remains challenging due to variable embryo formation depending on the state, size, and weight of the cell lines (
Han et al. 2002;
Jin et al. 2007). Only a few varieties of satsuma mandarin, such as ‘Miyagawa wase,’ ‘Haryejosaeng,’ and ‘Ishizuka wase,’ have established regeneration system to date (
Han et al. 2002;
Jin et al. 2007;
Jin et al. 2017;
Ling et al. 1990).
Somatic embryogenesis efficiency in citrus, primarily derived from nucellar tissue, is influenced by carbohydrate sources (
Gholami et al. 2013;
Tomaz et al. 2001) and genotypes (
Cardoso et al. 2014;
Starrantino et al. 1980). Carbohydrates regulate plant developmental processes (
Jin et al. 2007;
Shah et al. 2019;
Tomaz et al. 2001), with lactose (
Jin et al. 2007;
Jin et al. 2017) and high sucrose concentrations (
Ricci et al. 2002) effectively inducing somatic embryos in citrus. Sucrose hydrolyzes into fructose and glucose, while lactose comprises galactose and glucose. In citrus, glucose and fructose do not affect somatic embryo formation (
Nickell et al. 1970), whereas galactose inhibits UDP-glucose synthesis, affecting plant growth, and lactose enhances somatic embryo formation more effectively than galactose (
Gholami et al. 2013). Studies have indicated that the embryo formation rates in
C. unshiu and other
Citrus spp. with low somatic embryogenesis can be enhanced by increasing the agar concentration and using lactose as a carbohydrate source (
Jin et al. 2007;
Jin et al. 2017). However, these modifications may reduce cell proliferation and induce cell death, suggesting the need to evaluate media with a modified composition supporting rapid cell proliferation while maintaining high somatic embryogenesis rates. Sugars, such as sorbitol and maltose, although not typical carbohydrate sources for citrus, have been effective in promoting somatic embryogenesis. Maltose, in particular, is beneficial for generating glucose during metabolism and acts as an osmotic pressure agent (
Jin 2005;
Kayim et al. 2006;
Kazeem et al. 2020;
Liang et al. 2012;
Soria-Guerra et al. 2011).
Callus generally lose their regenerative ability due to somatic variation after prolonged subculturing (
Kim et al. 1995). While some citrus types, such as oranges, retain their ability to form somatic embryos and regenerate even after 10 years of subculturing (
Kochba et al. 1972), the callus of certain citrus fruits, including
C. unshiu, are influenced by the age, size, and weight of the cell line (
Han et al. 2002). To prevent the loss of these abilities from long-term subculturing, callus must be induced and propagated through new ovules and unfertilized and undeveloped seeds (
Jin et al. 2007;
Jin et al. 2017;
Jin et al. 2025). However, obtaining a new cell line using these methods is time-consuming, suggesting the need for alternatives to recover and improve the efficiency of somatic embryogenesis in such cell lines.
In this study, we aimed to explore the effects of various carbohydrates on somatic embryo proliferation and differentiation in mandarin cultivars. We also investigated the genetic relationships between the chloroplast DNA (cpDNA; trnL-trnF) and ribosomal DNA (Internal transcribed spacer [ITS]) regions and evaluated the regenerative ability of non-differentiating cells and plant cells.
Materials and Methods
Plant Materials and Callus Proliferation
Callus from four mandarin cultivars (three
C. unshiu cultivars ‘Miyagawa wase’ [MW], ‘Nankan 20 gou’ [NW], ‘Haryejosaeng’ [HW], and one
C. reticulata cultivar ‘Ootaponkan’ [PM]) were obtained via culturing immature fruit ovules. The callus were induced on MS1 medium (
Jin et al. 2017) supplemented with malt extract (500 mg/L) and sucrose (50 g/L) and cultured twice at 4-week intervals under dark conditions at 25 ± 2℃ to induce somatic embryos.
To examine the influence of the carbohydrate composition of the media on callus proliferation and somatic embryogenesis rates, callus samples (approximately 50 mg) from the four mandarin cultivars were cultured on MS basal media (
Murashige et al. 1962). The medium was supplemented with 500 mg/L malt extract and various concentrations (0, 100, 200, and 300 mM) of sorbitol, mannitol, galactose, or lactose (0, 90, 146, and 205 mM), either alone or in combination, and compared with MS1 media (
Jin et al. 2017) as a control. After 8 weeks of culture, the callus proliferation and somatic embryogenesis rates were assessed. Experiments were conducted in five replicates, and proliferation was measured at 4 (Optimal callus proliferation and subculture cycle period) and 8 (Maximum cell culture cycle and peak stage of somatic embryo formation) weeks by weighing the samples and subtracting the weight of the culture medium. Somatic embryogenesis was quantified by visually counting the number of embryos.
Each experiment included at least two biological and three technical replicates, and the data were analyzed using the SPSS software (IBM, Armonk, NY, USA). ANOVA followed by Duncan’s multiple range test was performed, with p < 0.05, indicating statistical significance.
Somatic Embryogenesis Cell Selection and Plant Regeneration
Callus with somatic embryogenesis from the four mandarin cultivars were selected using the Percoll (colloidal PVP coated with silica, Sigma-Aldrich Co., St. Louis, MO, USA) concentration gradient method (
Han et al. 2002;
Jin et al. 2007;
Jin et al. 2025), specifically from the 40% Percoll layer. These cells (approximately 50 mg) were cultured on MT basal media (
Jin et al. 2017;
Jin et al. 2025;
Murashige et al. 1969) with 500 mg/L malt extract, 70 g/L lactose, and 1.6% agar to induce callus formation. The cells were subcultured twice at 4-week intervals in the same medium to select those with high somatic embryogenesis potential.
To induce plant regeneration, selected somatic embryos were cultured on MS1 media supplemented with 0.05 M sorbitol, 0.05 M galactose, 0.5 mg/L GA3, and 0.2% gelrite for 5-6 weeks. Callus that formed roots and shoots were transferred to hormone-free MS1 media and subcultured every 4 weeks for 16 weeks to obtain normal plantlets.
Phylogenetic Relationship Analysis
Whole genomic DNA was extracted from the four mandarin cultivars using an Automatic Nuclear extraction kit (MX 16, Promega, Madison, WI, USA) and stored at -70℃. Phylogenetic analysis was conducted based on the nucleotide sequences of the trnL-trnF chloroplast region and the entire ITS region (ITS1-5.8S-ITS2) of nuclear ribosomal DNA, following (
Jin et al. 2016;
Jin et al. 2018). The trnL-trnF and ITS regions were amplified using the following primers: trnLF: 5'-AAAATCGTGAGGGTTCAAGTC-3' and trnFR 5'-GATTTGAACTGGTGACAC-GAG-3' (
Jung et al. 2005); ITS1F1: 5'-GAAGGATCATTGTCGACCTGCCAGCAGACG-3' and ITS2R2 5'-GACCTGGGGTCGCAATGCGAGCGCCGCTT-3' (
Jin et al. 2018). PCR products were verified using electrophoresis on 1.2% agarose gel at 100 V for 30 min, purified using the GeneAll® Gel Purification kit (GeneAll Biotechnology, Co., Seoul, Korea), cloned using the pLUG-Prime® TA-Cloning Vector kit (iNtRON, Korea), and transformed into Escherichia coli (
Jin et al. 2018). Sequencing was performed on three out of five white colonies from each transformation using SolGent (SolGent Co., Daejeon, Korea). The resulting sequences were then subjected to homology analysis using BLAST against the GenBank database, and one representative sequence was selected and edited using BioEdit (
Hall 1999). Finally, the nucleotide sequences from 18 citrus cultivars (
Table S1), including 12
Citrus unshiu mandarin cultivars registered in GenBank (
Kim et al. 2021), 2 hybrid mandarins reported in a previous study (
Jin et al. 2025), and 4 mandarin cultivars tested in this study were used for the analysis of phylogenetic relationships using the Molecular Evolutionary Genetics Analysis (MEGA) version 5.2 software (
Jin et al. 2018).
Data analysis was performed using analysis of variance (ANOVA), followed by Duncan’s multiple range test for mean comparisons. Differences were considered statistically significant at p < 0.05.
Results
Callus Proliferation Rates between Cultivars based on the Carbohydrate Source
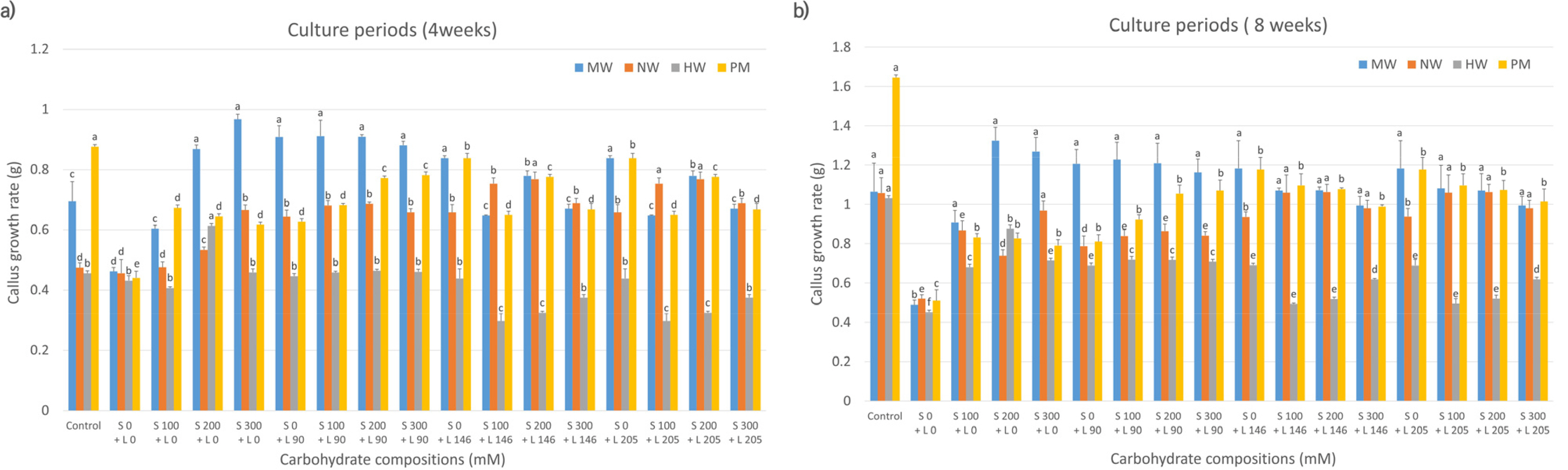
Evaluation of the callus proliferation rates of the four mandarin cultivars, based on carbohydrate sources, revealed that MW, NW, and HW had higher proliferation rates at 4 weeks of culture in media containing 200-300 mM sorbitol only than that in control media (
Fig. 1a). NW showed increased proliferation with a combination of 200 mM sorbitol and 146 mM lactose compared to the control. In contrast, PM cultured in media containing sorbitol or lactose alone or in combination exhibited lower proliferation rates. At 8 weeks (
Fig. 1b), MW showed elevated proliferation in media containing 200-300 mM sorbitol compared to that in control media; however, the difference was not statistically significant. The other three cultivars (NW, HW, and PM) had lower proliferation rates in sorbitol-supplemented media than those in control media.
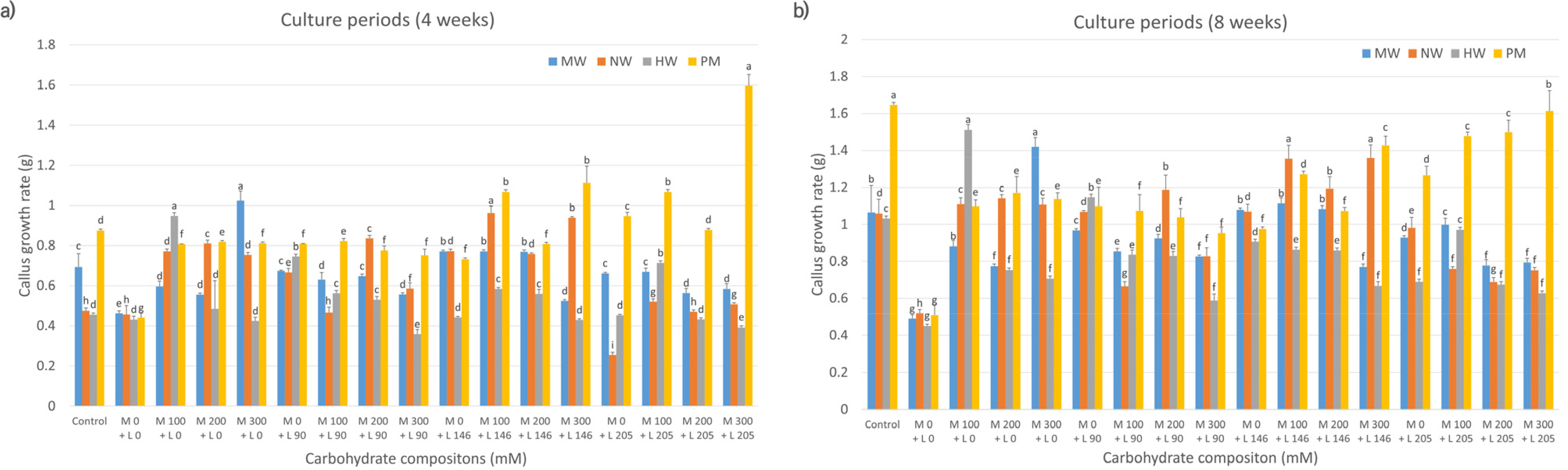
Fig. 2 shows a comparative analysis of callus proliferation rates in citrus cultivars treated with maltose, lactose, or their combination. At 4 weeks (
Fig. 2a), cell proliferation of MW was higher in the media supplemented with 300 mM maltose alone than those in control and other media compositions. HW showed higher proliferation in 100 mM maltose-alone media than in control and other media compositions. Conversely, compared to the control, NW and PM showed improved proliferation in media supplemented with a mixture of maltose and lactose in most combinations. The proliferation rate of NW grown in media containing 100 mM maltose and 146 mM lactose was twice as high (0.96) as that in control media (0.48). In contrast, PM exhibited the highest cell proliferation rate when grown in media containing 300 mM maltose and 205 mM lactose compared to other compositions (
Fig. 2a). At 8 weeks (
Fig. 2b), the proliferation rate of MW in 300 mM maltose was comparable to that observed at 4 weeks, showing superior proliferation on the same medium compared to the control and other media. Similarly, NW demonstrated the highest proliferation rate in media containing 100 mM maltose and 146 mM lactose and those with 300 mM maltose and 146 mM lactose, consistent with the results at 4 weeks, outperforming the rate in control and other media compositions. HW exhibited the highest cell proliferation rate in 100 mM maltose-only media, surpassing that in the control and other media, similar to that observed at 4 weeks. In contrast, the proliferation rate of PM was enhanced in media containing 300 mM maltose and 205 mM lactose at 4 weeks compared to that in the control and other media compositions. However, at 8 weeks, PM grown in control media exhibited a significantly higher proliferation rate.
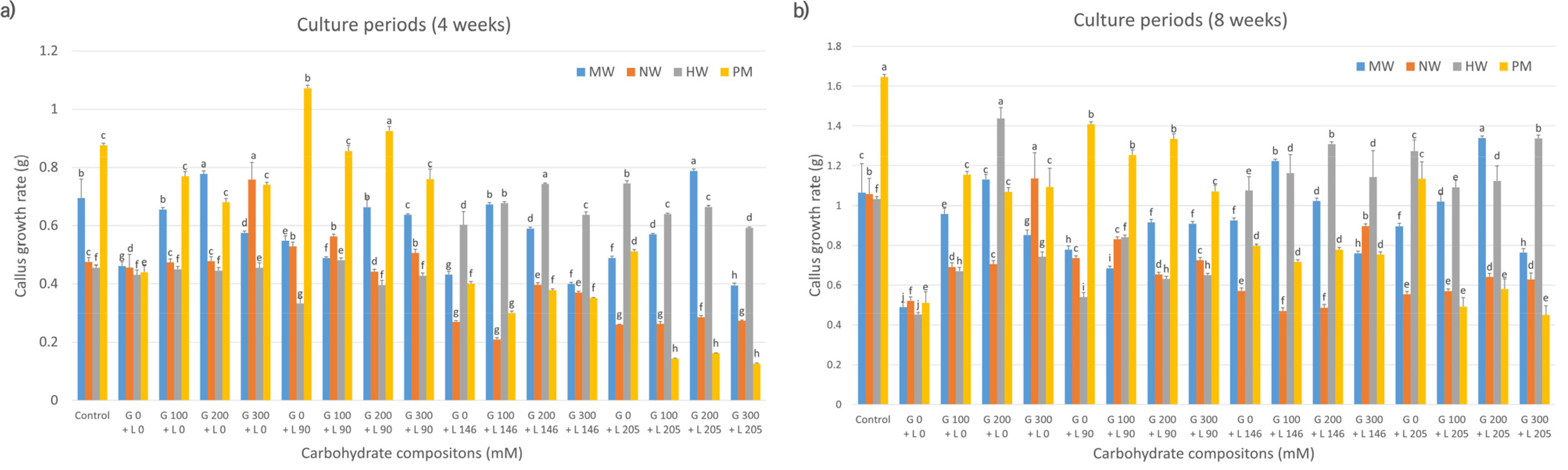
Next, we assessed callus proliferation rates among citrus varieties treated with galactose and lactose, individually and in combination. Four weeks post-culture, MW and HW had the highest proliferation rates in the combined treatments of 200 mM galactose and 205 mM lactose and 200 mM galactose and 146 mM lactose, respectively, compared with those in other compositions (
Fig. 3a). PM also showed maximum proliferation in media containing 200 mM galactose and 90 mM lactose. Conversely, NW performed better in 300 mM galactose-supplemented media. At 8 weeks post-culture, MW and NW exhibited higher cell proliferation rates in the same media compositions as at 4 weeks, surpassing those observed in the control and other treatments. In contrast, unlike at 4 weeks, HW demonstrated the highest proliferation rate in the medium supplemented solely with 200 mM galactose, outperforming the control and other media compositions. Additionally, unlike its response at 4 weeks, PM exhibited the highest proliferation rate in the control medium compared to those supplemented with either galactose or lactose alone or in combination (
Fig. 3b). These results were statistically significant compared to other compositions (
Fig. 3).
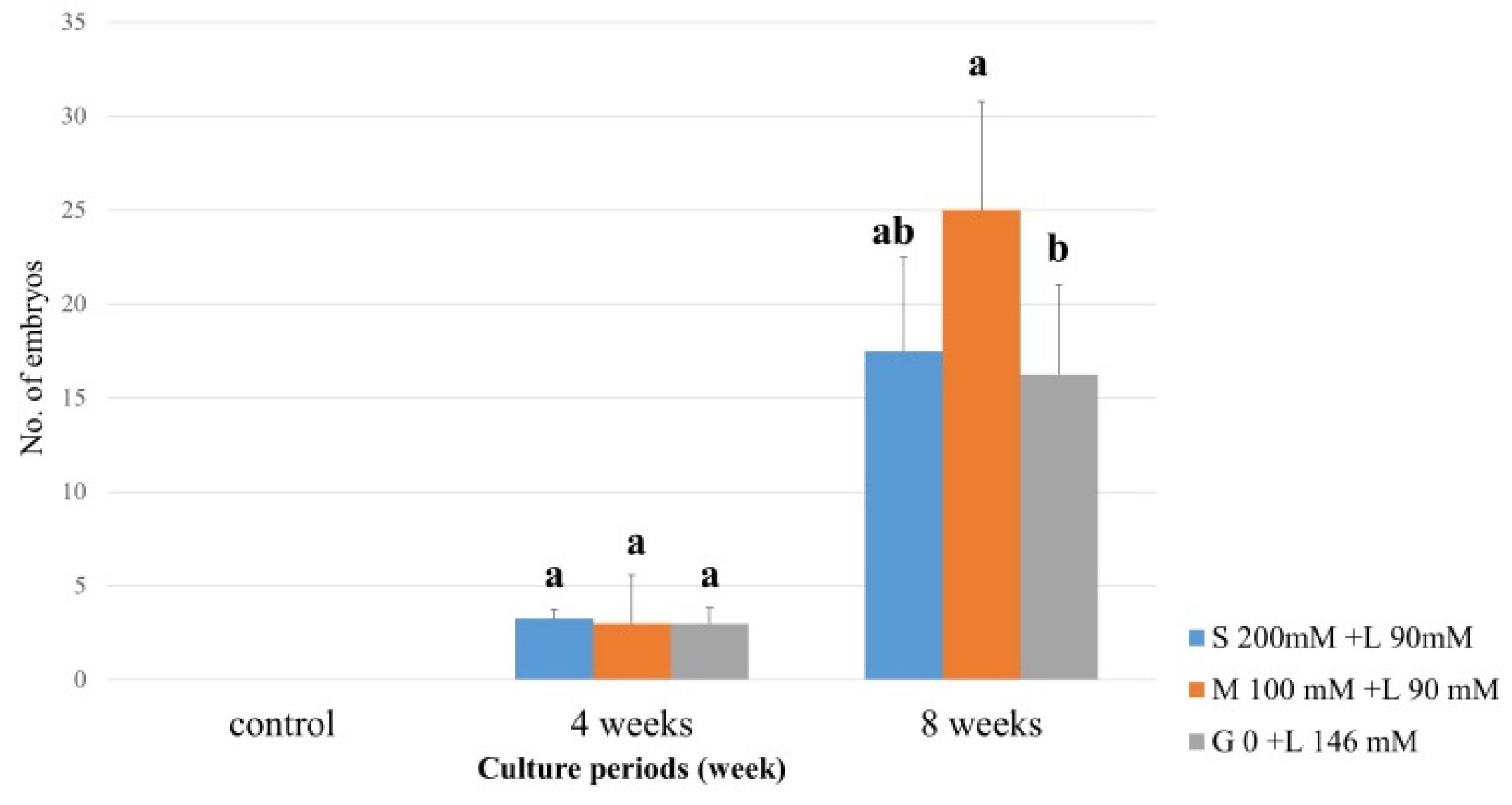
Evaluation of the rate of somatic embryo formation based on carbohydrate source composition revealed that MW formed somatic embryos when treated with a mixture of sorbitol (100 mM), lactose (90 mM), maltose (100 mM), lactose (90 mM), or lactose alone, starting at 4 weeks and becoming robust at 8 weeks (
Fig. 4). The rate of somatic embryo formation was highest in media containing 100 mM maltose and 90 mM lactose, compared with that in media containing lactose 146 mM alone (
Fig. 4).
Furthermore, MW exhibited considerable somatic embryo proliferation in media containing a mixture of galactose (100 mM) and lactose (146 mM;
Fig. 5a). In contrast, PM formed somatic embryos at a very low rate of ≤3 (
Fig. 5b) in media containing a mixture of galactose (200 mM) and lactose (146 mM). In contrast, the other two varieties did not form somatic embryos (
Figs. 5c,
5d). When sucrose was used as a control, it stimulated cell proliferation in all mandarin species; however, it did not induce somatic embryos.
Assessment of the regeneration abilities of the four cultivars revealed that, except for NW, the other varieties formed callus clusters and developed somatic embryos (
Table 1). The rates of somatic embryo formation were approximately 36 embryos per 50 mg of cells for MW, 60 embryos for PM, and 11 embryos for HW. Subsequent transfer of the embryos to MT medium for plant regeneration revealed transformation from the globular stage to the cotyledon stage after 5-6 weeks (
Fig. 6,
Table 1). The cotyledons were transferred to a hormone-free MS1 medium for 4 weeks, resulting in normal plant development (
Fig. 6). Plant regeneration rates were 35% (50/145), 25% (11/44), and 2% (5/240) for MW, HW, and PM, respectively (
Table 1).
To verify the maternal inheritance of the four mandarin species (‘Miyagawa wase’, ‘Nankan 20 gou’, ‘Haraejoseng’, and ‘Ootaponkan’), sequence divergences were calculated based on the nucleotide sequences of the trnL-trnF region of the chloroplast (
Supplementary Table 1). As shown in
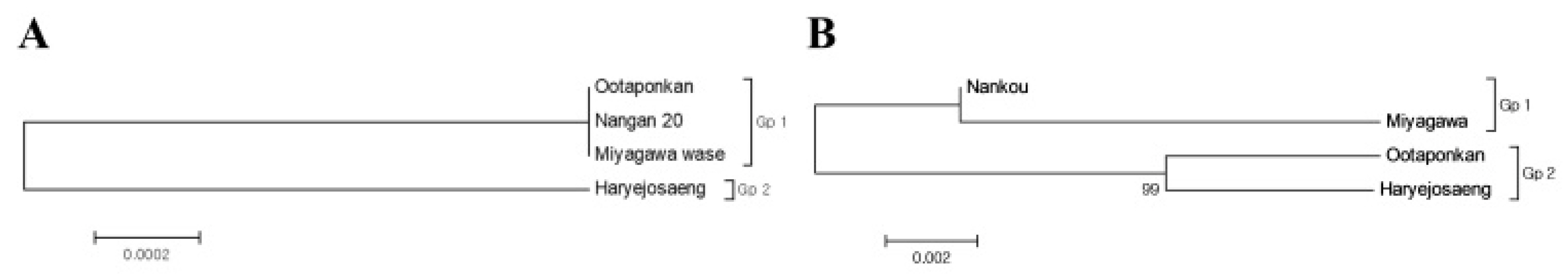
Table 2, the divergences of the three varieties, except for ‘Haraejoseng’ among the mandarins, were 0, indicating a close genetic relationship. The divergence value for ‘Haraejoseng’ was 0.002. Molecular phylogenetic relationships among citrus varieties were investigated using the Maximum Likelihood method based on nucleotide sequence data for the trnL-trnF intergenic spacer region. As a result, two classification groups, Gp1 and Gp2, were obtained, as shown in
Fig. 7a.
The changes in nuclear chromosomes of citrus species were determined based on the nucleotide sequences of the entire ITS (ITS1+5.8S rDNA+ITS2) region, and the branch lengths were calculated (
Table 3,
Table S1). As shown in
Table 3, the branch lengths ranged from 0.025 to 0.04, indicating genetic distance between the four species. Among the mandarin varieties, ‘Haryejosaeng’ from the
C. unshiu mandarin group showed genetic distance of 0.015 and 0.025 from ‘Nankan 20 gou’ and ‘Miyagawa wase’, respectively, while it was closer to ‘Ootaponkan’ with a distance of 0.009 compared to the other two
C. unshiu mandarin varieties. Molecular phylogenetic relationships among citrus varieties were also investigated using the Maximum Likelihood method, resulting in two classification groups, Gp1 and Gp2, as shown in
Fig. 7b. ‘Nankan 20 gou’ and ‘Miyagawa wase’ belonged to the same group (GP1), while ‘Ootaponkan’ and ‘Haryejosaeng’ belonged to the different group (Gp2).
Also, a phylogenetic analysis based on the trnL-trnF chloroplast region sequences (
Table S1) was conducted using MEGA version 5.2 to confirm maternal inheritance among 18 citrus varieties (
Table S2), including the four mandarin cultivars tested in this study (NW, HW, MW, PM). The sequence divergence within the trnL-trnF chloroplast region ranged from 0 to 0.012 (
Table S3). Among the four mandarin cultivars, MW, NW, and PM showed no sequence variation, suggesting a close genetic relationship. In contrast, HW was distinguished from the other three cultivars with a sequence divergence of 0.002. The genetic distance between these four mandarin cultivars and the remaining citrus varieties ranged from 0.010 to 0.012, with the highest divergence observed from the Tavares’ lime cultivar (
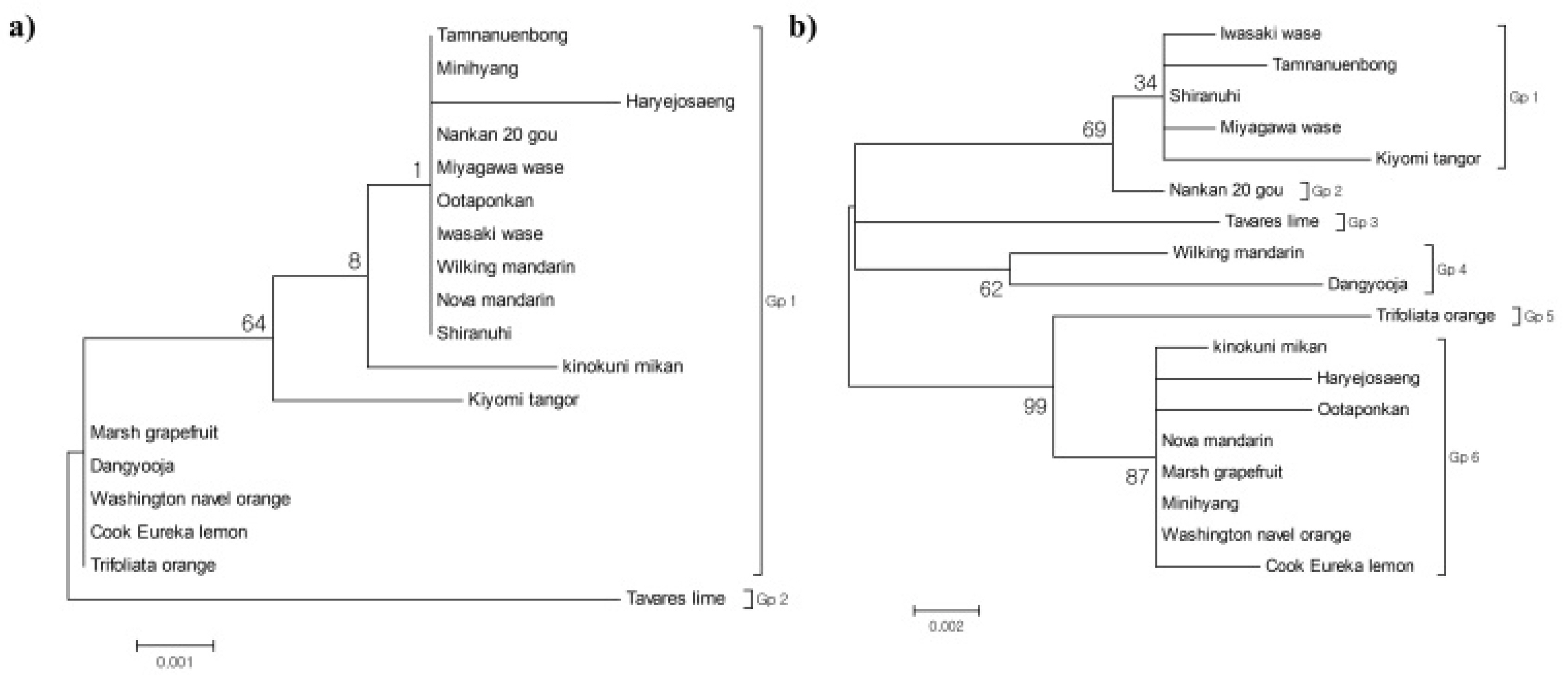
Table S3). Phylogenetic analysis using the Maximum Likelihood method grouped the varieties into two major clusters: Gp1, which included MW, NW, PM, HW, and 13 other citrus varieties, and Gp2, which contained Tavares’ lime (
Fig. 8a).
Nuclear chromosomal variation among citrus varieties was assessed based on the entire ITS region, including ITS1, 5.8S rDNA, and ITS2 sequences (
Table S1). Sequence divergence within the ITS region ranged from 0 to 0.031 (
Table S4). Genetic distances among the four mandarin cultivars varied from 0.003 to 0.026. NW exhibited the closest genetic relationship with ‘Shiranuhi’ (0.002) and the greatest divergence from ‘Trifoliate orange’ (0.024). HW displayed a genetic similarity of 0.005 with ‘Nova’ mandarin, ‘Marsh,’ ‘Minihyang,’ and ‘Washington navel’ orange, and the greatest divergence from ‘Dangyooja’ (0.029). Additionally, among the four mandarin varieties, PM showed the closest genetic relationship with a genetic distance of 0.010, while it exhibited a more distant genetic relationship with NW (0.023) and MW (0.026). MW showed the closest genetic distance to NW and ‘Shiranuhi’ (0.002) and the farthest from ‘Trifoliate orange’ (0.027). Among the four mandarin varieties, MW was found to have the closest genetic relationship with a genetic distance of 0.003. In contrast, HW and PM exhibited a more distant genetic relationship with a genetic distance of 0.026. NW exhibited the closest relationship with HW (0.006) but a more distant genetic relationship with MW and PM (0.026 and 0.010, respectively). PM was most closely related to HW, ‘Nova’ mandarin, ‘Marsh,’ ‘Minihyang,’ and ‘Washington navel’ orange (0.005) and showed the greatest divergence from ‘Dangyooja’ (0.029). Among the four mandarin cultivars, PM exhibited the closest genetic distance to HW (0.010), while NW and MW showed a genetic distance of 0.023 or 0.026 from PM, indicating a more distant relationship (
Table S4). Phylogenetic analysis classified the citrus varieties into six groups: Gp1 (including MW and four other citrus varieties), Gp2 (NW), Gp3 (Tavares’ lime), Gp4 (Wilking mandarin and Dangyooja), Gp5 (Trifoliate orange), and Gp6 (HW, PM, and six other citrus varieties;
Fig. 8b).
Discussion
Effect of Carbohydrate Source Composition on Cell Proliferation Rate by Citrus Cultivars
In this study, the effects of carbohydrate source composition on cell proliferation rates of four mandarin varieties were investigated. Among the three
C.
unshiu varieties, MW showed the highest proliferation rate in media containing 300 mM mannitol and proliferated well in media supplemented with 300 mM sorbitol and a mixture of galactose and lactose. NW achieved the best rate with 100 mM mannitol and 146 mM lactose, whereas HW proliferated well with 100 mM mannitol alone. These findings confirm that carbohydrate sources significantly affect citrus cell proliferation, similar to previous findings (
Gholami et al. 2013;
Haoa et al. 2004;
Kayim et al. 2006). Moreover, lactose and galactose are more effective than maltose, sorbitol, and glucose in the proliferation of citrus cultivars (
Hidaka et al. 1989;
Kayim et al. 2006). However, our findings showed that mannitol and sorbitol played a key role as carbohydrate sources in the cell proliferation of
C.unshiu mandarin cultivars.
In
C.
reticulata cultivar PM, carbohydrate sources did not affect its proliferation, as evident from comparable proliferation rates in 5% sucrose and other carbohydrate sources. This finding aligned with those of previous studies (
Giladi et al. 1977;
Jin 2005). Together, these findings demonstrate that carbohydrate sources other than sucrose significantly affect cell proliferation in satsuma cultivars but not that of other mandarin cultivars, which could be attributed to the genetic differences among the citrus cultivars (
Carimi et al. 1998).
The results of investigating the somatic embryogenesis induction rate of four mandarin cultivars by carbohydrate source composition showed that only the cells of MW and PM were capable of inducing somatic embryogenesis. According to
Ricci et al. (2002), lactose (75 mM) and high concentration maltose (150 mM) as single treatments were effective for somatic embryogenesis induction from cells of the ponkan mandarin (
Citrus reticulata, Blanco) cultivar, but in this study, the induction of somatic embryogenesis from PM cells was not achieved with these carbohydrate sources as single treatments. On the other hand, although somatic embryogenesis induction was not observed with other carbohydrate source compositions, it was confirmed that somatic embryogenesis could be induced to a small extent by mixing galactose (100 mM) and high concentration lactose (146 mM) as the carbohydrate source. However, in this mixed carbohydrate source composition, it was observed that somatic embryogenesis did not proliferate overall but tended to degenerate or deteriorate, possibly because galactose and lactose acted as toxins in growth, rather than as sugars necessary for plant growth (
Carimi et al. 1998;
Gaj 2004;
Gholami et al. 2013;
Hao YuJin et al. 2004;
Kayim et al. 2006;
Kochba et al. 1982;
Perez et al. 1998;
Sakai et al. 1990). It is therefore suggested that as the culture period lengthens, somatic embryogenesis may deteriorate.
According to
Kayim et al. (2006), there are differences in somatic embryos induction between citrus cultivars. In this study, it was observed that only the MW variety of mandarin showed an increase in somatic embryos culture from 4 weeks after culture to vigorous growth at 8 weeks when treated with a mixture of sorbitol, maltose, and lactose or lactose alone. In contrast, the other mandarin varieties did not show any increase in somatic embryos culture even when treated with high concentrations of carbohydrate sources. In particular, MW showed the highest rate of somatic embryos culture formation when maltose (100 mM) and lactose (90 mM) were mixed, rather than when only lactose (146 mM) was used. Previous studies (
Hidaka et al. 1989;
Perez et al. 1998;
Tomaz et al. 2001) have shown that galactose, lactose, and maltose are effective in inducing somatic embryos culture in citrus, with lactose being the most effective, especially for mandarin varieties (
Jin et al. 2007;
Kunitake et al. 1991;
Ling et al. 1990). This study also found that high concentrations of lactose could induce somatic embryos culture in mandarin, similar to the previous studies. In addition, the best results were obtained when maltose and lactose were mixed as carbohydrate sources, rather than when lactose alone was used, which is believed to be due to the stress factors required for somatic embryos culture induction in MW mandarin (
Lincy et al. 2009). To the best of our knowledge, there are no reports on the investigation of somatic embryos culture efficiency in citrus according to the composition of mixed carbohydrate sources.
Although, high concentrations of agar and lactose treatment as a water stress agent resulted in high efficiency of somatic embryo formation, it was observed that the cells undergo necrosis with prolonged culture duration (
Jin 2005;
Jin et al. 2007). Considering this, it was expected that the application of the method of mixing carbohydrate source ratios could not only reduce the percentage of cell necrosis but also increase the rate of somatic embryo formation.
Furthermore, based on the above results, callus proliferation in citrus shows high efficiency when sucrose is supplied within an appropriate range that does not impose stress on cell growth. In contrast, some citrus species have been reported to form somatic embryos when exposed to high concentrations of sucrose due to osmotic stress (
Ricci et al. 2002). However, cultivars with low regenerative capacity, such as Satsuma mandarin, reportedly induce somatic embryogenesis not through sucrose but rather through stress stimuli such as osmotic pressure, or by using lactose—which cannot be utilized as an energy source—as well as by increasing the concentration of solidifying agents in the medium (
Jin et al. 2007;
Jin et al. 2017;
Jin et al. 2025). These findings suggest that citrus genotypes vary considerably in their regenerative competence; while some genotypes regenerate readily, others require embryo-inducing media to successfully regenerate plants.
Previous studies have shown that long-term subculturing can impair the regeneration ability of plant-cultured cells due to somatic variation (
Kim et al. 1995;
Li et al. 2004). This study demonstrated that all four mandarin cultivars lost their regeneration ability. Although MW and PM formed some calli, the rate was too low. These findings align with those of
Jin et al. (2007), who showed that carbohydrate type and agar concentration are crucial for somatic embryogenesis under stress. Cells from the four mandarin cultivars were separated using a Percoll density gradient and cultured twice on media to regenerate cells with regeneration capability, inducing somatic embryogenesis. Consequently, from three mandarin lines, except NW, regained regeneration ability. NW was deemed completely lost its regeneration ability, as its cells neither formed callus nor proliferated. Thus, alternative propagation methods and optimized tissue culture conditions are recommended to enhance the regeneration of NW.
Evaluation of the plant regeneration capacity from somatic embryos derived from mandarin cultivars capable of callus formation revealed that MW displayed the highest regeneration rate (35%). Conversely, even though PM exhibited a cell division rate 2-6 times higher than that of other cultivars, its plant regeneration ability was very low, indicating that the cells that do not develop into normal plantlets may have undergone division. Furthermore, attempts to improve its regeneration ability using established methods were unsuccessful (
Dutt et al. 2010;
Kazmi et al. 2015;
Li et al. 2002). These findings imply that somatic variation has led to a loss of redifferentiation ability, as suggested by
Li et al. (2004). Therefore, securing a new cell line through alternative culture methods and further investigating the specific factors limiting its regeneration is warranted.
Many molecular marker techniques have been employed to elucidate the genetic relationships and origins among plant cultivars (
Tingey et al. 1993). Among these, widely used markers for citrus phylogenetic studies include the chloroplast intergenic region trnL-trnF (
Huh et al. 2011;
Jung et al. 2005;
Penjor et al. 2013) and the nuclear ribosomal DNA internal transcribed spacer (ITS) region (
Jin et al. 2016;
Sun et al. 2015). In contrast, phylogenetic analyses based on mitochondrial regions may be prone to errors in determining maternal lineage due to biparental inheritance of mitochondria (
Froelicher et al. 2011;
Jin et al. 2016). Therefore, in this study, we applied the trnL-trnF region (reflecting maternal inheritance) and the ITS region (reflecting biparental inheritance), which are commonly used to investigate interspecific genetic relationships and evolutionary origins, to the phylogenetic classification of citrus (
Baldwin et al. 1995;
Kim et al. 2021;
Moon et al. 2015).
The phylogenetic analysis of citrus revealed that HW and MW tangerines are genetically distinct. This finding aligns with those of
Kim et al. (2021). Additionally, previous studies have demonstrated that citrus cell formation is genotype-dependent (
Cardoso et al. 2014;
Carimi 2005;
Starrantino et al. 1980) and that genetic differences influence adventitious root formation (
Ha et al. 2002;
Hardwick 1979). Moreover, the
C. unshiu mandarin cultivars, which are known to have poor regeneration capacity (
Han et al. 2002;
Jin et al. 2007), belong to the mandarin lineage phylogenetically, but according to
Tanaka (1977) classification, these lineages are grouped under the
C. unshiu mandarin category. In other words, ‘Ponkan’ belongs to the mandarin lineage that shows good regeneration ability (
Perez et al. 1998), whereas the
C. unshiu mandarins, unlike typical mandarin lineages, exhibit poor regeneration. Consequently, differences in the efficiency of somatic embryogenesis due to variation in carbohydrate sources can be attributed to genetic differences between the cultivars even though they belong to the same genus. These findings suggest optimizing carbohydrate composition ratio could enhance regeneration efficiency in specific citrus varieties.
Conclusion
This study revealed that carbohydrate sources significantly influenced cell proliferation, somatic embryogenesis, and plant regeneration in mandarin cultivars, with varied responses. High mannitol and lactose concentrations enhanced cell proliferation, while sorbitol and maltose combinations promoted somatic embryo growth in specific cultivars. Phylogenetic analysis showed distinct genetic differences among the cultivars, affecting their regeneration capacities and highlighting the need for cultivar-specific carbohydrate optimization. Additionally, it emphasizes the importance of a selection method for cells to restore regeneration ability from those that have lost their regenerative capacity. However, this study evaluated a limited number of mandarin cultivars and carbohydrate sources, which may not fully capture the diversity of responses across all citrus species. Therefore, further studies with expanded sample sizes are necessary to generalize these findings. Furthermore, a comprehensive assessment of carbohydrate types and concentrations could provide deeper insights into their influence on citrus cell proliferation and somatic embryogenesis. The phylogenetic analysis was based on sequencing data of only three out of five white colonies per transformation event, which may have affected the accuracy of the inferred phylogenetic relationships. Lastly, the study did not investigate the molecular mechanisms underlying the observed differences in cell proliferation and regeneration. Future research should incorporate transcriptomic or metabolomic profiling to gain deeper insights into the pathways regulating these responses. Addressing these limitations will enhance the robustness and applicability of the findings for citrus tissue culture and genetic analysis.
Acknowledgments
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01735603),” Rural Development Administration, Republic of Korea.
Fig. 1Effects of carbohydrate sources (sorbitol and lactose combination) on callus growth rate in citrus cultivars. MW: ‘Miyagawa wase’; NW: ‘Nankan 20 gou’; HW: ‘Haryejosaeng’; PM: ‘Ootaponkan,’ S; sorbitol, L; lactose. The proliferation rates of callus were determined by measuring the sample weight at 4 (a) and 8 (b) weeks of culture and subtracting the weight of the culture medium. The control medium contained sucrose at 50 g/L and malt extract at 500 mg/L, while the medium without sorbitol and lactose contained only malt extract at 500 mg/L. Means shown with the same lowercase letter indicate no significant difference at the p < 0.05 by Duncan’s multiple range test; values are the mean ± SE (n = 5).
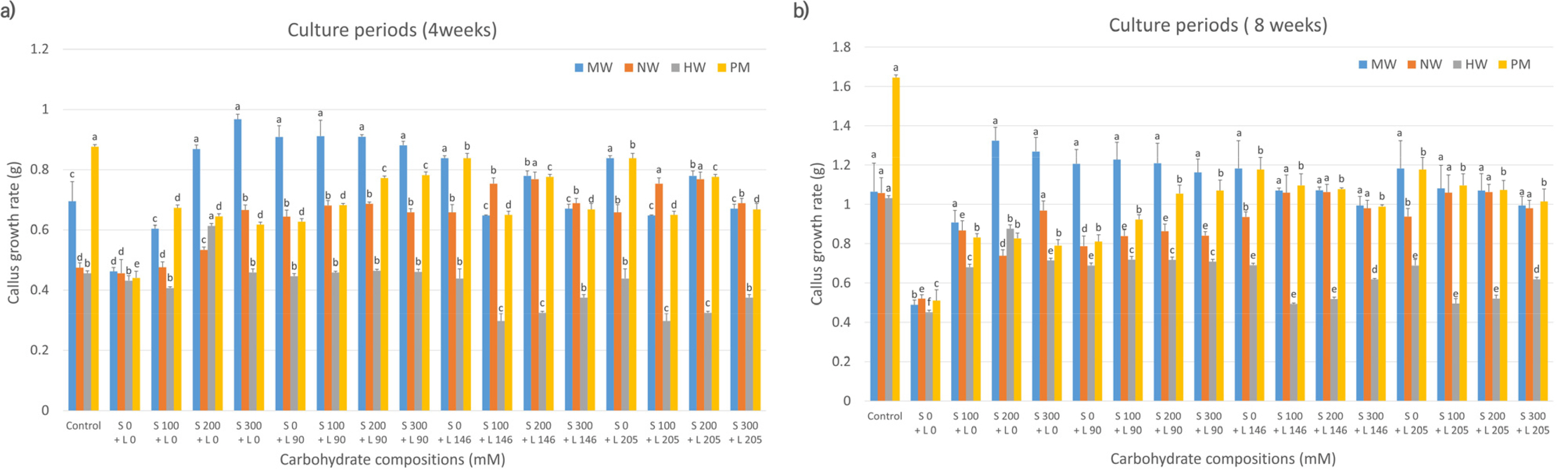
Fig. 2Effects of carbohydrate sources (maltose and lactose combination) on callus growth rate in citrus cultivars. MW: ‘Miyagawa wase’; NW: ‘Nankan 20 gou’; HW: ‘Haryejosaeng’; PM: ‘Ootaponkan,’ M; maltose, L; lactose. The proliferation rates of callus were determined by measuring the sample weight at 4 (a) and 8 (b) weeks of culture and subtracting the weight of the culture medium. The control medium contained sucrose at 50 g/L and malt extract at 500 mg/L, while the medium without maltose and lactose contained only malt extract at 500 mg/L. Means shown with the same lowercase letter indicate no significant difference at the p < 0.05 by Duncan’s multiple range test; values are the mean ± SE (n = 5).
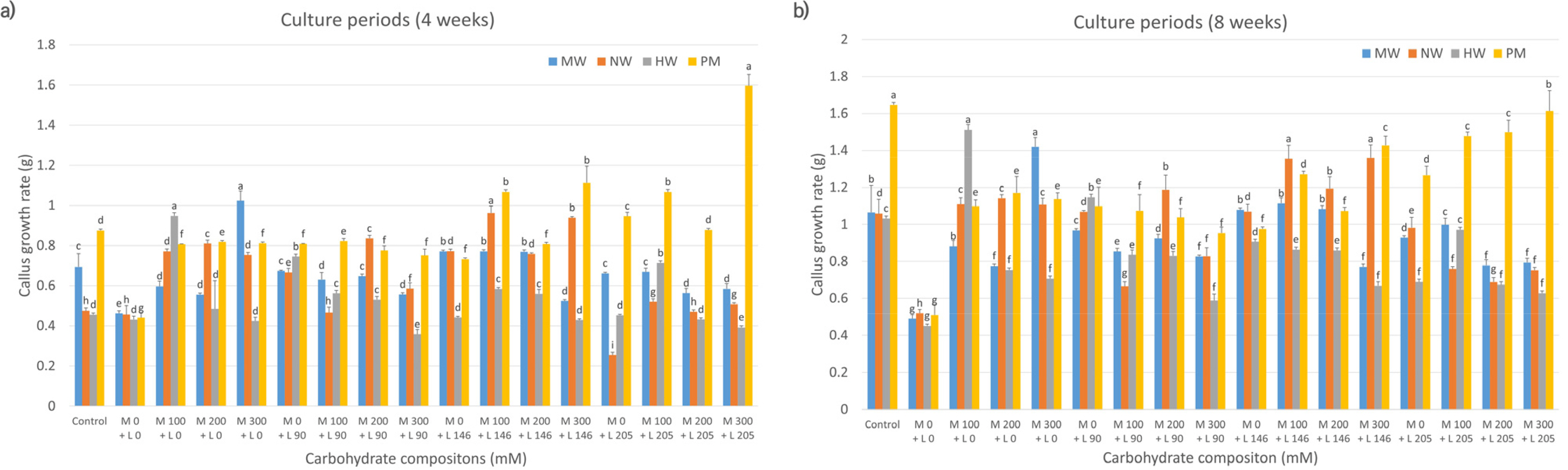
Fig. 3Effects of carbohydrate sources (galactose and lactose combination) on callus growth rate in citrus cultivars. MW: ‘Miyagawa wase’; NW: ‘Nankan 20 gou’; HW: ‘Haryejosaeng’; PM: ‘Ootaponkan,’ M; maltose, L; lactose. The proliferation rates of callus were determined by measuring the sample weight at 4 (a) and 8 (b) weeks of culture and subtracting the weight of the culture medium. The control medium contained sucrose at 50 g/L and malt extract at 500 mg/L, while the medium without galactose and lactose contained only malt extract at 500 mg/L. Means shown with the same lowercase letter indicate no significant difference at the p < 0.05 by Duncan’s multiple range test; values are the mean ± SE (n = 5).
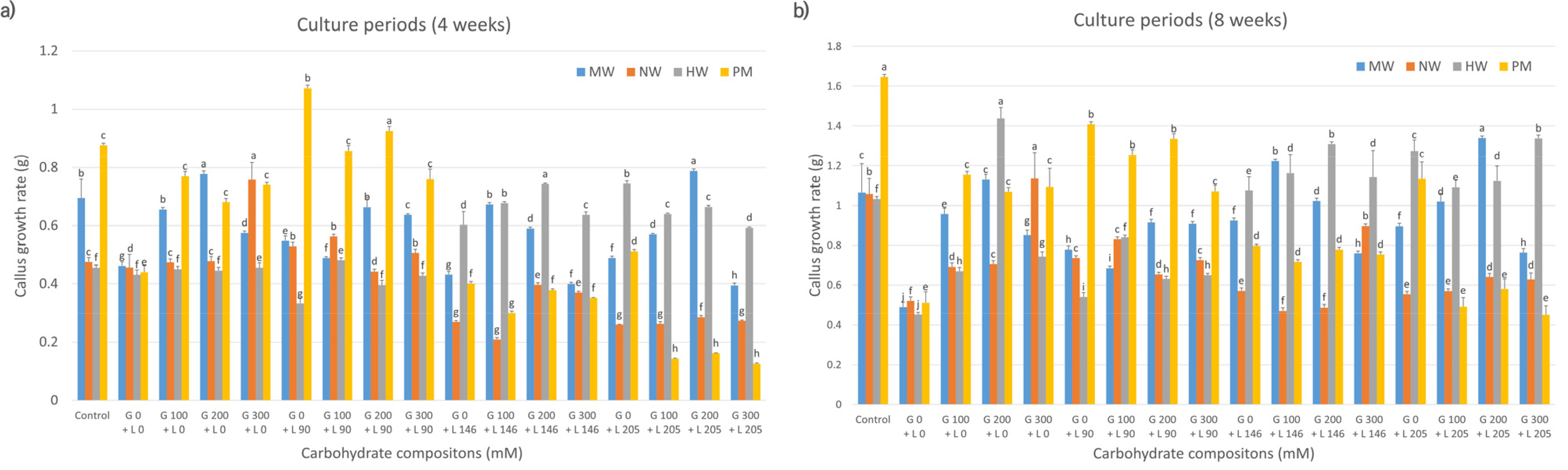
Fig. 4Carbohydrate impact on somatic embryo induction in C. unshiu ‘Miyagawa wase’ (MW). The induction rates of somatic embryo were determined by measuring at 4 and 8 weeks of culture. The control medium contained sucrose at 50 g/L and malt extract at 500 mg/L. Means shown with the same lowercase letter indicate no significant difference at the p < 0.05 by Duncan’s multiple range test; values are the mean ± SE (n = 4).
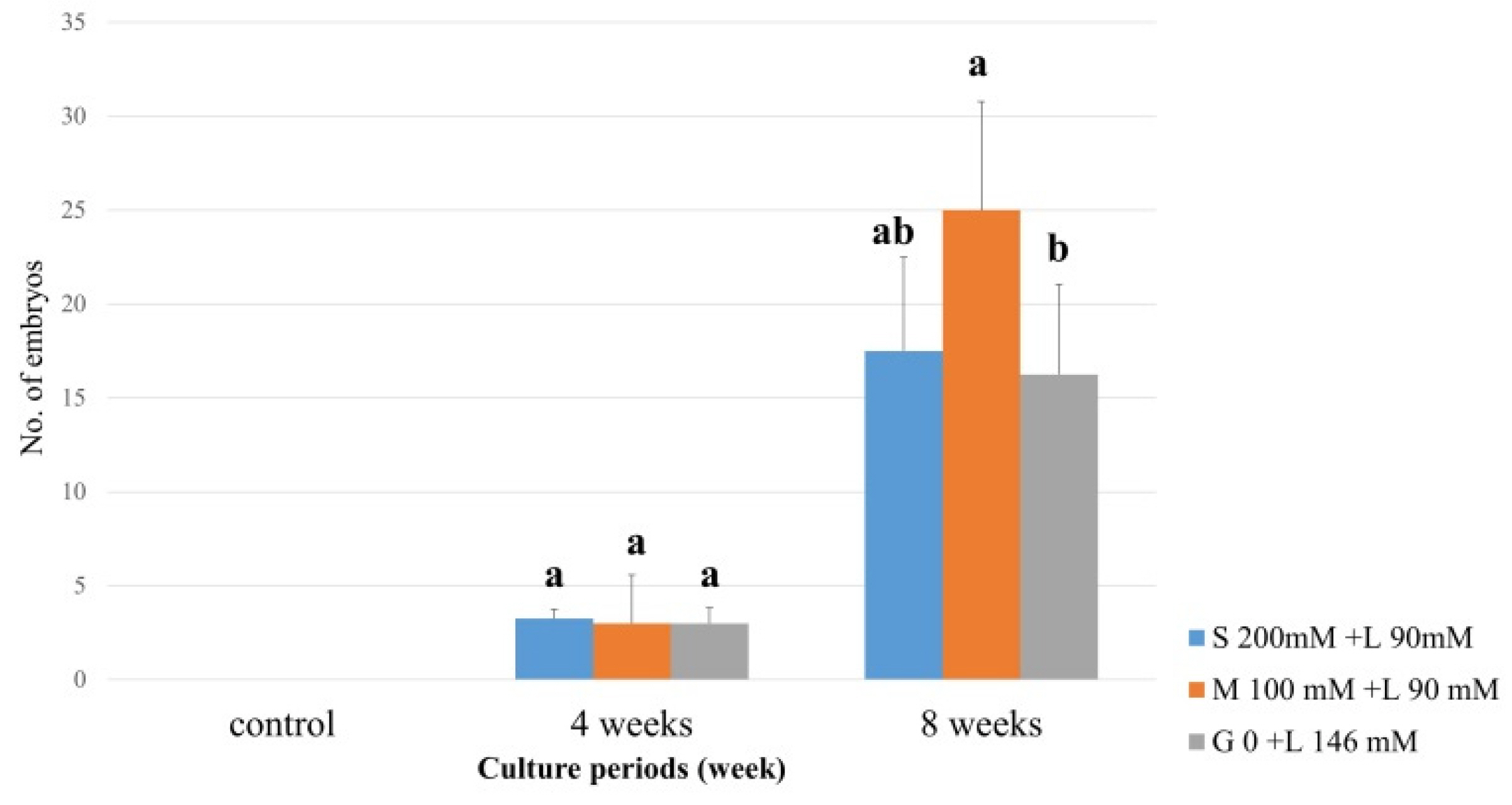
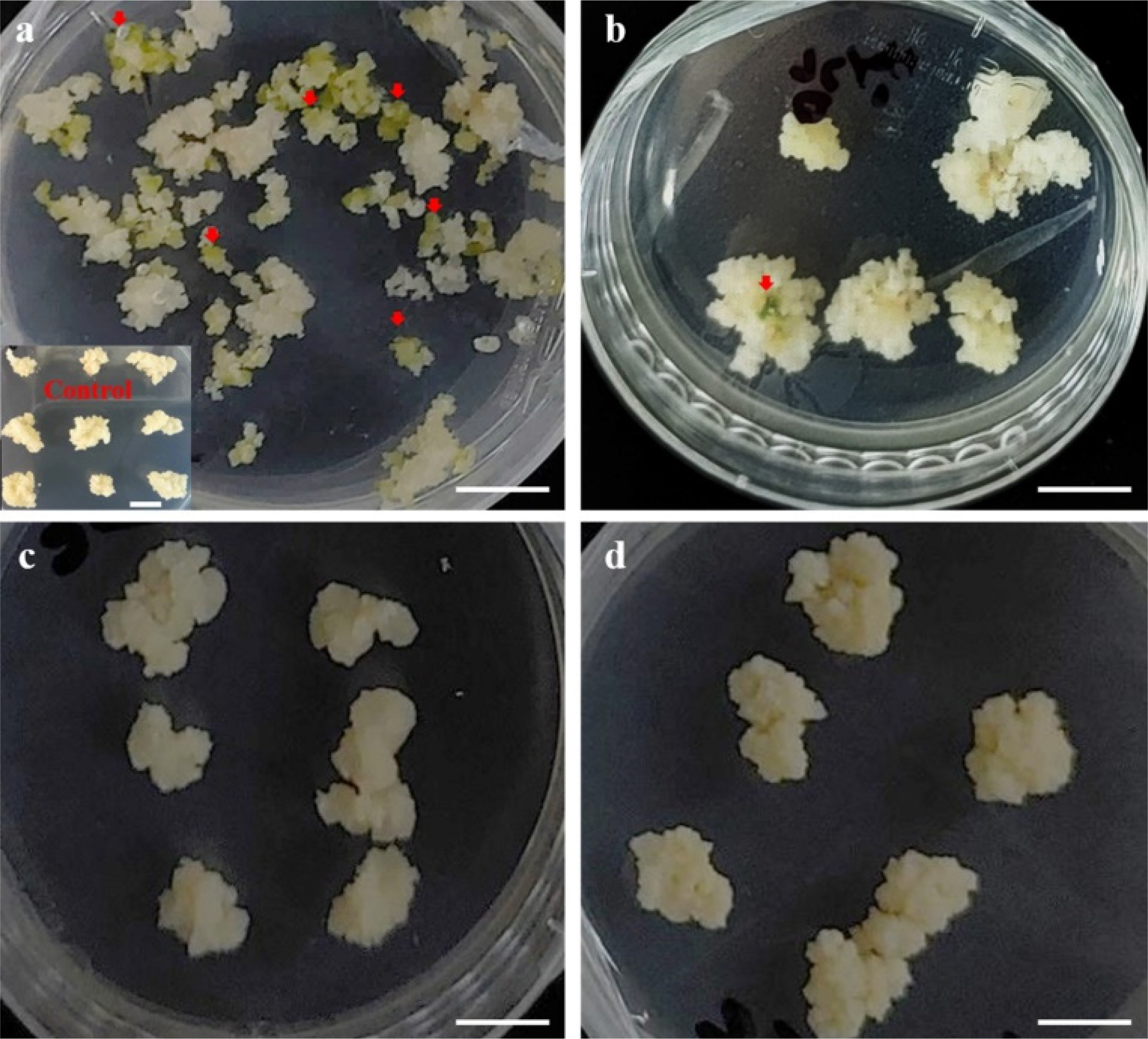
Fig. 5Carbohydrate effects on somatic embryo induction in citrus cultivars. Cells were cultured on MS1 medium supplemented with 100 mM maltose and 90 mM lactose for 8 weeks. (a) ‘Miyagawa wase’ (MW), (b) ‘Ootaponkan’ (PM); MS1 medium supplemented with 200 mM galactose and 146 mM lactose for 8 weeks., (c) ‘Haryejosaeng’ (HW), and (d) ‘Nankan 20 gou’ (NW). Experiments were conducted in four replicates. Scale bar is 1 cm.
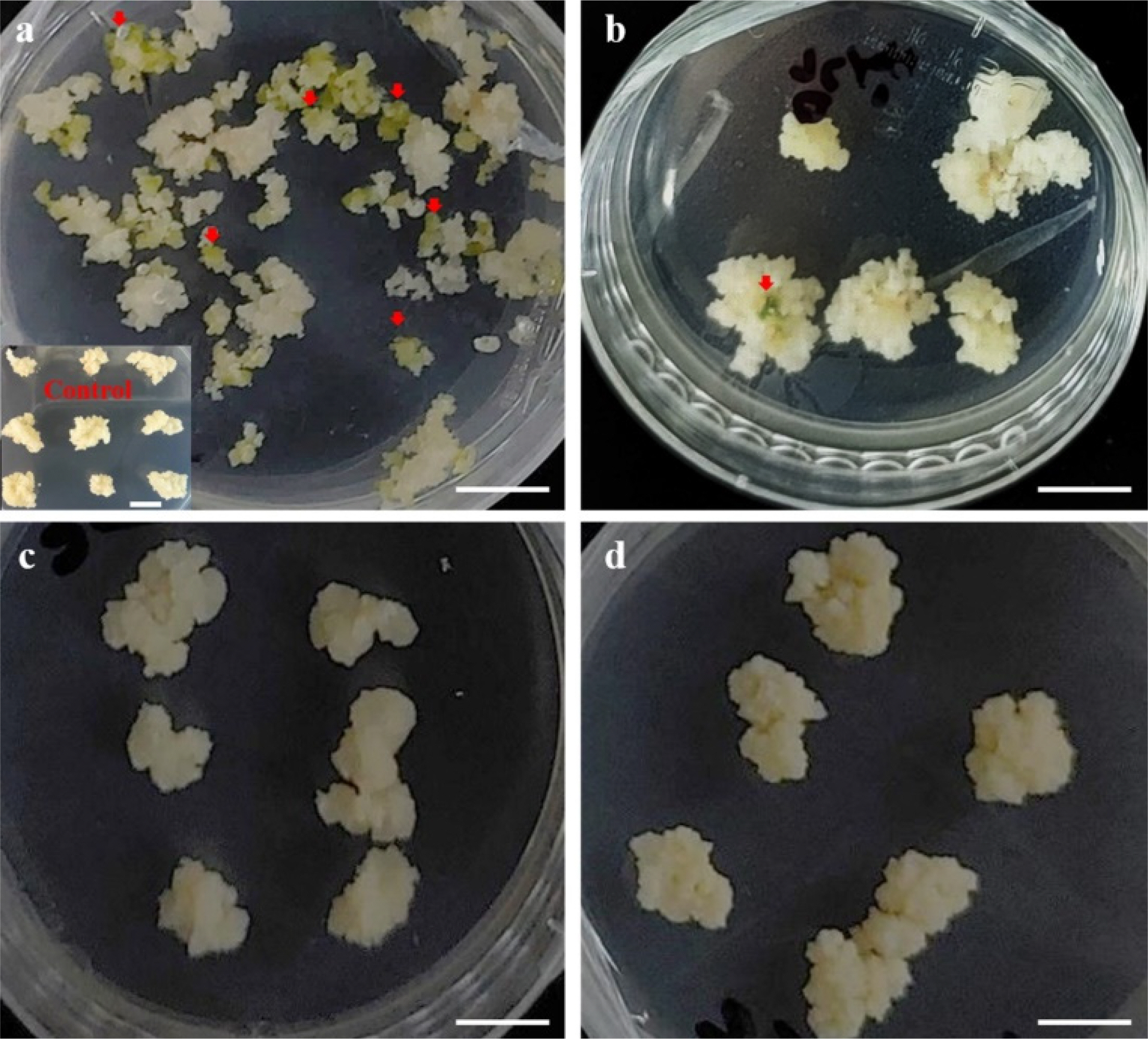
Fig. 6Somatic embryogenesis and regeneration in citrus cultivars. Step 1: Primary selection stage: cells on somatic embryogenesis induction medium from cells isolated from Percoll 40%; Cells were cultured on MT basal media with 500 mg/L malt extract, 146 mM lactose, and 1.6% agar for 8 weeks; Step 2: Secondary culture stage: 50 mg of cells selected from the primary selection stage cells that were cultured and propagated; Selected somatic embryos were cultured on MS1 media supplemented with 0.05 M sorbitol, 0.05 M galactose, 0.5 mg/L GA3, and 0.2% gelrite for 5-6 weeks; Steps 3: Somatic embryogenesis induction and plant regeneration stages; Normed roots and shoots were transferred to hormone-free MS1 media and subcultured every 4 weeks for 16 weeks. (a) ‘Miyagawa wase’, (b) ‘Haryejosaeng’, and (c) ‘Ootaponkan’ Scale bar is 500 ㎛. All experiments were repeated four times.

Fig. 7Maximum-likelihood phylogenetic tree of the four Citrus mandarin genotypes based on (a) chloroplast trnL-trnF intergenic spacer (IGS) and (b) total internal transcribed spacer (ITS; ITS1+5.8S rDNA+ ITS2) region sequences.
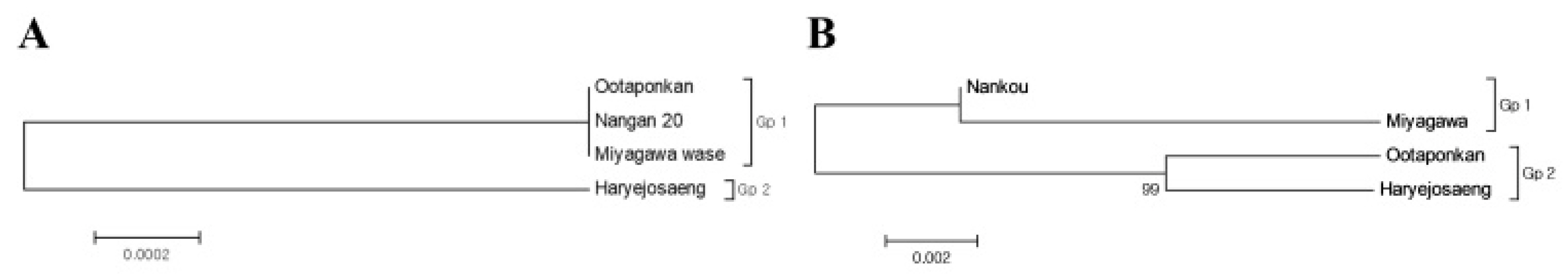
Fig. 8Phylogenetic analysis of citrus cultivars (n = 18), including the four citrus mandarin genotypes tested in this study. Maximum-likelihood trees based on (a) chloroplast trnL-trnF IGS and (b) ITS region sequences. Bootstrap values (%) indicated at nodes. The scale bar represents substitutions per site.
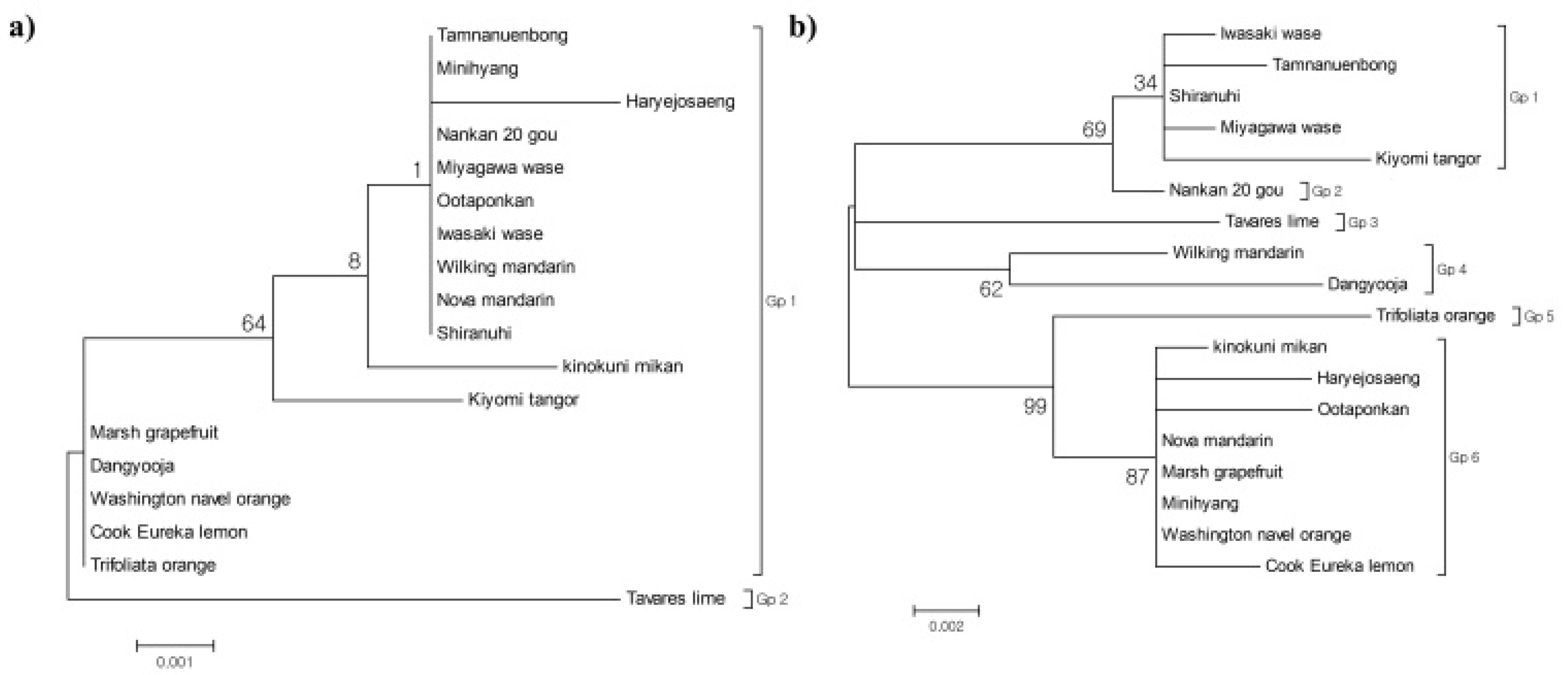
Table 1Somatic embryogenesis and plant regeneration abilities of different citrus cultivars.
Table 1
|
No. |
Cultivar |
Number of
somatic
embryo formationz)
|
Number of
regenerated plantY)
|
Plant regeneration rates
(Number of regenerated plants /total number of somatic embryos formed) |
|
1 |
‘Miyagawa wase’ |
36 ± 2.2b |
10 ± 1.4a |
35% (50/145) |
|
2 |
‘Nankan 20 gou’ |
0 ± 0d |
0 ± 0d |
0% (0/0) |
|
3 |
‘Haryejosaeng’ |
11 ± 1.8c |
2 ± 1.0b |
25% (11/44) |
|
4 |
‘Ootaponkan’ |
60 ± 7.0a |
1 ± 0.8c |
2% (5/240) |
Table 2Sequence divergences in the chloroplast trnL-trnFintergenic spacers (IGS)gene regions of between Citrus mandarin cultivars.
Table 2
|
No. |
Cultivar |
1 |
2 |
3 |
4 |
|
1 |
‘Nankan 20 gou’ |
|
0.000 |
0.000 |
0.002 |
|
2 |
‘Ootaponkan’ |
0.000 |
|
0.000 |
0.002 |
|
3 |
‘Miyagawa wase’ |
0.000 |
0.000 |
|
0.002 |
|
4 |
‘Haryejosaeng’ |
0.002 |
0.002 |
0.002 |
|
Table 3Sequence divergences in the total ITS (ITS1+5.8S rDNA+ ITS2) regions of between Citrus mandarin cultivars.
Table 3
|
No. |
Cultivar |
1 |
2 |
3 |
4 |
|
1 |
‘Nankan 20 gou’ |
|
0.005 |
0.004 |
0.005 |
|
2 |
‘Ootaponkan’ |
0.015 |
|
0.006 |
0.004 |
|
3 |
‘Miyagawa wase’ |
0.009 |
0.025 |
|
0.006 |
|
4 |
‘Haryejosaeng’ |
0.015 |
0.009 |
0.025 |
|
References
- An HJ, Jin S-B, Kang BC, Park HG. 2008. Production of somatic hybrids between satsuma mandarin (Citrus unshiu) and navel orange (Citrus sinensis) by protoplast fusion. J. Plant Biol. 51(3): 186-191.
- Baldwin BG, Sanderson MJ, Porter JM, Wojciechowski MF, Campbell CS, Donoghue MJ. 1995. The ITS region of nuclear ribosomal DNA: A valuable source of evidence on angiosperm phylogeny. Ann. Missouri Bot. Gard. 82(2): 247-277.
- Cardoso JC, Martinelli AP, Germana MA, Latado RR. 2014. In vitro anther culture of sweet orange (Citrus sinensis L. Osbeck) genotypes and of a C. clementina×C. sinensis 'Hamlin' hybrid. Plant Cell Tissue Organ Cult. 117(3): 455-464.
- Carimi F. 2005. Somatic embryogenesis protocol: Citrus. For. Sci. 77: 321-343.
- Carimi F, Tortorici MC, De Pasquale F, Crescimanno FG. 1998. Somatic embryogenesis and plant regeneration from undeveloped ovules and stigma/style explants of sweet orange navel group [Citrus sinensis (L.) Osb.]. Plant Cell Tissue Organ Cult. 54(3): 183-189.
- Dutt M, Vasconcellos M, Song K, Gmitter F Jr, Grosser J. 2010. In vitro production of autotetraploid Ponkan mandarin (Citrus reticulata Blanco) using cell suspension cultures. Euphytica. 173(2): 235-242.
- Froelicher Y, Mouhaya W, Bassene J-B, Costantino G, Kamiri M, Luro F, et al. 2011. New universal mitochondrial PCR markers reveal new information on maternal citrus phylogeny. Tree Genet. Genomes. 7(1): 49-61.
- Gaj MD. 2004. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 43(1): 27-47.
- Gholami AA, Alavi SV, Majd A, Fallahian F. 2013. Plant regeneration through direct and indirect somatic embryogenesis from immature seeds of citrus. Eur. J. Exp. Biol. 3(3): 307-310.
- Giladi I, Altman A, Goren R. 1977. Differential effects of sucrose, abscisic acid, and benzyladenine on shoot growth and callus formation in the abscission zone of excised citrus buds. Plant Physiol. 59(6): 1161-1164.
- Ha K-S, Han T-J. 2002. Adventitious root formation from cotyledon in soybean (Glycine max L.) cultivars. J. Plant Biotechnol. 29(1): 31-36.
- Hall TA. 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser.
- Han S-H, Kang S-K, An H-J, Kim H-Y. 2002. Effect of embryogenic callus conditions on plant regeneration in satsuma mandarin (Citrus unshiu Marc.). J. Plant Biotechnol. 4(1): 29-32.
- Hao Y-J, Wen X-P, Deng X-X. 2004. Genetic and epigenetic evaluations of citrus calluses recovered from slow-growth culture. J. Plant Physiol. 161(4): 479-484.
- Hardwick R. 1979. Leaf abscission in varieties of phaseolus vulgaris (L.) and glycine max (L.) merrill/emdash a-correlation with propensity to produce adventitious roots. J. Exp. Bot. 30(4): 795-804.
- Hidaka T, Omura M. 1989. Control of embryogenesis in citrus cell culture: Regeneration from protoplasts and attempts to callus bank. Bull. Fruit Tree Res. Stn. B. 16: 1-17.
- Huh M-K, Yoon H-J, Choi J-S. 2011. Phylogenic study of genus citrus and two relative genera in Korea by trnL-trnF sequence. J. Life Sci. 21(10): 1452-1459.
- Jin SB, Kim HB, Park S, Kim MJ, Choi CW, Yun S-H. 2020. Identification of the 'Haryejosaeng' mandarin cultivar by multiplex PCR-based SNP genotyping. Mol. Biol. Rep. 47(11): 8385-8395.
- Jin SB, Lee WJ, Park JH, Park SM, Lee DH, Yun SH. 2018. A phylogenic analysis of citrus cultivars native to Jeju using chloroplast DNA trnL-trnF and internal transcribed spacer region sequences. Hortic. Sci. Technol. 36(4): 585-597.
- Jin SB, Park JH, Park SM, Lee DH, Yun SH. 2016. Genetic phylogenetic relationship of the Jeju native Citrus 'Byungkyool' (Citrus platymamma Hort. ex Tanaka) using ITS (internal transcribed spacer) region of nuclear ribosomal DNA. Korean J. Breed. Sci. 48(3): 241-253.
- Jin SB, Park JH, Park SM, Lee DH, Yun SH. 2017. Production of citrus plants from ovule cell culture and verification of CTV-free plants. Hortic. Sci. Technol. 35(1): 121-130.
- Jin SB, Park SM, Park JS, Moon YE, Lee DH. 2025. Establishment of a regeneration system for 'Minihyang', a domestic citrus variety with high utility as a genetic resource. Plant Breed. 144(4): 529-539.
- Jin S-B, Song KJ, Riu KZ. 2007. Several factors affecting embryogenic culture maintenance and shoot regeneration in 'Miyagawa Wase' satsuma mandarin (Citrus unshiu). Hortic. Environ. Biotechnol. 48(3): 165-170.
- Jin S-B. 2005. Plant regeneration via somatic embryogenesis and agrobacterium-mediated transformation in 'Miayagawa Wase' satsuma mandarin. Cheju National University..
- Jung Y-H, Kwon H-M, Kang S-H, Kang J-H, Kim S-C. 2005. Investigation of the phylogenetic relationships within the genus citrus (Rutaceae) and related species in Korea using plastid trnL-trnF sequences. Sci. Hortic. 104(2): 179-188.
- Kayim M, Koc NK. 2006. The effects of some carbohydrates on growth and somatic embryogenesis in citrus callus culture. Scientia Horticulturae. 109(1): 29-34.
- Kazeem MI, Bankole HA, Oladokun TI, Bello AO, Maliki MA. 2020. Citrus aurantifolia (Christm.) Swingle (lime) fruit extract inhibits the activities of polyol pathway enzymes. EFood. 1(4): 310-315.
- Kazmi SK, Khan S, Kabir N, Mirbahar AA, Raziq M, Kauser N. 2015. Embryogenic callus induction, somatic embryogenesis, regeneration and histological studies of kinnow mandarin (Citrus reticulata blanco L.) From Nucellar Embryo and Epicotyl Region. Pak. J. Bot. 47(1): 305-310.
- Kepiro J, Roose M. 2007. Nucellar embryony. Pp. 141-149 in Citrus genetics. Breeding and Biotechnology: CAB International Wallingford UK..
- Kim HB, Kim JJ, Oh CJ, Yun S-H, Song KJ. 2016. Current status and prospects of molecular marker development for systematic breeding program in citrus. J. Plant Biotechnol. 43(3): 261-271.
- Kim MJ, Kim MS, Shin K, Park S, Choi C, Yun SH, et al. 2021. Comparative genetic analysis between the Jeju 'Inchangkyool' and Chinese 'Ichangensis' (Citrus ichangensis) using internal chloroplast trnL-trnF intergenic spacers and transcribed spacer sequence regions. Korean J. Breed. Sci. 53(1): 16-31.
- Kim M-J, Yun S-H, Park S-M, Jin S-B, Song K-J. 2020. Characterization of allotetraploids derived from protoplast fusion between navel orange and kumquat. In Vitro Cell Dev. Biol. Plant. 56(5): 634-643.
- Kim S, Jeong W, Min S, Bae K, Liu J. 1995. Plant regeneration from cryopreserved embryogenic cell suspension cultures of Korean rice (Oryza sativa L.) cultivars. Korean J. Plant Tissue Culture. 22(2): 115-119.
- Kochba J, Spiegel-Roy P, Neumann H, Saad S. 1982. Effect of carbohydrates on somatic embryogenesis in subcultured nucellar callus of citrus cultivars. Z. Pflanzenphysiol. 105(4): 359-368.
- Kochba J, Spiegel-Roy P, Safran H. 1972. Adventive plants from ovules and nucelli in citrus. Planta. 106(3): 237-245.
- Kunitake H, Kagami H, Mii M. 1991. Somatic embryogenesis and plant regeneration from protoplasts of 'Satsuma' mandarin (Citrus unshiu Marc. ). Sci. Hortic. 47(1-2): 27-33..
- Li CH, Lim J-D, Heo K, Kim M-J, Lee C-O, Lee J-G, et al. 2004. Long-term cold storage and plant regeneration of suspension cultured somatic embryos of eleutherococcus senticosus maxim. Korean J. Med. Crop Sci. 12(6): 494-499.
- Li D, Shi W, Deng X. 2002. Agrobacterium-mediated transformation of embryogenic calluses of Ponkan mandarin and the regeneration of plants containing the chimeric ribonuclease gene. Plant Cell Rep. 21(2): 153-156.
- Liang D, Cui M, Wu S, Ma F-W. 2012. Genomic structure, sub-cellular localization, and promoter analysis of the gene encoding sorbitol-6-phosphate dehydrogenase from apple. Plant Mol. Biol. Report. 30(4): 904-914.
- Lincy AK, Remashree AB, Sasikumar B. 2009. Indirect and direct somatic embryogenesis from aerial stem explants of ginger (Zingiber officinale Rosc.). Acta Bot. Croat. 68(1): 93-103.
- Ling J-T, Nito N, Iwamasa M, Kunitake H. 1990. Plant regeneration from protoplasts isolated from embryogenic callus of satsuma. HortScience. 25(8): 970-972.
- Liu Q, Zhu A, Chai L, Zhou W, Yu K, Ding J, et al. 2009. Transcriptome analysis of a spontaneous mutant in sweet orange [Citrus sinensis (L.) Osbeck] during fruit development. J. Exp. Bot. 60(3): 801-813.
- Moon A-R, Park J-M, Jang C-G. 2015. A phylogenetic analysis of the genus Pilea (Urticaceae) using nrDNA and cpDNA sequences. Korean J. Plant Taxon. 45(2): 158-168.
- Murashige T, Skoog F. 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15(3): 473-497.
- Murashige T, Tijcker D. 1969. Growth factor requirements of citrus tissue culture. Proc 1st Intl Citrus Symp. 3: 1156-1161.
- Nickell L, Maretzki A. 1970. The utilization of sugars and starch as carbon sources by sugarcane cell suspension cultures. Plant Cell Physiol. 11(1): 183-185.
- Park YC, Oh HW, Kang JH, Lee JS, Chin SC, Kang SH, et al. 2013. Characteristics of new satsuma mandarin cultivar 'Sangdojosaeng'. Korean J. Plant Res. 26(1): 143-147.
- Penjor T, Yamamoto M, Uehara M, Ide M, Matsumoto N, Matsumoto R, et al. 2013. Phylogenetic relationships of citrus and its relatives based on matK gene sequences. PloS One. 8(4): e62574
- Perez R, Galiana A, Navarro L, Durán-Vila N. 1998. Embryogenesis in vitro of several citrus species and cultivars. J. Hortic. Sci. Biotechnol. 73(6): 419-429.
- Praveen S, Pawar V, Ahlawat Y. 2003. Somatic embryogenesis and plant regeneration in Kinnow mandarin. J. Plant Biochem. Biotechnol. 12(2): 163-165.
- Ricci AP, Mourão Filho FdAA, Mendes BMJ, Piedade SMdS. 2002. Somatic embryogenesis in Citrus sinensis, C. reticulata and C. nobilis x C. deliciosa. Sci. Agric. 59: 41-46.
- Richa R, Kohli D, Vishwakarma D, Mishra A, Kabdal B, Kothakota A, et al. 2023. Citrus fruit: Classification, value addition, nutritional and medicinal values, and relation with pandemic and hidden hunger. J. Agric. Food Res. 14: 100718
- Sakai A, Kobayashi S, Oiyama I. 1990. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 9(1): 30-33.
- Shah D, Sajjad N, Ali R, Nazir N, Hassan S, Shah S. 2019. Sugar regulates plant growth and development under in vitro conditions. Plant signaling molecules. Woodhead Publishing. pp. 257-268.
- Soria-Guerra RE, Rosales-Mendoza S, Gasic K, Wisniewski ME, Band M, Korban SS. 2011. Gene expression is highly regulated in early developing fruit of apple. Plant Mol. Biol. Report. 29(4): 885-897.
- Starrantino A, Russo F. 1980. Seedlings from undeveloped ovules of ripe fruits of polyembryonic citrus cultivars1. HortScience. 15(3): 296-297.
- Sun Y-L, Kang H-M, Han S-H, Park Y-C, Hong S-K. 2015. Taxonomy and phylogeny of the genus citrus based on the nuclear ribosomal DNA its region sequence. Pak. J. Bot. 47(1): 95-101.
- Swingle WT. 1943. The botany of Citrus and its wild relatives in the orange subfamily. The Citrus Industry. 1: 128-474.
- Tanaka T. 1977. Fundamental discussion of citrus classification. Stud Citrol. 14: 1.
- Tingey SV, del Tufo JP. 1993. Genetic analysis with random amplified polymorphic DNA markers. Plant Physiol. 101(2): 349
- Tomaz ML, Januzzi Mendes BM, Mourão Filho fDAA, Demétrio CG, Jansakul N, Martinelli Rodriguez AP. 2001. Somatic embryogenesis in citrus spp.: Carbohydrate stimulation and histodifferentiation. In Vitro Cell Dev. Biol. Plant. 37(4): 446-452.
- Yun J-U, Kim B-K, Jin S-B, Ahn SY, Kim YH, Moon D-K, et al. 2006. Factors affecting embryogenic culture initiation from undeveloped seeds of satsuma mandarin (Citrus unshiu). Hortic. Environ. Biotechnol. 47(5): 276-279.
- Zhang M, Deng XX. 2006. Advances in research of citrus cultivars selected by bud mutation and the mechanism of formation of mutated characteristics. Int. J. Fruit Sci. 23(6): 871-876.