Abstract
Hairs on the leaf are an important agronomic characteristic for rice growth and farming. The segregation ratio of pubescence in the F2 population showed that the pubescence on the leaf is controlled by a single dominant gene. Fine mapping for the gene was carried out by producing an Indel (insertion-deletion) primer based on BSA-Seq data. Results of the analysis revealed within the candidate site the presence of HL6, a published pubescence gene. By haplotype analysis of HL6 in the core-collection data, SNPs were found in HL6’s exons between the plants. These sequence variations can be useful for marker-assisted selection.
-
Key words: Rice (Oryza sativa L.), Pubescence, BSA-Seq, HL6
INTRODUCTION
Leaf growth is related to many plant functions such as photosynthesis and transpiration which ultimately affect yield (
Wang et al. 2011). Many genes that control rice leaves’ morphological traits such as rolled leaf (rl), glabrous leaf (gl) have been identified and their mode of inheritance have been elucidated using mutant lines (
Kinoshita 1995). DNA markers were also used in detecting quantitative trait loci (QTLs) and map-based cloning of genes responsible for such traits (
Yonemaru et al. 2010).
Pubescence is an important characteristic for plant pro-tection, especially against insect infestation, drought, and UV radiation. Leaves with an abundance of macro-hairs has shown an increased in leaf surface temperature and water-use efficiencies (WUEp) by restricting leaf transpi-ration (
Hamaoka et al. 2017).
Pubescence is characterized by trichomes gathering on the plant surface, consisting of unicellular or multicellular exogenous outgrowths on leaves, stems, and root surfaces. Pubescence may be related to the development of glandular cells and thus are called glandular hairs. Glandular hairs can accumulate and secrete alkaloids such as nicotine and terpenoids as well as some other chemical substances against injurious insects. Most plants also have non-glandular hairs that can strengthen resistance to abiotic stress such as extremely temperatures, drought, and ultraviolet irradiation to promote normal growth (
Wang et al. 2009).
Glabrous rice varieties are widely cultivated in America and Africa while most varieties cultivated in Asia are pubescent (
Khush et al. 2001). In agriculture, glabrous rice is advantageous in the harvesting and packaging process due to the plants being less itchy and less prone to accu-mulate dust. However, trichomes are thought to be im-portant for plant defense against biotic and abiotic stresses. Understanding the
HL6 gene can help produce rice vari-eties that are suited for agricultural field workers while enhancing plant adaptations to stress.
MATERIALS AND METHODS
Plant materials and phenotype evaluation
Cheongsongaengmi4 (CH4, RWG-130) showing long leaf hairs japonica rice and Hatadani (HA, RWG-054) showing short leaf hairs indica rice were selected as parental to produce the F
2 population for phenotyping screening and genetic studies related to pubescence (
Fig. 1). Pubescence on the leaf surface can be easily observed when it is bent as the hairs reflect light under sunlight in field.
Total genomic DNA was extracted from leaves using modified Cetyl Trimethylammonium Bromide (CTAB) method as previously described by
Porebski et al. (1997). DNA concentration was quantified using a spectrophoto-meter (NanoDrop One, ThermoScientific). DNA samples were diluted to a concentration of 50 ng/uL and were used for genotyping.
L-bulk and S-bulk with 10 plants each whose DNA demonstrated differences with hairs in F
2 population were prepared. BSA-seq analysis was used QTL-seq program by Takagi in default procedure for evaluating the SNP-index and detecting association SNPs or indels for target traits. QTL mapping as a traditional method showed low efficiency due to its need to process data derived from samples across multiple generations of breeding. QTL-seq is a more rapid alternative to identify plant QTLs by whole-genome rese-quencing of DNAs from two populations showing extreme opposite trait values for a given phenotype in a segregating progeny (
Takagi et al. 2013).
To search the site for the pubescence gene, BSA-Seq was used with BIONEER’s next generation sequencing service to process the raw data from the sampled plants while using IRGSP 1.0 as reference.
Narrow down the QTL region and candidate gene
RESULTS AND DISCUSSION
Phenotype analysis
Out of the 208 F
2 plants, 155 plants showed longer hairs similar to CH4 and 53 plants showed shorter hairs following HA. This ratio of pubescence shows that the trait is con-trolled by one dominant gene following Mendelian genetics as verified using a chi-squared test at df = 1 (
Table 1).
A total of 10 plants for each distinct phenotype was prepared in bulk as source material for DNA collection. Differences between the two parents were also compared to obtain multiple SNP-index level. Total SNP-index level was calculated by subtracting the SNP-index values. Raw NGS data were processed using Python (
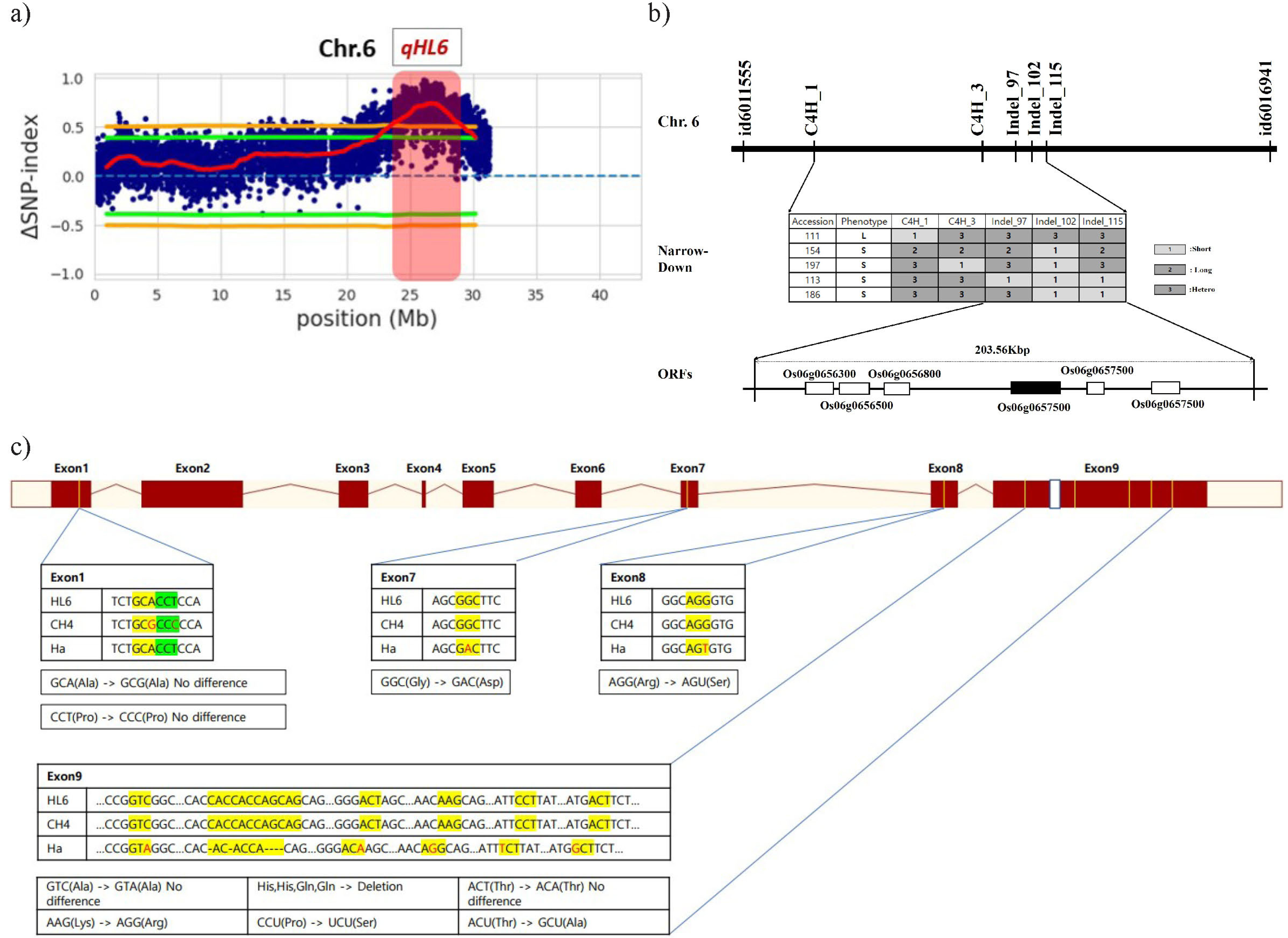
Sugihara et al. 2022). The threshold limit was set to 99% confidence for the selection of candidate regions and SNPs. BSA-Seq was carried out using the sequence information within CH4 and Ha by BIONEER’s NGS service. The candidate region (
qHL6) showing a SNP-index score of over 0.5 was iden-tified to be between 24.8Mb and 29.8 Mb in chromosome 6 (
Fig. 2a).
Narrowed-down results using indel markers show pubes-cence gene is place between Indel-102 and Indel-115. The size of region is around 203.56 Kbp. Within the specified region, a published gene controlling pubescence named
HL6 (
Os06g0657500) exists on 203.56 Kbp region. (
Sun et al. 2017;
Fig. 2b). According to existing studies,
HL6 was identified to be related to trichome formation which is reg-ulated by the AP2/ERF transcription factor. This gene reg-ulates trichome elongation by interaction with OsWOX3B, a homeodomain-containing protein related to the initiation of trichome formation. Our results confirmed that
HL6 lo-cated in chromosome 6 showed direct functions to control the glabrous leaf phenotype in the Indica and Japonica rice subspecies contrary to the
BKL which controls this phe-notype in wild rice (
Oryza nivara). In this study, we analyzed sequences of
HL6 in CH4 and HA to search which exons in
HL6 are directly affecting hair length production.
To search which exon is related to pubescence, rese-quencing and haplotype analysis for the exons of
HL6 in CH4, HA, and IRGSP1.0 (used as reference) were compared. Between CH4 and HA, 9 SNPs and 6 deletions were detected. A total of seven alterations of SNPs are responsible for changes in the translated amino acid sequence (
Fig. 2c).
ACKNOWLEDGEMENTS
This work was supported by the Rural Development Administration, Republic of Korea (RS-2022-RD010353).
Fig. 1Difference of leaf hairs in parents (Left: Hatadani, Right: Cheongsongaengmi4).

Fig. 2Genetic analysis of qHL6. (a) Distribution of △SNP-index from BSA-Seq on chromosome 6. (b) Linkage map construction and Narrow down the region of qHL6 and candidate gene screening. Information of primers are in
Supplementary Table 1. (c) Sequence variation between Hatadani and Cheongsongaengmi4 on the exons of HL6.
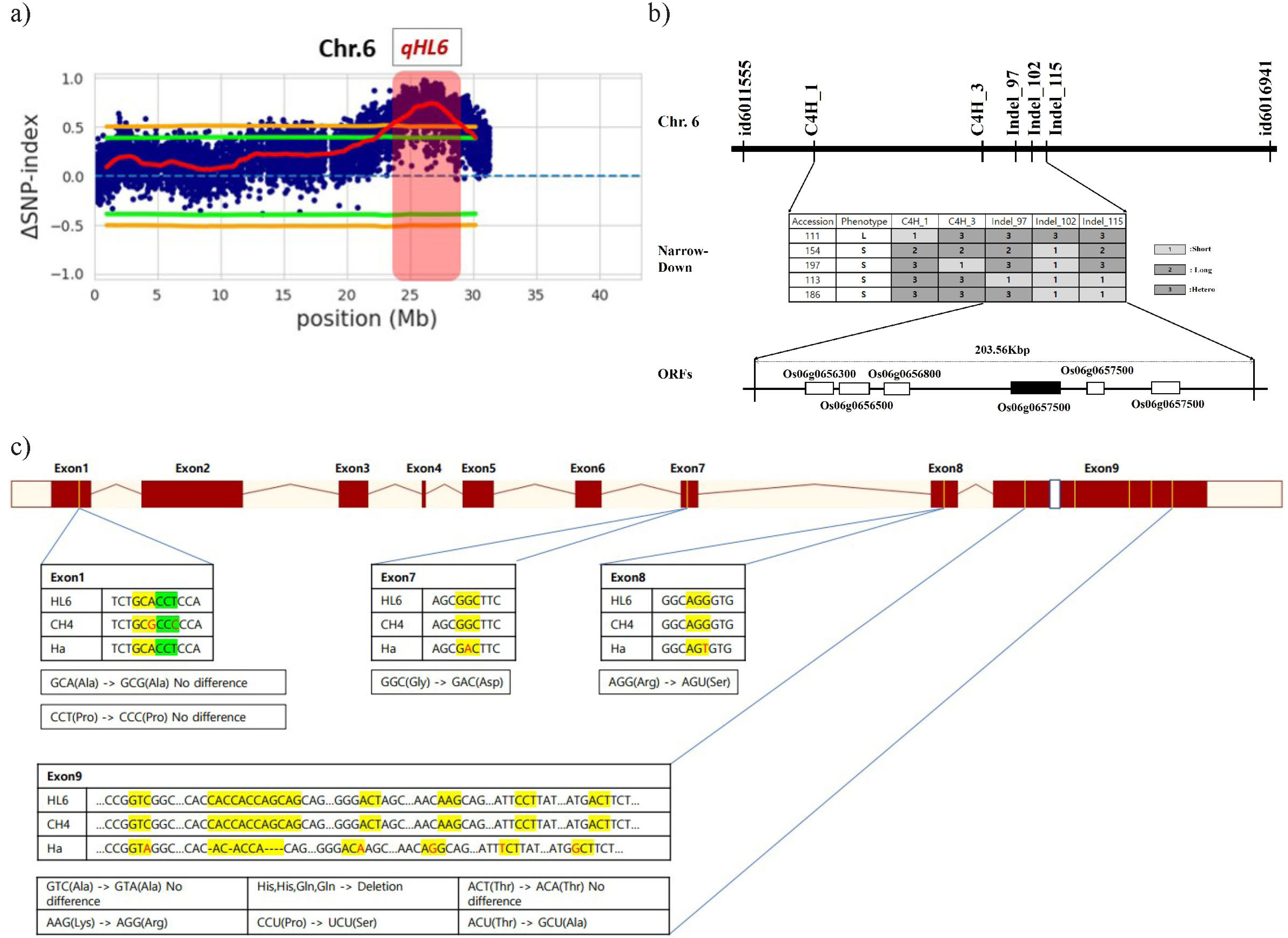
Table 1Segregation distribution of leaf hair length.
Table 1
|
Long leaf hair |
Short leaf hair |
Total |
x2
|
|
Actual |
155 |
53 |
208 |
0.026**
|
|
Expected |
156 |
52 |
208 |
References
- Hamaoka N, Yasui H, Yamagata Y, Inoue Y, Furuya N, Araki T, et al. 2017. A hairy-leaf gene, BLANKET LEAF, of wild Oryza nivara increases photosynthetic water use effciency in rice. Rice.. 10(1): 1-11.
- Khush GS, Brar DS, Hardy B, 2001. Rice genetics IV. Proceedings of the Fourth International Rice Genetics Symposium, 22-27 October 2000, Los Baños, Philippines. Enfield, NH (USA). editors. Rice genetics IV. International Rice Research Institute. Science Publishers Inc. and Los Baños (Philippines).
- Kinoshita T. 1995. Report of committee on gene symboli-zation, nomenclature and linkage groups. Rice Genet. Newslett.. 12: 9-154.
- Porebski S, Bailey LG, Baum BR. 1997. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol omponents. Plant Mol Biol Rep.. 15: 8-15.
- Sugihara Y, Young L, Yaegashi H, Natsume S, Shea DJ, Takagi H, et al. 2022. High performance pipeline for MutMap and QTL-seq. Peer J.. 10: e13170
- Sun W, Gao D, Xiong Y, Tang X, Xiao X, Wang C, Yu S. 2017. Hairy Leaf 6, an AP2/ERF Transcription Factor, Interacts with OsWOX3B and Regulates Trichome Formation in Rice. Molecular Plant.. 10(11): 1417-1433.
- Takagi H, Abe A, Yoshida K, Kosugi S, Natsume S, Mitsuoka C, et al. 2013. QTL-seq: rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J.. 74(1): 174-183.
- Wang D, Sun SX, Gao FY, Lu XJ, Li ZH, Ren GJ. 2009. Mapping a rice glabrous gene using simple sequence repeat markers. Rice Sci.. 16(2): 93-98.
- Wang P, Zhou G, Yu H, Yu S. 2011. Fine mapping a major QTL for flag leaf size and yield-related traits in rice. Theor Appl Genet.. 123: 1319-1330.
- Yonemaru JI, Yamamoto T, Fukuoka S, Uga Y, Hori K, Yano M. 2010. Q-TARO: QTL Annotation Rice Online Database. Rice.. 3(2): 194-203.
- Zeng Y, Zhu Y, Lian L, Xie H, Zhang J, Xie H. 2013. Genetic analysis and fine mapping of the pubescence gene GL6 in rice (Oryza sativa L.). Chin. Sci. Bull.. 58: 2992-2999.
Citations
Citations to this article as recorded by