Abstract
Phytophthora root and stem rot (PRR) of soybean is a severe disease that causes significant economic losses in soybean-growing countries. The causal agent, Phytophthora sojae, is a soil-borne oomycete that causes pre- and post-emergence damping-off of soybean in poorly drained soils. PRR has not been a severe problem in South Korea; however, as the cultivation of soybean in paddy fields recently increased, there is a growing interest because the incidence of PRR can be extensively expanded. R-gene mediated resistance is known as the main strategy for the management of this disease. On the other hand, partial resistance has also been known to provide more effective disease management than the Rps (Resistance to Phytophthora sojae) resistance. Responses of domestic soybean cultivars to a few P. sojae isolates were recently reported, while phenotypic distribution of numerous germplasms is mostly unknown. The objectives of this review are to summarize published studies conducted on PRR, to suggest future directions of genetic researches and breeding to the target pathogen. This review will discuss the followings; i) a brief introduction to PRR and the causal agent P. sojae, ii) types of genetic resistance to P. sojae and findings of genes/QTL conditioning to resistance, iii) changes in virulence pathotype of P. sojae populations, and iv) current status and perspective of genetic/genomic researches on the interaction of soybean and P. sojae in the South Korea.
-
Key words: Soybean, Phytophthora root and stem rot, Rps, Partial resistance, Virulence pathotype
INTRODUCTION
PRR was first identified in 1996 in Chungnam province in South Korea (
Jee et al. 1998). In recent years, incidences of soybean diseases caused by soil-borne pathogens, including PRR, have been more frequently reported in South Korea (
Kang et al. 2019;
Kang et al. 2020;
Ko et al. 2020). It is considered that as paddy-fields begin to be utilized in the soybean production, root rot diseases tends to occur more in such poorly drained soils. Consequently, attention for such soybean diseases is steadily increasing. The objectives of this review are to introduce soybean genetic researches and breeding efforts for resistance to PRR, and to suggest future directions of researches and breeding programs in South Korea to protect soybean production against the PRR.
P. sojae is known to be host-specific, but it can also infect the lupine species (
Lupinus spp.) (
Dorrance and Grünwald 2009). This pathogen predominantly infects soybean pre-emergence, but as oospores germinate throughout the growing season, soybean plants can be attacked by
P. sojae during any stage of the plant development (
Dorrance 2018). When it infects susceptible soybean at the early stage of growth, consequently, seed decay and seedling damping-off would be the critical disease symptoms (
Schmitthenner 1985). Root rot and brown stem lesions typically develop in susceptible plants during the later stage of growth, along with leaf yellowing, wilting, and finally, plant death, which subsequently results in reduced plant vigor and decreases in the seed yield (
Schmitthenner 1985).
It is a monocyclic disease, and
P. sojae can withstand freezing and cold temperatures as oospores in both soil and plant debris for several years (
Schmitthenner 1985). In warm and highly wet soils, oospores of
P. sojae germinate to form mycelia, and then the mycelia produce sporangia and zoospores under those wet conditions. Zoospores are the primary infectious agents of this process. They are attached to soybean roots via chemotaxis, which is done by isoflavones, such as genistein and daidzein (
Morris and Ward 1992). Zoospores encyst, germinate, and penetrate the cell wall of soybean roots within a few hours after the inoculation (
Morris and Ward 1992). Afterward,
P. sojae colonizes the root and stem tissues as reproducing oospores (
Schmitthenner 1985).
PRR is managed predominantly through the development of cultivars with one or two dominant resistance genes (
Rps). Although the
Rps genes are useful, such resistance is race-specific and, thus, they would be effective in limited numbers of
P. sojae isolates. Each of the
Rps genes have been often effective for less than 15 years as new
P. sojae isolates that can evade
Rps-mediated defense emerge (
Schmitthenner 1985;
Dorrance et al. 2003). Some of the
Rps genes (
Rps1a, 1b, 1c, 1k, 3a, and
6), individually or in combinations of two
Rps, have been deployed into cultivars (
Dorrance and Schmitthenner 2000;
Slaminko et al. 2010). Other
Rps genes (
1d, 2, 3b, 3c, 4, 5, and
7) have not been commercially deployed because
P. sojae populations that can overcome these
R genes already existed in fields (
Abney et al. 1997). Therefore, understanding the regional distribution of pathotypes of
P. sojae is vital for managing the PRR by deploying a few
Rps genes into new soybean cultivars.
The diversity of the
P. sojae population has been widely studied since the late 1960s in North America, including the United States and Canada (
Dorrance 2018). In these locations, the virulence pathotypes of
P. sojae isolates were determined based on the responses following the hypocotyl inoculation on 15 differential varieties (
Dorrance et al. 2004). The differentials consisted of universal susceptible “Williams” and near-isogenic lines carrying a single
Rps gene in the Williams background (
Dorrance et al. 2004).
Rps1a,
1d, and
1k allele had been widely used in controlling PRR during the 1980s, and subsequently, new races with
Rps1d and
1k virulence emerged during the 1990s (
Schmitthenner 1985;
Abney et al. 1997). Subsequently, the virulence prevalence of
Rps1a, 1b, 1c, and
1k has continued to increase in the majority of north-central states in the United States, resulting in that 30-100% of the collected isolates were virulent toward
Rps1a, 1b, 1c, and
1k (
Dorrance et al. 2016). Until 2000, there were 55 races reported based on the reactions of eight soybean differentials for the
Rps1a, 1b, 1c, 1d, 1k, 2, 3a, 6, and
7 genes, which were virulent to a few numbers of
Rps genes (
Grau et al. 2004). In the early 2000s, the pathotypes of
P. sojae isolates were observed to be more complex and diverse (
Kaitany et al. 2001;
Dorrance et al. 2003;
Jackson et al. 2004;
Malvick and Grunden 2004;
Nelson et al. 2008). In a study conducted during 2012-2013, over 200 unique virulence pathotypes were identified from more than 800 isolates, which were collected in the north-central region of the United States (
Dorrance et al. 2016).
Diversity or changes in pathogenecity of
P. sojae were also studied in other countries. In China, for instance,
P. sojae was first identified in Heilongjiang in 1991 (
Su and Shen 1993), and multiple races were subsequently identified until the early 2000s (
Ma et al. 2005). Until 2015, the incidence of the pathogen has been reported in the Inner Mongolia Autonomous Region, Fujian Province, Xinjiang Uygur Autonomous Region (
Wen and Chen 2002;
Zhu et al. 2003;
Chen et al. 2004;
Liu et al. 2006;
Wang et al. 2006;
Xiao et al. 2011). Between 2005 and 2007, a total of 96 isolates were collected and investigated for their virulence pathotypes in Heilongjiang Province (
Zhang et al. 2010). Of the 8 races identified, 4 races had new pathotypes and some known races were not discovered, implying that
P. sojae population in Heilongjiang Province became more diverse during a decade. None of the
P. sojae isolates found in Heilongjiang Province was virulent to
Rps1c until race 4 and race 5 were found, which can overcome
Rps1c (
Zhang et al. 2010).
In Japan, the first PRR was reported in 1977 in Hokkaido (
Tsuchiya et al. 1990). The virulence of 49 Japanese isolates was determined into 10 pathotypes based on 6 Japanese differential cultivars (
Tsuchiya et al. 1990), which revealed distinguished pathotypes from the known 55 American
P. sojae races (
Grau 2004). This result indicates that genetic differences exist among American and Japanese isolates.
Sugimoto et al. (2010) identified that pathotype E is most dominant in
P. sojae populations in Japan from screening of 164 isolates collected between 2002 and 2006. Later, more than 100
P. sojae isolates identified from 14 regions were assessed using differential cultivars for 14
Rps genes (
Moriwaki 2010).
Rps1a, 1b, 1c, 1d, 1k, 3b, 7, and
8 were found to provide resistance against 47-81% of the assessed isolates.
Rps1d and
1k were the most effective resistance genes (
Moriwaki 2010).
In Brazil, the
P. sojae population presents a different origin. Of 37 isolates collected in six Brazilian states, 17 pathotypes were determined and, surprisingly, all the Brazilian pathotypes differ from previously known pathotypes (
Costamilan et al. 2013). It probably has been under different selection pressure by
Rps genes that unknowingly exist in Brazilian soybean cultivars (
Costamilan et al. 2013). The isolates in the Brazilian population were highly variable for their virulence to seven different
Rps genes of the 14
Rps. Consequently,
Rps1a, 1c, and
1k were highly recommended to utilize in breeding programs in Brazil, while
Rps1a and
1c are not effective in the United States (
Costamilan et al. 2013).
Soybean genotypes have different reactions to different isolates of
P. sojae (
Kaufmann and Gerdemann 1958). With regards to fungicide application to the seeds, genetic resistance of soybeans has taken the roles in the disease management. Mainly, two types of genetic resistance are described in soybean–
P. sojae system;
R-gene mediated resistance and partial resistance (
Schmitthenner 1985).
R-gene mediated resistance, conditioned by single dominant
Rps genes, generally provides race-specific and complete resistance. Exceptionally, the
Rps2 gene was known to provide root-specific and incomplete resistance that limits the colonization of infection on the soybeans (
Mideros et al. 2007). As a result,
Rps-mediated resistance has provided effective protection against the majority of
P. sojae populations in the United States (
Dorrance et al. 2018).
Table 1 summarizes all the
Rps alleles that were reported to date, their genomic positions, and flanking molecular markers used in the genetic mapping. The first
Rps gene was identified in the 1950s (
Bernard et al. 1957). More than 30
Rps alleles have been identified and mapped to nine chromosomes as follow:
Rps1a, 1b, 1c, 1d, 1k, 7, 9, UN1, YU25, YD29, HN, Q, WY, HC18, X, and
Waseshiroge on chromosome 3;
Rps3a, 3b, 3c, 8, and
SN10 on chromosome 13;
Rps4, 5, 6, 12, and
JS on chromosome 18;
Rps2 and
UN2 on chromosome 16;
Rps10 on chromosome 17;
Rps11 on chromosome 7;
RpsZS18 on chromosome 2;
RpsSu on chromosome 10; and
RpsYB30 on chromosome 19 (
Table 1) (
Demirbas et al. 2001;
Weng et al. 2001;
Burnham et al. 2003a;
Sandhu et al. 2004;
Gordon et al. 2007;
Sugimoto et al. 2007;
Yu et al. 2010;
Sugimoto et al. 2011;
Sun et al. 2011;
Lin et al. 2013;
Sun et al. 2014;
Cheng et al. 2017;
Li et al. 2017;
Niu et al. 2017;
Sahoo et al. 2017;
Zhong et al. 2017).
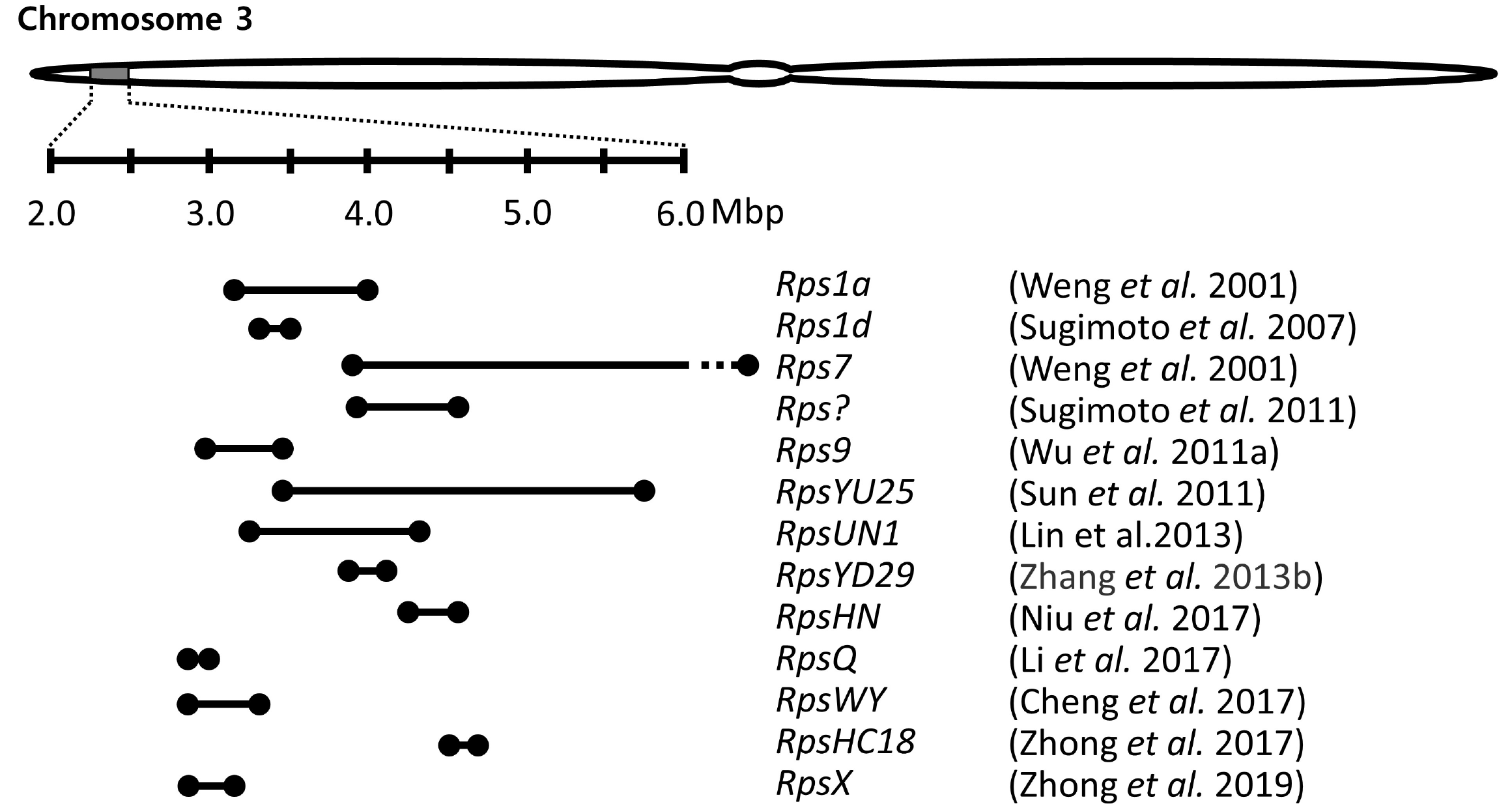
A few genomic regions have been repeatedly detected in multiple genetic mapping using bi-parent populations. Especially, a genomic region of ∼1.5 Mb on chromosome 3 is a hot spot, where resistance was identified in over ten previous studies using different resistance sources (
Fig. 1). Many copies of
R-gene type sequences were predicted in a recent genome annotation Wm82.a2.v1 (Glyma2.0) (available at
http://soybase.org).
Li et al. (2016) fine-mapped
RpsUN1 in a 151-kb interval on chromosome 3 and
RpsUN2 in a 36-kb interval on chromosome 16, where multiple copies of
R-gene type annotation are found. Relative abundance of transcripts of selected candidate genes for
RpsUN1 and
UN2 (
i.e. Glyma.03g034400 and Glyma.16g214900) also varied between the two parents (
i.e. Williams and PI 567139B). Nucleotide-binding site leucine-rich repeat (NBS-LRR) proteins are typically encoded by the majority of disease resistance genes in plants (
McHale et al. 2006). Thus, many studies highlighted the NBS-LRR containing genes in their candidate regions for
R genes.
Cheng et al. (2017) mapped
RpsWY in a 35 kb-region of chromosome 3 through a high-resolution genetic mapping with ∼200 F
7:8 recombinant inbred lines (RIL) and suggest four genes (Glyma03g04350, Glyma03g04360, Glyma03g04370, and Glyma03g04380) as candidates. Unlike
Li et al. (2016), non-NBS-LRR types of genes have also been proposed as candidates for another
Rps allele on chromosome 3 in this study (
Cheng et al. 2017). Of the published
Rps loci,
Rps1 was most characterized and known to contain a cluster of coiled-coil (CC)-NBS-LRR type
R genes (
Bhattacharyya et al. 2005). Four copies of closely related CC-NBS-LRR gene isolated from the
Rps1k genotype indicated that at least two of these copies (
Rps1k-1 and
Rps1k-2) function in steric resistance. Similarly, the
Rps4-Rps6 locus contains multiple copies of the NBS-LRR gene, and the deletion of the NBS-LRR sequence in
Rps4 locus was associated with loss of resistance to
P. sojae (
Sandhu et al. 2004). These studies imply that specificities of certain
Rps might be regulated by haplotypes or copy number variations, rather than single genes.
Rps-mediated resistance is known to follow gene-for-gene interaction (
Flor 1955). Individual isolates of
P. sojae may possess one or more avirulence (
Avr) genes corresponding to each of the existing
Rps genes of soybean in their genome (
Tyler 2007). Soybean
Rps genes are thought to activate an effector-triggered immune response, like the well-characterized
R-gene in other pathosystems (
Jones and Dangl 2006;
Dong et al. 2011). In
P. sojae-infected plants, the expression of defense responses was determined based on the interaction between the proteins encoded by the
Rps genes and the effector proteins encoded by the
Avr genes. Direct or indirect interaction between such resistance proteins and cognate
Avr proteins of pathogens leads to hypersensitive responses in the form of programmed cell death in infected cells; subsequently, the spread of the pathogen becomes limited locally (
Bent and Mackey 2007;
Tyler 2007).
Partial resistance to P. sojae in soybean
Partial resistance is an alternative form of genetic resistance that has been used in soybean fields against
P. sojae, which is quantitatively inherited by multiple genes, and effective against a wide range of
P. sojae races (
Schmitthenner 1985). It is also known as rate-limiting resistance, field tolerance, incomplete resistance, or non-race specific resistance (
Tooley and Grau 1984;
Walker and Schmitthenner 1984). This type of genetic resistance can be expressed in the form of reduced infection efficiency, reduced lesion size, and fewer oospores produced within the infected tissues (
Mideros et al. 2007). This resistance has been shown to reduce yield loss in the presence of
P. sojae, but not to negatively impact on yield even in the absence of the disease (
St. Martin et al. 1994).
Partial resistance had moderate to high heritability, thus can be improved through the selection of resistance. Typically, the levels of partial resistance are determined using lesion length measurement, tray test, root rot score, inoculum layer test, or field evaluation (
Tooley and Grau 1984;
Dorrance et al. 2008). The soybean cultivar ‘Conrad’ was extensively studied for the resistance through the quantitative trait locus (QTL) analysis with several biparental populations, because it does not have
Rps genes but exhibits high partial resistance (
Burnham et al. 2003b;
Han et al. 2008;
Li et al. 2010;
Wang et al. 2010;
Stasko et al. 2016). Some common QTLs were identified against three isolates of
P. sojae in a ‘Conrad’ × ‘Sloan’ population, indicating that common defense responses may occur in responsive to the individual isolates inoculated (
Stasko et al. 2016). Two loci on chromosome 19 were highlighted in
Wang et al. (2010) and
Stasko et al. (2016), which were highly significant and effective to multiple isolates.
Although the mechanisms of partial resistance are largely unknown, there is evidence that several studies provided evidences to support different mechanisms for partial resistance. Much higher levels of suberin were formed in the roots of Conrad and growth of hyphae of
P. sojae was retarded in Conrad in comparison of a susceptible line, suggesting that colonization of
P. sojae may be suppressed by pre-formed suberin (
Thomas et al. 2007;
Ranathunge et al. 2008). Expression of genes underlying the identified QTL revealed that signal transduction, hormone-associated defense pathway, and defeated Rps genes might be associated with high partial resistance in Conrad (
Wang et al. 2012). High levels of sequence variation were identified in the upstream, intron, and downstream regulatory regions of the genes within the QTL on chromosome 19 (
Wang et al. 2012). In two plant introductions (PIs) 398841 and 407861A, with high levels of partial resistance to
P. sojae, a few of QTL for the partial resistance were mapped near known
Rps loci; for instance, a region of
Rps1 and
Rps7 on chromosome 3 and a region of
Rps3 and
Rps8 on chromosome 13 (
Lee et al. 2013a,
2013b). These results support that weak or defeated form of
Rps genes may contribute to high levels of partial resistance to
P. sojae because the two PIs did not have any
Rps gene effective to the inoculated isolates.
Kang et al. (2019) reported the presence or absence of
Rps-mediated resistance in major soybean cultivars grown in S. Korea against 4 isolates, where the resistance reactions were observed only in ∼5 of the accessed 21 cultivars per isolate. Ten of the assessed cultivars were susceptible to all the inoculated isolates, which implies that major soybean cultivars are potentially under threat of this pathogen (
Kang et al. 2019). Genetic resistance is a desirable and effective method in crop protection from various biotic stresses; thus, it is important for researchers/breeders to research soybean–
P. sojae interaction via various approaches for successes in disease management of PRR.
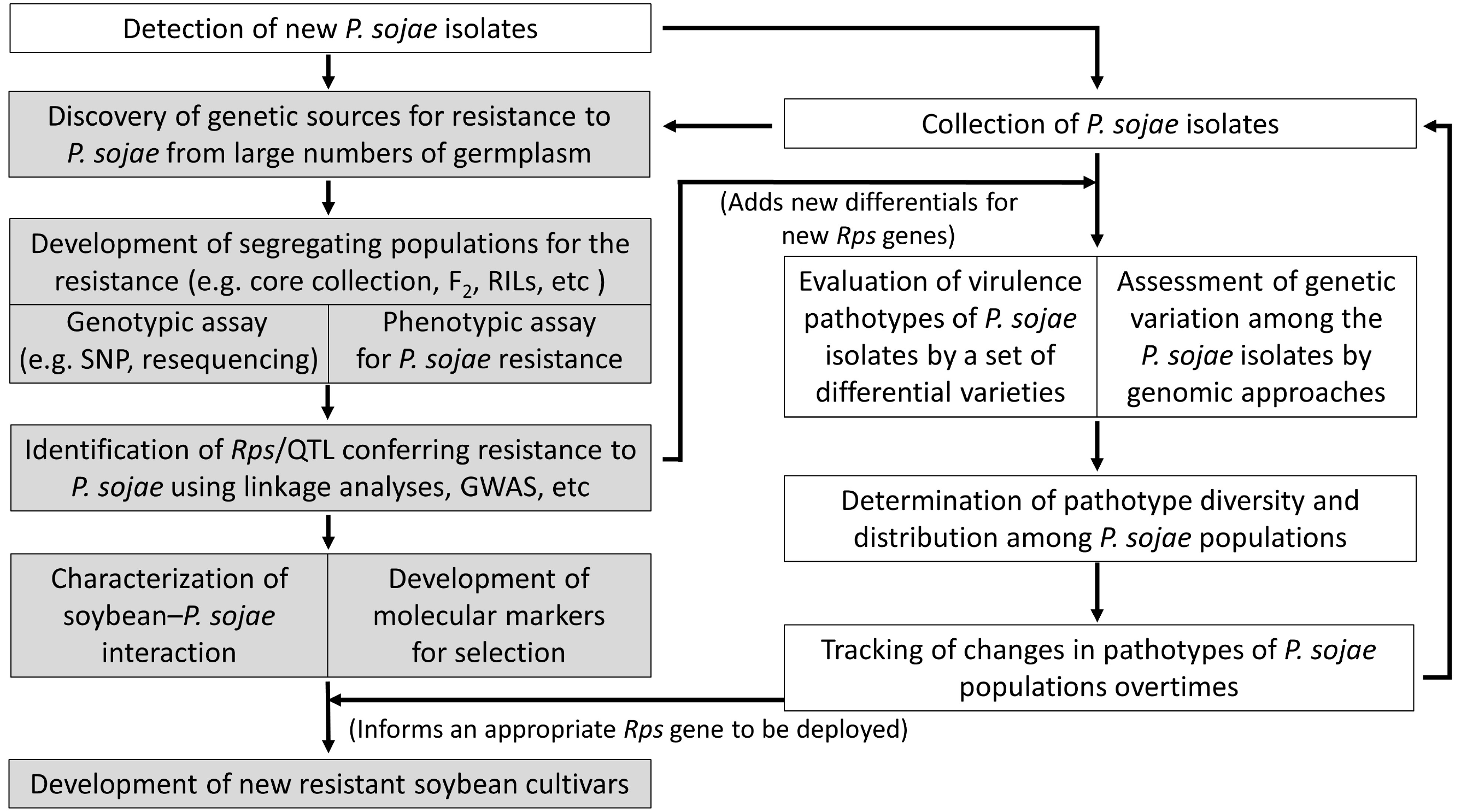
Fig. 2 highlights collaborative future researches to support better protection from PRR, which consists of two parts, i.e. soybean genetics/breedng and pathology.
Genetic diversity provides an essential key in plant breeding (
Carter et al. 2004). A vast number of soybean germplasm are available in South Korea, as an origin of soybean, and a wide range of genetic diversity exists among them (
Jeong et al. 2019). The germplasm have not been evaluated for
Rps or partial resistance to
P. sojae to date. Thus, it should be the first priority to identify soybean germplasm from continuous phenotypic screenings against many
P. sojae isolates. A core collection of Korean cultivated soybean germplasm was recently defined and used to evaluate phenotypic variations and to dissect genetic architectures of multiple traits such as seed composition, seed size/color, and agronomic traits (
Jeong et al. 2019). Since the core collection represents genetic diversity with minimal repeatability across the Korean soybean germplasm collection, it will be reasonable to conduct initial phenotypic screening with this representative of Korean cultivated soybean germplasm. Subsequently, the discovered resistant germplasm will need to be characterized to identify
R-genes/QTL associated with resistance to
P. sojae. This process can be conducted using a collection of hundreds of germplasm or bi-parental populations developed from segregating parents for the target traits. In addition to the core collection, the Korean soybean nested association mapping (NAM) population, which consisted of 27 F
6-derived RIL population, is another valuable resource for dissecting genetic architecture of complex traits. These plant materials were genotyped with hundred thousands of single nucleotide polymorphism (SNP) markers, which will facilitate high resolution of genetic analyses after disease phenotypes are obtained. Identificaiton of resistance genes/QTL will be followed by further omic-based characterization for soybean-
P. sojae molecular interactions as genetic research purposes. As well, fine mapping of the target gene and marker development for selection will lead to development of new
Phytophthora-resistant cultivars.
Deploying
Rps genes is a primary way to genetically improve existing susceptible varieties in a viewpoint of soybean breeding. Intense use of a few resistance varieties, however, has led selective pressures on the pathogen for changes in its ability to overcome the defenses (
Abney et al. 1997). The utilization of various
Rps genes will benefit to attenuate selective pressure on
P. sojae populations caused by respective deployed
Rps genes; consequently, increased occurrence of new virulence pathotypes is possibly delayed (
Schmitthenner 1985). The selection of partial resistance combined with a few
Rps genes is also recommended for the long-term management of PRR by avoiding boom and bust cycles in a single
Rps locus (
Buzzell and Anderson 1992). A recent study demonstrated genetic gains from selections for a major QTL for partial resistance to
P. sojae (
Karhoff et al. 2019). The introgressions of a resistance allele from the respective PI 427105B and PI 427106 improved the levels of resistance to
P. sojae by ∼20% and ∼40%, respectively, and the yield by 13-29% under diseased condition (
Karhoff et al. 2019).
On the other hand, understanding of the pathogen contributing valuable information for disease management because the pathogen also co-evolves as plant defense responses develop, a.k.a an evolutionary arm-race (
Anderson et al. 2010).
Kang et al. (2019) reported that although a small number of isolates were tested, the variability of virulence pathotypes also existed among the isolates, suggesting that dominant virulence pathotypes may differ across geographical regions. Therefore, it is highly requried to continue to collect
P. sojae isolates as many as possible from fields, and to evaluate them with the designated differential varieties. Monitoring the changes in pathotypes among
P. sojae populations overtimes will be a greatly valuable study because pathotype changes in
P. sojae populations have been one of the significant hurdles in combating against
P. sojae in several countries as stated above. Such tracking research will also assist to select a few effective
Rps genes based on the dominant pathotypes of
P. sojae observed in soybean fields. For this, at the same time, the addition of new differentials is strongly recommended as new
Rps genes were discovered. The differential varieties for 14
Rps genes, developed in the United States, have been used to characterize
P. sojae isolates in the majority of recent studies. Some recent studies have identified novel
Rps genes (
i.e. Rps10 and
Rps11) in genomic locations distinct from the previously known
Rps loci (
Table 1). These additional differentials will improve the ability of diagnoses for the variability of pathotypes in
P. sojae populations for current and future characterization.
CONCLUDING REMARKS
The study on the interaction between soybean and soil-borne oomycete pathogen P. sojae has relatively long histories in the United States, China, Japan, and other countries. This review provided a general introduction to PRR and its causal agent and summarized the results of published researches regarding Rps genes and QTL analysis to understand the resistance and pathotype diversity of P. sojae population. It is evident that PRR and other possible soil-borne diseases threaten the soybean production in South Korea. In this circumstance, proposed genetic researches should be conducted to provide a framework for effective management of newly-emerging diseases and to develop disease-resistant soybean varieties, which will be helpful to reduce yield losses caused by P. sojae in the future.
ACKNOWLEDGEMENTS
This research was funded by a grant from Chungnam National University.
-
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
Fig. 1A genomic region on chromosome 3 where several resistance genes to Phytophthora sojae were mapped. Lines between the two black points present the marker interval of each Rps gene/allele as reported in the publications. The dashed part of the interval presents outside of 2.0-6.0 Mbp.
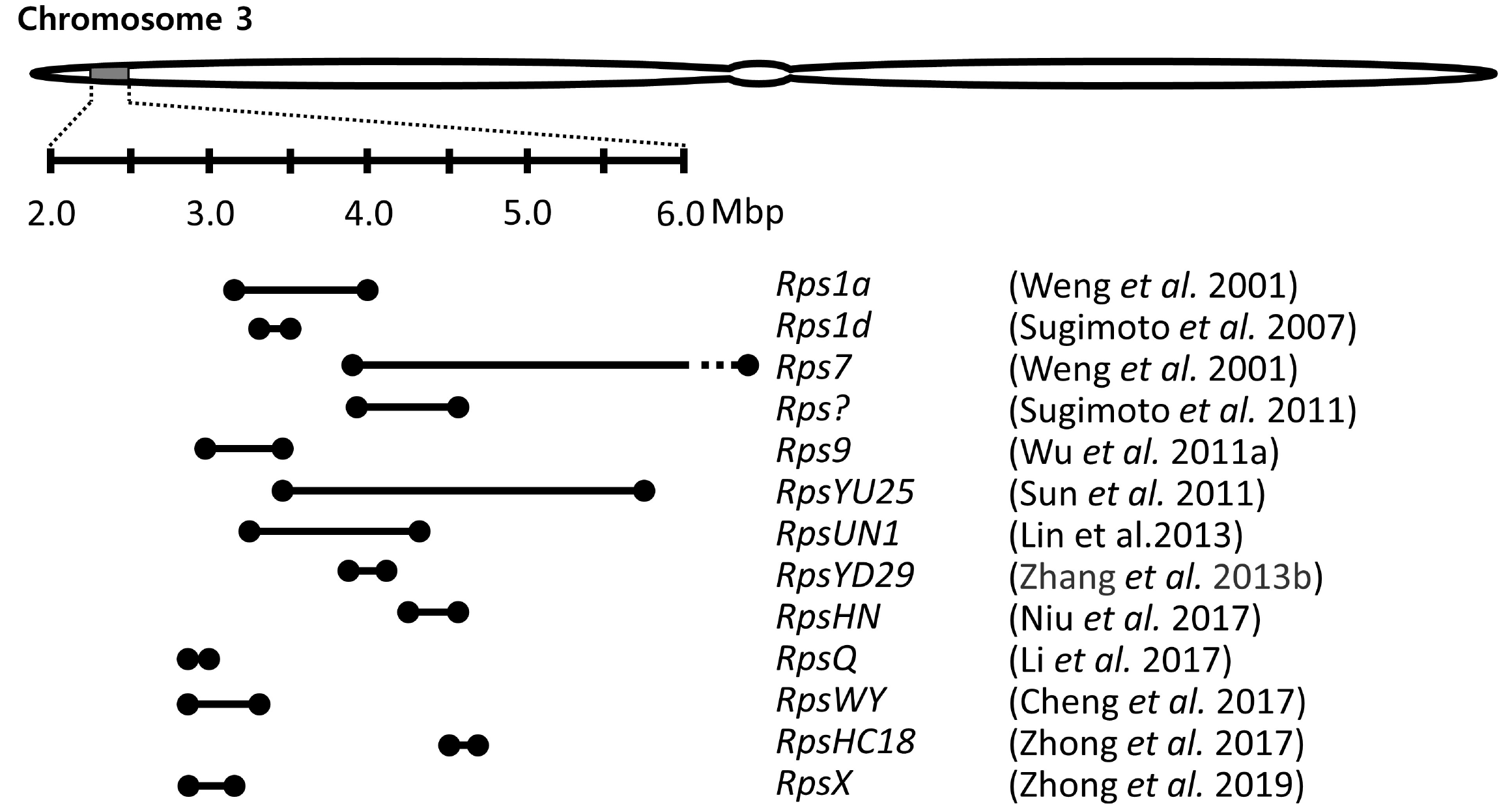
Fig. 2Schematic diagram of interdisciplinary researches suggested for soybean resistance to Phytophthora sojae in South Korea.
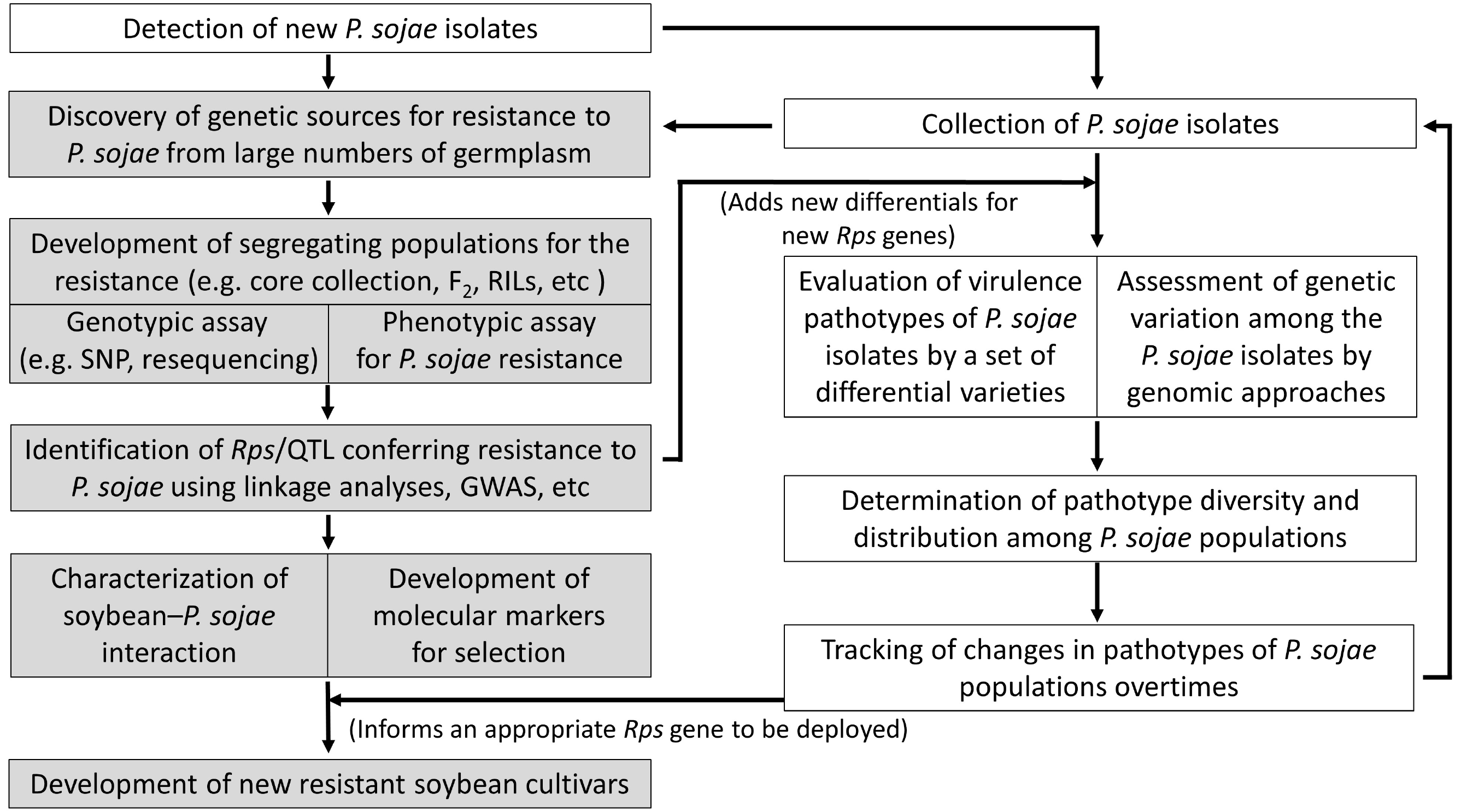
Table 1A list of Rps genes/alleles and associated SNP/SSR markers reported in the previous studies.
Table 1
|
Chr.z) (LG) |
Rpsy)
|
Position1 (Mbp)x)
|
Position2 (Mbp)x)
|
Flanking marker 1y)
|
Flanking marker 2y)
|
Reference |
|
1 (D1a) |
- |
44.4 |
44.6 |
ss715579469 |
ss715579474 |
, Niu et al. 2018
|
|
2 (D1b) |
RpsZS18
|
46.6*
|
44.3 |
Sat_069 (BARCSOYSSR_02_1540) |
Sat_183 (BARCSOYSSR_02_1602) |
, Yao et al. 2010
|
|
3 (N) |
Rps1a
|
3.2 |
3.9 |
Satt159 (BARCSOYSSR_03_0180) |
Satt009 (BARCSOYSSR_03_0226) |
, Weng et al. 2001
|
|
3 (N) |
Rps1b
|
3.4 |
5.7 |
Satt152 (BARCSOYSSR_03_0192) |
Satt530 |
, Demirbas et al. 2001
|
|
3 (N) |
Rps1c
|
3.4 |
9.2 |
Satt152 (BARCSOYSSR_03_0192) |
Satt584 (BARCSOYSSR_03_0442) |
, Demirbas et al. 2001
|
|
3 (N) |
Rps1d
|
3.4 |
3.5*
|
Satt152 (BARCSOYSSR_03_0192) |
Sat_186 (BARCSOYSSR_03_0204) |
, Sugimoto et al. 2007
|
|
3 (N) |
Rps7
|
3.9 |
18.4 |
Satt009 (BARCSOYSSR_03_0226) |
Satt125 (BARCSOYSSR_03_0564) |
, Weng et al. 2001
|
|
3 (N) |
Rps9
|
2.9 |
3.5*
|
Satt631 (BARCSOYSSR_03_0162) |
Sat_186 (BARCSOYSSR_03_0204) |
, Wu et al. 2011a
|
|
3 (N) |
RpsUN1
|
3.2 |
4.3 |
Satt159 (BARCSOYSSR_03_0180) |
BARCSOYSSR_03_0250 |
, Lin et al. 2013
|
|
3 (N) |
RpsYU25
|
3.5*
|
5.7 |
Sat_186 (BARCSOYSSR_03_0204) |
Satt530 |
, Sun et al. 2011
|
|
3 (N) |
RpsYD29
|
3.9 |
4.1 |
SattWM82-50 |
Satt1k4b |
, Zhang et al. 2013b
|
|
3 (N) |
RpsHN
|
4.2 |
4.5 |
SSRSOYN-25 |
SSRSOYN-44 |
, Niu et al. 2017
|
|
3 (N) |
RpsQ
|
3.0 |
3.1 |
BARCSOYSSR_03_0165 |
InDel281 |
, Li et al. 2017
|
|
3 (N) |
- |
3.9 |
4.5 |
Satt009 (BARCSOYSSR_03_0226) |
T0003044871 |
, Sugimoto et al. 2011
|
|
3 (N) |
RpsWY
|
2.9 |
3.4 |
Satt631 (BARCSOYSSR_03_0162) |
Satt152 (BARCSOYSSR_03_0192) |
, Cheng et al. 2017
|
|
3 (N) |
RpsHC18
|
4.5 |
4.6 |
BARCSOYSSR_03_0269 |
BARCSOYSSR_03_0272 |
, Zhong et al. 2017
|
|
3 (N) |
RpsX
|
2.9 |
3.2 |
InDelxz6 |
BARCSOYSSR_03_0175 |
, Zhong et al. 2019
|
|
7 (M) |
Rps11
|
5.1 |
5.2 |
BARCSOYSSR_07_0266 |
BARCSOYSSR_07_0278 |
, Ping et al. 2015
|
|
10 (O) |
RpsSu
|
1.0 |
39.4 |
Satt358 |
Sat_242 (BARCSOYSSR_10_1104) |
, Wu et al. 2011b
|
|
13 (F) |
Rps3
|
31.8 |
32.7 |
Satt510 (BARCSOYSSR_13_1219) |
Satt335 (BARCSOYSSR_13_1271) |
, Gordon et al. 2007
|
|
13 (F) |
Rps8
|
24.3 |
28.9 |
Satt425 (BARCSOYSSR_13_0784) |
Satt114 (BARCSOYSSR_13_1055) |
, Gordon et al. 2006
|
|
13 (F) |
RpsSN10
|
16.6 |
16.9 |
Satt423 (BARCSOYSSR_13_0264) |
Satt149 (BARCSOYSSR_13_0245) |
, Yu et al. 2010
|
|
16 (J) |
Rps2
|
1.6 |
34 |
Satt287 (BARCSOYSSR_16_0090) |
Satt547 (BARCSOYSSR_16_1165) |
, Demirbas et al. 2001
|
|
16 (J) |
RpsUN2
|
36.9 |
37.3 |
BARCSOYSSR_16_1275 |
Sat_144 (BARCSOYSSR_16_1294) |
, Lin et al. 2013
|
|
16 (J) |
-
|
4.0*
|
- |
BARC-014467-01559 |
- |
, Huang et al. 2016
|
|
17 (D2) |
Rps10
|
30.8 |
31.1 |
Sattwd15-24 |
Sattwd15-47 |
, Zhang et al. 2013a
|
|
18 (G) |
Rps4
|
54.5 |
56.3 |
Satt191 (BARCSOYSSR_18_1750) |
Sat_064 (BARCSOYSSR_18_1858) |
, Sandhu et al. 2004
|
|
18 (G) |
Rps5
|
- |
53.9 |
- |
Satt472 (BARCSOYSSR_18_1708) |
, Sahoo et al. 2017
|
|
18 (G) |
Rps6
|
54.5 |
- |
Satt191 (BARCSOYSSR_18_1750) |
Sat_372 |
, Gordon et al. 2007
|
|
18 (G) |
Rps12
|
56 |
56.3 |
BARCSOYSSR_18_1840 |
Sat_064 (BARCSOYSSR_18_1858) |
, Sahoo et al. 2017
|
|
18 (G) |
RpsJS
|
56.3 |
56.6 |
BARCSOYSSR_18_1859 |
BARCSOYSSR_18_1864 |
, Sun et al. 2014
|
|
19 (L) |
RpsYB30
|
33.9 |
34.8 |
Satt497 (BARCSOYSSR_19_0760) |
Satt313 (BARCSOYSSR_19_0788) |
, Zhu et al. 2007
|
|
20 (I) |
-
|
46.6*
|
- |
BARC-013645-01207 |
- |
, Huang et al. 2016
|
Table 2A list of QTL for partial resistance to Phytophthora sojae in the previous studies.
Table 2
|
Chr. (LG)z)
|
Position1 (bp)y,x)
|
Position2 (bp)y,x)
|
Flanking marker1 |
Flanking marker2 |
PVE (%)w)
|
Plant materials (Mapping population) |
Reference |
|
1 (D1a) |
50164447 |
50295635 |
BARC_2.0_Gm01_50164447 |
BARC_2.0_Gm01_50295635 |
4.5 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
1 (D1a) |
50206347 |
50287274 |
BARC_2.0_Gm01_50206347 |
BARC_2.0_Gm01_50287274 |
8.2 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
1 (D1a) |
50560774 |
51398886*
|
BARCSOYSSR_01_1400 |
BARC-020113-04470 |
4.6 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
1 (D1a) |
50560774 |
50157973*
|
BARCSOYSSR_01_1400 |
BARC-054071-12319 |
5.6 |
Combined population |
, Lee et al. 2014
|
|
1 (D1a) |
50572171 |
50797061 |
BARC_2.0_Gm01_50572171 |
BARC_2.0_Gm01_50797061 |
7.6 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
2 (D1b) |
4549894*
|
5455404*
|
BARC-065787-19749 |
BARC-056237-14178 |
10.7-11.4 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
2 (D1b) |
653563*
|
4901498*
|
BARC-041773-08087 |
BARC-016573-02145 |
2.3 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
2 (D1b) |
14288241 |
19688108 |
Satt266 (BARCSOYSSR_02_0727) |
Satt579 (BARCSOYSSR_02_0855) |
15.9 |
Conrad × Harosoy |
, Burnham et al. 2003b
|
|
2 (D1b) |
19688108 |
29355267 |
Satt579 (BARCSOYSSR_02_0855) |
Satt600 (BARCSOYSSR_02_1048) |
10.6 |
Conrad × Sloan |
, Burnham et al. 2003b
|
|
2 (D1b) |
19688108 |
29355267 |
Satt579 (BARCSOYSSR_02_0855) |
Satt600 (BARCSOYSSR_02_1048) |
20.7 |
Conrad × Williams |
, Burnham et al. 2003b
|
|
2 (D1b) |
19688108 |
34875449 |
Satt579 (BARCSOYSSR_02_0855) |
Sat_089 (BARCSOYSSR_02_1152) |
5-28 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
2 (D1b) |
27699285 |
29355267 |
Satt005 (BARCSOYSSR_02_0998) |
Satt600 (BARCSOYSSR_02_1048) |
11-22 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
2 (D1b) |
- |
45267040 |
OPL18800
|
Satt274 (BARCSOYSSR_02_1663) |
2-10 |
Conrad × OX760-6-1 |
, Han et al. 2008
|
|
2 (D1b) |
50529114*
|
51243188*
|
BARC-042881-08448 |
BARC-019805-04379 |
12.1-14.6 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
PI399036 × AR3 |
|
2 (D1b) |
51243188*
|
51550018*
|
BARC-019805-04379 |
BARC-906743-01012 |
5.6 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
3 (N) |
2993784*
|
- |
BARC-028645-05979 |
BARC-010837-00763 |
2.8 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
3 (N) |
2993784*
|
5656713 |
BARC-028645-05979 |
BARCSOYSSR_03_0317 |
3.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
3 (N) |
3225968*
|
- |
Gm03_3225968 |
- |
21.1 |
Germplasm (n=169) |
, Ludke et al. 2019
|
|
3 (N) |
3852827 |
- |
ss715585712 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
3 (N) |
3865669 |
- |
ss715585728 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
3 (N) |
4276473 |
- |
ss715586320 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
3 (N) |
4277319 |
- |
ss715586321 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
3 (N) |
4315451 |
- |
ss715586376 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
3 (N) |
5217414*
|
- |
Gm03_5217414 |
- |
13.9 |
Germplasm (n=169) |
, Ludke et al. 2019
|
|
3 (N) |
37047526*
|
38032013*
|
BARC-050433-09624 |
BARC-010179-00543 |
6.7-6.9 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
3 (N) |
38834669 |
44771010*
|
Sat_091 (BARCSOYSSR_03_1348) |
Sat_125 (BARCSOYSSR_03_1546) |
4.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
3 (N) |
41065116 |
41146728*
|
Sat_241 (BARCSOYSSR_03_1469) |
BARC-020101-04452 |
3.9 |
Combined population |
, Lee et al. 2014
|
|
4 (C1) |
57542*
|
3930569*
|
BARC-038359-10052 |
BARC-021219-04011 |
2.5 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
4 (C1) |
57542*
|
2069791*
|
BARC-038359-10052 |
BARC-054289-12451 |
2.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
4 (C1) |
8061903*
|
38604410*
|
BARC-024445-04886 |
BARC-061079-17031 |
2.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
4 (C1) |
38604410*
|
43684322*
|
BARC-061079-17031 |
BARC-042189-08197 |
1.9 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
4 (C1) |
45977762 |
46204517 |
BARC_2.0_Gm04_45977762 |
BARC_2.0_Gm04_46204517 |
3.2 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
4 (C1) |
46096228 |
46536196 |
BARC_2.0_Gm04_46096228 |
BARC_2.0_Gm04_46536196 |
3.2 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
5 (A1) |
33679928*
|
34294649*
|
BARC-031361-07059 |
BARC-018011-02495 |
7.9 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
6 (C2) |
17218677 |
- |
Satt277 (BARCSOYSSR_06_0920) |
Satt365 |
9-21 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
6 (C2) |
23848501 |
31490622 |
Satt489 (BARCSOYSSR_06_1129) |
Satt100 (BARCSOYSSR_06_1202) |
5-21 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
6 (C2) |
44049891 |
46820673 |
Satt460 (BARCSOYSSR_06_1456) |
Satt307 (BARCSOYSSR_06_1581) |
4-7 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
6 (C2) |
7023397*
|
20218893 |
Satt520 (BARCSOYSSR_06_0386) |
Satt557 (BARCSOYSSR_06_1041) |
4.3 |
Su88-M21 × Xinyixiaoheidou |
, Wu et al. 2011c
|
|
6 (C2) |
21775764*
|
46063373*
|
BARC-040475-07751 |
BARC-062515-17881 |
4.7-5.5 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
7 (M) |
2471949*
|
6404814*
|
BARC-029825-06442 |
BARC-042815-08424 |
11.6-12.4 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
7 (M) |
5899427*
|
14931953*
|
BARC-028455-05917 |
BARC-065353-19384 |
2.4 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
8 (A2) |
17232172 |
- |
Satt233 (BARCSOYSSR_08_0960) |
Satt437 |
4-17 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
8 (A2) |
9492376*
|
21435976*
|
BARC-057257-14650 |
BARC-060405-16674 |
14.5-15.9 |
PI 399036 × AR3 |
, Abeysekara et al. 2016
|
|
8 (A2) |
24848378*
|
44053323*
|
BARC-051883-11286 |
BARC-042715-08379 |
7.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
9 (K) |
15487393 |
- |
BARC_2.0_Gm09_15487393 |
- |
4.5-7.4 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
9 (K) |
1280743*
|
2330196*
|
BARC-007972-00189 |
BARC-051275-11075 |
7.1 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
9 (K) |
37144807*
|
38300154*
|
BARC-017625-02635 |
BARC-055533-13402 |
29.9 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
9 (K) |
38300154*
|
38383165*
|
BARC-055533-13402 |
BARC-007999-00186 |
21 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
10 (O) |
10091607 |
43788256 |
Satt420 (BARCSOYSSR_10_0507) |
Sat_274 (BARCSOYSSR_10_1353) |
7.7 |
Su88-M21 × Xinyixiaoheidou |
, Wu et al. 2011c
|
|
10 (O) |
39359642*
|
44753498*
|
BARC-060257-16508 |
BARC-015925-02017 |
4.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
11 (B1) |
34173104 |
- |
Satt453 (BARCSOYSSR_11_1468) |
Satt484 |
5-14 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
12 (H) |
1687387 |
- |
Satt353 (BARCSOYSSR_12_0068) |
OSU31 |
4.6 |
Conrad × Sloan |
, Wang et al. 2010
|
|
12 (H) |
7494659*
|
9770096*
|
BARC-019775-04370 |
BARC-025943-05179 |
5.7 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
12 (H) |
14578917*
|
33153095*
|
BARC-061985-17608 |
BARC-018895-03034 |
4.0 |
Combined population |
, Lee et al. 2014
|
|
13 (F) |
6216988 |
13134055 |
Satt509 (BARCSOYSSR_11_0342) |
Satt030 (BARCSOYSSR_13_0445) |
6-13 |
Conrad × OX760-6-1 |
, Han et al. 2008
|
|
13 (F) |
8587948 |
10392903 |
Satt325 (BARCSOYSSR_13_0639) |
Satt343 (BARCSOYSSR_13_0518) |
9-10 |
Conrad × Hefeng25 |
, Li et al. 2010
|
|
13 (F) |
10392903 |
- |
Satt343 (BARCSOYSSR_13_0518) |
OPG16600 |
2-8 |
Conrad × OX760-6-1 |
, Han et al. 2008
|
|
13 (F) |
16454986 |
16855019 |
Satt252 (BARCSOYSSR_13_0272) |
Satt149 (BARCSOYSSR_13_0245) |
32.4 |
Conrad × Sloan |
, Burnham et al. 2003b
|
|
13 (F) |
16454986 |
16600399 |
Satt252 (BARCSOYSSR_13_0272) |
Satt423 (BARCSOYSSR_13_0264) |
35.0 |
Conrad × Harosoy |
, Burnham et al. 2003b
|
|
13 (F) |
16454986 |
16855019 |
Satt252 (BARCSOYSSR_13_0272) |
Satt149 (BARCSOYSSR_13_0245) |
21.4 |
Conrad × Williams |
, Burnham et al. 2003b
|
|
13 (F) |
16855019 |
17875691 |
Satt149 (BARCSOYSSR_13_0245) |
Satt160 (BARCSOYSSR_13_0196) |
2.0 |
Conrad × Sloan |
, Wang et al. 2010
|
|
13 (F) |
27656895 |
30236183 |
Sat_234 (BARCSOYSSR_13_0981) |
BARCSOYSSR_13_1131 |
16.1 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
13 (F) |
27656895 |
30236183 |
Sat_234 (BARCSOYSSR_13_0981) |
BARCSOYSSR_13_1131 |
8.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
13 (F) |
28506083 |
35072147 |
Sat_154 (BARCSOYSSR_13_1029) |
Sat_375(BARCSOYSSR_13_1385) |
20.1-35.8 |
S99-2281 × PI408105A |
, Nguyen et al. 2012
|
|
13 (F) |
28912864 |
31802559 |
Satt114 (BARCSOYSSR_13_1055) |
Satt510 (BARCSOYSSR_13_1219) |
7.0 |
V71-370 × PI407162 |
, Tucker et al. 2010
|
|
13 (F) |
29685828 |
29647017 |
BARCSOYSSR13_1106 |
BARCSOYSSR13_1103 |
10.6 |
Combined population |
, Lee et al. 2014
|
|
13 (F) |
30739608 |
30739608*
|
Sct_033 (BARCSOYSSR_13_1230) |
Sct_033 (BARCSOYSSR_13_1230) |
5.4 |
Conrad × Sloan |
, Wang et al. 2010
|
|
13 (F) |
30765997 |
- |
ss715615031 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
13 (F) |
38566546*
|
- |
BARC-061571-17276 |
BARC-013325-00484 |
5.7 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
14 (B2) |
7132730*
|
8339923*
|
BARC-050249-09527 |
BARC-064873-18956 |
5.9 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
14 (B2) |
8642719*
|
13086766 |
Satt416 (BARCSOYSSR_14_0485) |
Satt304 (BARCSOYSSR_14_0646) |
4.7 |
Conrad × Sloan |
, Wang et al. 2010
|
|
15 (E) |
2740854*
|
2893732*
|
BARC-055329-13210 |
BARC-062899-18147 |
4.6 |
PI399036 × AR3 |
, Abeysekara et al. 2016
|
|
15 (E) |
2740854*
|
3581354 |
BARC-055329-13210 |
BARCSOYSSR_15_0160 |
7.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
15 (E) |
3639988 |
3591774 |
BARC_2.0_Gm15_3639988 |
BARC_2.0_Gm15_3591774 |
2.0 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
15 (E) |
4415019*
|
5522858*
|
BARC-054257-12408 |
BARC-028907-06042 |
6.7-7.3 |
PI399036 × AR2 |
, Abeysekara et al. 2016
|
|
15 (E) |
6823519 |
13653981 |
Satt651 (BARCSOYSSR_15_0306) |
Satt598 (BARCSOYSSR_15_0645) |
15.9 |
Su88-M21 × Xinyixiaoheidou |
, Wu et al. 2011
|
|
15 (E) |
1541381*
|
5522858*
|
BARC-018923-03037 |
BARC-028907-06042 |
2.6 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
15 (E) |
18422604*
|
- |
Gm15_18422604 |
- |
18.4 |
Germplasm (n=169) |
, Ludke et al. 2019
|
|
16 (J) |
486741 |
807114 |
BARC_2.0_Gm16_486741 |
BARC_2.0_Gm16_807114 |
5.1 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
16 (J) |
3124736 |
3362395 |
BARC_2.0_Gm16_3124736 |
BARC_2.0_Gm16_3362395 |
3.5-5.4 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
16 (J) |
8909747 |
23417256 |
Satt414 |
Satt529 (BARCSOYSSR_16_0703) |
22-32 |
V71-370 × PI407162 |
, Tucker et al. 2010
|
|
16 (J) |
8909747 |
14155157 |
Satt414 |
Satt596 |
13-21 |
Conrad × OX760-6-1 |
, Weng et al. 2007
|
|
16 (J) |
36544070*
|
36732606*
|
BARC-011625-00310 |
BARC-048135-10500 |
3.9 |
Combined population |
, Lee et al. 2014
|
|
17 (D2) |
7756014*
|
8360833*
|
BARC-058841-15463 |
BARC-052295-11407 |
7.6-12.7 |
PI 399036 × AR3 |
, Abeysekara et al. 2016
|
|
17 (D2) |
18425834 |
31915278*
|
Satt514 (BARCSOYSSR_17_0930) |
Satt574 (BARCSOYSSR_17_1164) |
7.1 |
Conrad × Sloan |
, Wang et al. 2010
|
|
17 (D2) |
20378955 |
20352435*
|
Sat_300 (BARCSOYSSR_17_0988) |
BARC-023721-03465 |
7.5-8.8 |
S99-2281 × PI408105A |
, Nguyen et al. 2012
|
|
18 (G) |
2296656*
|
2833147*
|
BARC-025777-05064 |
BARC-047665-10370 |
19.4 |
Combined population |
, Lee et al. 2014
|
|
18 (G) |
2419864 |
- |
Sat_163 (BARCSOYSSR_18_0143) |
SLP142 |
10-11 |
V71-370 × PI407162 |
, Tucker et al. 2010
|
|
18 (G) |
56710850 |
56876857 |
BARC_2.0_Gm18_56710850 |
BARC_2.0_Gm18_56876857 |
13.6 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
18 (G) |
56710850 |
56766936 |
BARC_2.0_Gm18_56710850 |
BARC_2.0_Gm18_56766936 |
5.3 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
18 (G) |
54810869*
|
57797198*
|
BARC-040163-07672 |
BARC-041331-07965 |
3.0 |
OX20-8 × PI407861A |
, Lee et al. 2013b
|
|
18 (G) |
57797198*
|
60488428*
|
BARC-041331-07965 |
BARC-044363-08678 |
3.6 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
|
19 (L) |
42821735*
|
46116996 |
BARC-047496-12943 |
BARC_2.0_Gm19_46116996 |
4.6 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
19 (L) |
43036918 |
- |
Satt527 (BARCSOYSSR_19_1214) |
OSU10 |
4.6 |
Conrad × Sloan |
, Wang et al. 2010
|
|
19 (L) |
43533689 |
44370710 |
BARCSOYSSR_19_1243 |
BARCSOYSSR_19_1286 |
3.1 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
19 (L) |
44370710 |
46116996 |
BARCSOYSSR_19_1286 |
ss715635553 |
9.1 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
19 (L) |
47528116 |
47787869 |
BARCSOYSSR_19_1452 |
Glyma.19G226100 |
4.1-7.8 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
19 (L) |
47114567*
|
48609875*
|
BARC-064609-18739 |
BARC-039977-07624 |
8.5-8.9 |
PI399036 × AR2 |
, Abeysekara et al. 2016 |
|
19 (L) |
49121197 |
- |
ss715635897 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
49461521 |
- |
ss715635934 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50305134 |
50222676*
|
BARC_2.0_Gm19_50305134 |
BARC-014385-01342 |
6.6 |
Conrad × Sloan |
, Stasko et al. 2016
|
|
19 (L) |
50544302 |
- |
ss715636056 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50555372 |
- |
ss715636059 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50604872 |
- |
ss715636064 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50663405 |
- |
ss715636073 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50666502 |
- |
ss715636076 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50668601 |
- |
ss715636077 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50680353 |
- |
ss715636083 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
19 (L) |
50681202 |
- |
ss715636084 |
- |
- |
Germplasm (n=797) |
, Schneider et al. 2016
|
|
20 (I) |
343168*
|
1747664*
|
BARC-042281-08231 |
BARC-057033-14543 |
6.8-7.4 |
PI 399036 × AR3 |
, Abeysekara et al. 2016
|
|
20 (I) |
25275083 |
- |
Satt239 (BARCSOYSSR_20_0543) |
Sat_105 |
7-12 |
V71-370 × PI407162 |
, Tucker et al. 2010
|
|
20 (I) |
26484795*
|
35176184*
|
BARC-046570-12662 |
Sat_268 (BARCSOYSSR_20_0855) |
1.9 |
OX20-8 × PI398841 |
, Lee et al. 2013a
|
References
- Abeysekara NS, Matthiesen RL, Cianzio SR, Bhattacharyya MK, Robertson AE. 2016. Novel sources of partial resistance against Phytophthora sojae in soybean PI 399036. Crop Sci.. 56: 2322-2335.
- Abney T, Melgar J, Richards T, Scott D, Grogan J, Young J. 1997. New races of Phytophthora sojae with Rps 1-d virulence. Plant Dis.. 81: 653-655.
- Allen TW, Bradley CA, Sisson AJ, Byamukama E, Chilvers MI, Coker CM, et al. 2017. Soybean yield loss estimates due to diseases in the United States and Ontario, Canada, from 2010 to 2014. Plant Health Prog.. 18: 19-27.
- Anderson JP, Gleason CA, Foley RC, Thrall PH, Burdon JB, Singh KB. 2010. Plants versus pathogens: an evolutionary arms race. Funct. Plant Biol.. 37: 499-512.
- Bent FB, Mackey D. 2007. Elicitors, effectors, and R genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol.. 45: 399-436.
- Bernard R, Smith P, Kaufmann M, Schmitthenner A. 1957. Inheritance of resistance to Phytophthora root and stem rot in the soybean. Agron. J.. 49: 391-391.
- Bhattacharyya M, Narayanan N, Gao H, Santra D, Salimath S, Kasuga T, et al. 2005. Identification of a large cluster of coiled coil-nucleotide binding site-leucine rich repeat-type genes from the Rps1 region containing Phytophthora resistance genes in soybean. Theor. Appl. Genet.. 111: 75-86.
- Burnham K, Dorrance A, Francis D, Fioritto R, St Martin. 2003a. Rps8, a new locus in soybean for resistance to Phytophthora sojae. Crop Sci.. 43: 101-105.
- Burnham K, Dorrance A, VanToai T, St Martin. 2003b. Quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop Sci.. 43: 1610-1617.
- Buzzell R, Anderson T. 1992. Inheritance and race reaction of a new soybean Rps1 allele. Plant Dis.. 76: 600-601.
- Carter TE, Nelson RL, Sneller CH, Cui Z. Boerma HR., Specht JE., 2004. Genetic diversity in soybean. editors. Soybeans: improvement, production, and uses. 3rd edition. ASA-CSSA-SSSA. Madison, WI, U.S.A.: pp. 303-416.
- Chen Q, Weng Q, Wang Y, Zheng X. 2004. Identification and sequencing of ribosomal DNA-ITS of Phytophthora sojae in Fujian. Acta. Phytopathol. Sin.. 34: 112-116.
- Cheng Y, Ma Q, Ren H, Xia Q, Song E, Tan Z, et al. 2017. Fine mapping of a Phytophthora-resistance gene RpsWY in soybean (Glycine max L.) by high-throughput genome-wide sequencing. Theor. Appl. Genet.. 130: 1041-1051.
- Costamilan LM, Clebsch CC, Soares RM, Seixas CDS, Godoy CV, Dorrance AE. 2013. Pathogenic diversity of Phytophthora sojae pathotypes from Brazil. Eur. J. Plant Pathol.. 135: 845-853.
- Demirbas A, Rector B, Lohnes D, Fioritto R, Graef G, Cregan P, et al. 2001. Simple sequence repeat markers linked to the soybean Rps genes for Phytophthora resistance. Crop Sci.. 41: 1220-1227.
- Dong S, Yin W, Kong G, Yang X, Qutob D, Chen Q, et al. 2011. Phytophthora sojae avirulence effector Avr3b is a secreted NADH and ADP-ribose pyrophosphorylase that modulates plant immunity. PLoS Pathog.. 7: e1002353.
- Dorrance A, Schmitthenner A. 2000. New sources of resistance to Phytophthora sojae in the soybean plant introductions. Plant Dis.. 84: 1303-1308.
- Dorrance A, McClure S, DeSilva A. 2003. Pathogenic diversity of Phytophthora sojae in Ohio soybean fields. Plant Dis.. 87: 139-146.
- Dorrance AE, Jia H, Abney TS. 2004. Evaluation of soybean differentials for their interaction with Phytophthora sojae. Online. Plant Health Prog.
- Dorrance AE, Berry SA, Anderson TR, Meharg C. 2008. Isolation, storage, pathotype characterization, and evaluation of resistance for Phytophthora sojae in soybean. Plant Health Prog.. 10: 1094-1101.
- Dorrance A, Grünwald NJ. Lamour K., Kamoun S., 2009. Phytophthora sojae: among and within populations. editors. Oomycete genetics and genomics: diversity, interactions, and research tools. Wiley. Hoboken, NJ, U.S.A.: pp. 197-212.
- Dorrance A, Kurle J, Robertson A, Bradley C, Giesler L, Wise K, et al. 2016. Pathotype diversity of Phytophthora sojae in eleven states in the United States. Plant Dis.. 100: 1429-1437.
- Dorrance AE. 2018. Management of Phytophthora sojae of soybean: a review and future perspectives. Can. J. Plant Pathol.. 40: 210-219.
- Flor HH. 1955. Host-parasite interaction in flax rust - Its genetics and other implications. Phytopathology. 45: 680-685.
- Gordon SG, Martin S, Dorrance AE. 2006. Rps8 maps to a resistance gene rich region on soybean molecular linkage group F. Crop Sci.. 46: 168-173.
- Gordon S, Kowitwanich K, Pipatpongpinyo W, Martin S, Dorrance A. 2007. Molecular marker analysis of soybean plant introductions with resistance to Phytophthora sojae. Phytopathology. 97: 113-118.
- Grau CR, Dorrance AE, Bond J, Russin JS. Boerma HR., Specht JE., 2004. Fungal diseases. editors. Soybeans: improvement, production, and uses. 3rd edition. ASA-CSSA-SSSA. Madison, WI, U.S.A.: pp. 679-763.
- Han Y, Teng W, Yu K, Poysa V, Anderson T, Qiu L, et al. 2008. Mapping QTL tolerance to Phytophthora root rot in soybean using microsatellite and RAPD/SCAR derived markers. Euphytica. 162: 231-239.
- Huang J, Guo N, Li Y, Sun J, Hu G, Zhang H, et al. 2016. Phenotypic evaluation and genetic dissection of resistance to Phytophthora sojae in the Chinese soybean mini core collection. BMC Genet.. 17: 85
- Jackson T, Kirkpatrick T, Rupe J. 2004. Races of Phytophthora sojae in Arkansas soybean fields and their effects on commonly grown soybean cultivars. Plant Dis.. 88: 345-351.
- Jee H, Kim W, Cho W. 1998. Occurrence of Phytophthora root rot on soybean (Glycine max) and identification of the causal fungus. RDA J. Crop Prot.. 40: 16-22.
- Jeong N, Kim KS, Jeong S, Kim JY, Park SK, Lee JS, et al. 2019. Korean soybean core collection: Genotypic and phenotypic diversity population structure and genome-wide association study. PLoS One. 14: e0224074.
- Jones JD, Dangl JL. 2006. The plant immune system. Nature. 444: 323-329.
- Kaitany R, Hart L, Safir G. 2001. Virulence composition of Phytophthora sojae in Michigan. Plant Dis.. 85: 1103-1106.
- Kang IJ, Kang S, Jang IH, Jang YW, Shim HK, Heu S, et al. 2019. Identification of new isolates of Phytophthora sojae and the reactions of Korean soybean cultivars following hypocotyl inoculation. Plant Pathol. J.. 35: 698
- Kang I, Shim H, Heu S, Kim K. 2020. First report of soybean root and stem rot caused by Fusarium graminearum in South Korea. Plant Dis.. 104: 568-568.
- Karhoff S, Lee S, Mian MAR, Ralston TI, Niblack TL, Doorance AE, et al. 2019. Phenotypic characterization of a major quantitative disease resistance locus for partial resistance to Phytophthora sojae. Crop Sci.. 59: 968-980.
- Kaufmann MJ, Gerdemann J. 1958. Root and stem rot of soybean caused by Phytophthora sojae n. sp. Phytopathology. 48: 4.
- Ko YM, Choi J, Lee YH, Kim HT. 2020. First report of Charcoal rot caused by Macrophomina phaseolina on Glycine max in Korea. Res. Plant Dis.. 26: 29-37.
- Koenning SR, Wrather JA. 2010. Suppression of soybean yield potential in the continental United States by plant diseases from 2006 to 2009. Plant Health Prog.. 11: 5
- Lee S, Mian MR, McHale LK, Wang H, Wijeratne AJ, Sneller CH, et al. 2013a. Novel quantitative trait loci for partial resistance to Phytophthora sojae in soybean PI 398841. Theor. Appl. Genet.. 126: 1121-1132.
- Lee S, Mian R, McHale LK, Sneller CH, Dorrance AE. 2013b. Identification of quantitative trait loci conditioning partial resistance to Phytophthora sojae in soybean PI 407861A. Crop Sci.. 53: 1022-1031.
- Lee S, Mian MR, Sneller CH, Wang H, Dorrance AE, McHale LK. 2014. Joint linkage QTL analyses for partial resistance to Phytophthora sojae in soybean using six nested inbred populations with heterogeneous conditions. Theor. Appl. Genet.. 127: 429-444.
- Li X, Han Y, Teng W, Zhang S, Yu K, Poysa V, et al. 2010. Pyramided QTL underlying tolerance to Phytophthora root rot in mega-environments from soybean cultivars 'Conrad'and 'Hefeng 25'. Theor. Appl. Genet.. 121: 651-658.
- Li L, Lin F, Wang W, Ping J, Fitzgerald JC, Zhao M, et al. 2016. Fine mapping and candidate gene analysis of two loci conferring resistance to Phytophthora sojae in soybean. Theor. Appl. Genet.. 129: 2379-2386.
- Li Y, Sun S, Zhong C, Wang X, Wu X, Zhu Z. 2017. Genetic mapping and development of co-segregating markers of RpsQ, which provides resistance to Phytophthora sojae in soybean. Theor. Appl. Genet.. 130: 1223-1233.
- Lin F, Zhao M, Ping J, Johnson A, Zhang B, Abney TS, et al. 2013. Molecular mapping of two genes conferring resistance to Phytophthora sojae in a soybean landrace PI 567139B. Theor. Appl. Genet.. 126: 2177-2185.
- Liu FP, Gao ZM, Wang XF, Yu C, Pan YM. 2006. On identification and biological characteristics of the causal organism of Phytophthora root rot of soybean in Anhui. J. Guangxi Agric. Biol. Sci.. 25: 48-51.
- Ludke WH, Schuster I, Da Silva, Montecelli TDN, Soares BDA, De Oliverira, et al. 2019. SNP markers associated with soybean partial resistance to Phytophthora sojae. Crop Breed. Appl. Biot.. 19: 31-39.
- Ma SM, Ding JJ, Zheng TQ, Gu X. 2005. The identification of physiological races of Phytophthora megasperma. Soybean Sci.. 24: 260-262.
- Malvick D, Grunden E. 2004. Traits of soybean-infecting Phytophthora populations from Illinois agricultural fields. Plant Dis.. 88: 1139-1145.
- McHale L, Tan X, Koehl P, Michelmore RW. 2006. Plant NBS-LRR proteins: adaptable guards. Genome Biol.. 7: 212
- Mideros S, Nita M, Dorrance AE. 2007. Characterization of components of partial resistance, Rps 2, and root resistance to Phytophthora sojae in soybean. Phytopathology. 97: 655-662.
- Moriwaki J. 2010. Aiming of the construction of the race distinction system of Phytophthora sojae. Plant Protection. 64: 508-510.
- Morris PF, Ward E. 1992. Chemoattraction of zoospores of the soybean pathogen, Phytophthora sojae, by isoflavones. Physiol. Mol. Plant P.. 40: 17-22.
- Nelson B, Mallik I, McEwen D, Christianson T. 2008. Pathotypes, distribution, and metalaxyl sensitivity of Phytophthora sojae from North Dakota. Plant Dis.. 92: 1062-1066.
- Nguyen V, Vuong T, VanToai T, Lee J, Wu X, Mian M, et al. 2012. Mapping of quantitative trait loci associated with resistance to Phytophthora sojae and flooding tolerance in soybean. Crop Sci.. 52: 2481-2493.
- Niu J, Guo N, Sun J, Li L, Cao Y, Li S, et al. 2017. Fine mapping of a resistance gene RpsHN that controls Phytophthora sojae using recombinant inbred lines and secondary populations. Front. Plant Sci.. 8: 538
- Niu J, Guo N, Zhang Z, Wang Z, Huang J, Zhao J, et al. 2018. Genome-wide SNP-based association mapping of resistance to Phytophthora sojae in soybean (Glycine max (L.) Merr.). Euphytica 214. 10: 187
- Ping J, Fitzgerald JC, Zhang C, Lin F, Bai Y, Wang D, et al. 2015. Identification and molecular mapping of Rps11, a novel gene conferring resistance to Phytophthora sojae in soybean. Theor. Appl. Genet.. 129: 445-451.
- Ranathunge K, Thomas RH, Fang X, Peterson CA, Gijzen M, Bernards MA. 2008. Soybean root suberin and partial resistance to root rot caused by Phytophthora sojae. Phytopathology. 98: 1179-1189.
- Ryley M, Obst N, Irwin J, Drenth A. 1998. Changes in the racial composition of Phytophthora sojae in Australia between 1979 and 1996. Plant Dis.. 82: 1048-1054.
- Sahoo DK, Abeysekara NS, Cianzio SR, Robertson AE, Bhattacharyya MK. 2017. A novel Phytophthora sojae resistance Rps12 gene mapped to a genomic region that contains several Rps genes. PLoS One. 12: e0169950.
- Sandhu D, Gao H, Cianzio S, Bhattacharyya MK. 2004. Deletion of a disease resistance nucleotide-binding-site leucine-rich-repeat-like sequence is associated with the loss of the Phytophthora resistance gene Rps4 in soybean. Genetics. 168: 2157-2167.
- Schmitthenner A. 1985. Problems and progress in control of Phytophthora root rot of soybean. Plant Dis.. 69: 362-368.
- Schneider R, Rolling W, Song Q, Cregan P, Dorrance AE, McHale LK. 2016. Genome-wide association mapping of partial resistance to Phytophthora sojae in soybean plant introductions from the Republic of Korea. BMC Genomics. 17: 607
- Slaminko TL, Bowen CR, Hartman GL. 2010. Multi-year evaluation of commercial soybean cultivars for resistance to Phytophthora sojae. Plant Dis.. 94: 368-371.
- St. Martin S, VanToai T, Beuerlein A, Schmitthenner S. 1994. Genetic variability for flooding tolerance in soybeans. Crop Sci.. 34: 1112-1115.
- Stasko AK, Wickramasinghe D, Nauth BJ, Acharya B, Ellis ML, Taylor CG, et al. 2016. High-density mapping of resistance QTL toward Phytophthora sojae, Pythium irregulare, and Fusarium graminearum in the same soybean population. Crop Sci.. 56: 2476-2492.
- Su Y, Shen C. 1993. The discovery and biological characteristic studies of Phytophthora megasperma f. sp. glycinea on soyabean in China. Acta. Phytopathol. Sin.. 23: 341-347.
- Sugimoto T, Yoshida S, Aino M, Watanabe K, Shiwaku K, Sugimoto M. 2006. Race distribution of Phytophthora sojae on soybean in Hyogo, Japan. J. Gen. Plant Pathol.. 72: 92-97.
- Sugimoto T, Yoshida S, Watanabe K, Aino M, Kanto T, Maekawa K, et al. 2007. Identification of SSR markers linked to the Phytophthora resistance gene Rps1-d in soybean. Plant Breed.. 127: 154-159.
- Sugimoto T, Yoshida S, Aino M, Maekawa K, Kanto T, Matoh T, et al. 2010. Pathogenic races of Phytophthora sojae in Glycine max cv. Tanbakuro producing areas in Hyogo and selection of parental soybeans for breeding new resistant cultivars. J. Crop Res.. 55: 19-25.
- Sugimoto T, Yoshida S, Kaga A, Hajika M, Watanabe K, Aino M, et al. 2011. Genetic analysis and identification of DNA markers linked to a novel Phytophthora sojae resistance gene in the Japanese soybean cultivar Waseshiroge. Euphytica. 182: 133
- Sun S, Wu X, Zhao J, Wang Y, Tang Q, Yu D, et al. 2011. Characterization and mapping of RpsYu25, a novel resistance gene to Phytophthora sojae. Plant Breed.. 130: 139-143.
- Sun J, Li L, Zhao J, Huang J, Yan Q, Xing H, et al. 2014. Genetic analysis and fine mapping of RpsJS, a novel resistance gene to Phytophthora sojae in soybean [Glycine max (L.) Merr.]. Theor. Appl. Genet.. 127: 913-919.
- Thomas R, Fang X, Ranathunge K, Anderson TR, Peterson CA, Bernards MA. 2007. Soybean root suberin: anatomical distribution, chemical composition, and relationship to partial resistance to Phytophthora sojae. Plant Physiol.. 144: 299-311.
- Tooley P, Grau C. 1984. Field characterization of rate-reducing resistance to Phytophthora megasperma f. sp. glycinea in soybean. Phytopathology. 74: 1201-1208.
- Tsuchiya S, Tanaka F, Adachi T. 1990. Classification of race of Phytophthora sojae and screening of resistant soybean cultivars to Phytophthora stem rot. Ann. Phytopathol. Soc. Jpn.. 56: 144.
- Tucker D, Maroof S, Mideros S, Skoneczka J, Nabati D, Buss G, et al. 2010. Mapping quantitative trait loci for partial resistance to Phytophthora sojae in a soybean interspecific cross. Crop Sci.. 50: 628-635.
- Tyler BM. 2007. Phytophthora sojae: root rot pathogen of soybean and model oomycete. Mol. Plant Pathol.. 8: 1-8.
- Walker A, Schmitthenner A. 1984. Heritability of tolerance to Phytophthora rot in soybean. Crop Sci.. 24: 490-491.
- Wang H, Li G, Zhan Y, Wang P, Zhang P. 2006. Identification of Phytophthora root rot of soybean in Xinjiang. Xinjiang Agricultural Sciences. 43: 106-108.
- Wang H, Waller L, Tripathy S, St Martin, Zhou L, Krampis K, et al. 2010. Analysis of genes underlying soybean quantitative trait loci conferring partial resistance to Phytophthora sojae. Plant Genome. 3: 23-40.
- Wang H, St Martin, Dorrance AE. 2012. Comparison of phenotypic methods and yield contributions of quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Crop Sci.. 52: 609-622.
- Wen J, Chen H. 2002. Races and virulence phenotypes of Phytophthora sojae in Heilongjiang province and Mongolia Region of China. Chinese Journal of Oil Crop Sciences. 24: 63-66.
- Weng C, Yu K, Anderson T, Poysa V. 2001. Mapping genes conferring resistance to Phytophthora root rot of soybean, Rps1a and Rps7. J. Hered.. 92: 442-446.
- Weng C, Yu K, Anderson TR, Poysa V. 2007. A quantitative trait locus influencing tolerance to Phytophthora root rot in the soybean cultivar 'Conrad'. Euphytica. 158: 81-86.
- Wrather J, Stienstra W, Koenning S. 2001. Soybean disease loss estimates for the United States from 1996 to 1998. Can. J. Plant Pathol.. 23: 122-131.
- Wrather JA, Koenning SR. 2006. Estimates of disease effects on soybean yields in the United States 2003 to 2005. J. Nematol.. 38: 173
- Wrather JA, Koenning SR. 2009. Effects of diseases on soybean yields in the United States 1996 to 2007. Plant Health Prog.. 10: 24
- Wu X, Zhang B, Shi S, Zhao J, Feng Y, Na G, et al. 2011a. Identification, genetic analysis and mapping of resistance to Phytophthora sojae of Pm28 in soybean. Agr. Sci. China. 10: 1506-1511.
- Wu X, Zhou B, Sun S, Zhao J, Chen S, Gai J, et al. 2011b. Genetic analysis and mapping of resistance to Phytophthora sojae of Pm14 in soybean. Scientia Agricultura Sinica. 44: 456-460.
- Wu X, Zhou B, Zhao J, Guo N, Zhang B, Yang F, et al. 2011c. Identification of quantitative trait loci for partial resistance to Phytophthora sojae in soybean. Plant Breed.. 130: 144-149.
- Xiao J, Miao M, Gao K, Xiang G, Yang S, Dong S, et al. 2011. Genetic diversity of Phytophthora sojae in China based on RFLP. Scientia Agricultura Sinica. 44: 4190-4198.
- Yao HY, Wang XM, Wu XF, Xiao YN, Zhu ZD. 2010. Molecular mapping of Phytophthora resistance gene in soybean cultivar Zaoshu18. J. Plant Genet. Resour.. 11: 213-217.
- Yu A, Xu P, Wang J, Zhang S, Wu J, Li W, et al. 2010. Genetic analysis and SSR mapping of gene resistance to Phytophthora sojae race 1 in soybean cv Suinong 10. Chinese Journal of Oil Crop Sciences. 32: 462-466.
- Zhang S, Xu P, Wu J, Xue AG, Zhang J, Li W, et al. 2010. Races of Phytophthora sojae and their virulences on soybean cultivars in Heilongjiang, China. Plant Dis.. 94: 87-91.
- Zhang J, Xia C, Duan C, Sun S, Wang X, Wu X, et al. 2013a. Identification and candidate gene analysis of a novel Phytophthora resistance gene Rps10 in a Chinese soybean cultivar. PLoS One. 8: e69799.
- Zhang J, Xia C, Wang X, Duan C, Sun S, Wu X, et al. 2013b. Genetic characterization and fine mapping of the novel Phytophthora resistance gene in a Chinese soybean cultivar. Theor. Appl. Genet.. 126: 1555-1561.
- Zhong C, Sun S, Li Y, Duan C, Zhu Z. 2017. Next-generation sequencing to identify candidate genes and develop diagnostic markers for a novel Phytophthora resistance gene, RpsHC18, in soybean. Theor. Appl. Genet.. 131: 525-538.
- Zhong C, Li Y, Sun S, Duan C, Zhu Z. 2019. Genetic mapping and molecular characterization of a broad-spectrum Phytophthora sojae resistance gene in Chinese soybean. Int. J. Mol. Sci.. 20: 1809
- Zhu Z, Wang H, Wang X, Chang R, Wu X. 2003. Distribution and virulence diversity of Phytophthora sojae in China. Agr. Sci. China. 3: 116-123.
- Zhu Z, Huo Y, Wang X, Huang J, Wu X. 2007. Molecular identification of a novel Phytophthora resistance gene in soybean. Acta Agron. Sin.. 33: 154-157.