Abstract
Bread wheat (Triticum aestivum L.) is a fundamental crop worldwide, playing a crucial role in ensuring food security for human beings. Understanding the genetic diversity of wheat cultivars and developing genetic passports through DNA fingerprinting is essential for preventing varietal duplication enhancing breeding diversity and protecting breeders' rights globally. This study investigated the genetic diversity, phylogenetic relationships, and allelic frequencies of 32 Uzbekistan bread wheat cultivars using 66 polymorphic simple sequence repeat (SSR) markers. Unique marker alleles were identified and used to develop genetic passports for the cultivars. The results showed that the primers Barc181, Wmc522, Wmc407, Wms18, Cfa2201, Wms294, Cfd76, Wmc73, Wmc486, Cfa220, Wmc314, And Wms495 are reliable SSR markers for assessing genetic polymorphism in Uzbek wheat cultivars.
-
Key words: Bread wheat, Triticum aestivum L., SSR markers, genetic diversity, DNA fingerprinting, genetic passport
Introduction
Over the years, the selection of crops for certain traits to meet human needs has led to a narrowing of genetic diversity (
Baboeva 2023;
Joshi et al. 2023). Therefore, molecular genetic studies on germplasm accessions using genomic technologies provide an opportunity to assess existing diversity. In particular, using DNA markers in biological research is crucial for determining the genetic diversity of studied organisms and identifying valuable economic traits.
Standardized DNA barcoding methods provide a powerful tool for creating an international database of living organisms, pinpointing specific DNA regions, and accurately determining species identity. In conjunction with genomic technologies, DNA barcoding enables large-scale genetic data analysis (
Newmaster et al. 2008;
Saarela et al. 2013). This integrated approach offers solutions to complex questions within traditional breeding and genetics.
Typically, DNA barcoding utilizes standardized short DNA sequences (400-800 bp) designed for easy generation and characterization across all species on Earth (
Kress et al. 2008). The creation of an online digital DNA barcoding library allows for streamlined molecular analysis of unknown plants, facilitating species and genus identification (
Costion et al. 2016;
Yu et al. 2011). Within plant science, DNA barcoding has applications in species identification (
Valentini et al. 2009), breeding, the molecular identification of new varieties, and international copyright validation and protection (
Korir et al. 2013).
Among molecular markers, SSRs (
Simple Sequence Repeats or microsatellites) play a significant role in analyzing plant species, studying multi-allelic properties, tracking trait inheritance, and determining plant variability (
Abdurakhmonov IY 2007). Their wide genomic coverage makes them invaluable (
Kress 2017). Researchers appreciate SSR markers for their highly polymorphic nature and accuracy (
Hasnaoui N 2012). SSRs offer stability for genetic diversity studies and have applications in genetic mapping, population studies, and even individual identification through genome fingerprinting (
Agrama et al. 2007). They are widely used for genetic identification across various crops, including wheat varieties (
Adylova et al. 2018;
Turaev et al. 2023). Microsatellite (SSR) markers are short repeating sequences that may vary in the number of repeats in some regions (
Tautz 1989). Co-dominance, a wide range of allele numbers, distribution throughout the whole genome, and automation of this technology make them popular among users (
Powell 1996). Over the past few years, SSR markers have been used in the analysis of genetic diversity (
Chen 2010;
Gonzalez MA 2009;
Shiferaw E 2012), in species identification (
Shirasawa K 2013), in the evaluation of parental samples (
Malysheva L 2003) and widely used in genetic mapping (
Lu JJ 2013;
Temnykh S 2000). However, these SSR markers have also been used in the genetic research of wheat varieties.
Bread wheat (
Triticum aestivum L.), an allohexaploid species with the genome formula AABBDD (
Loginova 2018), has been extensively studied. Its entire genome has been analyzed, leading to the creation of molecular markers tailored to specific genomic regions. These markers are now available in databases and used for practical applications. Researchers employ them to identify traits in wheat samples (
Bakhadirov US. 2024;
Parmentier 2013), investigate valuable economic characteristics (
Pei N 2011), and assess genetic diversity based on geographic origin (
Huang XC 2015).
The economic value of wheat varieties is traditionally assessed through morpho-biological and quality indicators. However, these characteristics are susceptible to environmental influences, leading to phenotypic changes that may not reflect underlying genetic inheritance (
Bakhadirov U.Sh. 2024;
Sapakhova Z. 2022;
Turaev et al. 2023). Molecular identification of wheat varieties provides a robust solution, enabling rapid genetic characterization that is independent of external factors. The International Union for the Protection of New Varieties of Plants (UPOV) acknowledges the use of DNA markers to determine the distinctness, uniformity, and stability of new plant varieties (UPOV 2013).
This research aims to analyze the genetic diversity and phylogenetic relationships among Uzbek wheat varieties using SSR markers. The specific goal is to identify unique alleles associated with bread wheat cultivars and develop individual genetic passports for these varieties using DNA barcoding methods.
Materials and Methods
Plant materials
This study investigates 32 elite local wheat varieties cultivated across diverse agro-climatic zones in Uzbekistan (
Table 1). These zones range from the fertile Fergana Valley in the east to the arid plains of Karakalpakstan in the west and the mountainous regions in the south. The selected wheat varieties represent these different agro-climatic zones, ensuring a comprehensive assessment of genetic diversity across Uzbekistan’s wheat germplasm. Andijan region, located in the Fergana Valley, is characterized by a warm, continental climate with fertile irrigated lands, making it suitable for high-yielding wheat varieties. Wheat varieties cultivated in this region often exhibit resistance to diseases such as yellow rust and powdery mildew, which are prevalent in warm and humid conditions. Jizzakh region, situated in the central part of Uzbekistan, experiences a more arid climate with hot summers and cold winters. Wheat varieties cultivated in this region are often drought-tolerant and adapted to rainfed conditions. They also tend to have resistance to diseases such as leaf rust and stem rust, which are common in arid and semi-arid environments. Kashkadarya region, located in the south, has a hot, dry climate with limited rainfall. Wheat varieties grown in this region must be highly drought-tolerant and adapted to arid environments. They often exhibit resistance to diseases such as Fusarium head blight, which can thrive in hot and dry conditions.
Fresh wheat leaves were collected from the field nursery and immediately placed in liquid nitrogen until stored in a -80℃ ultra-low temperature freezer. Then the genomic DNA was isolated using the CTAB method with minor modifications (
Doyle 1987).
For polymerase chain reaction (PCR) analysis, 144 simple sequence repeat (SSR) markers with known chromosome locations were used based on the references. The PCR analysis was performed using the Hot-start program (
Adylova et al. 2018) in the ProFlex PCR System (Applied Biosystems, USA). The PCR reaction was carried out in a total volume of 10 µL per sample, using the following mixture: 1 µL genomic DNA, 1 µL 10X PCR buffer, 0.8 µL dNTPs (25 mM), 0.2 µL of each primer (forward and reverse), 0.2 µL bovine serum albumin (BSA), 0.2 µL Taq DNA polymerase (5 units/µL) and 6.6 µL nuclease-free water.
PCR amplicons were run on a 3.5% high-resolution agarose gel (Condalab, Spain) at 120 W for approximately 20-30 minutes. The gel was then visualized and documented using an Alpha Imager 3400 (Alpha Innotech, USA). Genotyping of the obtained data was performed using GelAnalyzer 19.1 software.
Data analysis
The polymorphism information content (PIC) and heterozygosity (He) of SSR markers were calculated according to the methods described by (
Collard 2005;
Collard et al. 2005). The range of PIC and He values are indicators used to determine genetic diversity, and these values can be used to distinguish the genetic diversity of wheat varieties.
Here, i - allel, n - the number of alleles at the marker, p - the frequency of alleles.
The analysis of genetic diversity is of great importance in the evaluation and utilization of plant germplasm resources and the development of new varieties. In recent years, most research on plant genetic diversity has focused primarily on SSR markers. The PIC value is determined by analyzing the polymorphism of samples using molecular markers. It measures the presence of this genetic polymorphism, specifically the structure of a genetic variant at a certain locus (location). Generally, a PIC value≥0.5 is considered highly polymorphic, 0.25≤PIC<0.5 moderately polymorphic, and PIC<0.25 low polymorphic (Botstein et al., 1980).
The He value represents the expected ratio of homozygotes and heterozygotes in a population. It calculates the heterozygote distribution of genotypes at a locus and serves as a measure of genetic diversity that can indicate a population's origin.
Essentially, He measures the probability that two randomly selected alleles from a population will be different (heterozygous) at a given locus. This reflects genetic variation at the population level, with higher He values indicating greater genetic diversity.
The range and mean values of PIC and He serves as effective indicators of genetic diversity in wheat cultivars. Higher PIC and He values indicate greater genetic variability, which may improve selection potential and adaptability to different climatic conditions. Conversely, low PIC and He values indicate genetic homogeneity, which may limit selection opportunities and make populations more susceptible to environmental influences. For statistical analysis, cluster analyses were performed using the UPGMA method in MEGA X software, and clustered heatmaps were generated using NCSS 2022 software.
Results
Banding pattern interpretation and SSR genotyping
All wheat leaf samples were collected from field nurseries at the Center of Genomics and Bioinformatics. These samples were previously provided by various regional research institutions in Uzbekistan (
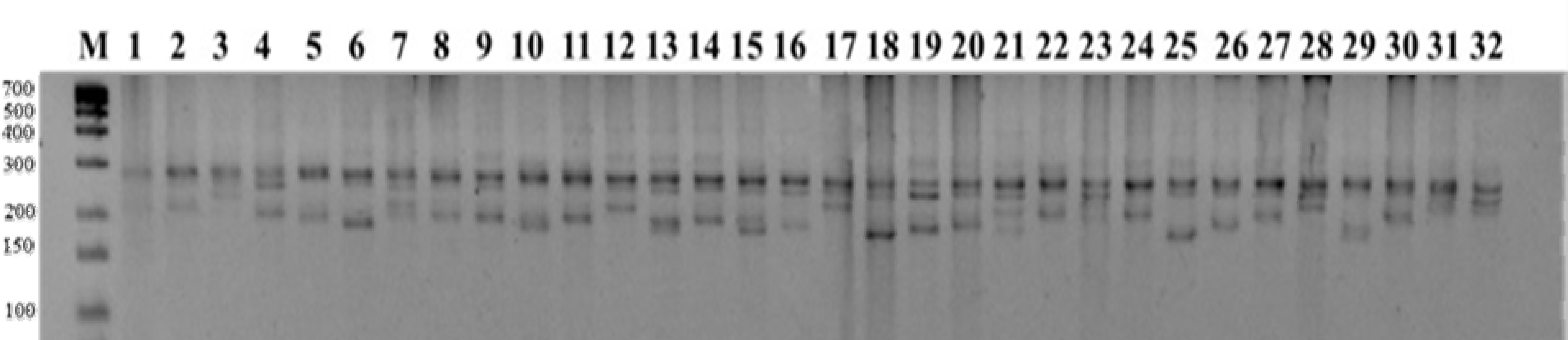
Table 1). In this study, the isolated genomic DNAs were of high quality and suitable for PCR amplification with SSR markers. Out of 144 SSR markers tested, 97 produced clear banding patterns with the DNA samples from the 32 wheat varieties and were selected for further analysis. Of these 97 amplified markers, 66 were polymorphic and produced 267 amplicons (
Fig. 1), 31 markers not amplified at any wheat genotypes, while the remaining 47 failed to amplify.
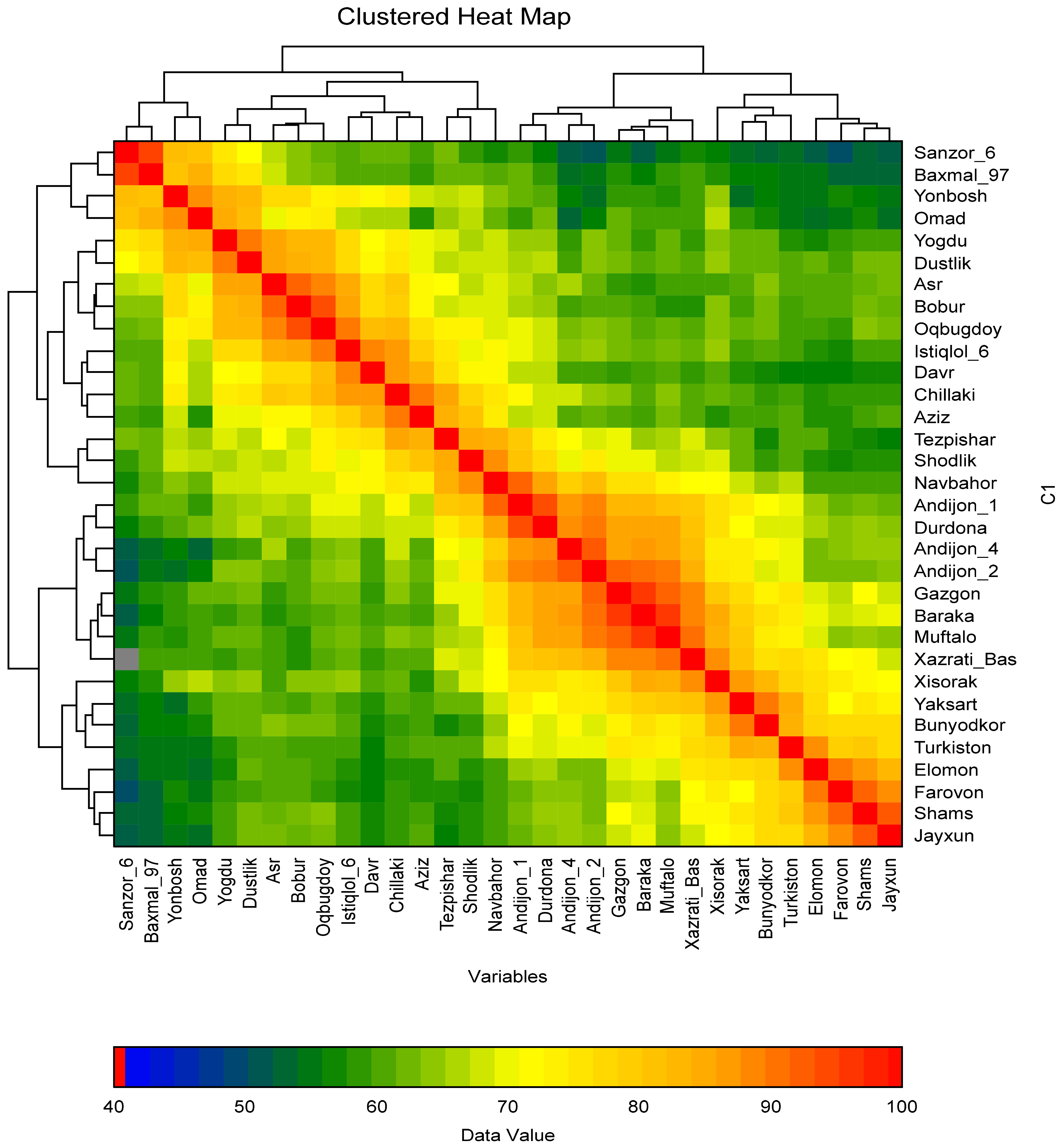
In the present study, the polymorphism information content (PIC), heterozygosity (He), and several effective alleles (Ne) of DNA makers were identified. The polymorphic markers amplified between 2 and 12 alleles. Specifically, the WMC522 marker amplified the most alleles (12), while the BARC187 marker amplified the fewest (2). The PIC value of SSR markers ranged from 0.22 (WMS18) to S0.85 (BARC181), with an average of 0.51 across all markers. The He value of the markers was also determined to assess genetic diversity among the study samples. The He values ranged from 0.12 (BARC187) to 0.86 (BARC181), with an average of 0.57. Markers with a higher effective number of alleles (NE) are better at distinguishing allele size differences between varieties. In this study, markers with NE values of 0.3 or greater indicated high diversity between varieties. Markers with 3 to 4 alleles exhibited NE values between 0.3 and 0.7 (Table S1). A Heat Map cluster was constructed to identify similarities between wheat cultivars using polymorphic markers. Cultivars that are genetically close to each other in this cluster are marked in dark red. It can be observed that the colors change, resembling the colors of the rainbow, as the genetic proximity between cultivars increases (
Fig. 2).
The heat map cluster analysis shows that the Sanzar-6 variety is genetically closest to the Bakhmal-97 and Shams varieties, but distant from the Farovon and Andijan-4 varieties. Andijon-1, Durdona, Andijon-2, Andijon-4, Gozgon, Baraka, and Muftalo cultivars were found to be genetically close to each other compared to other varieties. The genetic distances of the remaining varieties were also determined. Data analysis revealed mutual genetic polymorphisms among the local wheat varieties. Their level of genetic relatedness was represented by constructing a phylogenetic tree, and the position of each variety within the tree was determined. This phylogenetic tree was built using a total of 66 polymorphic markers, 22 with an effective allele number, and 6 microsatellite markers.
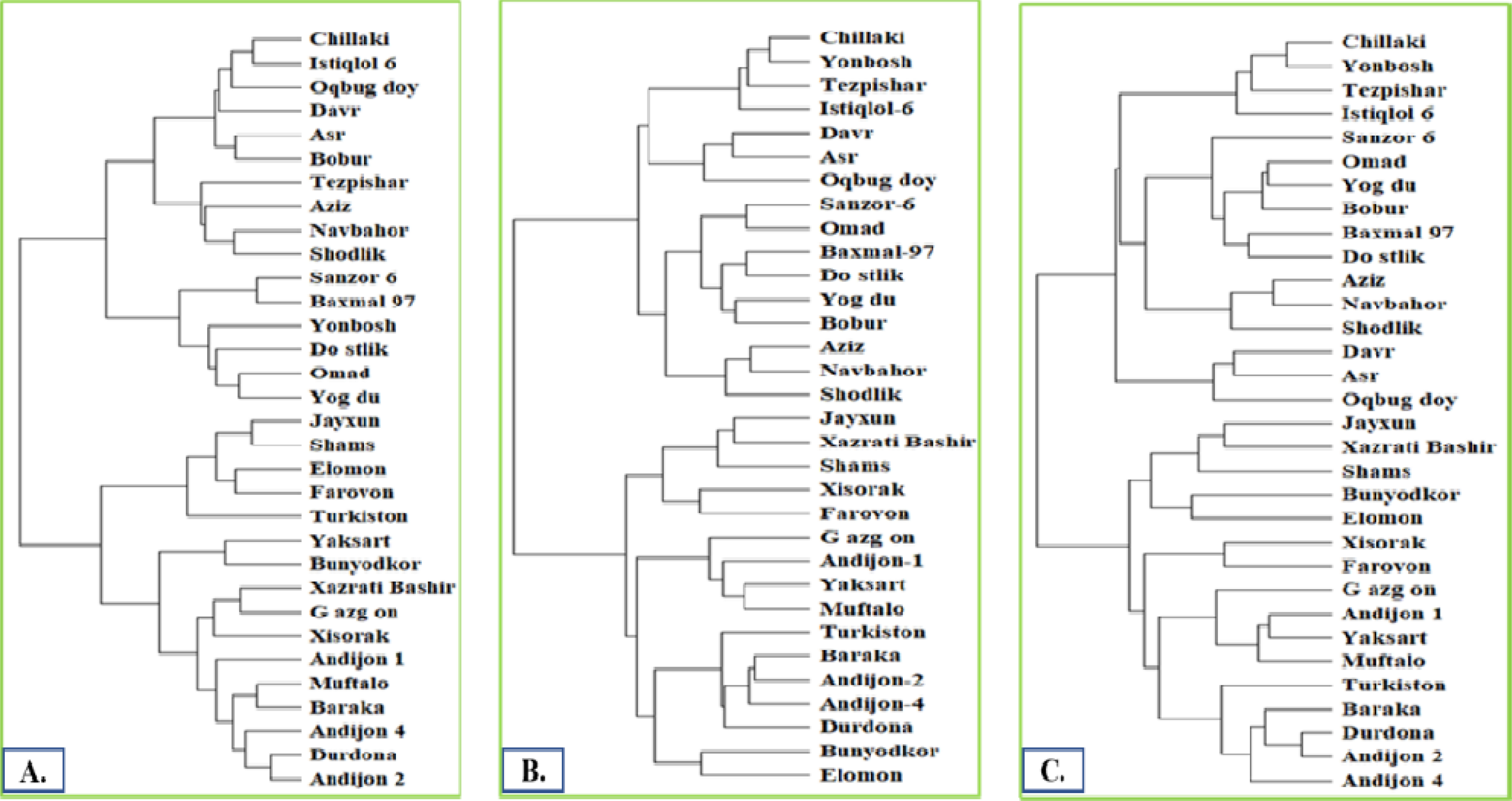
Phylogenetic analysis
Phylogenetic trees, constructed from the SSR marker data, revealed the genetic relationships among the 32 wheat varieties (
Fig. 3). Distinct clusters within the trees corresponded to the varieties' genetic similarities and differences. This information is valuable for understanding their genetic background and for future breeding programs.
The phylogenetic tree constructed using the 66 markers served as a control for further study of phylogenetic relationships. The number of markers in each phylogenetic tree was systematically reduced, starting by excluding markers with low levels of polymorphism. This process continued until changes in the phylogenetic pattern were observed. No major group changes were seen until the number of markers was reduced to 6. However, a dramatic shift in phylogenetic relationships occurred when only 5 markers remained.
DNA fingerprinting
A set of markers with unique alleles was identified and used to create genetic passports for the wheat varieties. These genetic passports provide a detailed genetic profile for each variety, helping in the identification and differentiation of the varieties. This can prevent the issue of similarity among varieties and increase the genetic diversity available for breeding
https://bise.uz/biocollection/user/12.
The main focus of this section was DNA barcoding. Microsatellite markers for each variety were recorded using letters in alphabetical sequence, and their corresponding base pairs were recorded as numerical indices (Table S2). The genetic passports of the varieties listed in the table are explained below (Table S3). A set of markers with unique alleles was identified and used to create genetic passports for the wheat varieties. These genetic passports provide a detailed genetic profile for each variety, aiding in their identification and differentiation. This helps prevent issues of varietal similarity and increases the genetic diversity available for breeding programs.
Discussion
The application of DNA barcoding for species identification has proven effective across various plant species worldwide, and this study on Uzbekistan wheat varieties contributes to that global effort. The identification of unique alleles and the development of genetic passports align with similar initiatives in other regions, such as Turkey, where SSR markers have been employed to analyze the genetic diversity and population structure of bread wheat genotypes (
Türkoğlu 2023). The consistency in findings across different regions highlights the universal applicability of molecular markers in exploring the genetic complexities of wheat cultivars.
A particularly noteworthy aspect of this study is its emphasis on DNA barcoding as a robust tool for cultivar identification and protection. The development of genetic passports based on unique marker alleles offers a reliable method for distinguishing wheat varieties, thereby safeguarding breeders' rights and preventing varietal duplication. This approach aligns with the principles advocated by the International Union for the Protection of New Varieties of Plants (UPOV), which underscores the importance of DNA markers in determining the distinctness, uniformity, and stability of new plant varieties (UPOV 2013). The study's contributions are thus significant, offering practical solutions for cultivar identification and protection in Uzbekistan.
Furthermore, the study's results are consistent with those obtained in similar research efforts worldwide. For example, the genetic diversity observed among Uzbek wheat cultivars parallels findings from studies on Turkish wheat genotypes, where SSR markers revealed substantial genetic variation (
Türkoğlu 2023). These consistent outcomes across different studies affirm the effectiveness of SSR markers in genetic diversity research and highlight their critical role in advancing wheat breeding programs. The study also identified specific SSR markers, such as BARC181, WMC522, WMC407, WMS18, and others, as reliable tools for assessing genetic polymorphism in Uzbek wheat cultivars, further contributing to the development of efficient breeding strategies.
In conclusion, this study represents a valuable contribution to wheat genetics, demonstrating the power of molecular markers in revealing the genetic diversity and phylogenetic relationships among Uzbekistani wheat cultivars. The identification of unique marker alleles and the creation of genetic passports provide crucial tools for cultivar identification, protection, and future breeding efforts. The findings align with broader trends in plant genetics, emphasizing the universal applicability of molecular markers in genetic analysis and cultivar characterization. Moreover, the research underscores the importance of DNA barcoding in safeguarding breeders' rights and promoting genetic diversity in wheat, contributing to global efforts in plant genetics and breeding.
Acknowledgments
The authors express their gratitude for the funding provided by the Agency for Innovative Development under the Ministry of Higher Education, Science, and Innovation of the Republic of Uzbekistan, which supported the completion of this research through project No. AL-8523122265.
Fig. 1PCR amplification results with SSR marker WMC522. Lane M: 25 bp DNA ladder. Lanes 1-32: Wheat cultivars arranged in the same order as listed in
Table 1.
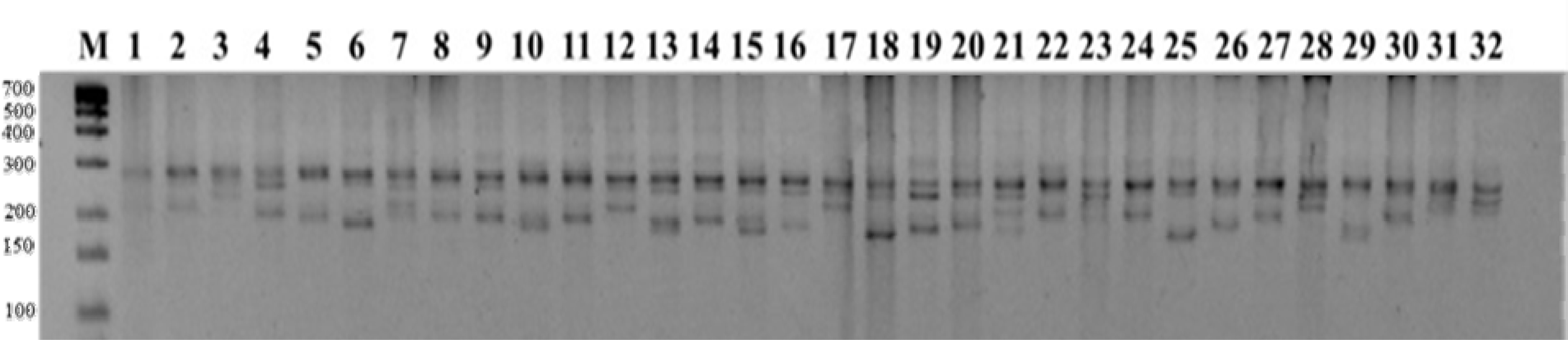
Fig. 2Heat map that describes the genetic relationships between wheat varieties.
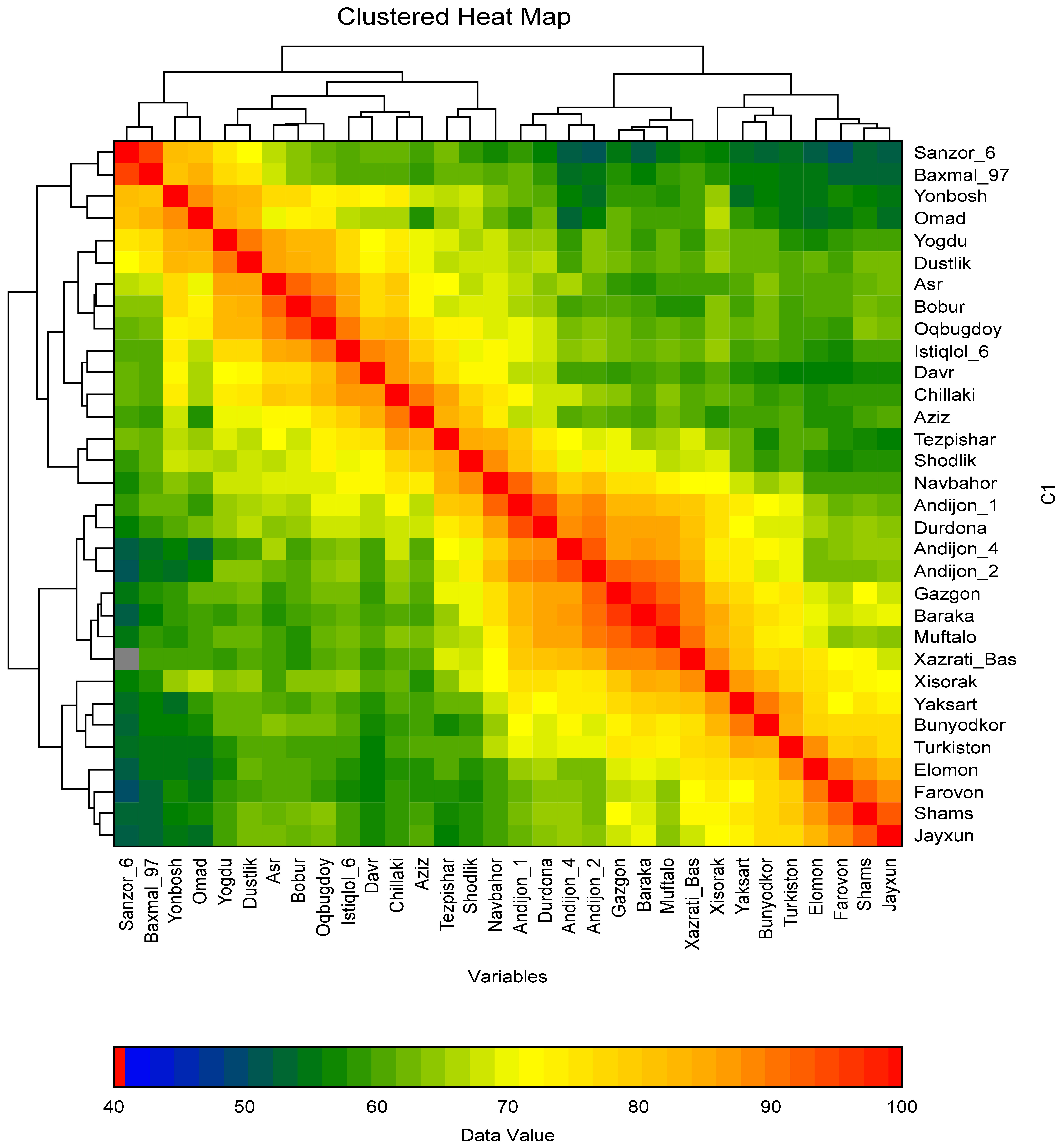
Fig. 3Phylogenetic trees were constructed based on 66 (A), 22 (B), and 6 (C) polymorphic markers.
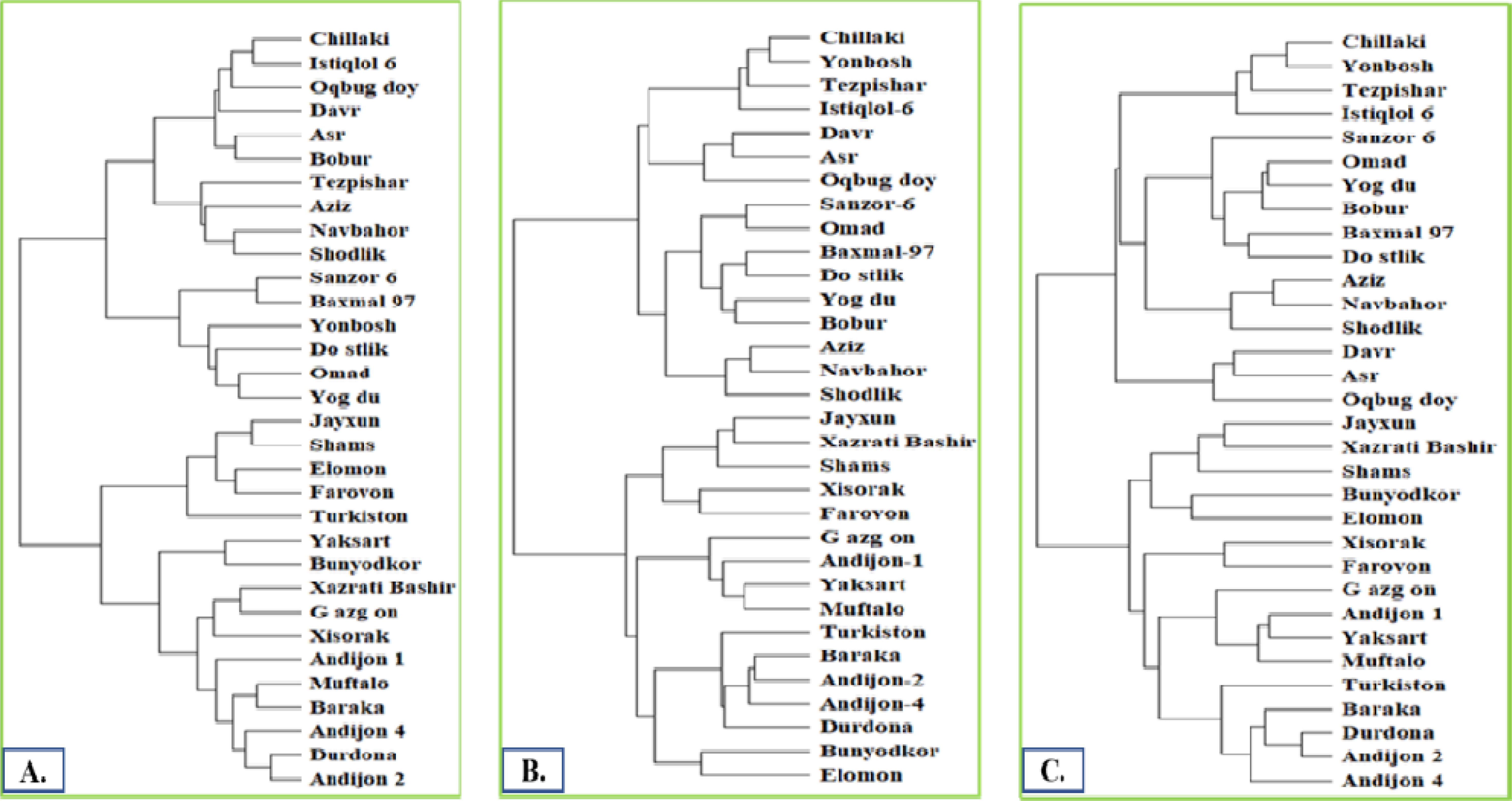
Table 1List of wheat (Triticum aestivum) germplasm accessions used in the study.
Table 1
|
No |
Wheat cultivars |
Main characteristics of cultivars provided by the authors |
Region of Origin |
Collection |
|
1 |
Chillaki |
Early maturity, moderately resistant to yellow rust, resistant to powdery mildew |
Andijan, Uzbekistan |
RICLCI
|
|
|
2 |
Yonbosh |
Early maturity, yellow rust resistant, drought tolerant |
Jizzakh, Uzbekistan |
RIRAII
|
|
|
3 |
Jaykhun |
Medium maturity, moderately tolerant to cold, highly resistant to yellow and leaf rust |
Jizzakh, Uzbekistan |
RIRA |
|
|
4 |
Tezpishar |
Facultative cultivar, early maturity, cold and heat tolerant, leaf rust and powdery mildew resistant, moderately resistant to yellow rust |
Jizzakh, Uzbekistan |
RIRA |
|
|
5 |
Istiklol-6 |
Early maturity, highly resistant to yellow and leaf rust, powdery mildew, cold and drought tolerant |
Jizzakh, Uzbekistan |
RIRA |
|
|
6 |
Hisorak |
Medium maturity, moderately cold tolerant, highly resistant to yellow and leaf rust |
Kashkadarya, Uzbekistan |
RIASRIII
|
|
|
7 |
Shams |
Medium maturity, moderately tolerant to cold and drought, moderately resistant to yellow and leaf rust, resistant to powdery mildew |
Kashkadarya, Uzbekistan |
RIASR |
|
|
8 |
Khazrati Bashir |
Early maturity, moderately tolerant to cold, heat and drought tolerant, moderately resistant to yellow and leaf rust, resistant to powdery mildew |
Kashkadarya, Uzbekistan |
RIASR |
|
|
9 |
Davr |
Medium maturity, moderately susceptible to leaf rust, resistant to yellow rust, moderately resistant to cold and highly resistant to drought |
Andijan, Uzbekistan |
RICLC |
|
|
10 |
Okbugdoy |
Medium maturity, moderately susceptible to yellow rust. |
Andijan, Uzbekistan |
RICLC |
|
|
11 |
Asr |
Medium maturity, resistant to powdery mildew, leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
12 |
Gozgon |
Medium maturity, highly tolerant to cold, heat and drought, highly resistant to yellow rust, resistant to powdery mildew. |
Kashkadarya, Uzbekistan |
RIASR |
|
|
13 |
Andijan-1 |
Medium maturity, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
14 |
Yaksart |
Medium maturity, highly tolerant to cold, drought tolerant, resistant to powdery mildew. |
Kashkadarya, Uzbekistan |
RIASR |
|
|
15 |
Sanzar-6 |
Facultative cultivar, moderately resistant to yellow rust, highly resistant to powdery mildew, cold and drought tolerant. |
Jizzakh, Uzbekistan |
RIRA |
|
|
16 |
Muftalo |
Medium maturity, resistant to leaf and yellow rust, moderately tolerant to cold and drought |
Andijan, Uzbekistan |
RICLC |
|
|
17 |
Omad |
Early maturity, resistant to leaf and yellow rust, moderately tolerant to cold and drought |
Andijan, Uzbekistan |
RICLC |
|
|
18 |
Bakhmal-97 |
Facultative cultivar, weakly resistant to yellow rust |
Jizzakh, Uzbekistan |
RIRA |
|
|
19 |
Dustlik |
Early maturity, resistant to yellow and leaf rust |
Jizzakh, Uzbekistan |
RIRA |
|
|
20 |
Aziz |
Early maturity, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
21 |
Turkiston |
Medium maturity, cold tolerant, heat, drought and salt tolerant, resistant to powdery mildew |
Kashkadarya, Uzbekistan |
RIASR |
|
|
22 |
Baraka |
Early maturity, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
23 |
Navbakhor |
Medium maturity, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
24 |
Bunyodkor |
Early maturity, moderately tolerant to cold, heat and drought tolerant, highly resistant to yellow and leaf rust, resistant to powdery mildew. |
Kashkadarya, Uzbekistan |
RIASR |
|
|
25 |
Durdona |
Medium maturity |
Andijan, Uzbekistan |
RICLC |
|
|
26 |
Yogdu |
Early maturity, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
27 |
Andijon-4 |
Early maturity, moderately resistant to leaf and stem rust, powdery mildew, and moderately resistant to fusarium head blight |
Andijan, Uzbekistan |
RICLC |
|
|
28 |
Shodlik |
Medium maturity, moderately resistant to drought and cold, resistant to leaf and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
29 |
Andijon-2 |
Medium maturity, resistant to leaf and stem rust, susceptible to powdery mildew and yellow rust |
Andijan, Uzbekistan |
RICLC |
|
|
30 |
Bobur |
Early maturity, yellow and leaf rust resistant |
Andijan, Uzbekistan |
RICLC |
|
|
31 |
Elomon |
Early maturity, leaf and stem rust resistant, fusarium head blight resistant, highly tolerant to heat and drought |
Kashkadarya, Uzbekistan |
RIASR |
|
|
32 |
Farovon |
Early maturity, moderately tolerant to cold, drought tolerant, moderately resistant to yellow and leaf rust, resistant to powdery mildew |
Jizzakh, Uzbekistan |
RIRA |
References
- Abdurakhmonov IY BZT, Saha S, Pepper AE, Musaev JA, Almatov A, Shermatov SE, et al. 2007. Microsatellite markers associated with lint percentage trait in cotton, Gossypium hirsutum. Euphytica. 156: 141-156.
- Adylova AT, Norbekov GK, Khurshut EE, Nikitina EV, Kushanov FN. 2018. SSR analysis of the genomic DNA of perspective Uzbek hexaploid winter wheat varieties. Vavilovskii Zhurnal Genet Selektsii. 22: 634-639.
- Agrama HA, Eizenga GC, Yan W. 2007. Association mapping of yield and its components in rice cultivars. Mol Breeding. 19: 341-356.
- Baboeva SS. 2023. Climate change impact on Chlorophyll content and grain yield of bread wheat (Triticum Aestivum L.). SABRAO J Breed Genet. 55: 1930-1940.
- Bakhadirov US, YUK , Turaev OS, Erjigitov DS, Mansurov KG, Iskandarov AA, Khidirov MT, et al. 2024. Determining close relationships between wheat cultivars (Triticum Aestivum) using DNA markers. Front Health Inform. 13: 5807-5814.
- Bakhadirov USh, Turaev OS, Erjigitov DSh, Dolimov AA, Tursunmurodova BT, Fayzullaev AZ, et al. 2024. Determining aphid resistance genes in bread wheat (Triticum cesium L.) cultivates using DNA markers. SABRAO J Breed Genet. 56: 582-590.
- Buckler ES, Holland JB, Bradbury PJ, Acharya CB, Brown PJ, Browne C, et al. 2009. The genetic architecture of maize flowering time. Science. 325: 714-718.
- Chen S, Yao H, Han J, Liu C, Song J, Shi L, et al. 2010. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS One. 5(1): e8613
- Collard BCY, Jahufer MZZ, Brouwer JB, Pang ECK. 2005. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica. 142: 169-196.
- Costion C, Lowe A, Rossetto M, Kooyman R, Breed M, Ford A, et al. 2016. Building a plant DNA barcode reference library for a diverse tropical flora: An example from queensland, Australia. Diversity. 8(1): 5
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin. 19: 11-15.
- Edwards DAH, Taylor D, Savolainen V, Hawkins JA. 2008. DNA barcoding of a large genus, Aspalathus L. (Fabaceae). Taxon. 57: 1317-1327.
- Gonzalez MA, Baraloto C, Engel J, Mori SA, Pétronelli P, Riéra B, et al. 2009. Identification of Amazonian trees with DNA barcodes. Plos one. 14: e7483
- Hasnaoui NBA, Sebastiani F, Mars M, Zhang D, Vendramin GG. 2012. Molecular genetic diversity of Punica granatum L. (pomegranate) as revealed by microsatellite DNA markers (SSR). Gene. 493: 105-112.
- Huang XC, Ci XQ, Conran JG, Li J. 2015. Application of DNA barcodes in Asian tropical trees- A case study from Xishuangbanna nature reserve, Southwest China. Plos one. 10: e0129295
- Jaakola L, Suokas M, Häggman H. 2010. Novel approaches based on DNA barcoding and high-resolution melting of amplicons for authenticity analyses of berry species. Food Chem. 123: 494-500.
- Joshi BK, Ghimire KH, Neupane SP, Gauchan D, Mengistu DK. 2023. Approaches and advantages of increased crop genetic diversity in the fields. Diversity. 15: 603
- Korir NK, Han J, Shangguan L, Wang C, Kayesh E, Zhang Y, et al. 2013. Plant variety and cultivar identification: Advances and prospects. Crit Rev Biotechnol. 33: 111-125.
- Kress WJ. 2017. Plant DNA barcodes: Applications today and in the future. J Syst Evol. 55: 291-307.
- Kress WJ, Erickson DL. 2008. DNA barcodes: genes, genomics, and bioinformatics. Proc Natl Acad Sci USA. 105: 2761-2762.
- Kumar S, Kahlon T, Chaudhary S. 2011. A rapid screening for adulterants in olive oil using DNA barcodes. Food Chem. 127: 1335-1341.
- Loginova DS, Olga . 2018. The genome of bread wheat triticum aestivum L.: Unique structural and functional properties. Russ J Genet. 4: 403-414.
- Lu JJ KJ, Feng SG, Zhao HY, Liu JJ, Wang HZ. 2013. Transferability of SSR markers derived from Dendrobium nobile expressed sequence tags (ESTs) and their utilization in Dendrobium phylogeny analysis. Sci Hortic. 158: 8-15.
- Malysheva LST, Matzk F, Röder M, Ganal M. 2003. Molecular cytogenetic analysis of wheat-barley hybrids using genomic in situ hybridization and barley microsatellite markers. Genome. 46: 314-322.
- Newmaster SG, Fazekas AJ, Steeves RA, Janovec J. 2008. Testing candidate plant barcode regions in the Myristicaceae. Mol Ecol Resour. 8: 480-490.
- Parmentier I, Duminil J, Kuzmina M, Philippe M, Thomas DW, Kenfack D, et al. 2013. How effective are DNA barcodes in the identification of African rainforest trees? PloS one. 8: e54921
- Pei N, Lian JY, Erickson DL, Swenson NG, Kress WJ, Ye WH, et al. 2011. Exploring tree-habitat associations in a Chinese subtropical forest plot using a molecular phylogeny generated from DNA barcode loci. PloS one. 6: e21273
- Powell W, Machray GC, Provan J. 1996. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1: 215-222.
- Saarela JM, Sokoloff PC, Gillespie LJ, Consaul LL, Bull RD. 2013. DNA barcoding the Canadian Arctic flora: core plastid barcodes (rbcL + matK) for 490 vascular plant species. Plos one. 8: e77982
- Sapakhova Z, Irkitbay A, Madenova A, Suleimanova G. 2022. Mitigation effect of salicylic acid on wheat (Triticum cesium L.) under drought stress. Res Crops. 23: 267-275.
- Shiferaw E Pè ME, Porceddu E, Ponnaiah M. 2012. Exploring the genetic diversity of Ethiopian grass pea (Lathyrus sativus L.) using EST-SSR markers. Mol Breed. 30: 789-797.
- Shirasawa K, Ishii K, Kim C, Ban T, Suzuki M, Ito T, et al. 2013. Development of Capsicum EST-SSR markers for species identification and in silico mapping onto the tomato genome sequence. Mol Breed. 31: 101-110.
- Stoeckle MY, Gamble CC, Kirpekar R, Young G, Ahmed S, Little DP. 2011. Commercial teas highlight plant DNA barcode identification successes and obstacles. Sci Rep. 1: 42
- Tautz D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res. 17: 6463-6471.
- Temnykh S, Park WD, Ayres N, Cartinhour S, Hauck N, Lipovich L. 2000. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor. Appl. Genet. 100: 697-712.
- Türkoğlu A, Haliloğlu K, Mohammadi SA, Öztürk A, Bolouri P, Özkan G, et al. 2023. Genetic diversity and population structure in Türkiye bread wheat genotypes revealed by simple sequence repeats (SSR) markers. Genes (Basel). 14: 1182
- Turaev OS, Baboev SK, Ziyaev ZM, Norbekov JK, Erjigitov DS, Bakhadirov US, et al. 2023. Present status and future perspectives of wheat (Triticum Aestivum L.) research in Uzbekistan. SABRAO J Breed Genet. 55: 1463-1475.
- Valentini A, Pompanon F, Taberlet P. 2009. DNA barcoding for ecologists. Trends Ecol Evol. 24: 110-117.
- Yu J, Xue JH, Zhou SL. 2011. New universalmatKprimers for DNA barcoding angiosperms. J Syst Evol. 49: 176-181.