Abstract
This study aimed to establish a positive selection system for the genetic transformation of cauliflower using the phosphomannose isomerase (PMI) gene. PMI catalyzes the reversible interconversion of mannose 6-phosphate and fructose 6-phosphate. Thus, only plant cells transformed with the PMI gene can survive on synthetic medium containing mannose as a carbon source. Cotyledon explants from 6-day-old seedlings were infected with Agrobacterium tumefaciens strain LBA4404 harboring binary vector pNWB-JMT containing the jasmonic acid carboxyl methyltransferase (JMT) gene with full codon modification. After co-cultivation with Agrobacterium, positive selection was conducted on MS medium supplemented with 2 mg/L of BAP, 1 mg/L of NAA, 0.6% (w/v) mannose, and 2% (w/v) sucrose. Over 40 putative transgenic plants were obtained in the repetitive transformation experiments. The overall transformation efficiency was 1.2% in the mannose-based selection. Polymerase chain reaction along with Southern and Northern blotting analyses were used to confirm and characterize the transgenic plants containing the integrated JMT gene. After soil acclimatization, transgenic plants were successfully grown to maturity in a greenhouse. T1 seeds were successfully collected from these transgenic plants. This is the first successful report of cabbage transformation using a mannose-based selection system. Therefore, the positive selection system established in this study could be applied as an alternative tool for the rapid selection of elite lines for the purposes of breeding and reduction of antibiotic use in order to improve human health and environmental safety.
-
Key words: Molecular biological analysis, Mannose concentration, Transgenic plant, Selectable marker, Transformation
INTRODUCTION
Cabbage (
Brassica oleracea var. capitata), belonging to the family of Brassicaceae, is one of the most widely consumed vegetables worldwide. Similar to other brassicaceous plants, cabbage is an excellent source of fiber, vitamins, and minerals (
Björkman et al. 2011). It also produces antioxidant phenolics with significant free radical scavenging activity, ferric-reducing ability, and capacity to inhibit lipid peroxidation (
Llorach et al. 2003). Moreover, cabbage contains many nutrients preventing a wide range of diseases from cancer to heart diseases (
Kirsh et al. 2007;
Björkman et al. 2011). These health properties are largely attributed to its content of secondary metabolites such as glucosinolates (
Verkerk et al. 2009), which are linked to a reduced incidence of risk for degenerative diseases (
Virgili and Marino 2008;
Björkman et al. 2011).
Agrobacterium-mediated transformation is widely used for gene delivery in plants. Genetic transformation of
Brassica oleracea using
Agrobacterium tumefaciens has been reported by a number of groups (
Srivastava et al. 1988;
De Block et al. 1989;
Metz et al. 1995;
Jin et al. 2000;
Pius et al. 2000;
Tsukazaki et al. 2002;
Bhattacharya et al. 2004;
Sparrow et al. 2004;
Liu et al. 2008). Despite considerable advances in transformation procedures in recent years, the routine transformation of Brassica still suffers from genotype restrictions and low transformation frequencies (
Bhalla and Singh 2008).
Sparrow et al. (2004) reported a positive correlation between tissue culture phenotypes and the transformation efficiencies of B. oleracea. Optimization of transformation procedures is also an alternative for achieving higher transformation efficiencies (
Rafat et al. 2010). However, it is difficult to determine the differences among tissue culture phenotypes as well as achieve the required skill for optimization. Thus, more reproducible and reliable selection methods are required to obtain elite transgenic plants. A recent development is based on the use of selective genes, which provide transformed cells a metabolic advantage compared to untransformed cells that are starved with a concomitant reduction of viability.
To minimize public concerns over the use of genes conferring antibiotic or herbicide resistance, many studies have attempted to develop alternative selection systems. Traditionally, transformed plants are selected using antibiotics-resistant genes, such as neomycin phosphotransferase (nptII) and hygromycin phosphotransferase (hpt), or herbicides-tolerant genes, such as phosphinothricin acetyl transferase (bar) (Bowen 1993). However, many methods have been developed to avoid the use of such antibiotics or herbicides as a selection marker. One alternative is a positive selection system using the phosphomannose isomerase (PMI) gene. The phosphomannose isomerase (PMI, E. C. 5.3. 1.8) enzyme catalyzes the reversible interconversion of mannose-6-phosphate (Man-6-P) and fructose-6-phosphate (Fru-6-P), thereby providing a link between glycolysis and the mannose metabolic pathway in both prokaryotic and eukaryotic organisms (
Stoykova and Stoeva-Popova 2011;
Joersbo et al. 1999). Thus, plant cells lacking this enzyme are incapable of surviving on synthetic medium containing mannose as a carbon source (
Reed et al. 2001). Due to the inherent safety of the mannose selection system, mannose-based selection has already been successfully established for the production of transgenic crops, including rice (
Lucca et al. 2001;
He et al. 2004), maize (
Wang et al. 2000;
Negrotto et al. 2000;
Wright et al. 2001;
Reed et al. 2001), wheat (
Gadaleta et al. 2008), sorghum (
Gao et al. 2005;
Gurel et al. 2009;
Grootboom et al. 2010), onion (
Aswath et al. 2006), Chinese cabbage (
Ku et al. 2006;
Min et al. 2007), cucumber (
He et al. 2006), lettuce (
Briza et al. 2010), rapeseed (
Wallbraun et al. 2009), potato (
Briza et al. 2008), tomato (
Sigareva et al. 2004;
Briza et al. 2008), Arabisopsis (
Todd et al. 2001), tobacco (
Barb et al. 2003), and orchid (
Thiruvengadam et al. 2011). Further, the transformation frequency of the mannose selection system has been shown to be higher than that of a kanamycin selection system (
Joersbo et al. 1998;
Reed et al. 2001).
Methyl jasmonate is a plant volatile that acts as an intracellular regulator, a diffusible intercellular signal transducer, and an airborne signal mediating intra- and interplant communications (
Seo et al. 2001). Jasmonates are critical in the defense responses of herbivory and plant to multiple abiotic and biotic stresses (
Farmer and Ryan 1990). They also control diverse plant developmental processes, such as seed germination, flower, fruit and seed development, senescence, and tuberization of potato (
Sohn et al. 2011). A key enzyme of jasmonate-regulated plant responses is jasmonic acid carboxyl methyltransferase (JMT) (
Seo et al. 2001). Therefore, it is expected that introduction of the JMT gene into crops through genetic transformation could be useful for overcoming multiple abiotic and biotic stresses. Recently, a few studies attempted to explore the role of JMT in crops (
Sohn et al. 2011;
Stitz et al. 2011). In a previous study, we already confirmed that the PMI gene isolated from Escherichia coli is a good candidate as a positive selection marker in the transformation of Chinese cabbage (
Min et al. 2007). Despite the horticultural and medicinal importance of cabbage, however, genetic transformation of cabbage has not been reported using a mannose selection system. Thus, this study aimed to introduce the JMT gene into cabbage via
Agrobacterium using a PMI/mannose selection system for the development of a stress-resistant cultivar.
MATERIAL AND METHODS
Plant material
To establish the genetic transformation system of cabbage (
Brassica oleracea L. var. capitata), inbred cabbage seeds (AD BENTAM) were purchased from Nong Woo Bio seed Company. Seeds were surface-sterilized with 70% ethanol for 1 min and then disinfected for 15 min with 2% sodium hypochlorite solution. Seeds were then washed more than three times with sterilized water. After the removal of remaining moisture with filter paper, surface-sterilized seeds were transferred onto half-strength MS (
Murashige and Skoog 1962) basal medium without any plant growth regulators. For germination, cultures were kept at 25°C in the dark for 5 days. Unless mentioned otherwise, all cultures were maintained at 25°C in the dark.
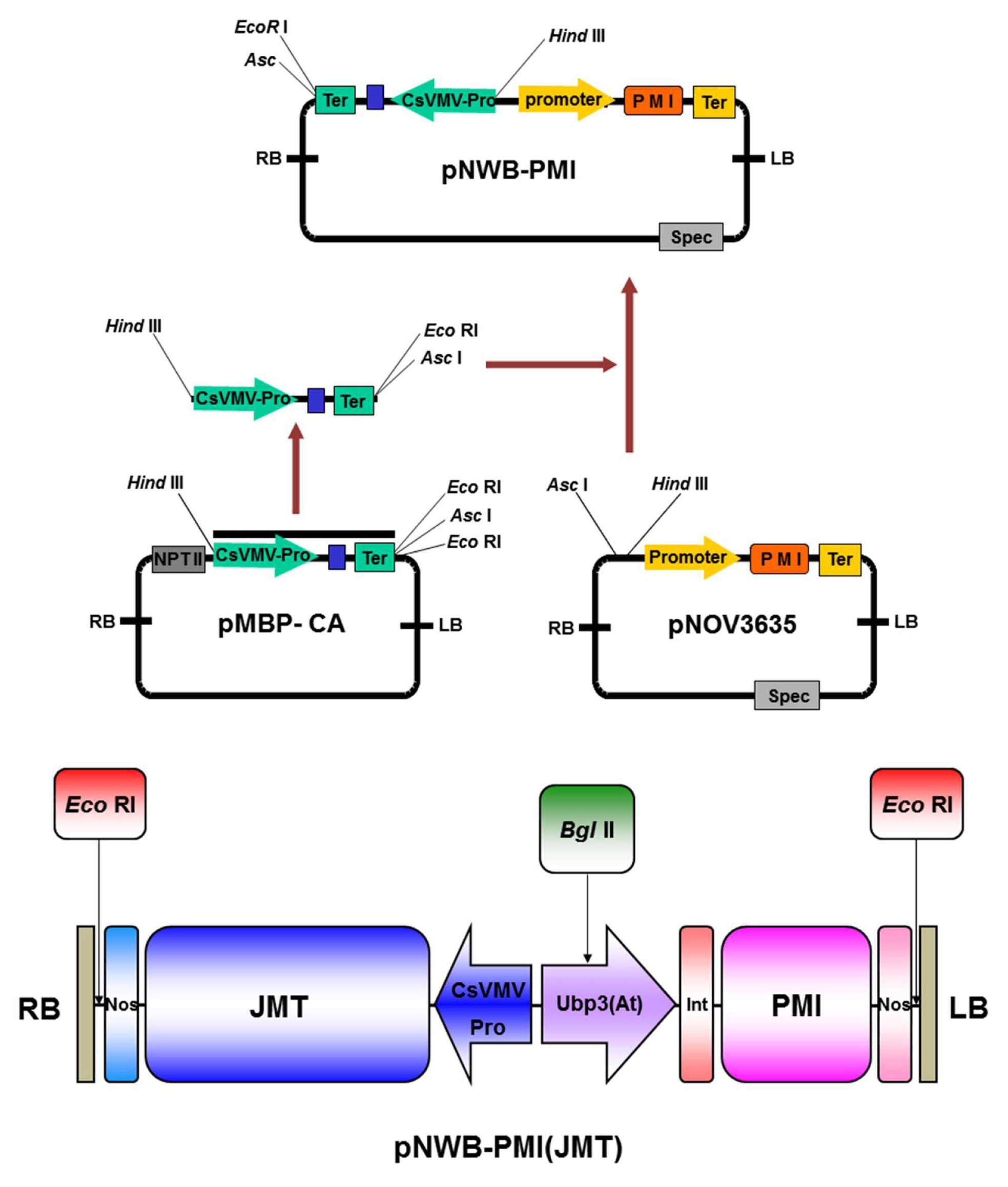
Agrobacterium tumefaciens strain LBA4404 harboring binary vector pNWB-JMT, which contains the JMT and PMI genes, was routinely used in our laboratory for genetic transformation. Full-length JMT cDNA (
Seo et al. 2001) in sense orientation was isolated from
Arabidopsis using RT-PCR (reverse transcription polymerase chain reaction) and then cloned into pMBP-1 vector. By cloning this gene into the
HindIII and
AscI sites of pNOV3635 linked to the PMI gene (provided by Syngenta) of
Escherichia coli, vector pNWB-JMT could be developed. In this vector, the JMT gene is controlled using the CsVMV (cassava mosaic virus) 35S promoter and nopaline synthase terminator (
Fig. 1).
Determination of mannose concentration for positive selection
Determining the suitable concentration of mannose is very important for selecting transformed plants. First, cotyledons and hypocotyls from 5-day-old seedlings were cut and planted on shoot induction medium consisting of MS salts supplemented with 2 mg/L of BAP and 1 mg/L of NAA. To determine the most suitable concentration of mannose for positive selection, cotyledon and hypocotyl explants were cultured onto shoot induction medium supplemented with 0–10 g/L of mannose. To increase the selection pressure, sucrose was simultaneously supplied to shoot induction medium containing mannose at several different concentrations (0, 10, 20, 30 g/L), respectively. For each treatment, 10 explants (total 2640 explants) were examined with three replicates. For determination of shoot formation frequency, the number of explants with newly formed shoots was counted after 4 weeks of incubation in the light.
Transformation and selection of transformants
A single cell colony of Agrobacteria was grown in 4 ml of YEP liquid medium containing 50 mg/L of kanamycin for 12 h, after which 2 ml was transferred to an additional 50 ml of YEP liquid medium containing 50 mg/L of kanamycin for another 12 h at 28°C. Agrobacteria were cultured in YEP medium until the OD value reached between 0.8 to 1.0. The bacterial cells were pelleted by centrifugation (4000 rpm, 15 min), after which the pellet was dissolved in liquid shoot induction medium containing 100 μmol of acetosyringone. Cotyledons (about 5 mm2) and hypocotyls (10 mm in length) from 5-day-old seedlings of cauliflower were cut into segments on a clean bench. The explants were then co-cultured in liquid shoot induction medium for 2 days in the dark at 21°C. After co-cultivation, the explants were rinsed with liquid shoot induction medium supplemented with 500 mg/L of cefatoxime and cultivated on solid shoot induction medium supplemented with 1 mg/L of AgNo3, 200 mg/L of lilacillin, 20 g/L of sucrose, and 6 g/L of mannose (mannose selection medium). After 6 to 8 weeks of incubation, adventitious shoots had formed on the cut edges of explants. The regenerated shoots were then cut and subcultured onto fresh mannose selection medium. Well grown shoots were transferred to MS medium supplemented with 2 mg/L of NAA for root induction. Successfully rooted plantlets were transplanted into potting soil (vermiculite: perlite, 3:1 mixture) and maintained in a plastic container for 2 weeks to acclimate. After the emergence of new leaves from transplanted plants, plants were transferred and maintained in a growth chamber (25°C day/22°C night, 80 μmol m−2s−1 from cool white fluorescent lamps, a 16 h photoperiod, and 50–70% RH).
PCR and Southern blot analysis of transformants
In general, PCR is used to confirm the presence or absence of transgenes in primary transformants (
Pastori et al. 2001). To confirm the presence of the transgene, total genomic DNAs of leaves from putative transgenic and untransformed wild-type cauliflowers were extracted according to a modified method from
McGarvey and Kaper (1991). PCR was carried out using specific forward/reverse primers designed for the
JMT gene. The gene-specific primers used for genomic PCR were as follows: 5′-GAGATGGAGGTAATGCGAGTTCTT-3 (sense) and 5′-ACACGATCAACCGGTTCTAACGAG-3 (antisense). After denaturation for 5 min at 94°C, samples were subjected to 35 cycles of PCR under the following conditions: 94°C for 1 min, 55°C for 1 min, and 72°C for 1 min. At least two replicates were carried out for each PCR analysis. The PCR product obtained through these processes was analyzed by 0.8% agarose gel electrophoresis.
For Southern blot analysis, genomic DNA was isolated from leaves according to the method of
Cao et al. (1992). Exactly 10 μg of genomic DNA was digested overnight with
BamHI restriction enzyme (Roche Diagnostics GmbH, Germany), followed by fractionation on 0.8% agarose gel in 1X TAE buffer for 12 h at 60 V and blotting onto Hybond-N
+ membranes (Amersham, USA). Hybridization was performed in hybridization reagents (1X HSB, 0.23 Denhardt’s III, 0.05 mg ml
−1 of salmon sperm DNA) at 65°C for 16 h using the full JMT gene as a probe, which was labeled with
32P-dCTP by a random priming method (Promega, USA). Following hybridization, the membranes were washed twice at low stringency (2X SSC and 0.5% SDS) and then twice at high stringency (0.2X SSC and 0.5% SDS). All washes were done for 15 min at 65°C. The membrane was then exposed to a storage phosphor screen for 2 to 3 days, and image data were obtained on a Molecular Dynamics Phosphor Imager (Amersham Inc, USA).
For Northern blot hybridization, isolation of total RNA from the young leaves of transformed cabbage plants was performed according to the method of
Logemann et al. (1987). Electrophoresis was performed in 1.2% formaldehyde agarose gel, after which the full JMT gene was labeled with
32P-dCTP and then used as a probe according to the method reported by
Sambrook et al. (1989). As a positive control r-RNA was used. Hybridization was performed using 50% formamide, 5 X SSC, 0.05 M phosphate buffer, and Denhardt’s solution at 42°C. The membrane was then exposed to a storage phosphor screen for 2–3 days, and image data were obtained on a Molecular Dynamics Phosphor Imager (Amersham Inc, USA).
Student’s t-test and one-way analysis of variance (ANOVA) was performed to compare the effects of different concentrations of mannose and sucrose on selection efficiency as well as the frequency of shoot formation from cotyledon and hypocotyl explants (total 2640 explants) using Origin (ver 9.0). Significance (P-value) was evaluated at the 0.05 level for all comparisons. Each experiment was conducted with three replicates. For each treatment, the standard deviation (SD) was calculated based on at least three biological replicates.
RESULTS
Determination of mannose and sucrose concentrations for positive selection
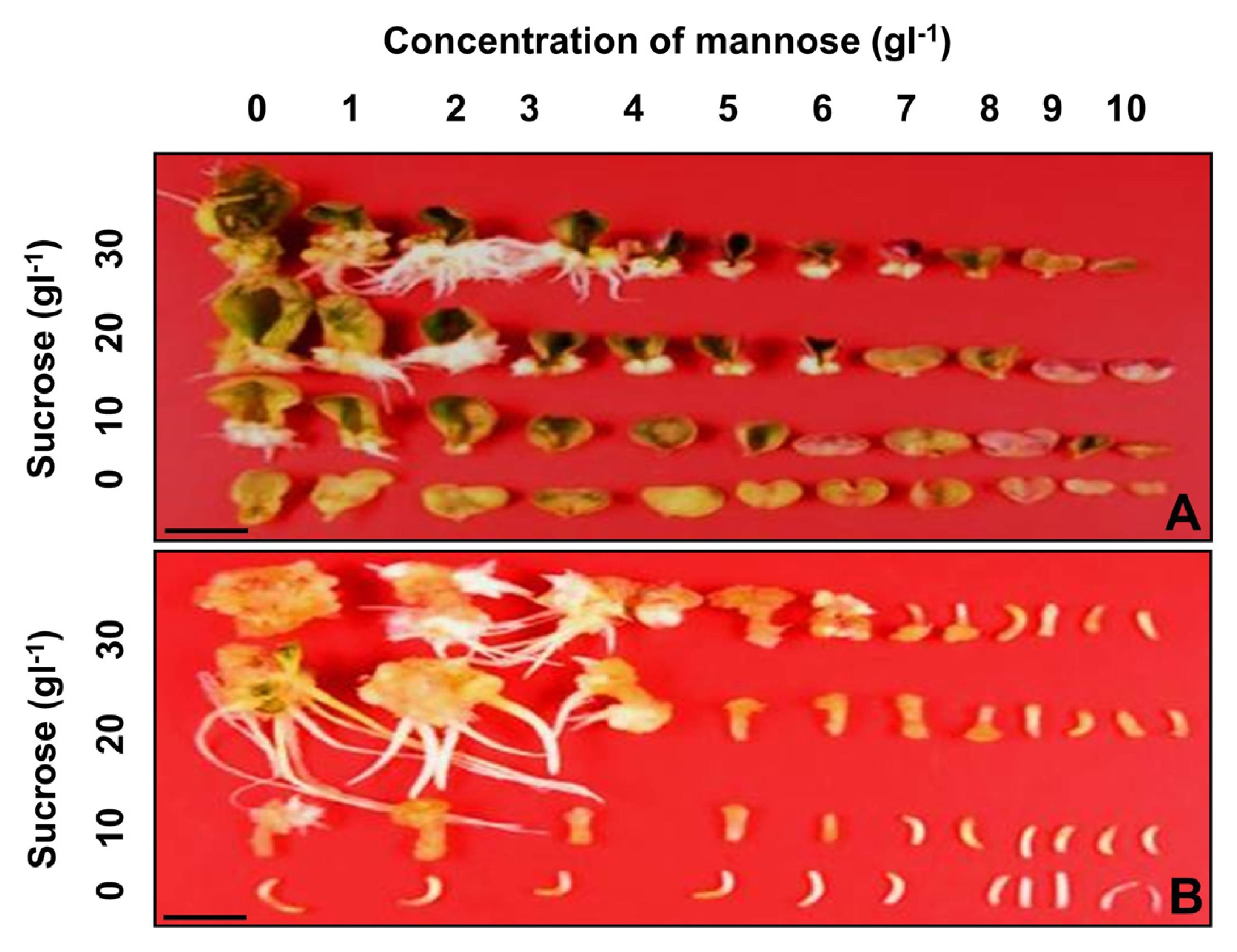
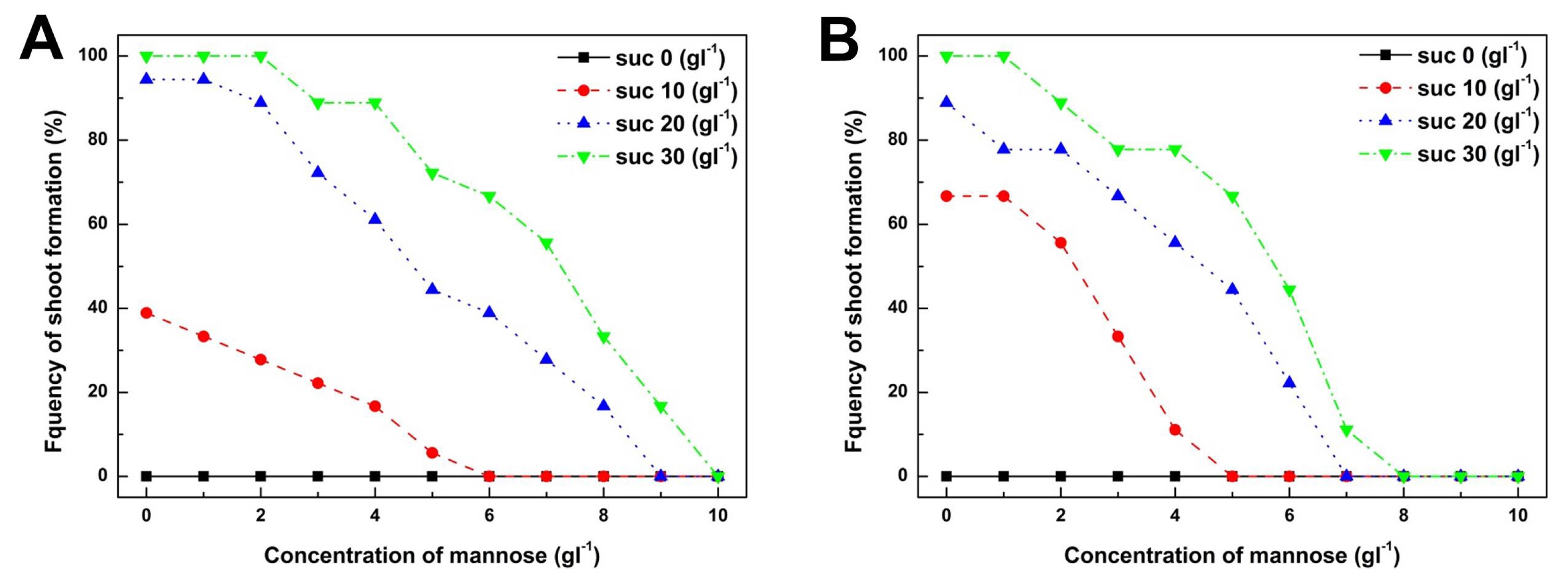
To determine the suitable concentrations of mannose and sucrose for positive selection of cabbage transgenic plants, the frequency of shoot formation was examined from cotyledon and hypocotyl explants cultured on shoot induction medium supplemented with various combinations of mannose and sucrose (
Figs. 2,
3). Green adventitious shoots and white calluses were formed on the cut surfaces of cotyledon explants cultured on shoot induction medium without mannose (
Fig. 2A). However, the frequency of shoot formation from cotyledon explants was reduced when the concentration of mannose was increased from 0 to 10 g/L (
Fig. 3A). At a mannose concentration greater than 6 g/L, the frequency of shoot formation from explants decreased almost to zero when cultured on shoot induction medium supplemented with less than 10 g/L of sucrose (
Fig. 3A). However, cotyledon explants cultured on shoot induction medium containing 30 g/L of sucrose were able to form shoots at a frequency of 16.7% even at a mannose concentration of 9 g/L. These results show that a higher concentration of carbon sources reversed the inhibitory effect of mannose on shoot formation. Furthermore, we observed that inhibition of shoot formation due to mannose could be potentiated by reducing the amount of sucrose in the shoot induction medium. These results imply that shoot formation from cotyledon explants sharply decreased when a suitable concentration of carbon source was not supplied to the shoot induction medium. Therefore, we suggest that a suitable supply of carbon source for shoot formation is the primary requirement to establish a mannose-based positive selection system.
The effect of mannose concentration on shoot formation from hypocotyl explants was very similar compared to shoot formation from cotyledons (
Figs. 2B,
3B). Green adventitious shoots and white calluses were formed on the cut surfaces of hypocotyl explants cultured on shoot induction medium without mannose (
Fig. 2B). Inhibition of shoot formation from hypocotyl explants was much greater than that of cotyledon explants (
Fig. 3B). At a mannose concentration greater than 8 g/L, shoot formation from hypocotyl explants had completely disappeared regardless of whether or not a higher concentration of sucrose was supplemented into the shoot induction medium (
Fig. 3B). These results imply that hypocotyl explants of cabbage were more susceptible to mannose toxicity compared to cotyledon explants. Furthermore, these results show that mannose could be used as a positive selection agent from hypocotyl explants in the same manner as cotyledon explants of cabbage. In conclusion, our combined results on cotyledon and hypocotyl explants of cabbage show that combinational treatment of sucrose at 20 g/L and mannose at 6 g/L was the most optimal to establish a positive selection system for cabbage transformation.
A transgenic plant regeneration system based on mannose selection from hypocotyl explants of cabbage was established (
Fig. 4). Green calluses were formed on the cut edges of hypocotyl explants after 2 weeks of incubation on shoot induction medium containing 6 g/L of mannose and 20 g/L of sucrose under light conditions (
Fig. 4A). After an additional 2 to 3 weeks of incubation without subculture, green globular structures had formed on the green calluses (
Fig. 4B). These hypocotyl explants with green calluses and globular structures were transferred to fresh mannose selection medium and then subcultured at 2-week intervals. Non-transformed calluses and shoots gradually turned brown after 2 to 3 weeks of incubation in the same mannose selection medium. After the second round of mannose selection, several small shoots had begun to form onto the green calluses and globular structures (
Fig. 4C). Non-transformed calluses and shoots turned dark brown and degenerated into death at this stage. These results clearly show that growth of calluses and shoots was strongly suppressed at a higher mannose concentration of greater than 6 g/L, resulting in browning and withering. These results also represent that mannose could be used as a strong selection reagent to remove non-transformed plants. To accelerate plant growth after mannose selection, newly formed shoots were transferred to selection medium without mannose. After 2 weeks of incubation, these green shoots had turned dark green and leaves had expanded (
Fig. 4D). Well grown shoots were transferred to MS medium supplemented with 2 mg/L of NAA for root induction. Most shoots rooted within 2 weeks of incubation. The successfully rooted plantlets were then transplanted into potting soil (vermiculite: perlite, 3:1 mixture) and maintained in a plastic container for 2 weeks to acclimate (
Fig. 4E). After the emergence of new leaves, transgenic plants were transferred and maintained in a growth chamber (25°C day/22°C night, 80 μmol m
−2s
−1 from cool white fluorescent lamps, a 16 h photoperiod, and 50–70% RH) (
Fig. 4F). Although most transformed plants showed normal phenotypes, 2–3% showed abnormal phenotypes such as longer stigma. This could be attributed to somaclonal variations during the tissue culture. These transgenic plants grew into adult plants without any severe morphological abnormality (
Fig. 4G).
In summary, 57 transgenic plants were obtained from hypocotyl transformation experiments, representing an overall transformation efficiency of 1.2% (
Table 1). To our knowledge, this is the first successful report of whole plant regeneration of transgenic cabbage via a mannose-based positive selection system.
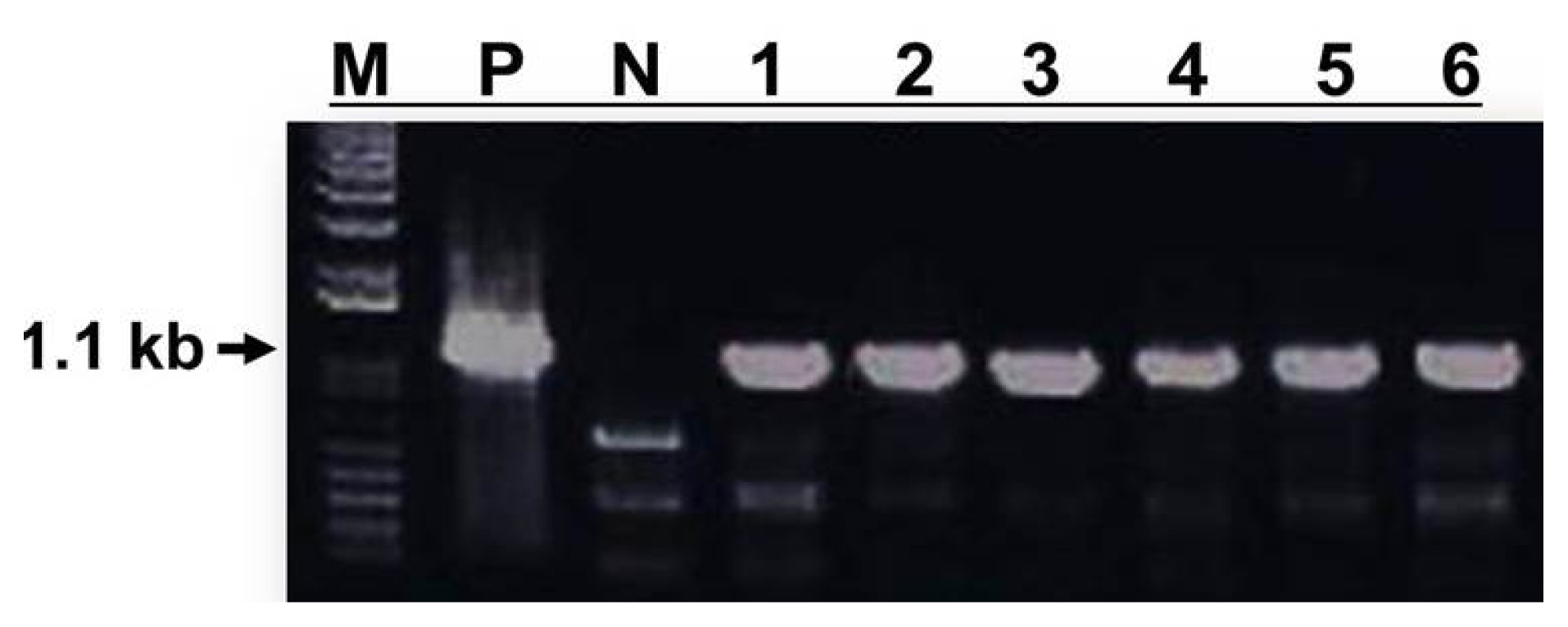
The JMT gene was introduced into cabbage as a transgene in this study. To confirm introduction of the JMT gene into the genome of cabbage, PCR (
Fig. 5), Southern (
Fig. 6), and Northern blot analyses (
Fig. 7) were performed using genomic DNA isolated from the leaves of transformed cabbage plants. To confirm the presence of the target transgene in the putative transgenic plants, the T
0 plants were subjected to PCR analysis with primers specific for the JMT gene (
Fig. 5). All putative transgenic plant lines showed the expected 1.1 kb coding fragment of the
JMT gene following agarose gel electrophoresis of PCR products (
Fig. 5), whereas non-transformed lines did not. These results clearly show that the mannose-based selection system worked efficiently as a selection tool.
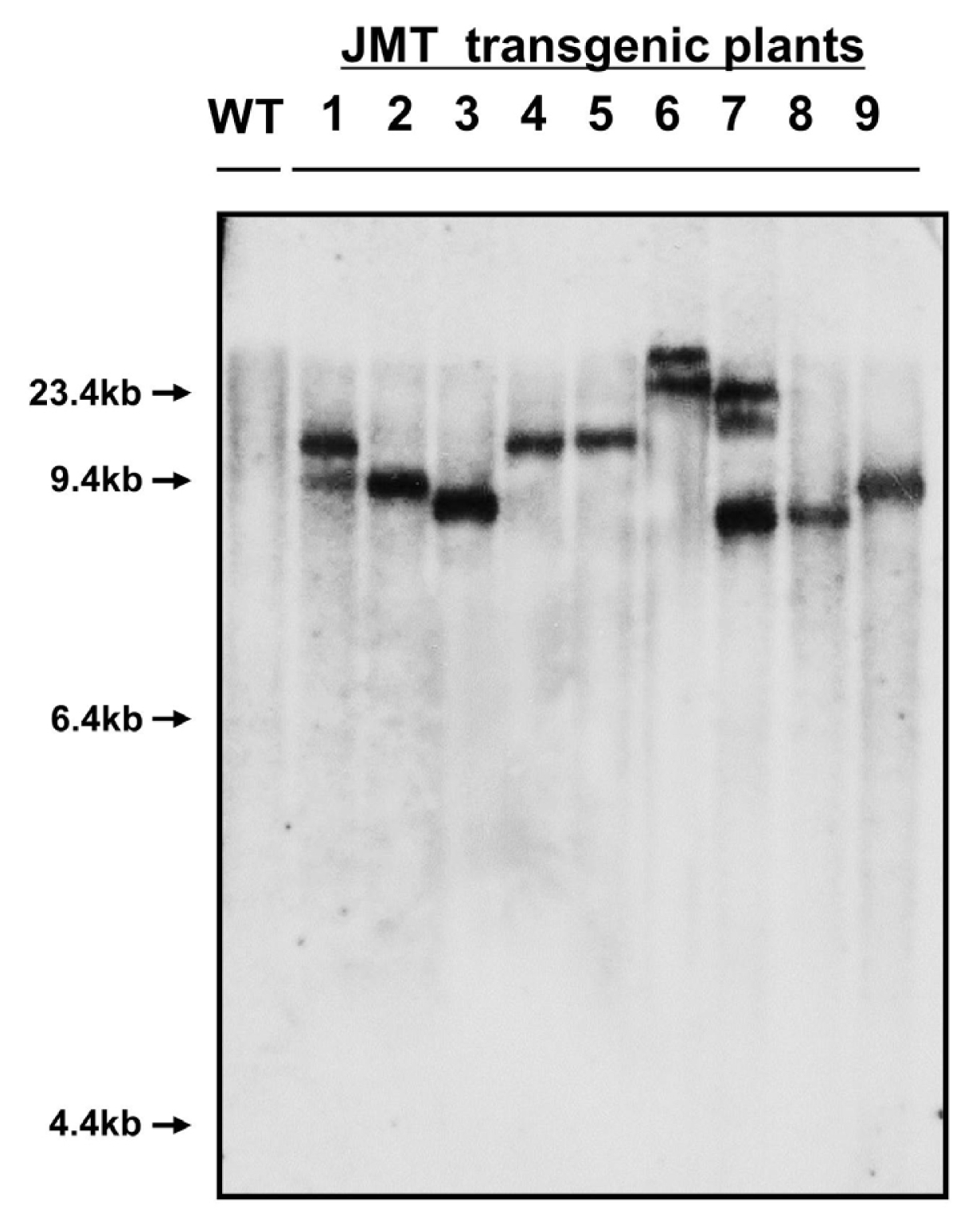
Southern blot analysis was also performed to confirm the presence of the JMT gene in the genomes of transgenic plants chosen as mannose-resistant regenerants (
Fig. 6). Genomic DNA was cut by treatment with
BamHI restriction enzyme. Southern hybridization of restricted DNA products from transgenic plants using a JMT gene-specific cDNA probe confirmed stable integration of the JMT gene. Southern hybridization for the JMT gene was performed on nine PCR-positive plants using a non-transformed plant as a control. As expected, PCR-positive plant lines gave hybridization signals, whereas no signal was observed in the non-transformed control plant. The number of hybridizing bands varied from one to two copys, thereby reflecting the number of inserted loci of the transgene in the plant genome (
Fig. 6). Southern blot analysis also showed a consistent integration pattern of the transgene in cabbage plants.
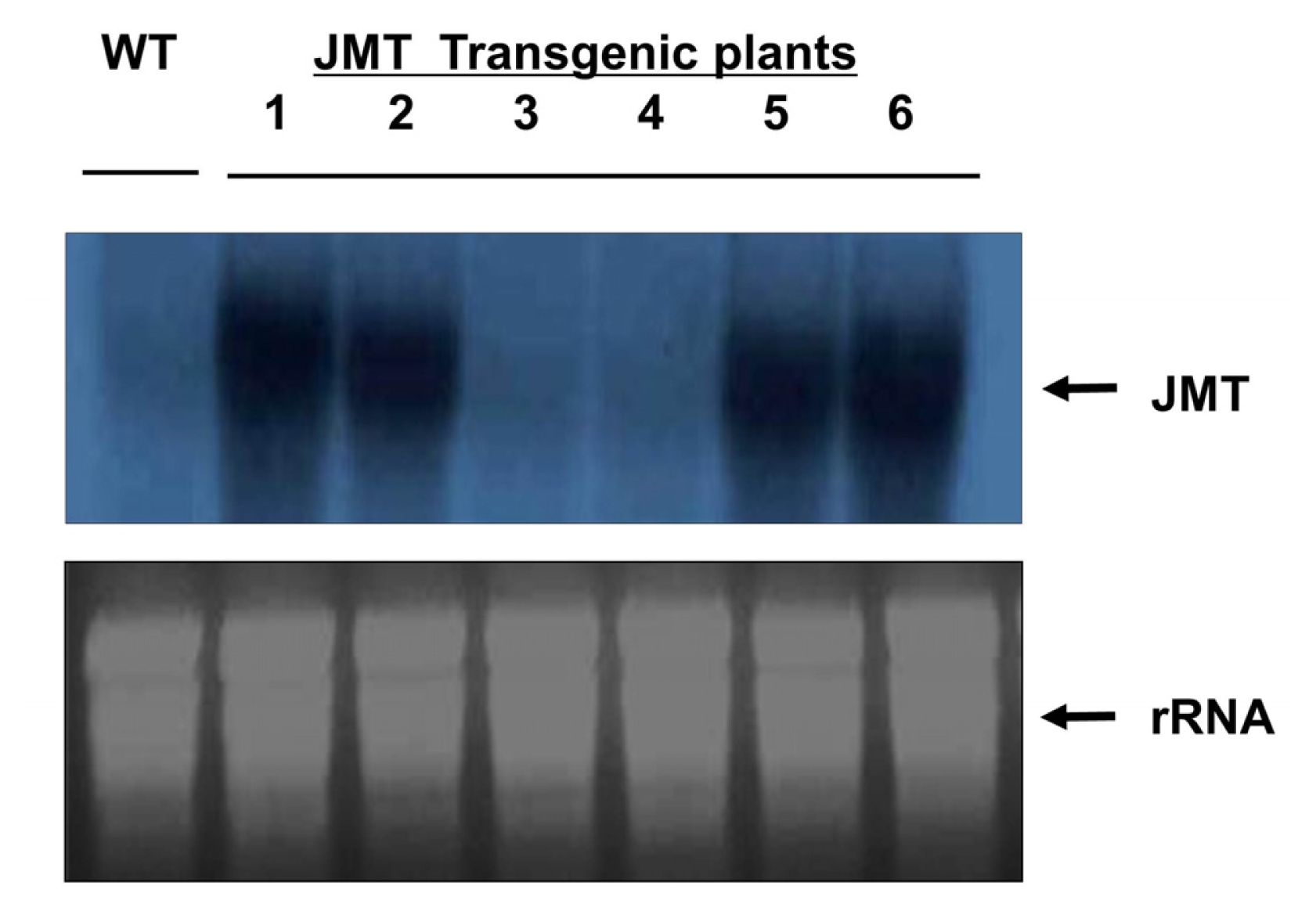
To investigate whether or not the JMT gene introduced into cabbage was expressed, transgene expression was analyzed by Northern blot analysis (
Fig. 7). Northern analysis showed that the JMT gene was stably transcribed in four transgenic plant lines. No signal was detected in either of the transgenic plant lines or the non-transformed control plant. These results demonstrate that the JMT gene was successfully introduced and expressed in transgenic cabbage.
The T1 seed segregation ratio was also examined. T0 cabbages expressing the JMT gene were self-fertilized, and T1 seeds produced from the self-fertilization were planted. To determine the segregation ratio of T1 plants, DNA was isolated from leaves and PCR analysis performed using JMT gene-specific primers. As expected, the segregation ratio between the transformed and non-transformed plants was 3:1 (33:12), which was consistent with that of Mendel (data not shown). This result also showed that there is only one copy of the JMT gene in transformed plants of the T1 generation.
In conclusion, we performed the genetic transformation of cabbage using a mannose selection marker. The positive selection system established in this study could be applied as an alternative to reduce antibiotics for human health and environmental safety during the genetic transformation of higher plants. At present, we plan to examine the stress resistance of transgenic cabbages.
DISCUSSION
Determination of mannose and sucrose concentrations for positive selection
Sucrose is the most representative chemical used as a carbon source in plant tissue culture. In this study, we found that there was a positive correlation between the concentration of sucrose in the shoot induction medium and the frequency of shoot formation from hypocotyl explants. It could be inferred that a higher concentration of sucrose had a positive role in reducing the toxicity of mannose in shoot formation. This result was similar to a previous report on Chinese cabbage exhibiting reduced mannose toxicity in shoot formation in the presence of a higher concentration of sucrose (
Min et al. 2007). Furthermore, these results were in good agreement with many previous studies regarding the requirement for a carbon source in a mannose selection system (
Joersbo et al. 1998;
Sigareva et al. 2004; O’Kennedy
et al. 2004; Zhu
et al. 2005;
Aswath et al. 2006;
Min et al. 2007). Therefore, we suggest that both determination of a suitable concentration of carbon source as well as a toxic concentration of mannose are simultaneously required to establish an efficient mannose selection system in cabbage.
In comparison to previous studies, the suitable mannose concentration determined in this work was lower than that of other crops (
Joersbo et al. 1998;
Negrotto et al. 2000;
Wright et al. 2001;
Sigareva et al. 2004; O’Kennedy
et al. 2004; Zhu
et al. 2005;
Aswath et al. 2006;
He et al. 2006;
Min et al. 2007). We could not elucidate why cruciferous plants such as cabbage were more sensitive to mannose toxicity in this study. However, it could be inferred that the rigidity or firmness of explants during tissue culture, including shoot organogenesis, caused absorbable differences in mannose intake or inherent differences in mannose sensitivity.
Generally, cabbage is known to be difficult in genetic transformation. As observed during the preliminary experiment of this study, its transformation rate was less than 1% when transformed using kanamycin as a selection reagent (data not shown). In comparison to previous studies reporting genetic transformation of cabbage using antibiotic selection markers (
Tsukazaki et al. 2002;
Bhalla and Singh 2008;
Liu et al. 2008;
Rafat et al. 2010), the transformation efficiency shown in this study was much lower. Moreover, the transformation efficiency of cabbage was slightly lower than those of tomato (
Sigareva et al. 2004), cucumber (
He et al. 2006), sweet orange (
Boscariol et al. 2003), and monocot plants (
Reed et al. 2001), whereas it was similar to that (1.4%) of chinese cabbage belonging to the same cruciferous family using the same mannose selection system (
Ku et al. 2006;
Min et al. 2007). To elucidate the exact mechanism behind reduction of transformation efficiency in cabbage, further studies should be performed. Previously, the overall transformation efficiency of cabbage was shown to be below 1% even though an antibiotic marker was used for selection (
Jin et al. 2000). Therefore, reduction of transformation efficiency can be attributed to the relative difference among genotype-independent regeneration potentials in cabbage and not entirely to mannose toxicity. Further studies are needed to increase the transformation efficiency based on the mannose selection system. Therefore, considering the safety problems with antibiotics, the mannose-based selection system established in this study could be applied as a useful tool for genetic improvement of cabbage.
Fig. 1Schematic map of pNWB-PMI(JMT) binary vector. The cloned PCR fragment was subcloned into the AscI and HindIII sites of pNOV3635 harboring the E. coli PMI gene (provided by Syngenta). The JMT gene was placed under the control of the constitutive cassava mosaic virus 35S promoter and a 3′ nopaline synthase terminator. The pNWB-PMI(JMT) plasmid was used for Agrobacterium-mediated transformation of cabbage. RB: right border, LB: left border
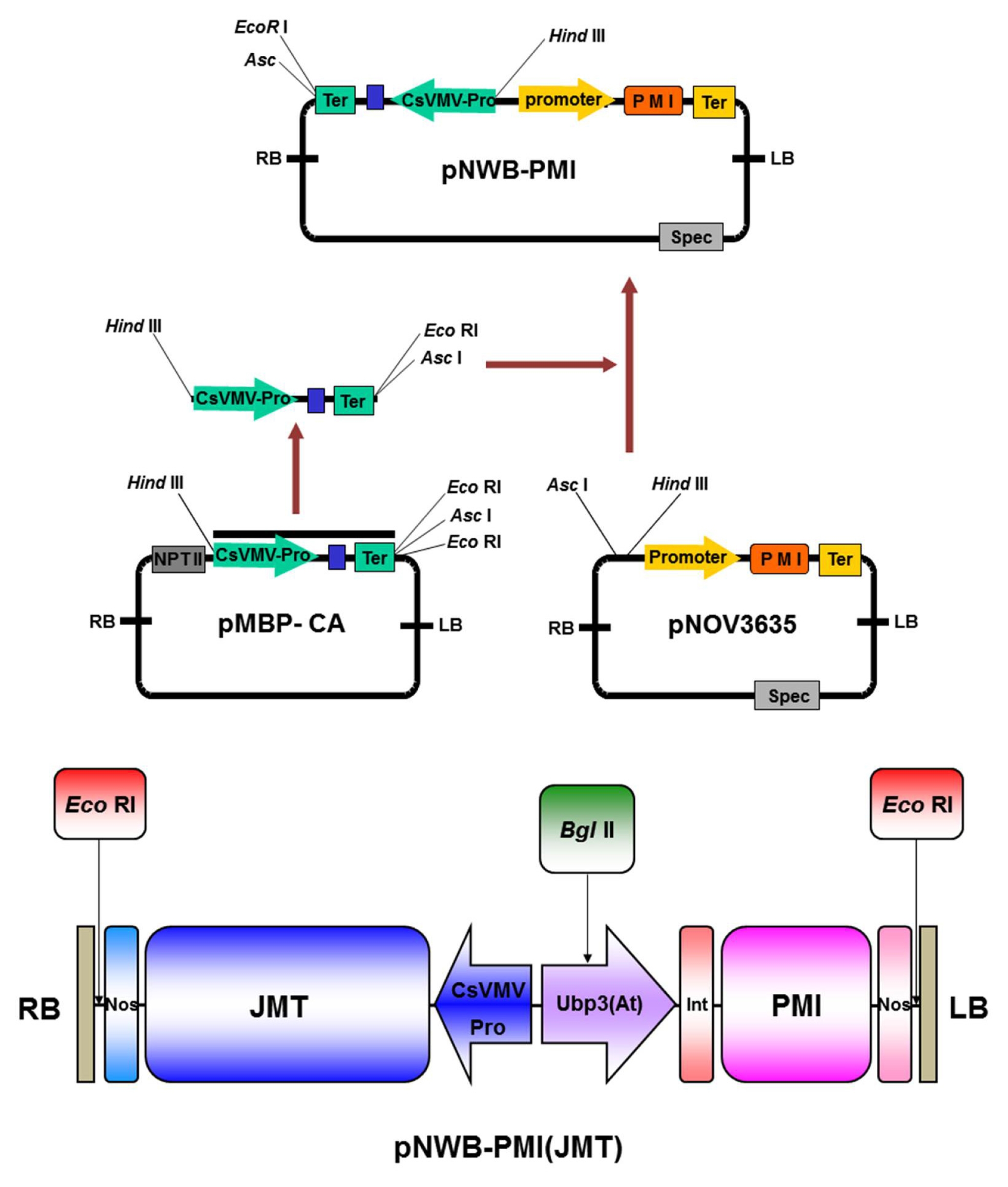
Fig. 2Effects of mannose and sucrose concentrations on shoot formation from cotyledon (A) and hypocotyl explants (B) of cabbage. Each explant was cultured on shoot induction medium supplemented with 0–10 g/L of mannose and several different concentrations (0, 10, 20, 30 g/L), respectively. Scale bars represent 10 mm (A, B).
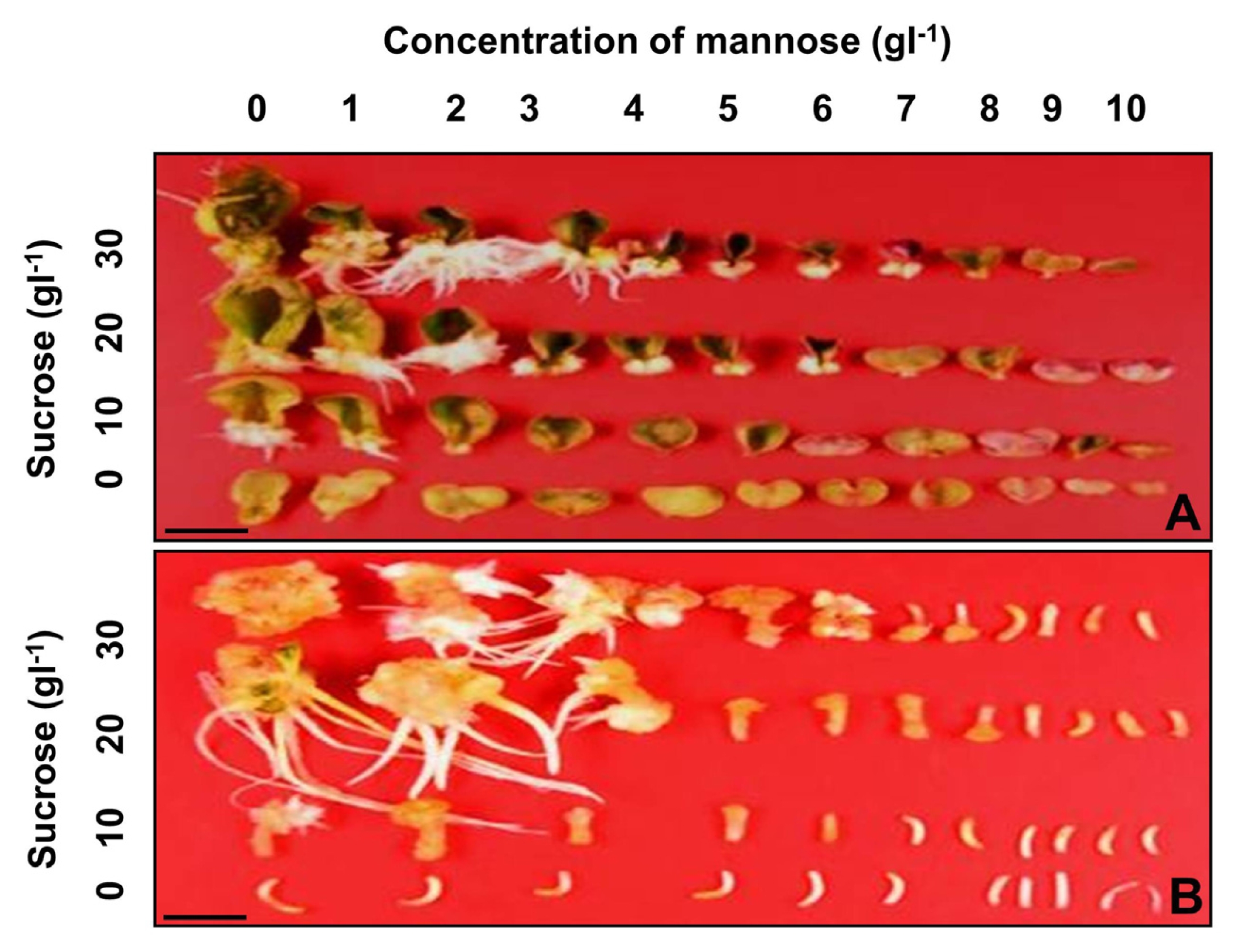
Fig. 3Effects of mannose and sucrose concentrations on the frequency of shoot formation from cotyledon (A) and hypocotyl explants (B) of cabbage. Each explant was cultured on shoot induction medium supplemented with 0–10 g/L of mannose and several different concentrations (0, 10, 20, 30 g/L), respectively. Each symbol represents the concentration of sucrose.
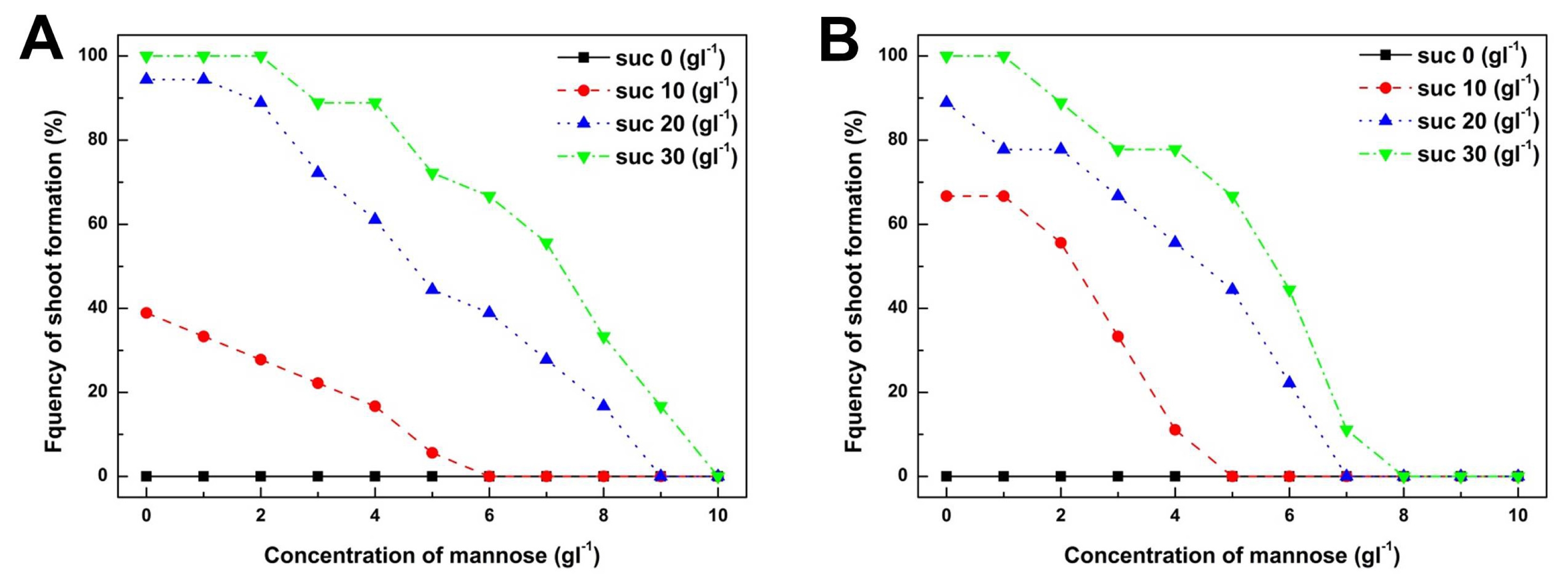
Fig. 4Transgenic plant regeneration from hypocotyls of cabbage. A: Green callus formation from hypocotyls after 2 weeks of incubation on mannose selection medium. B: Green globular structure formation from green calluses after 4 weeks of incubation on mannose selection medium. C: Shoot development from green globular structures after 6 weeks of incubation. D: Shoot elongation after 6 weeks of incubation. E: Soil transfer and acclimation of plantlets after 2 months of incubation. F: Young plant regeneration. G: Mature plants regenerated from hypocotyls of cabbage in a greenhouse. Scale bars represent 5 mm (A–D), 0.2 cm (E–F), and 10 cm (G).

Fig. 5PCR amplification of 1.1 kb JMT gene fragments from genomic DNA of mannose-resistant plants transformed with pNWB-PMI(JMT) binary vector. Capital letters and numbers represent each sample. M: 1 kb DNA ladder, P: Agrobacterium tumefaciens strain LBA4404 harboring pNWB-PMI(JMT) vector as a positive control. N: Non-transformed wild type plant as a negative control. 1–6: Independent mannose-resistant transformed plants.
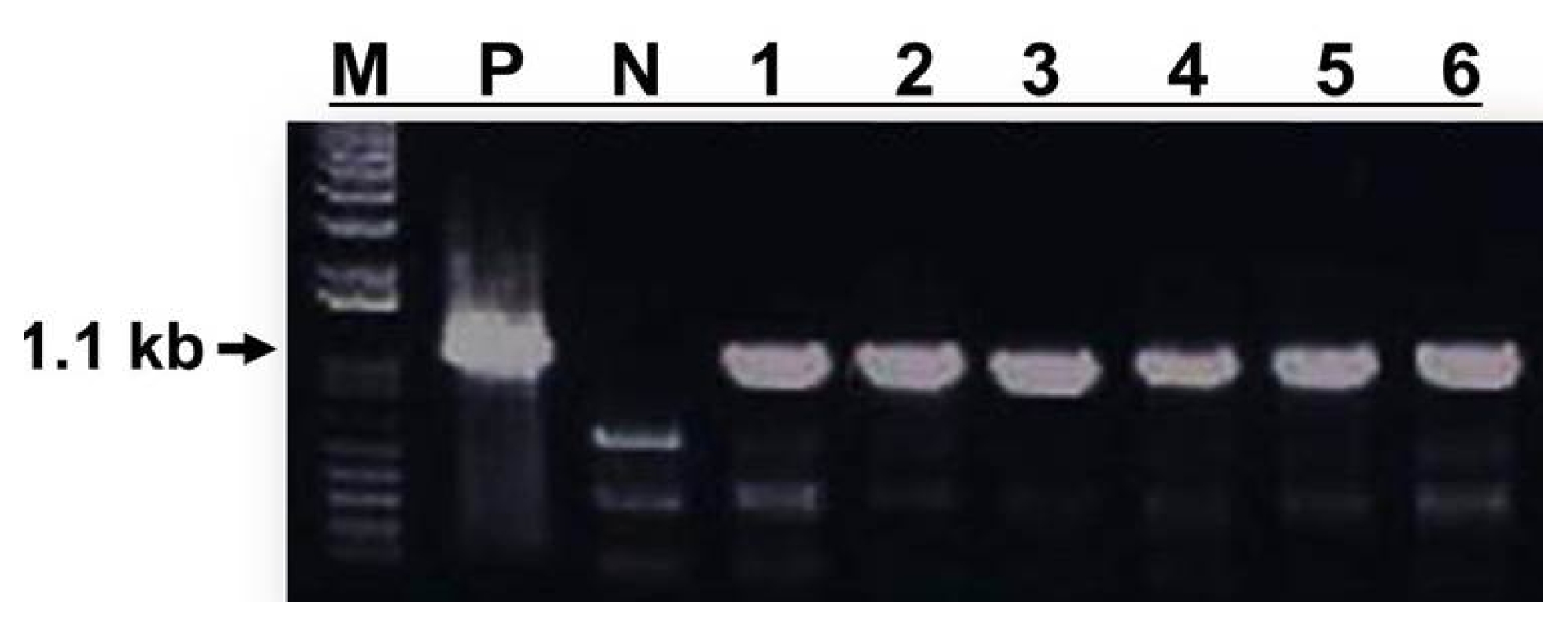
Fig. 6Southern blot analysis of cabbage transformed by the JMT gene. Ten micrograms of genomic DNA was digested with BamHI restriction enzyme and then blotted on a nylon membrane. The blot was hybridized with the 1.1 kb JMT coding region labeled with 32P-dCTP as a probe. Capital letters and numbers represent each sample. WT: Non-transformed wild type plant as a negative control. 1–9: Independent JMT transformed plants.
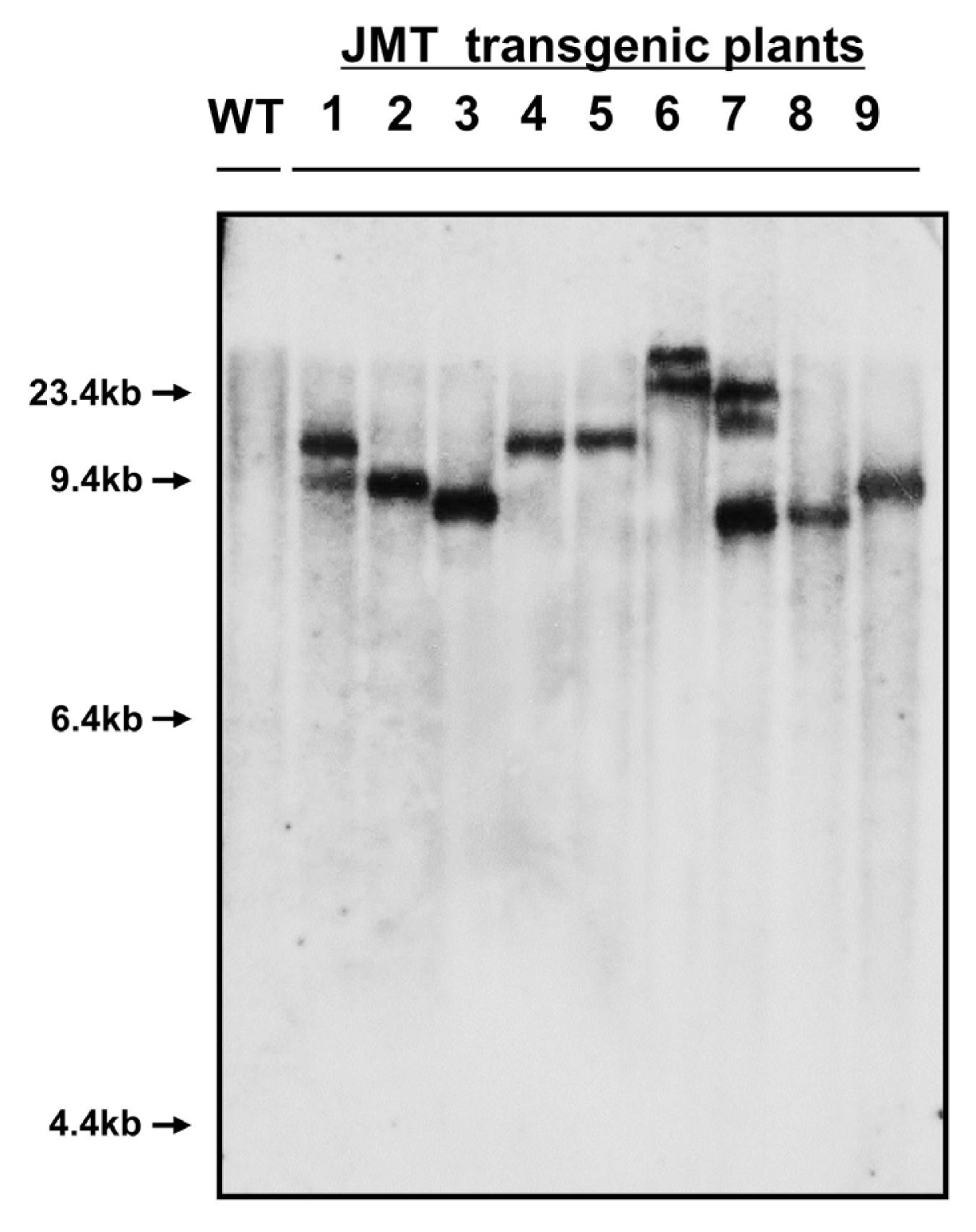
Fig. 7RNA gel blot analysis of transgenic cabbage plants. In each lane, 2 μg of mRNA was fractionated on a formaldehyde-containing agarose gel and blotted onto a nylon membrane. The membrane was then hybridized with a 32P-labeled 1.1 kb JMT fragment. Capital letters and numbers represent each sample. WT: Non-transformed wild type plant as a negative control. 1–6: Independent JMT transformed plants
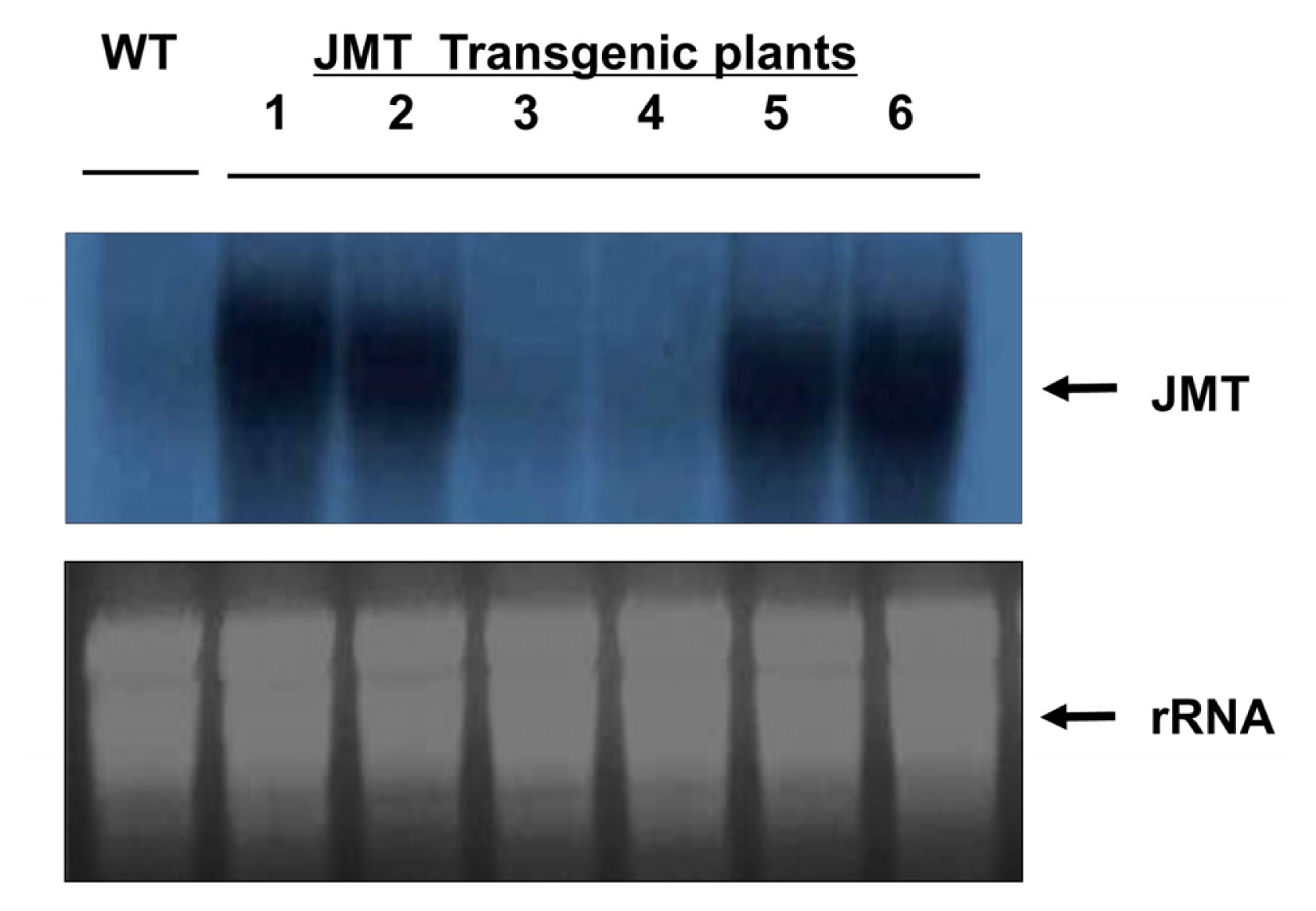
Table 1Transformation efficiency of cabbage by the mannose selection system.
Table 1
|
Target gene |
No. of explants tested |
No. of transformed plants |
Transformation efficiency (%) |
|
JMT |
4723 |
57 |
1.2 |
References
- Aswath CR, Mo SY, Kim DH, Park SW. 2006. Agrobacterium and biolistic transformation of onion using non-antibiotic selection marker phosphomannose isomerase. Plant Cell Rep. 25: 92-99.
- Barb AW, Pharr DM, Williamson JD. 2003. A Nicotiana tabacum cell culture selected for accelerated growth on mannose has increased expression of phosphomannose isomerase. Plant Sci. 165: 639-648.
- Bhalla PL, Singh MB. 2008. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat Protoc. 3: 181-189.
- Bhattacharya RC, Maheswari M, Dineshkumar V, Kirti PB, Bhat SR, Chopra VL. 2004. Transformation of Brassica oleracea var. capitata with bacterial betA gene enhances tolerance to salt stress. Sci Hortic. 100: 215-227.
- Björkman M, Klingen I, Birch ANE, Bones AM, Bruce TJA, Johansen TJ, Meadow R, Mølmann J, Seljasen R, Smart LE, Stewart D. 2011. Phytochemicals of Brassicaceae in plant protection and human health - Influences of climate environment and agronomic practice. Phytochemistry. 72: 538-556.
- Boscariol RL, Almeida WAB, Derbyshire MTVC, Mourao Filho FAA, Mendes BMJ. 2003. The use of the PMI/mannose selection system to recover transgenic sweet orange plants. Plant Cell Rep. 22: 122-128.
- Briza J, Pavingerova D, Prikrylova P, Gazdova J, Vlasak J, Niedermeierova H. 2008. Use of phosphomannose isomerase-based selection system for Agrobacterium-mediated transformation of tomato and potato. Biol Plant. 52: 453-461.
- Briza J, Ruzickova N, Niedermeierova H, Dusbabkova J, Vlasak J. 2010. Phosphomannose isomerase gene for selection in lettuce (Lactuca sativa L.) transformation. Acta Biochim Pol. 57: 63-68.
- Cao J, Duan X, McElroy D, Wu R. 1992. Regeneration of herbicide resistant transgenic rice plants following microprojectile mediated transformation of suspension culture cells. Plant Cell Rep. 11: 586-591.
- De Block M, Tenning P, De Brouwer D. 1989. Transformation of Brassica napus and Brassica oleracea using Agrobacterium tumefaciens and the expression of the bar and neo genes in the transgenic plants. Plant Physiol. 91: 694-701.
- Farmer EE, Ryan CA. 1990. Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc Natl Acad Sci USA. 87: 7713-7716.
- Gadaleta A, Giancaspro A, Blechl AE, Blanco A. 2008. A transgenic durum wheat line that is free of marker genes and expresses 1Dy10. J Cereal Sci. 48: 439-445.
- Gao Z, Xie X, Ling Y, Muthukrishnan S, Liang GH. 2005. Agrobacterium tumefaciens-mediated sorghum transformation using a mannose selection system. Plant Biotechnol J. 3: 591-599.
- Grootboom AW, Mkhonza NL, O’Kennedy MM, Chakauya E, Kunert K, Chikwamba RK. 2010. Biolistic mediated sorghum (Sorghum bicolor L. Moench) transformation via mannose and bialaphos based selection systems. Int J Bot. 6: 89-94.
- Gurel S, Gurel E, Kaur R, Wong J, Meng L, Tan HQ, Lemaux PG. 2009. Efficient, reproducible Agrobacterium-mediated transformation of sorghum using heat treatment of immature embryos. Plant Cell Rep. 28: 429-444.
- He Z, Duan Z, Liang W, Chen F, Yao W, Liang H, Yue C, Sun Z, Chen F, Dai J. 2006. Mannose selection system used for cucumber transformation. Plant Cell Rep. 25: 953-958.
- He Z, Fu Y, Si H, Hu G, Zhang S, Yu Y, Sun Z. 2004. Phosphomannose-isomerase (pmi) gene as a selectable marker for rice transformation via Agrobacterium. Plant Sci. 166: 17-22.
- Jin RG, Liu YB, Tabashnik BE, Borthakur D. 2000. Development of transgenic cabbage (Brassica oleracea var. capitata) for insect resistance by Aagrobacterium tumefaciens-mediated transformation. In Vitro Cell Dev Biol Plant. 36: 231-237.
- Joersbo M, Donaldson L, Kreiberg J, Petersen SG, Brunstedt J, Okkels FT. 1998. Analysis of mannose selection used for transformation of sugar beet. Mol Breed. 4: 111-117.
- Joersbo M, Petersen SG, Okkels FT. 1999. Parameters interacting with mannose selection employed for the production of transgenic sugar beet. Physiol Plant. 105: 109-115.
- Kirsh VA, Peters U, Mayne ST, Subar AF, Chatterjee N, Johnson CC, Hayes RB. 2007. Prospective study of fruit and vegetable intake and risk of prostate cancer. J Natl Cancer Inst. 99: 1200-1209.
- Ku JJ, Park YH, Park YD. 2006. A non-antibiotic selection system uses the phosphomannose-isomerase (PMI) gene for Agrobacterium-mediated transformation of Chinese cabbage. J Plant Biol. 49: 115-122.
- Liu CW, Lin CC, Yiu JC, Chen JJW, Tseng MJ. 2008. Expression of a Bacillus thuringiensis toxin (cry1Ab) gene in cabbage (Brassica oleracea L. var. capitata L.) chloroplasts confers high insecticidal efficacy against Plutella xylostella. Theor Appl Genet. 117: 75-88.
- Llorach R, Espín JC, Tomás-Barberán FA, Ferrere F. 2003. Valorization of cabbage (Brassica oleracea L. var. botrytis) by-products as a source of antioxidant phenolics. J Agric Food Chem. 51: 2181-2187.
- Logemann J, Schell J, Willmitzer L. 1987. Improved method for the isolation of RNA from plant tissue. Anal Biochem. 163: 16-20.
- Lucca P, Ye X, Potrykus I. 2001. Effective selection and regeneration of transgenic rice plants with mannose as selective agent. Mol Breed. 7: 43-49.
- McGarvey P, Kaper JM. 1991. A simple and rapid method for screening transgenic plants using the PCR. BioFeedback. 4: 428-432.
- Metz TD, Dixit R, Earle ED. 1995. Agrobacterium tumefaciens mediated transformation of broccoli (Brassica oleracea var. italica) and cabbage (B. oleracea var. capitata). Plant Cell Rep. 15: 287-292.
- Min BW, Cho YN, Song MJ, Noh TK, Kim BK, Chae WK, Park YS, Choi YD, Harn CH. 2007. Successful genetic transformation of Chinese cabbage using phosphomannose isomerase as a selection marker. Plant Cell Rep. 26: 337-344.
- Murashige T, Skoog F. 1962. A revisedr medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 15: 473-497.
- Negrotto D, Jolley M, Beer S, Wenck AR, Hansen G. 2000. The use of phosphomannose-isomerase as a selectable marker to recover transgenic maize plants (Zea mays L.) via Agrobacterium transformation. Plant Cell Rep. 19: 798-803.
- Pastori GM, Wilkinson MD, Steele SH, Sparks CA, Jones HD, Parry MAJ. 2001. Age-dependent transformation frequency in elite wheat varieties. J Exp Bot. 52: 857-863.
- Pius PK, Achar PN. 2000. Agrobacterium-mediated transformation and plant regeneration of Brassica oleracea var. capitata. Plant Cell Rep. 19: 888-892.
- Rafat A, Aziz MA, Rashid AA, Abdullah SNA, Kamaladini H, Sirchi MHT, Javadi MB. 2010. Optimization of Agrobacterium tumefaciens-mediated transformation and shoot regeneration after co-cultivation of cabbage (Brassica oleracea subsp. capitata) cv. KY Cross with AtHSP101 gene. Sci Hortic. 124: 1-8.
- Reed J, Privalle L, Powell ML, Meghji M, Dawson J, Dunder E, Suttie J, Wenck A, Launis K, Kramer C, Chang YF, Hansen G, Wright M. 2001. Phosphomannose isomerase: An efficient selectable marker for plant transformation. In Vitro Cell Dev Biol Plant. 37: 127-132.
- Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning A laboratory manual. Cold Spring Harbor Press. NY.
- Seo HS, Song JT, Cheong JJ, Lee YH, Lee YW, Hwang I, Lee JS, Choi YD. 2001. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses. Proc Nat Acad Sci USA. 98: 4788-4793.
- Sigareva M, Spivey R, Willits MG, Kramer CM, Chang YF. 2004. An efficient mannose selection protocol for tomato that has no adverse effect on the ploidy level of transgenic plants. Plant Cell Rep. 23: 236-245.
- Sohn HB, Lee HY, Seo JS, Jung C, Jeon JH, Kim JH, Lee YW, Lee JS, Cheong JJ, Choi YD. 2011. Overexpression of jasmonic acid carboxyl methyltransferase increases tuber yield and size in transgenic potato. Plant Biotechnol Rep. 5: 27-34.
- Sparrow PAC, Dale PJ, Irwin JA. 2004. The use of phenotypic markers to identify Brassica oleracea genotypes for routine high-throughput Agrobacterium-mediated transformation. Plant Cell Rep. 23: 64-70.
- Srivastava V, Reddy AS, Guha-Mukherjee S. 1988. Transformation of Brassica oleracea mediated by oncogenic Agrobacterium tumefaciens. Plant Cell Rep. 7: 504-507.
- Stitz M, Gase K, Baldwin IT, Gaquerel E. 2011. Ectopic expression of AtJMT in Nicotiana attenuata: creating a metabolic sink has tissue-specific consequences for the jasmonate metabolic network and silences downstream gene expression. Plant Physiol. 157: 341-354.
- Stoykova P, Stoeva-Popova P. 2011. PMI (manA) as a nonantibiotic selectable marker gene in plant biotechnology. Plant Cell Tiss Organ Cult. 105: 141-148.
- Thiruvengadam M, Hsu WH, Yang CH. 2011. Phosphomannose-isomerase as a selectable marker to recover transgenic orchid plants (Oncidium Gower Ramsey). Plant Cell Tiss Organ Cult. 104: 239-246.
- Todd R, Tague BW. 2001. Phosphomannose Isomerase: A Versatile Selectable Marker for Arabidopsis thaliana Germ-Line Transformation. Plant Mol Biol Rep. 19: 307-319.
- Tsukazaki H, Kuginuki Y, Aida R, Suzuki T. 2002. Agrobacterium-mediated transformation of a doubled haploid line of cabbage. Plant Cell Rep. 21: 257-262.
- Verkerk R, Schreiner M, Krumbein A, Ciska E, Holst B, Rowland I, de Schrijver R, Hansen M, Gerhäuser C, Mithen R, Dekker M. 2009. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol Nutr Food Res. 53(SUPPL 2): 219-265.
- Virgili F, Marino M. 2008. Regulation of cellular signals from nutritional molecules: a specific role for phytochemicals, beyond antioxidant activity. Free Radic Biol Med. 45: 1205-1216.
- Wallbraun M, Sonntag K, Eisenhauer C, Krzcal G, Wang YP. 2009. Phosphomannose-isomerase (pmi) gene as a selectable marker for Agrobacterium-mediated transformation of rapeseed. Plant Cell Tiss Organ Cult. 99: 345-351.
- Wang AS, Evans RA, Altendorf PR, Hanten JA, Doyle MC, Rosichan JL. 2000. A mannose selection system for production of fertile transgenic maize plants from protoplasts. Plant Cell Rep. 19: 654-660.
- Wright M, Dawson J, Dunder E, Suttie J, Reed J, Kramer C, Chang Y, Novitzky R, Wang H, Artim-Moore L. 2001. Efficient biolistic transformation of maize (Zea mays L.) and wheat (Triticum aestivum L.) using the phosphomannose isomerase gene, pmi, as the selectable marker. Plant Cell Rep. 20: 429-436.