Abstract
In this study, two near-isogenic lines (NILs) were selected from a series of chromosome segment substitution lines developed from an interspecific cross between Oryza sativa ssp. japonica cultivar ‘Hwaseongbyeo’ as a recipient parent and wild rice (Oryza grandiglumis) as the donor parent. Compared with Hwaseongbyeo control, one NIL (CR222) had reduced plant height and the other NIL (CR7501) displayed thicker basal and first internodes leading to increased lodging tolerance in the field condition. QTL analysis using progeny from crosses between Hwaseongbyeo and the NILs indicated that QTL for culm length and internode diameter leading to erect panicle were located on chromosomes 1 and 2, respectively. The major QTL for culm length on chromosome 1 was allelic to sd1 locus and the NIL showed the same sequence as the sd1 mutant cultivar ‘Reimei’. The QTL for the internode diameter was delimited to a 1-Mbp region on chromosome 2 and this QTL region harbors GW2 gene controlling grain width. The NIL with thick internode trait showed no undesirable pleiotropic effects and therefore has potential value for rice breeding for increasing lodging resistance.
-
Key words: Culm length, Internode diameter, QTL, Rice, Near Isogenic Lines
INTRODUCTION
Rice (
Oryza sativa) is one of the most important food crops for mankind. Rice productivity was significantly increased in the 1960s and 1970s by the introduction and use of the semi-dwarfing gene
sd1 (
Khush 1999;
Sasaki et al. 2002). The recessive semidwarf 1 (
sd1) gene in rice confers a shortened culm and a greater harvest index, resulting in lodging resistance and responsiveness to fertilizer.
sd1 has been introduced into high-yielding rice varieties in the tropics and other areas since 1960’s (
Suh and Heu 1978;
Ashikari et al. 2002;
Spielmeyer et al. 2002). The Green Revolution is a direct achievement of an intensive use of the
SD1 gene originated from the Chinese cultivar ‘Dee-geo-woo-gen’ (DGWG). In Korea, Tongil rice (IR667-98-1-2) is the first semi-dwarf variety obtained by a three-way cross of
indica/japonica varieties (
Suh and Heu 1978;
Kim et al. 2014). It is also known that the γ-ray-induced semi-dwarf cultivars ‘Reimei’ in Japan and ‘Calrose 76’ in the USA (
Foster and Rutger 1978) have been widely used in rice breeding programs.
Lodging resistance is one of the major targets in rice breeding because it is involved in grain yield and quality, and mechanical harvesting efficiency (
Hoshikawa and Wang 1990;
Setter et al. 1997). For stem lodging resistance, it is important to increase the physical strength of the basal culm which is determined by the morphology and the quality of its components. Because stem diameter and weight have been directly correlated with lodging resistance and the breaking strength of the stem among morphological traits, increasing the stem strength of the lower internodes of rice plants is the major focus for improving lodging resistance (
Terashima et al. 1994;
Islam et al. 2007;
Kashiwagi et al. 2008). A number of QTL associated with culm strength and diameter have been identified from
indica cultivars including
SCM2 (
Kashiwagi et al. 2008;
Ookawa et al. 2010;
Yano et al. 2015). Considering that culm diameter is the potential target in breeding for better lodging resistance, additional efforts are needed to identify new genes for culm diameter to improve lodging resistance in rice.
Wild rice not only possesses a large number of potentially useful agronomic characters but also has a large amount of genetic diversity (
Xiao et al. 1998). The wild rice species (
O. grandiglumis) possess beneficial traits for increased tolerance to biotic and abiotic stresses (
Vaughan et al. 2003). However, efforts to introduce useful genes from
O. grandiglumis into cultivated rice to improve high-yielding Asian cultivated rice are limited due to difficulty in interspecific crosses with
O. sativa and tall growth, which has limited their use to date in rice breeding.
In our previous research, we developed a near-isogenic line (NIL) from an interspecific cross between japonica cultivar ‘Hwaseongbyeo’ as a recipient parent and wild rice (O. grandiglumis) as the donor parent. The NIL displayed reduced plant height and thicker internode compared to Hwaseongbyeo. We performed morphological observations and QTL mapping using progeny from a cross between Hwaseongbyeo and NIL to map QTL underlying these traits. Also, sequencing of the SD1 gene was also carried out to know the sequence difference between two parents with the goal of utilizing the gene for culm length from O. grandiglumis into O. sativa for use in rice breeding programs.
MATERIALS AND METHODS
Plant materials
In our previous study, QTL for agronomic traits including grain shape traits, were detected in the 150 F
2:3 families (
Yoon et al. 2006). For fine mapping of genes for culm length and first internode diameter, two NILs CR222 and CR7501 were selected based on the phenotype and crossed with Hwaseongbyeo to produce F
2, F
3 and F
4 populations, respectively. These populations were used as the mapping populations in this study.
For CR222 population, 659 F2 plants and 25 F3 lines were grown in the field during the summer of 2016 and 2017 at Chungnam National University, Daejeon, respectively. For CR7501 population, 65 F3 lines and F4 lines were grown in the field during the summer of 2016 and 2017 at Chungnam National University, Daejeon, respectively. F2 plants were planted with 15 cm between plants and 30 cm between rows. For the F3 and F4 population, each family was represented by a single row of thirty 30-day-old seedlings planted with 15 cm between plants and 30 cm between rows in a completely randomized block design with two replications. Three traits, culm length, days to heading and first internode diameter were evaluated in the F2, F3 and F4 populations, respectively. Days to heading (DTH) was evaluated as the number of days from seeding until the first panicle flowered. For culm length each plant was measured. For first internode diameter, ten plants per line in the middle were measured, and the average of the measurements was used as the phenotype of each line. Culm length (CL) was measured in centimeters from the soil surface to the neck of tallest panicle. First internode diameter (FID) was measured 10 mm below the panicle neck in millimeters. For substitution mapping of the FID QTL, 18 F4 lines were employed.
DNA extraction, SSR genotyping and sequencing
DNA was extracted from fresh leaves of F
2 plants as described in
Causse et al. (1994). SSR markers showing polymorphisms between Hwaseongbyeo and two NILs in the
O. grandiglumis introgression regions were used for genotyping of F
2 plants (
McCouch et al. 2002). SSR analysis was performed according to the method described in
Panaud et al. (1996), with the following modification in the PCR profile: 94°C for 5 minutes, followed by 35 cycles of 94°C for 1 minute, 55°C for 1 minute, and 72°C for 1 minute, and lastly 5 minutes at 72°C. Analysis of the
SD1 gene sequences including exon and intron of both parents were carried out in Macrogen, Korea (
www.macrogen.co.kr).
One-way ANOVA was performed to determine the effect of each marker on each trait. QTL was declared if the phenotype was associated with a marker locus at P < 0.005 or with two adjacent marker loci at P < 0.05. The proportion of the total phenotypic variation explained by each QTL was calculated as an R2 value, from the regressions of each marker/phenotype combination. QTLs were fine mapped by comparing the phenotypic means of the F3 lines with different O. grandiglumis introgressions within the target region using the ANOVA feature in the SAS 9.0 program.
RESULTS
Genetic background of NILs
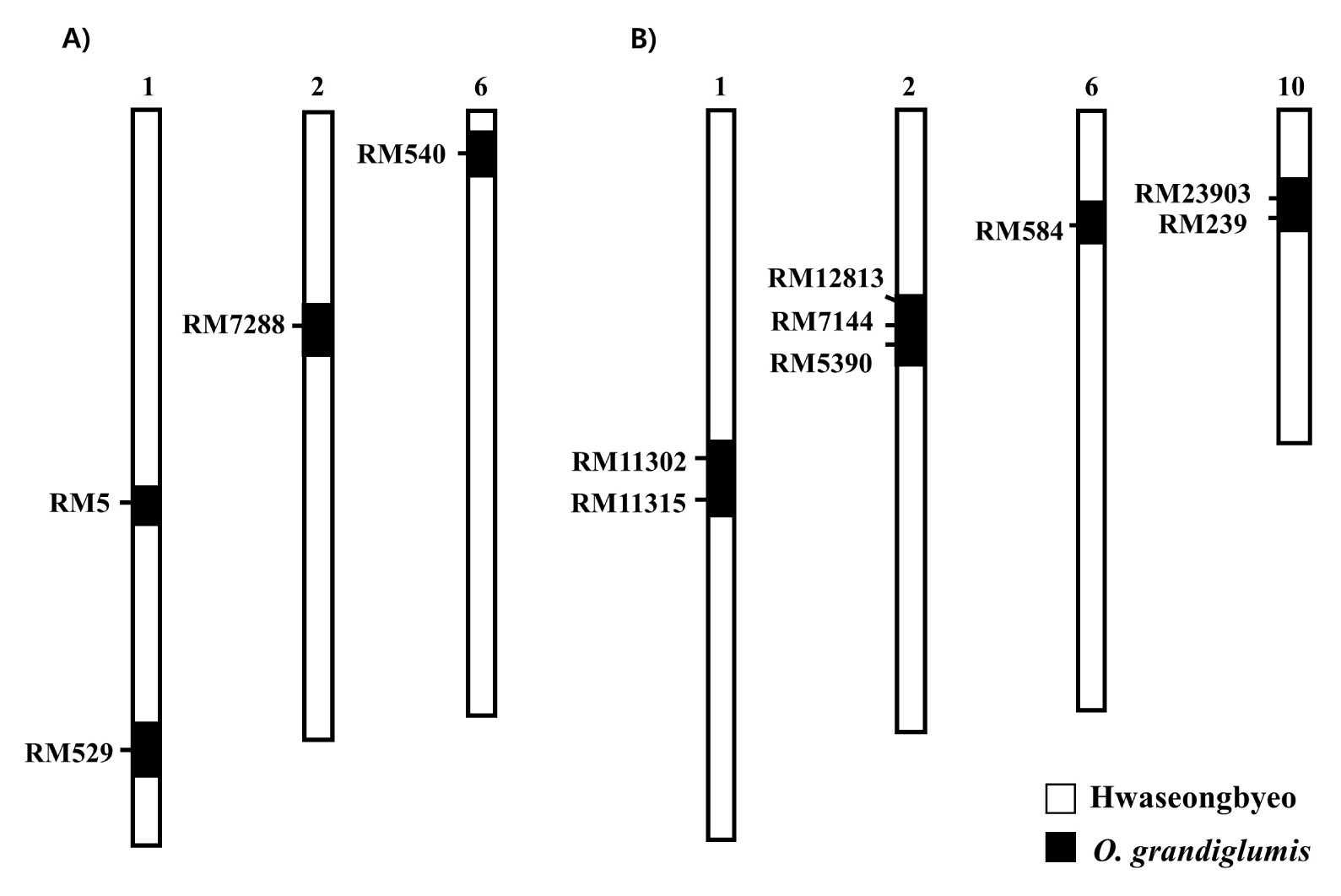
A total of four
O. grandiglumis chromosome segments were detected in CR222 whereas four introgressions were detected in CR7501 (
Fig. 1). The introgressed segments were distributed on chromosomes 1, 2 and 6 in CR222 and on chromosomes 1, 2, 6 and 10 in CR7501, respectively (
Fig. 1). It could not be determined whether each introgressed segment was of CC or DD genome origin from
O. grandiglumis. SSR markers located in the target regions were used to genotype F
2 plants.
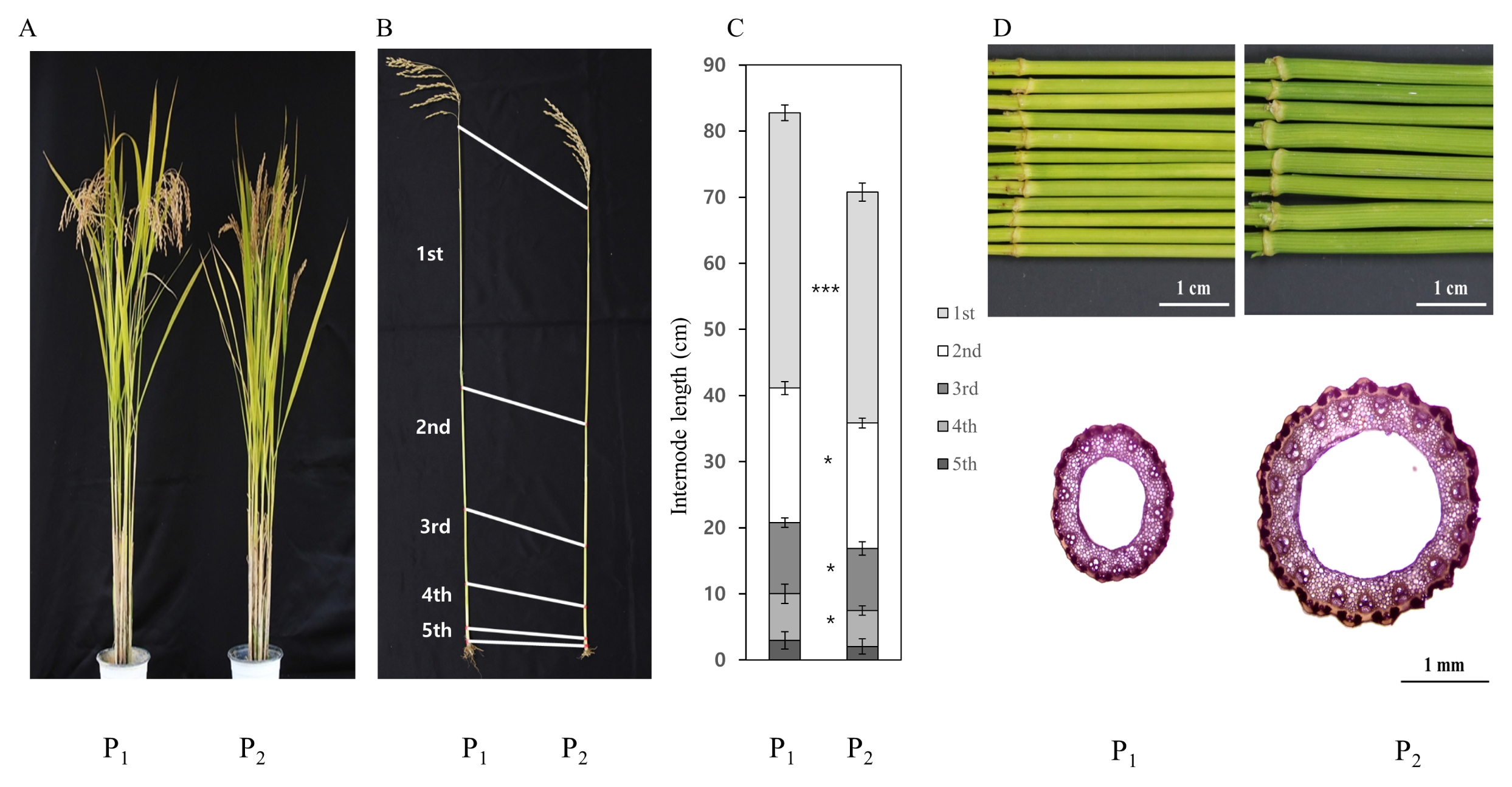
Significant difference for culm length, and first internode diameter was observed between Hwaseongbyeo and CR222, and Hwaseongbyeo and CR7501 (
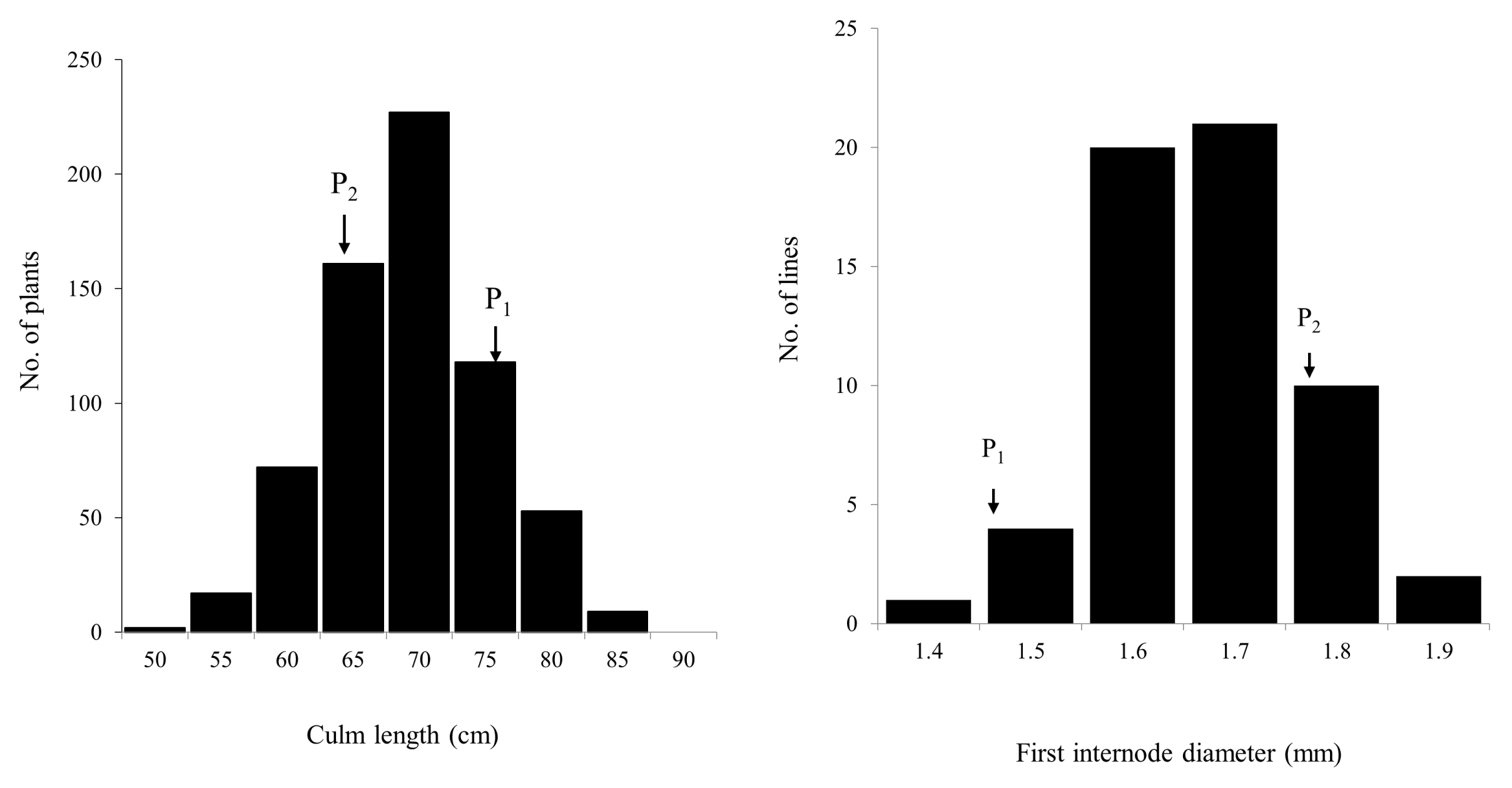
Fig. 2). Frequency distribution for two traits (CL in the F
2 and FID in the F
3) in the population showed nearly normal variation (
Fig. 3). Most of the F
2 plants fell within the range of the parents but some transgressive segregants exceeded the value of CR222. Days to heading for Hwaseongbyeo and CR222 were nearly the same (108 days).
SSR markers on the introgressed segments were used to genotype the F
2 individuals. In the F
2 population, two QTL for culm length were detected on chromosome 1 (
Table 1). The
O. grandiglumis alleles decreased culm length at two QTL loci. The two QTL,
qCL1.1 and
qCL1.2 explained 12.6% and 28.5% of the total phenotypic variance for CL, respectively. 65 F
3 lines were employed for mapping QTL for FID and one QTL was mapped near the SSR marker, RM7144 on chromosome 2. The phenotypic variance explained by this QTL on chromosome 2 was 18.9% in the F
3 population.
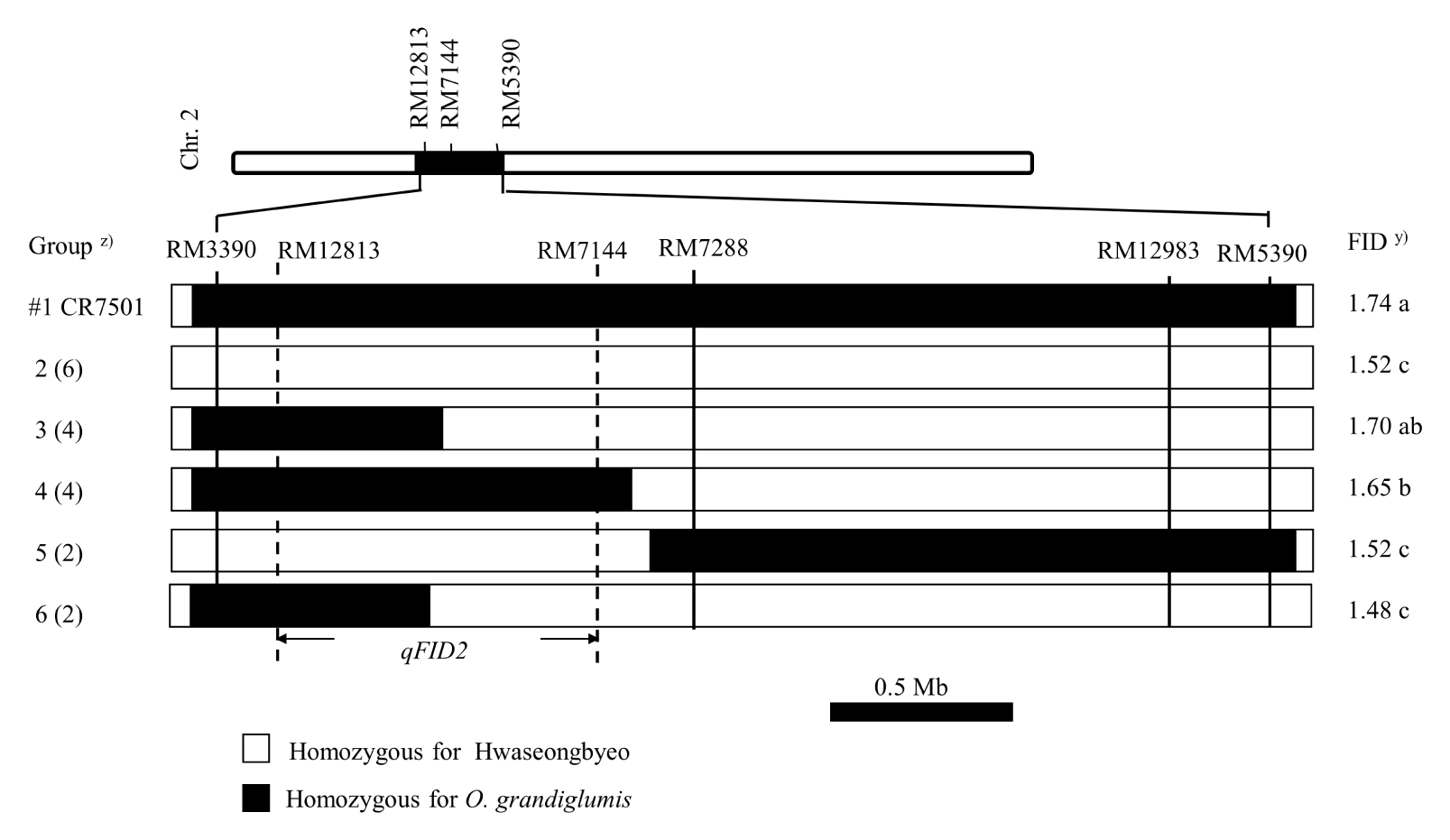
Eighteen recombinant F
4 lines with different
O. grandiglumis segments were selected and employed for the substitution mapping of
qFID2 (
Fig. 4). Plants of two groups (3 and 4) showed significant higher FID values than those of groups #2, 5, and 6. Two groups 3 and 4 had the segment between two SSR markers RM3390 and RM7288. Although group 6 had the segment between RM3390 and RM7288, FID of this group plants was shorter than those of plants in group 3. These results indicate that
qFID2 is located between RM12813 and RM7144, about 1.0-Mb apart.
CR222 plants exhibited an obviously shortened stature relative to Hwaseongbyeo (
Fig. 2). Heading date of the NIL was similar with Hwaseongbyeo (data not shown). In order to investigate the differences between two parents in plant height, we compared internode lengths between these two lines at maturity. In rice, internode elongation occurs from the bottom at the panicle initiation stage. Elongations of the first (uppermost), second and third internodes were most strongly inhibited in CR222 (
Fig. 2).
SD1 was regarded as the possible target gene considering its high effect on culm length based on the finding that the culm length QTL
qCL1.2 was detected near SSR marker, RM529 for F
2 and F
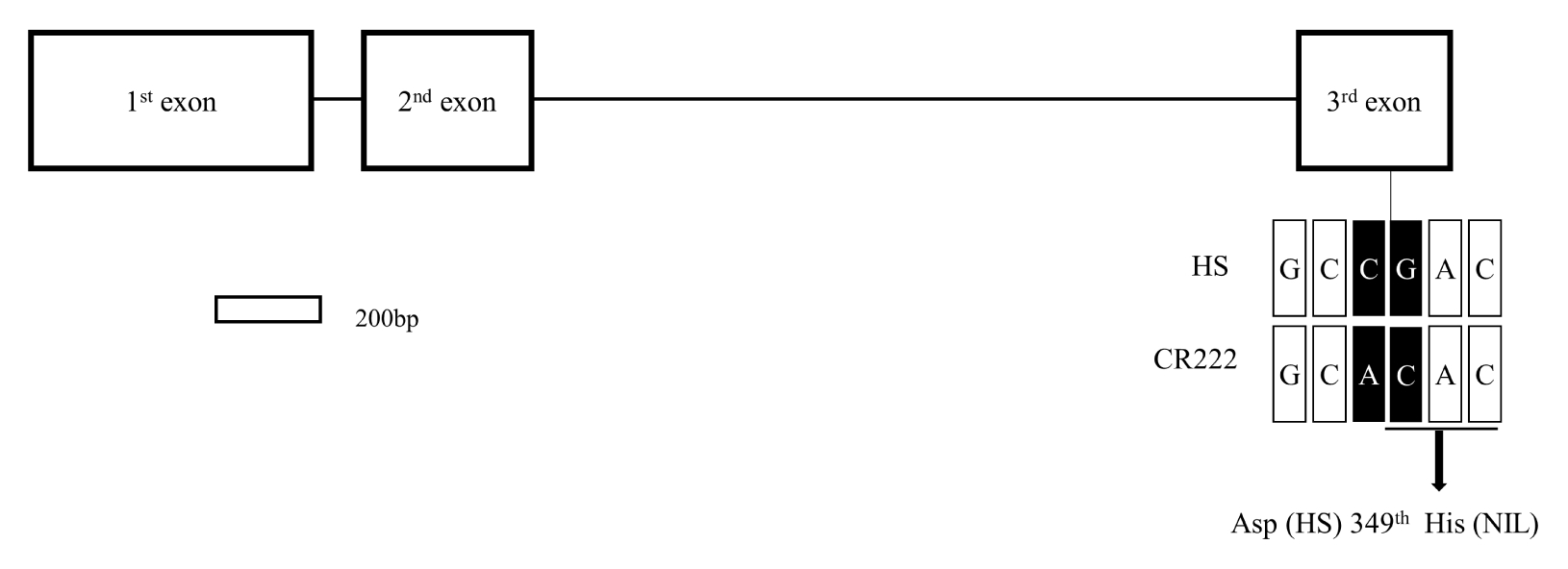
3 generations. Sequence analysis of two parents revealed that there was two SNPs in the
SD1 DNA sequence between Hwaseongbyeo and CR222 allele. The SNP located in the third exon did lead to amino acid change from aspartic acid to histidine at the 349
th amino acid position whereas the other SNP did not lead to amino acid change (
Fig. 5). The CR222
sd1 allele shared the same SNP leading to amino acid change in the 3
rd exon as the one in the semi-dwarf mutant line ‘Reimei’ (
Asano et al. 2007).
DISCUSSION
Plant height is an important agronomic trait in crop breeding because an appropriate plant height is important for improving lodging resistance leading to yield increase (
Sasaki et al. 2002;
Kashiwagi et al. 2008). In this study, we used isogenic lines from an interspecific cross between Hwaseongbyeo as the recurrent parent and wild rice (
O. grandiglumis) as the donor. The results showed that
sd1 gene was associated with reduced culm length in CR222 and the QTL for internode diameter was located in a 1-Mb region on chromosome 2.
Semi-dwarfism has been introduced into high-yielding rice varieties in the tropics and other areas since 1960’s. The Green Revolution is a direct achievement of an intensive use of the
SD1 gene. Comprehensive studies have indicated that most of the short-statured varieties possess the same gene for semi-dwarfism, because a Taiwan-native variety Dee-geo-woo-gen (DGWG) has been used as the common gene source (
Hargrove 1979;
Mackill and Rutger 1979;
Chang and Li 1980). The loss-of-function of
SD1 was successful in
japonica rice because several groups indicated that
sd1 alleles have been widely used in breeding programs (
Ashikari et al. 2002;
Spielmeyer et al. 2002).
Asano et al. (2007) reported that 38 out of 53 semi-dwarf high-yielding lines from China, USA and Japan shared an
sd1 allele and these
sd1 alleles could be grouped into a total of seven different types. To examine the
sd1 allele type of the NIL, we determined the
SD1 sequence of the NIL. Sequence analysis of two parents revealed that one SNP was detected in the third exon between Hwaseongbyeo and CR222 allele. This SNP located in the third exon induced a single amino acid change from aspartic acid to histidine at the 349
th amino acid position. The mutation in CR222
sd1 allele shared by Reimei may result in a partial loss of the GA20ox activity and would be suitable for producing rice lines with the desired height (
Futsuhara 1967;
Asano et al. 2007).
CR222 plants exhibited a shortened stature relative to Hwaseongbyeo although no difference was observed in heading date between Hwaseongbyeo and CR222. In order to investigate the differences between two parents in plant height, we compared internode lengths between these two lines at maturity. Lengths of the first (uppermost), second, third and fourth internodes in CR222 were significantly shorter than those in Hwaseongbyeo whereas no difference was observed in the fifth internode. More than 70 dwarf and semi-dwarf genes have been reported and cloned in rice (
Yang et al. 2014). Studies reported that some rice dwarfing genes showed specific patterns in inhibiting the elongation of internode(s) (
Takeda 1977). Among them, the
sd1 allele of Reimei reduces the length of the upper three internodes proportionally and this result is similar to the previous report (
Wu et al. 2000).
Several studies indicate that stem (internode) diameter correlated with breaking strength is currently the potential target in breeding for better lodging resistance in rice (
Kashiwagi et al. 2008). A number of QTL for culm strength were mapped and isolated using progeny from diverse cross combinations (
Kashiwagi et al. 2008;
Ookawa et al. 2010;
Yano et al. 2015).
Kashiwagi et al. (2008) mapped QTL for stem diameter using NILs from a cross between Nipponbare and Kasalath and demonstrated that NIL28 with a combination of four QTL had greater diameter at all internodes compared with NIL114 with one QTL.
Ookawa et al. (2010) isolated a
SCM2 QTL using progeny from a cross between Sasanishiki and Habataki which was found to be identical to
Aberrant Panicle Organization1 (
APO1) gene (
Ikeda et al. 2007).
SCM3 responsible culm section module was isolated progeny from an intersubspecific cross between Koshihikari and Habataki (
Yano et al. 2015). However, no study reported QTL for internode thickness in the 1-Mb region on the short arm of chromosome 2 and this
qFID2 appears to be a new QTL for internode diameter. This 1-MB chromosomal region also harbors
GW2 gene associated with grain width and CR7501 plants have larger grains than Hwaseongbyeo. The
qFID2 QTL for increased internode diameter coming from an accession of
O. grandiglumis not widely utilized in rice breeding programs would make a contribution for developing lines with better lodging resistance in combination with other QTL detected in other reports. Additional crosses are underway to fix this QTL in a Hwaseongbyeo genetic background with as little additional
O. grandiglumis (donor) chromosome as possible and to isolate the gene underlying this trait.
ACKNOWLEDGEMENTS
This study was financially supported by research fund of Chungnam National University in 2016.
Fig. 1Graphical genotypes of two NILs, (A) CR222 and (B) CR7501. The white bar and black bar show segments of the Hwaseongbyeo and O. grandiglumis.
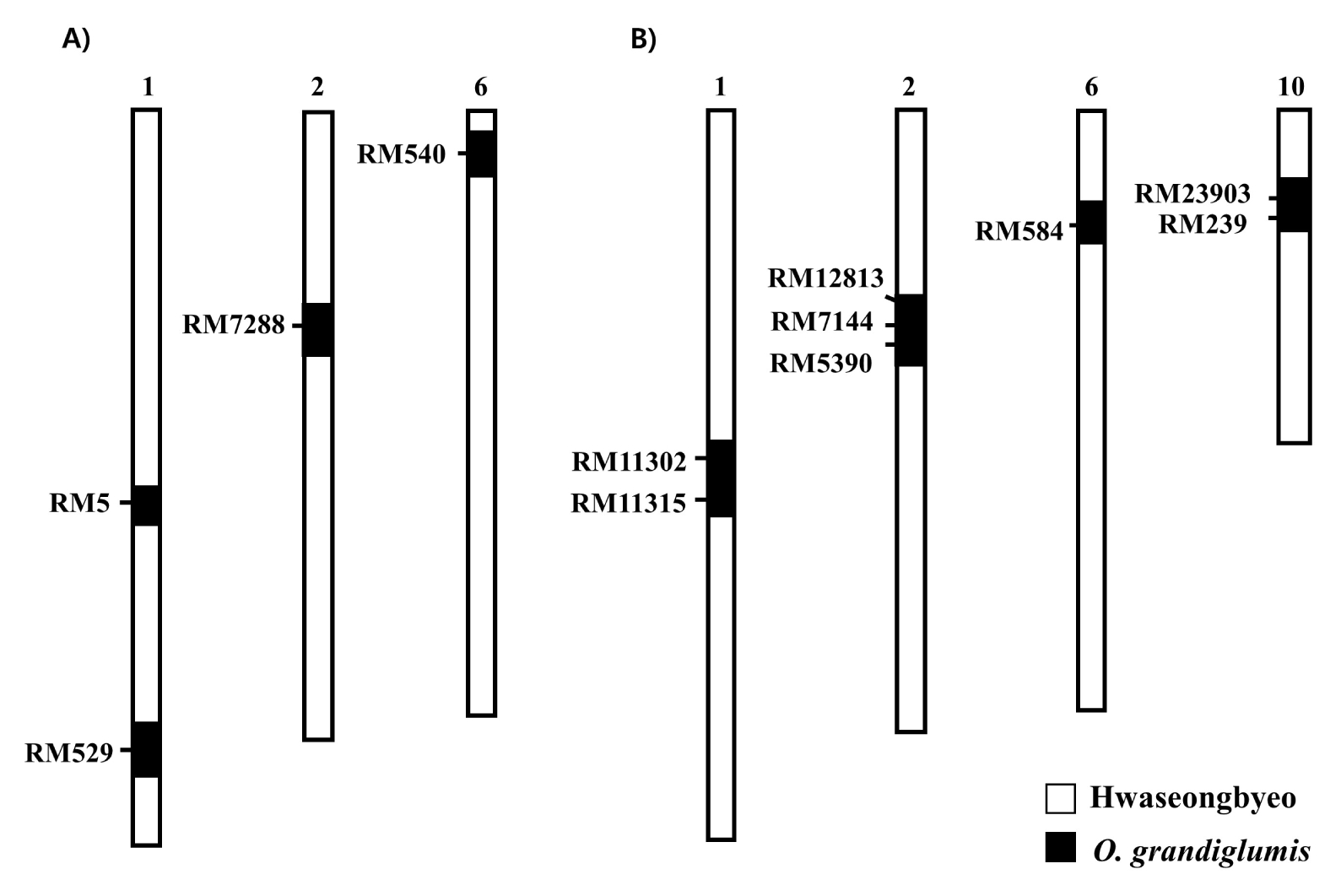
Fig. 2Phenotypic comparison between Hwaseongbyeo and two NILs, CR222 and CR7501. (A) Phenotype at the maturing stage. (B) Designation of 1st to 5th internode. (C) Length comparison of each internode between Hwaseongbyeo and CR222. Error bars represent mean ± SD (n = 8, * and *** indicate significantly different at the 0.05 and 0.005 probability level, respectively, using t-test). (D) Comparison of first internode diameter (above) and cross-dissection of the internode. P1: Hwaseongbyeo, P2: CR222 (A, B, C), CR7501 (D).
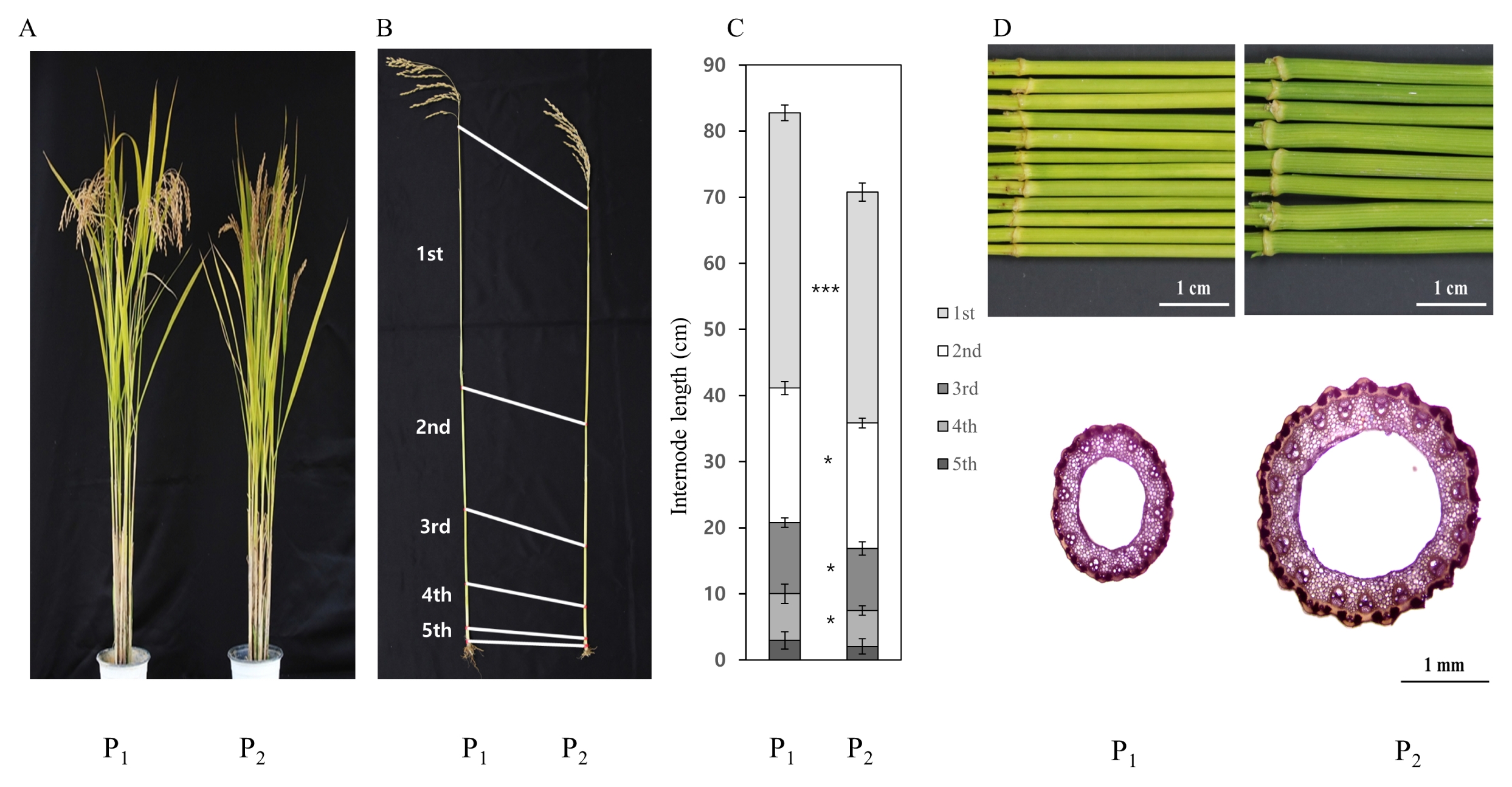
Fig. 3Frequency distributions of culm length in F2 (CR222) and first internode diameter in F3 (CR7501) populations. (P1: Hwaseongbyeo, P2: CR222 or CR7501).
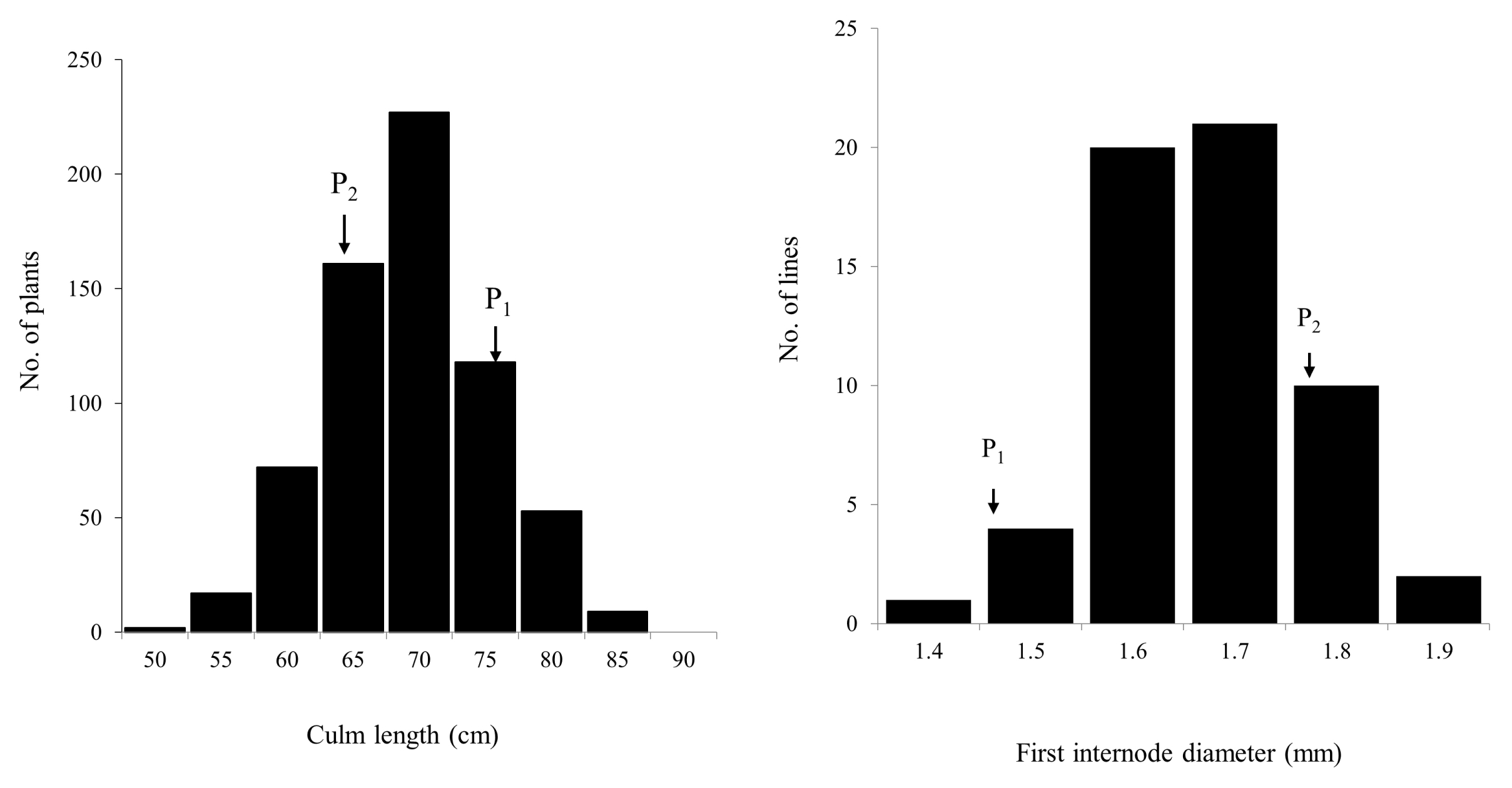
Fig. 4Substitution mapping of the qFID2 QTL. FID was measured with 10 panicles in 2 replicates. z)Values in parentheses are number of lines in each group. y)Numbers followed by the same letter in each column are not significantly different at the 0.05 probability level based on Tukey test.
Fig. 5Sequence comparison of two parental lines in sd1 gene. The structure of sd1 gene located on target region on chromosome 1. The white boxes and black lines between white boxes indicate exons and introns, respectively. The arrow indicates nucleotides substitution resulting in missense mutation. HS: Hwaseongbyeo, NIL: CR222.
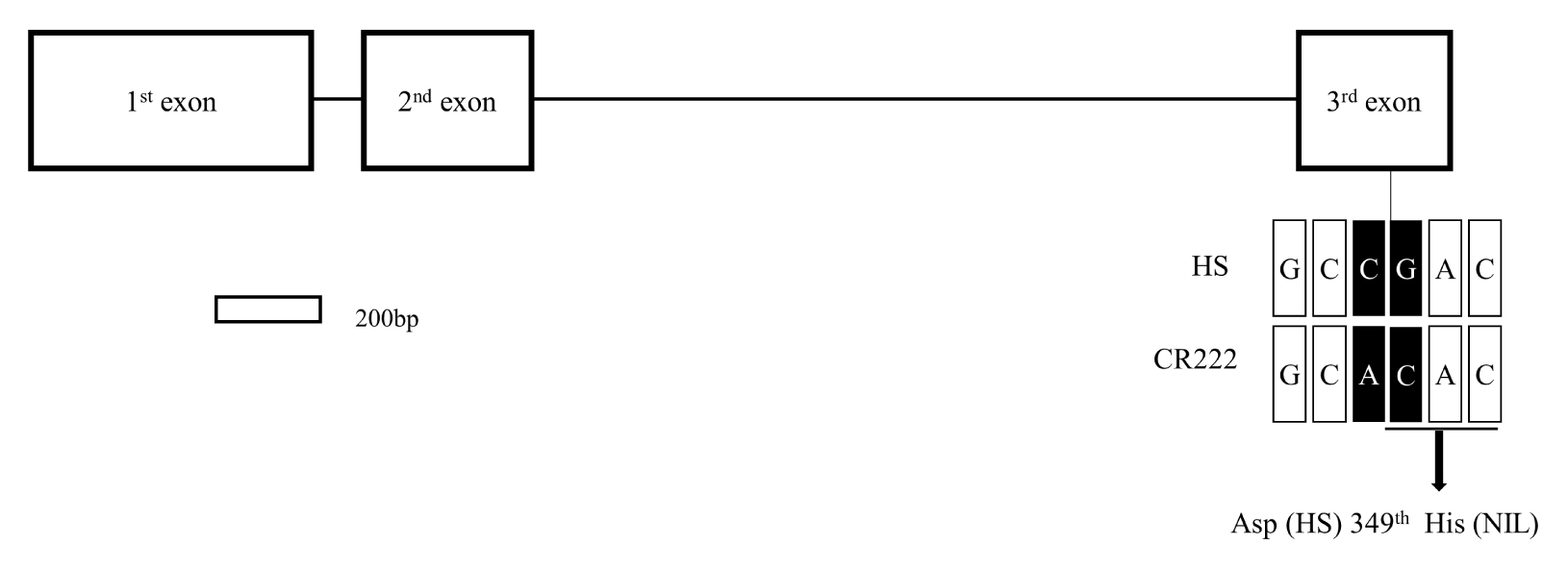
Table 1QTL for culm length and first internode diameter traits in the segregating populations.
Table 1
|
Traitz)
|
Gen. |
QTL |
Chr. |
Marker |
P-value |
R2 (%) |
Meany)
|
|
|
H/H |
H/G |
G/G |
|
CL (cm) |
F2
|
qCL1.1
|
1 |
RM5–RM488 |
0.01 |
12.6 |
69.8 |
67.6 |
65.5 |
|
F2
|
qCL1.2
|
1 |
RM529 |
0.005 |
28.5 |
72.1 |
67.2 |
66.2 |
|
FID (mm) |
F3
|
qFID2
|
2 |
RM7144 |
0.010 |
18.9 |
1.60 |
- |
1.66 |
|
F4
|
qFID2
|
2 |
RM12813 |
0.001 |
27.9 |
1.54 |
- |
1.68 |
References
- Asano K, Takashi T, Miura K, Qian Q, Kitano H, Matsuoka M, et al. 2007. Genetic and molecular analysis of utility of sd1 alleles in rice breeding. Breed Sci. 57: 53-58.
- Ashikari M, Sasaki A, Ueguchi-Tanaka M, Itoh H, Nishimura A, Datta S, et al. 2002. Loss-of-function of a rice gibberellin biosynthetic gene, GA20 oxidase (GA20ox-2), led to the rice ‘green revolution’. Breed Sci. 52: 143-150.
- Causse MA, Fulton TM, Cho YG, Ahn SN, Chunwongse J, Wu K, et al. 1994. Saturated molecular map of the rice genome based on an interspecific backcross population. Genetics. 138: 1251-1274.
- Chang TT, Li CC. 1980. Genetics and breeding. pp. 87-146. Luh BS, editor. Rice: production and utilization. AVI Publishing Company, Inc. Westport. Connecticut.
- Foster KW, Rutger JN. 1978. Inheritance of semidwarfism in rice, Oryza sativa L. Genetics. 88: 559-574.
- Futsuhara Y. 1967. Breeding a new rice variety Reimei by gamma-ray irradiation. Gamma Field Symp. 7: 87-109.
- Hargrove TR. 1979. Diffusion and adoption of semidwarf rice cultivars as parents in Asian rice breeding programs. Crop Sci. 19: 571-574.
- Hoshikawa K, Wang SB. 1990. Studies lodging in rice plants. I. A general observation on lodged rice culms. Jpn J Crop Sci. 59: 809-814.
- Ikeda K, Ito M, Nagasawa N, Kyozuka J, Nagato Y. 2007. Rice ABERRANT PANICLE ORGANIZATION 1, encoding an F-box protein, regulates meristem fate. Plant J. 51: 1030-1040.
- Islam MS, Peng S, Visperas RM, Ereful N, Bhuiya MSU, Julfiquar AW. 2007. Lodging related morphological traits of hybrid rice in a tropical irrigated ecosystem. Field Crops Res. 101: 240-248.
- Kashiwagi T, Togawa E, Hirotsu N, Ishimaru K. 2008. Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.). Theor Appl Genet. 117: 749-757.
- Khush GS. 1999. Green revolution: preparing for the 21st century. Genome. 42: 646-655.
- Kim B, Kim DG, Lee G, Seo J, Choi IY, Choi BS, et al. 2014. Defining the genome structure of ‘Tongil’ rice, an important cultivar in the Korean “Green Revolution”. Rice. 7: 22
- Mackill DJ, Rutger JN. 1979. The inheritance of induced-mutant semidwarfing genes in rice. J Hered. 70: 335-341.
- McCouch SR, Teytelman L, Xu Y, Lobos KB, Clare K, Walton M, et al. 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 9: 199-207.
- Ookawa T, Hobo T, Yano M, Murata K, Ando T, Miura H, et al. 2010. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat Comm. 1: 132
- Panaud O, Chen X, McCouch SR. 1996. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol Gen Genet. 252: 597-607.
- Sasaki A, Ashikari M, Ueguchi-Tanaka M, Itoh H, Nishimura A, Swapan D, et al. 2002. Green revolution: a mutant gibberellin-synthesis gene in rice. Nature. 416: 701-702.
- Setter TL, Laureles EV, Mazaredo AM. 1997. Lodging reduces yield of rice by self-shading and reduction in canopy photosynthesis. Field Crops Res. 49: 95-106.
- Spielmeyer W, Ellis MH, Chandler PM. 2002. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc Natl Acad Sci USA. 99: 9043-9048.
- Suh HS, Heu MH. 1978. The segregation mode of plant height in the cross of rice varieties, VI. Linkage analysis of the semi-dwarfness of the rice variety “Tongil”. Korean J Breed. 10: 1-6.
- Takeda K. 1977. Internode elongation and dwarfism in some gramineous plant. Gama Field Symp. 16: 1-18.
- Terashima K, Ogata T, Akita S. 1994. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation. II. Root growth characteristics of tolerant cultivars to root lodging. Jpn J Crop Sci. 63: 34-41.
- Vaughan DA, Morishima H, Kadowaki K. 2003. Diversity in the Oryza genus. Curr Opin Plant Biol. 6: 139-146.
- Wu X, Saeda T, Takeda K, Kitano H. 2000. Dominant gene, Ssi1 expresses semidwarfism by inhibiting the second internode elongation in rice. Breed Sci. 50: 17-22.
- Xiao J, Li J, Grandillo S, Ahn SN, Yuan L, Tanksley SD, et al. 1998. Identification of trait improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics. 150: 899-909.
- Yang XM, Jiang L, Yang C, Li H, Liu LL, Wan JM. 2014. Semidwarf gene sdk has pleiotropic effects on rice (Oryza sativa L.) plant architecture. Plant Breeding. 133: 335-340.
- Yano K, Ookawa T, Aya K, Ochiai Y, Hirasawa T, Ebitani T, et al. 2015. Isolation of a novel lodging resistance QTL gene involved in strigolactone signaling and its pyramiding with a QTL gene involved in another mechanism. Mol Plant. 8: 303-314.
- Yoon DB, Kang KH, Kim HJ, Ju HG, Kwon SJ, Suh JP, et al. 2006. Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population between Oryza grandiglumis and the O. sativa japonica cultivar Hwaseongbyeo. Theor Appl Genet. 112: 1052-1062.