Abstract
An Agrobacterium-mediated transformation system was developed for the ornamental cherry species Prunus incisa. This system uses both an antibiotic resistance gene (NPTII) and a visible selectable marker, the green fluorescent protein (GFP), to select plants. Cells from leaf and root explants were transformed with a NPTII/GFP fusion gene, and selected visually using fluorescence microscopy. Transformed cells were then induced to undergo embryogenesis and reselected by growing on media containing kanamycin. The presence of the GFP/NPTII fusion gene in all parts of transgenic plants grown in the greenhouse for one year was confirmed by PCR and Southern blot analysis. This transformation and selection system will be useful in future work to introduce genes for pathogen resistance and ornamental traits into flowering cherry germplasm.
-
Key words: Flowering cherry, Selectable marker, Green fluorescent protein, Transformation
INTRODUCTION
The flowering cherry (Prunus species) is a popular ornamental plant in many temperate parts of the world. The breeding program at the U.S. National Arboretum focuses on broadening the genetic base of ornamental Prunus and developing new cultivars with superior disease and pest resistance and tolerance to environmental stresses. Due to quarantine restrictions, it has become increasingly difficult to bring new Prunus germplasm into the U.S. for breeding; hence, genetic engineering may be one of the few options to introduce genes for resistance to microbial and insect pests into Prunus.
MATERIALS AND METHODS
Plant material
Actively growing
in vitro shoots of
Prunus incisa ‘February Pink’ (
Cheong and Pooler 2004) were transferred to MS medium containing 3% sucrose, 0.2% Gelrite, and 1uM IBA, to induce roots. Leaves and elongated roots were taken from the shoots after 3–4 weeks. Leaves were cut through the mid-vein, and the leaf margins were cut off to create a wound. The terminal 1 cm of roots was harvested and root tips were removed. Roots were rinsed with sterile liquid MS medium to remove substrate medium, and roots were wounded by gentle squeezing with forceps.
The Agrobacterium binary vector pLC101 containing the EGFP/NPTII fusion gene driven by the double CaMV35S promoter (provided by Dr. Dennis Gray, University of Florida) was used in binary transformation. Agrobacterium tumefaciens strain EHA105 was used for transformation.
Cocultivation, Embryogenesis and Selection
Explants were incubated in bacterial resuspension solution (MS medium with 0.1M acetosyringone) with gentle shaking for 30 min to induce infection. Tissues were co-cultivated on solidified (0.2% Gelrite) induction medium (MS medium with 10 uM 2,4-D plus 3% sucrose) without antibiotics for three days. After co-cultivation, explants were washed with liquid MS medium containing 300 mg/L carbenicillin and 100 mg/L cefotaxime, then cultivated on induction medium with antibiotics as above. Cultures were kept in the dark at 22 ± 2°C. After 7 days of culture, calli emitting green light were selected under a fluorescence microscope using 475 nm excitation and 510 nm emission filters.
Cell clusters emitting fluorescent green light (expressing GFP) were isolated and cultivated in liquid induction medium for a week. Suspended cells were then plated onto solid media and placed in the dark until cells formed callus at least 1 mm in diameter. GFP-expressing callus was then reselected and transferred to fresh medium every two weeks. Somatic embryos were induced from this callus on embryo induction medium consisting of MS medium with 10 uM 2,4-D (
Cheong and Pooler 2004) plus antibiotics as above. Induced somatic embryos were transferred to hormone-free MS medium with 50 mg/L kanamycin for selection.
Leaves, stems, and somatic embryos were used for PCR analysis of the GFP gene in transgenic plants. DNA from transgenic plants and non-transgenic controls was extracted using the DNeasy Plant Maxi Kit (Qiagen, Valencia, CA). Two primer sets were used to amplify the intact whole section (1.54 kb, 5′-GTGAGCAAGGGCGAGGAGCT-3′ and 5′-CGGCCGCTTATCAGAAGAAC-3′.) or internal section (0.6 kb, 5′-ATCAAGGTGAACTTCAAGAT-3′ and 5′-GATGGATACTTTCTCGGCAG-3′) of the EGFP/NPTII fusion gene (
Fig. 1). PCR products were separated on 1.5% agarose gel and visualized by staining with ethidium bromide.
For Southern blot analysis, 5 μg of the DNA from transformed and untransformed plants was digested with
BamHI at 37°C overnight, separated on a 0.8% agarose gel, and blotted onto a membrane (Amersham-Pharmacia Biotech, Sunnyvale, CA). To prepare the probe, the 542 bp NPTII fragment was amplified from the original DNA plasmid using specific NPTII primers (5′ GTTGTCACTG-AAGCGGGAAG 3′ and 5′ ACTCGTCAAGAAGGCGATAG 3′) and labeled with
32P-dATP. PCR amplification conditions were as follows: 94°C for 5 min, followed by 30 cycles of denaturation for 30 sec at 94°C, annealing for 30 s at 55°C, extension for 1 min at 72°C, followed by an additional 10 min extension at 72°C. Probe hybridization and washing of blots were performed under medium stringency conditions according to
Keates et al. (2003). Blots were imaged at 200 μm resolution on a Typhoon 8600 Variable Mode Imager (Molecular Dynamics/Amersham-Pharmacia Biotech, Sunnyvale, CA).
RESULTS
Transformation
Within two weeks of culture in the dark, 1–2 mm cell clusters had formed on the cut edges of leaves. Cell clusters on roots were difficult to distinguish from normal epidermal cells on the surfaces. However, transformed cells expressing the green fluorescent protein gene were visible under an inverted microscope in both explants, and were found in several locations on an explant. Transformation efficiency was 7.5% for leaves and 2.6% for roots (
Table 1). Despite the wounds on the root surfaces to aid bacterial infection, there were fewer transformed cells in roots than in leaf explants, although shoots development from somatic embryogenesis from root-derived cells was slightly higher than that for cells from leaves (
Table 1). Cells expressing the GFP gene were selected visually using a fluorescent microscope (
Fig. 2). A total of 42 cell lines were selected from leaf explants and were cultivated in liquid medium and then plated onto solidified somatic embryo induction medium. Cells grown in liquid medium for one week showed vigorous growth as opposed to cells in agar-solidified medium (data not shown). Plated cells formed callus and then roots. Eight of these callusing cell lines formed embryos and roots. From these embryos, 12 plantlets were developed and were transferred to the greenhouse for acclimatization and subsequently used for transgene detection.
One-year-old plants in the greenhouse did not express visually detectable levels of GFP from any plant part. However, transgenic plants were verified by both PCR and Southern blot analysis of the whole plant. Successful PCR amplification of transgenic DNA was obtained using PCR primers for the entire transformation vector and the internal EGFP/NPTII gene (
Fig. 3A). The presence of the transgene was confirmed by Southern Blot analysis using the NPTII gene as a probe (
Fig. 3B).
DISCUSSION
Improvements to many agricultural and horticultural crops have been made using
Agrobacterium-mediated transformation (
Gelvin 2003). Several factors affect the efficiency of transformation, including
Agrobacterium strain, explant type, and organogenesis system. For example,
Agrobacterium strain EHA105 has been shown to be an effective agent for genetic transformation of recalcitrant fruit tree species (
De Bondt et al. 1994;
Mourgues et al. 1996;
Pena et al. 1995;
Ramesh et al. 2006;
Scorza et al. 1994), and proved to be effective in our study with ornamental
Prunus as well. Explant type and organogenesis system are also important factors in transformation efficiency. Direct or indirect shoot regeneration from leaf discs was generally used in previous work with
Prunus species (
Miguel and Oliveira 1999;
Ramesh et al. 2006) and other crops (
De Bondt et al. 1994;
Mourgues et al. 1996;
Pena et al. 1995), while root pieces had rarely been used for transformation. The high rate of embryogenesis from roots in
Prunus incisa (
Cheong and Pooler 2004) facilitated transformation in our study. The relatively high transformation rates (2.6–7.5%) may be due to the high embryogenic efficiency from root tissue, which seemed to be unique to this species.
Using GFP as a reporter gene and a selectable marker was also advantageous in our study. Unlike other reporter genes such as GUS, cells containing GFP could be visualized non-destructively at an early stage of culture, which also allowed for early selection of transgenic cells. By optimizing bacterial strain, regeneration system, and selectable marker, we were able to obtain confirmed transgenic plants of Prunus incisa.
Fig. 1Schematic diagram of the pLC101 vector and probe used for Southern blot analysis. LB = left border; RB, = right border; Ter = transcriptional terminator sequence; 35S P = cauliflower mosaic virus (CaMV) 35S promoter; NPTII = neomycin phosphotransferase, EGFP = enhanced green fluorescent protein.

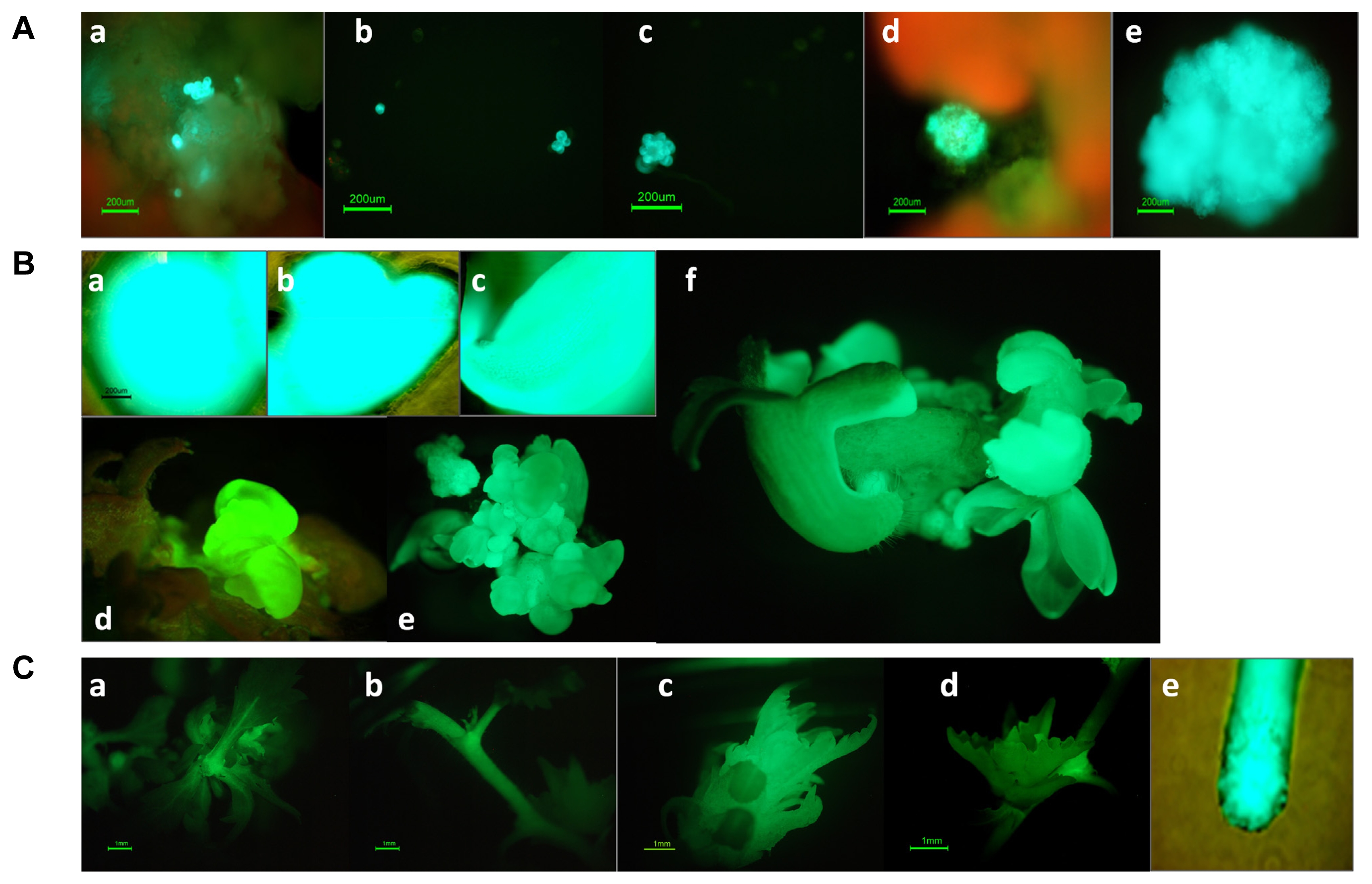
Fig. 2
Callus induction and somatic embryo development of transformed Prunus incisa cells and GFP expression in transgenic plants.
(A) GFP expression in (a) single cells and cell clusters; (b and c) suspension culture and plated cultures; and (d and e) propagated callus. (B) Development of somatic embryos of transformed cells at the (a) globular stage; (b) heart-shaped stage; (c) cotyledon stage; and (d, e, and f) germination stage. (C) GFP expression in various organs of transgenic plants (a) somatic embryo; (b) plant stem; (c) apical bud; (d) axillary bud; and (e) root tip.
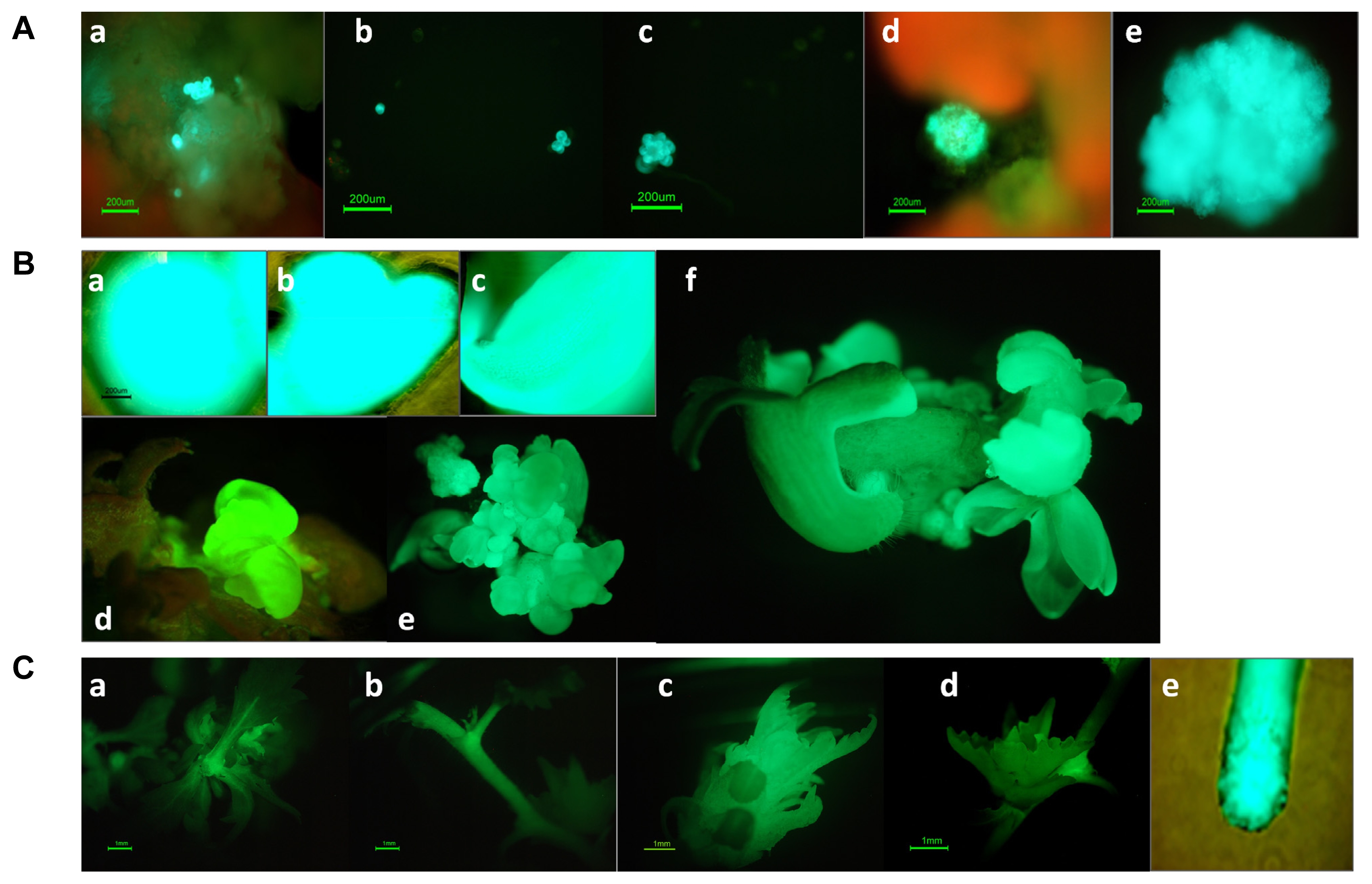
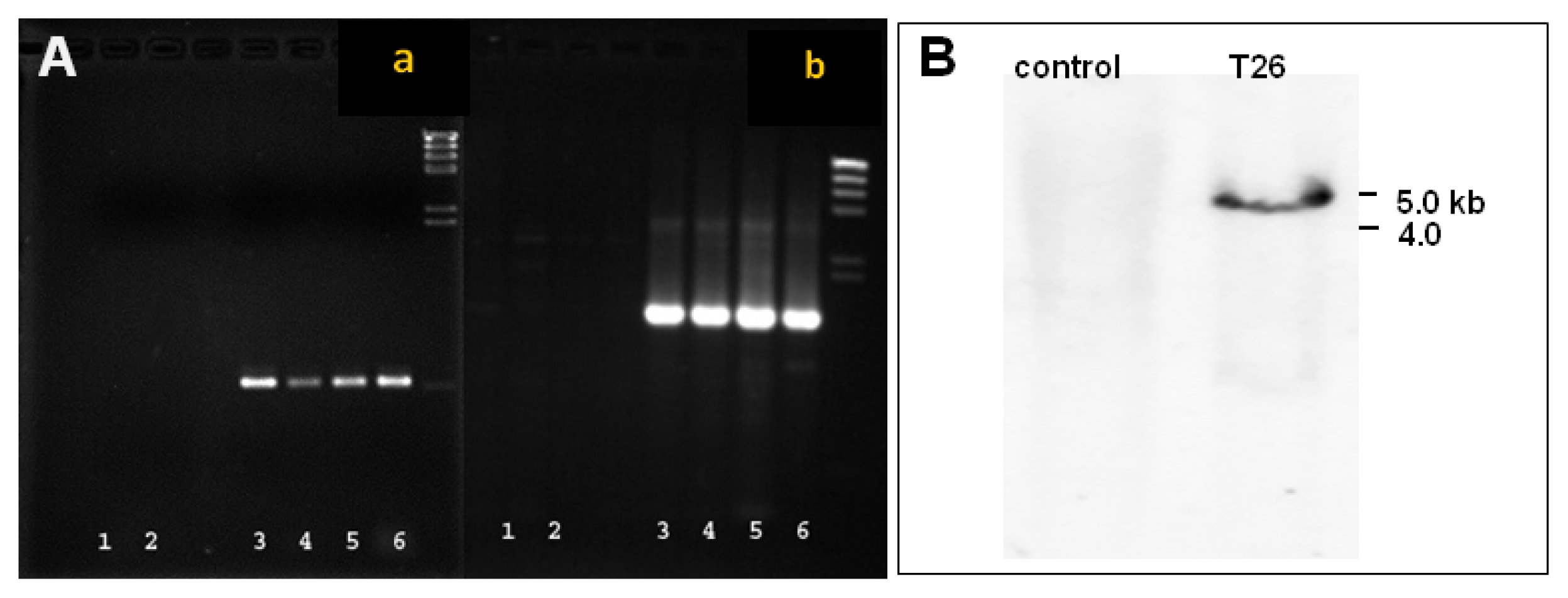
Fig. 3Detection of the GFP gene in transgenic plants. (A) PCR analysis where a = amplification of the whole EGFT/NPTII fusion gene (1.54 kb) and b = amplification of the internal EGFP/NPTII fusion gen (0.6 kb). Lanes 1 and 2 represent non-transformed stems and leaves, respectively; Lanes 3–6 represent transformed stems, leaves, somatic embryos, and positive control plasmid DNA, respectively. (B) Southern blot analysis of non-transformed control and transformed line T26 probed with NPTII.
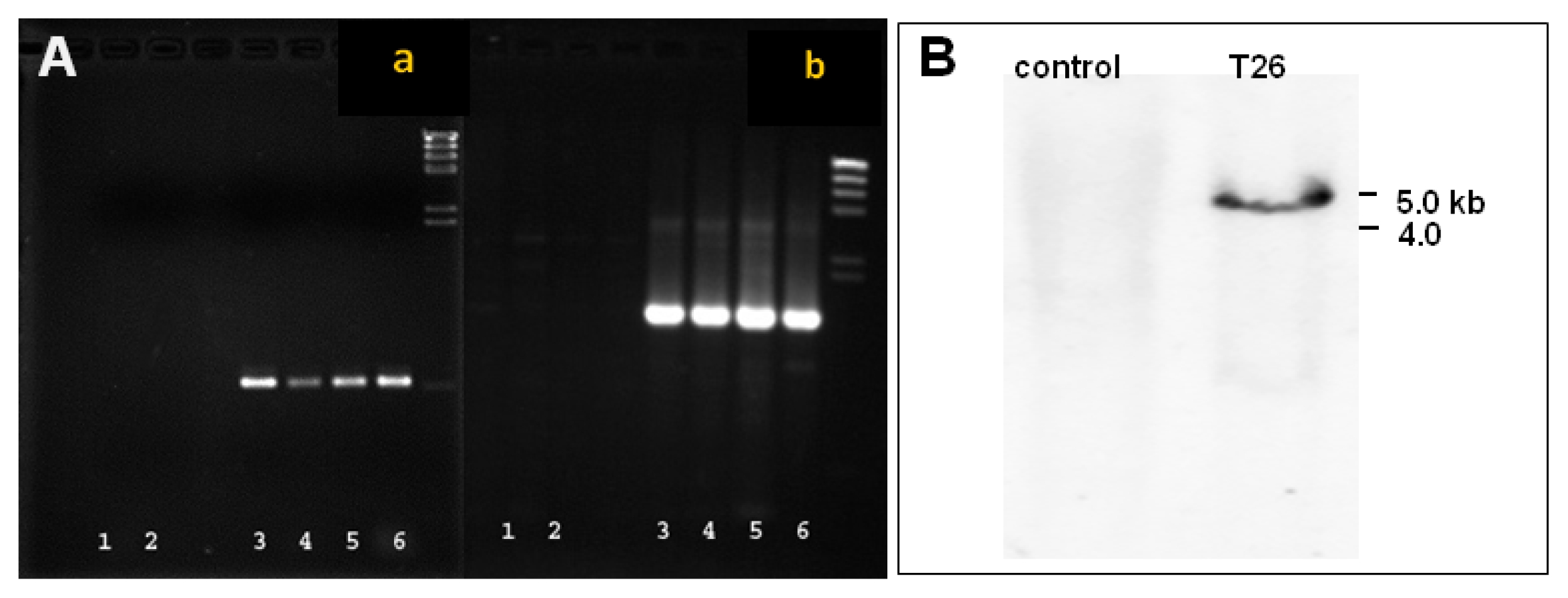
Table 1Efficiency of callus and root induction and GFP gene transformation for leaf and root explants of Prunus incisa.
Table 1
|
Explant Type |
Total No. of treated explants |
No. of explants forming callus (%) |
No. of explants expressing GFP (%) |
No. of transformed plantlets |
|
Leaf |
173 |
83 (48%) |
13 (7.5%) |
6 |
|
Root |
153 |
52 (34%) |
4 (2.6%) |
6 |
References
- Camara Machado A, Puschmann M, Puhringer H, Kremen R, Katinger H, Camara Machado ML. 1995. Somatic embryogenesis of Prunus subhirtella autumno rosa and regeneration of transgenic plants after Agrobacterium-mediated transformation. Plant Cell Rep. 14: 335-340.
- Cheong EJ, Pooler MR. 2004. Factors affecting embryogenesis in Prunus incisa cv. February Pink. Plant Cell Rep. 22: 810-815.
- Dai W, Magnusson V, Johnson C. 2007. Agrobacterium-mediated transformation of chokecherry (Prunus virginiana L). HortScience. 42: 140-142.
- De Bondt A, Eggermont K, Durat P, De Vil M, Vandeleyden J, Broekaert WF. 1994. Agrobacterium-mediated transformation of apple (Malus x domestica): an assessment of factors affecting gene transfer efficiency during early transformation steps. Plant Cell Rep. 13: 587-593.
- Druart P, Delporte F, Brazda M, Ugarte-Ballon C, Camara Machado A, Camara Machado ML, Jacquemin J, Watillon B. 1998. Genetic transformation of cherry trees. Acta Horticulturae. 468: 71-76.
- Escobar MA, Park JI, Polito VS, Leslie CA, Uratsu SL, McGranahan GH, Dankerar AM. 2000. Using GFP as a scorable marker in walnut somatic embryo transformation. Annals of Botany. 85: 831-835.
- Gelvin SB. 2003. Agrobacterium-mediated plant transformation: the Biology behind the “Gene-Jockeying” tool. Microbiology Molecular Biology Reviews. 67: 16-37.
- Ghorbel R, Juarez J, Navarro L, Pena L. 1999. Green fluorescent protein as a screenable marker to increase the efficiency of generating transgenic woody fruit plants. Theor Appl Genet. 99: 350-358.
- Jordan MC. 2000. Green fluorescent protein as a visual marker for wheat transformation. Plant Cell Rep. 19: 1069-1075.
- Kaeppler HF, Menon GK, Skadsen RW, Nuutila AM, Carlson AR. 2000. Transgenic oat plants via visual selection of cell expressing green fluorescent protein. Plant Cell Rep. 19661-666.
- Keates SE, Kostman TA, Anderson JD, Bailey BA. 2003. Altered gene expression in three plant species in response to treatment with Nep1, a fungal protein that causes necrosis. Plant Physiol. 132: 1610-1622.
- Mante S, Morgens PH, Scorza R, Cordts JM, Callahan AM. 1991. Agrobacterium-mediated transformation of plum (Prunus domestica L.) hypocotyls slices and regeneration of transgenic plants. BioTechnology. 9: 853-857.
- Maximova SN, Dandekar AM, Guiltinan MJ. 1998. Investigation of Agrobacterium-mediated transformation of apple using green fluorescent protein: high transient expression and low stable transformation suggest that factors other than T-DNA transfer are rate-limiting. Plant Molecular Biology. 37: 549-559.
- Miguel CM, Oliveira MM. 1999. Transgenic almond (Prunus dulcis Mill.) plants obtained by Agrobacterium-mediated transformation of leaf explants. Plant Cell Rep. 18: 387-393.
- Molinier JC, Hahne G. 2000. Use of green fluorescent protein for detection of transformed shoots and homozygous offspring. Plant Cell Rep. 19: 219-223.
- Mourgues F, Chevreau E, Lambert C, De Bondt A. 1996. Efficient Agrobacterium-mediated transformation and recovery of transgenic plants from pear (Pyrus communis L). Plant Cell Rep. 16: 245-249.
- Pena L, Cerevera M, Juarez J, Ortega C, Pina JA, Duran-Vila N, Navarro L. 1995. High efficiency Agrobacterium-mediated transformation and regeneration of citrus. Plant Sci. 104: 183-191.
- Perez-Clemente RM, Perez-Sanjuan A, Garcia-Ferriz L, Beltran J, Canas LA. 2004. Transgenic peach plants (Prunus persica L.) produced by genetic transformation of embryo sections using the green fluorescent protein (GFP) as an in vivo marker. Molecular Breeding. 14: 419-427.
- Phillips JA, Collins GG, Sedgley M. 2001. Factors affecting Agrobacterium-mediated gene transfer and the selection of transgenic calli in paper shell almond (Prunus dulcis Mill). The Journal of Horticultural Science & Biotechnology. 76: 522-528.
- Ramesh SA, Kaiser BN, Franks T, Collins G, Sedgley M. 2006. Improved methods in Agrobacterium-mediated transformation of almond using positive (mannose/pmi) or negative (kanamycin resistance) selection-based protocols. Plant Cell Rep. 25: 821-828.
- Scorza R, Morgens PH, Cordts JM, Mante S, Callahan AM. 1990. Agrobacterium-mediated transformation of peach (Prunus persica L. Batsch) leaf segments, immature embryos, and long-term embryogenic callus. In Vitro Cell Dev Biol. 26: 829-834.
- Scorza R, Ravelonandro M, Callahan AM, Cordts JM, Fuchs M, Dunez J, Gonzalves D. 1994. Transgenic plums (Prunus domestica L.) express the plum pox virus coat protein gene. Plant Cell Rep. 14: 18-22.
- Sidorov VA, Kasten D, Pang SZ, Hajdukiewicz PTJ, Staub JM, Nehra NS. 1999. Stable chloroplast transformation in potato: use of green fluorescent protein as a plastid marker. The Plant Journal. 19: 209-216.
- Tian L, Ali Canli F, Wang X, Sibbald S. 2009. Genetic transformation of Prunus domestica L. using the hpt gene coding for hygromycin resistance as the selectable marker. Scientia Horticulturae. 119: 339-343.
- Urtubia C, Devia J, Castro A, Zamora P, Aguirre C, Tapia E, Barba P, Dell Orto P, Moynihan MR, Petri C, Scorza R, Prieto H. 2008. Agrobacterium-mediated genetic transformation of Prunus salicina. Plant Cell Rep. 27: 1333-1340.
Citations
Citations to this article as recorded by