Abstract
To utilize the favorable gene(s) from winter wheat for genetic improvement of spring wheat, this study was carried out to identify the quantitative trait loci (QTL) associated with grain protein content (GPC) and grain hardness (GH) by analysis of recombinant inbred lines (RILS) derived from a cross between spring wheat and spring version of winter wheat. A genetic map of 334 loci was constructed which covered 1575.30cM on all 21 chromosomes. Two QTLs on 3B and 5B chromosome were detected for GPC. A QTL identified barc77 on chromosome 3B had additive effect of 0.17 and the other QTL identified by gwm499 on chromosome 5B had additive effect of 0.19. There were two major QTLs for GH identified on Chromosome 1B and chromosome 5A. The QTL on 1B was localized within a 18.7cM region flanked by wmc719 and wmc367-1 with 1.75 additive effect. The QTL on chromosome 5A flanked by SNP markers, IWA6573 and IWA2363, had additive effect of 1.44.
-
Key words: Quantitative trait loci (QTL), Recombinant inbred line (RIL), Grain protein content, Grain hardness, Winter wheat, Spring wheat
INTRODUCTION
The grain storage proteins in bread wheat (
Triticum aestivum L.) are very important for the visco-elastic properties of bread dough (
Turner et al. 2004). The improvement in grain protein content (GPC) and its composition has been a major concern of wheat breeders. The composition and amount of protein in bread wheat will determine the end-use quality after harvest. There are many genetic studies about wheat grain protein which make up the gluten, and their functionality has been investigated (
Shewry et al. 1997;
Rogers et al. 2001). However, the genetics of protein quantity is not well understood. Grain protein content is controlled by a complex genetic system and is strongly influenced by several environmental factors, making genetic analysis much more difficult.
Grain texture has been extensively studied because of its influence on bread-making quality (
Groos et al. 2004). The texture of the endosperm results mostly from the expression of a major gene designed Hardness (Ha) located on the short arm of chromosome 5D (
Law et al. 1978).
Sourdille et al. (1996) mapped the Ha gene at chromosome 5DS, close to loci of puroindoline proteins.
Symes (1965) reported that grain hardness is mainly due to a single gene. However, the same study also indicated the existence of minor genes that modify the action of the main hardness gene. Although Ha/ha has been reported to be important for kernel hardness, its gene product is not yet known with certainty. Genetic analyses have shown the main effect of chromosomes 2A, 2D, 3A, 4A, 4B, 5A, 5B, 6D and 7A on hardness but there also were minor effects between hard and soft wheat genotypes (
Campbell et al. 1999;
Morrison et al. 1989;
Sourdille et al. 1996;
Symes 1965).
Many of the economically important traits in crop plants are quantitative in nature and are controlled by many genes or gene complexes, those are described as quantitative trait loci (QTL). Mapping of the QTL has been greatly facilitated due to the availability of various molecular markers and the development of improved and powerful statistical methods.
Spring and winter wheat have been selected for many of the same traits, and the germplasm pools have been largely separate, it is likely that favorable genes would exist in one germplasm base that may be useful in the other group. Crosses between spring wheat and winter segregate for growth habit, and thus progeny from such a cross cannot be evaluated in a common environment. Thus, it has been impossible to conduct genetic analysis to identify favorable genes for quantitative traits that may be transferred between winter and spring wheat. The recent development of molecular markers for the Vrn genes provides an opportunity to develop populations that have the same growth habit, yet are segregating for genes from winter and spring wheat.
In this study, the population derived from cross between ‘Choteau’ and spring version of ‘Yellowstone’ was subjected to QTL analysis of grain protein content and grain hardness to use the favorable gene(s) from the different germplasm pool.
MATERIALS AND METHODS
Plant materials and data collection
Recombinant inbred lines (RILs) were developed by single-seed descent starting with F2 generation from the cross between spring wheat ‘Choteau’ and spring version of winter wheat ‘Yellowstone’. The parents, ‘Choteau’ is widely grown hard red spring wheat cultivar which has high protein content in Montana, while ‘Yellowstone’ is a high yielding winter cultivar which has high grain hardness. A total of 97 RILs were derived from individual F6 plants. To make the spring version of ‘Yellowstone’, we backcrossed the spring allele at Vrn-A1 into the winter wheat variety ‘Yellowstone’ using molecular markers.
The RIL population was grown in replicated field trials under four environments, grown under rainfed and irrigated condition, from 2011 to 2012 in Bozeman, Montana (
Table 1). A subsample of seed was taken from each plot and was analyzed using the Single Kernel Characterization System 4100 (Perten, Huddings, Sweden) to determine grain hardness. Grain protein content was obtained on whole grain samples using a Foss Infratec 1241 Grain Analyzer (Tecator, Höganäs, Sweden).
The PCR reactions with simple sequence repeats (SSRs) which were selected all over the 21 wheat chromosomes were performed in a total volume of 25 μL containing 50–100 ng genomic DNA, 1X PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTPs, 1 unit Taq DNA polymerase, and 0.2 ug/ul each primer. After an initial denaturation at 94°C for 5 min., 10 cycles (decrease 1°C per cycle) were performed for 1 min at 94°C, 1 min at 63°C, 1 min at 72°C, followed by 35 cycles of 1 min at 94°C, 1 min at 55°C, 1 min at 72°C and final extension at 72°C for 10 min. The PCR products were loaded on a 12% (w/v) acrylamide gels and stained with Ethidium Bromide. The single nucleotide polymorphism (SNP) marker analysis was carried out at Washington State University.
QTL mapping
MapDisto (
http://mapdisto.free.fr) was used to develop a frame map for initial QTL detection. The map covered 1575.30cM and included 334 loci on all 21 chromosomes, comprising 257 SSRs and 77 SNP markers. QTL analysis was conducted for grain protein content and grain hardness using the entry means for average of total environments by QTL Cartographer (
http://statgen.ncsu.edu/qtlcart/WQTLCart.htm). QTLs were investigated with composite interval mapping (CIM) (
Zeng 1993,
1994) using standard CIM model, ‘Forward and Backward Regression method’. The walking speed for all QTL was 2cM. Centimorgan units were calculated using Kosambi mapping function. Logarithm of the odds (LOD) value was set by 1,000 permutations at an experiment-wise P < 0.01. A QTL was claimed to be significant at a LOD value of 2.5.
RESULTS
Ninety-seven RILs were developed from the cross between ‘Choteau’ and spring version of ‘Yellowstone’ following the SSD methods.
The mean performance of grain protein content (GPC) and grain hardness (GH) in four environments are given in
Table 2. Significant differences in GPC and GH were observed between two parents, ‘Choteau’ and spring version of ‘Yellowstone’. Across the four environments, ‘Choteau’ had a higher GPC (15.2%) than spring version of ‘Yellowstone’ (13.4%) while spring version of ‘Yellowstone’ had higher GH score than that of ‘Choteau’ by 9.2%. The GPC and GH in RILs ranged from 13.0% to 15.9% and from 50.03% to 83.29%, respectively, which showed transgressive segregants in both traits (
Table 2).
QTL analysis was conducted for the ‘Choteau’/spring version of ‘Yellowstone’ RIL population using the genetic map generated by MapDisto (
http://mapdisto.free.fr).
All putative QTLs are presented in
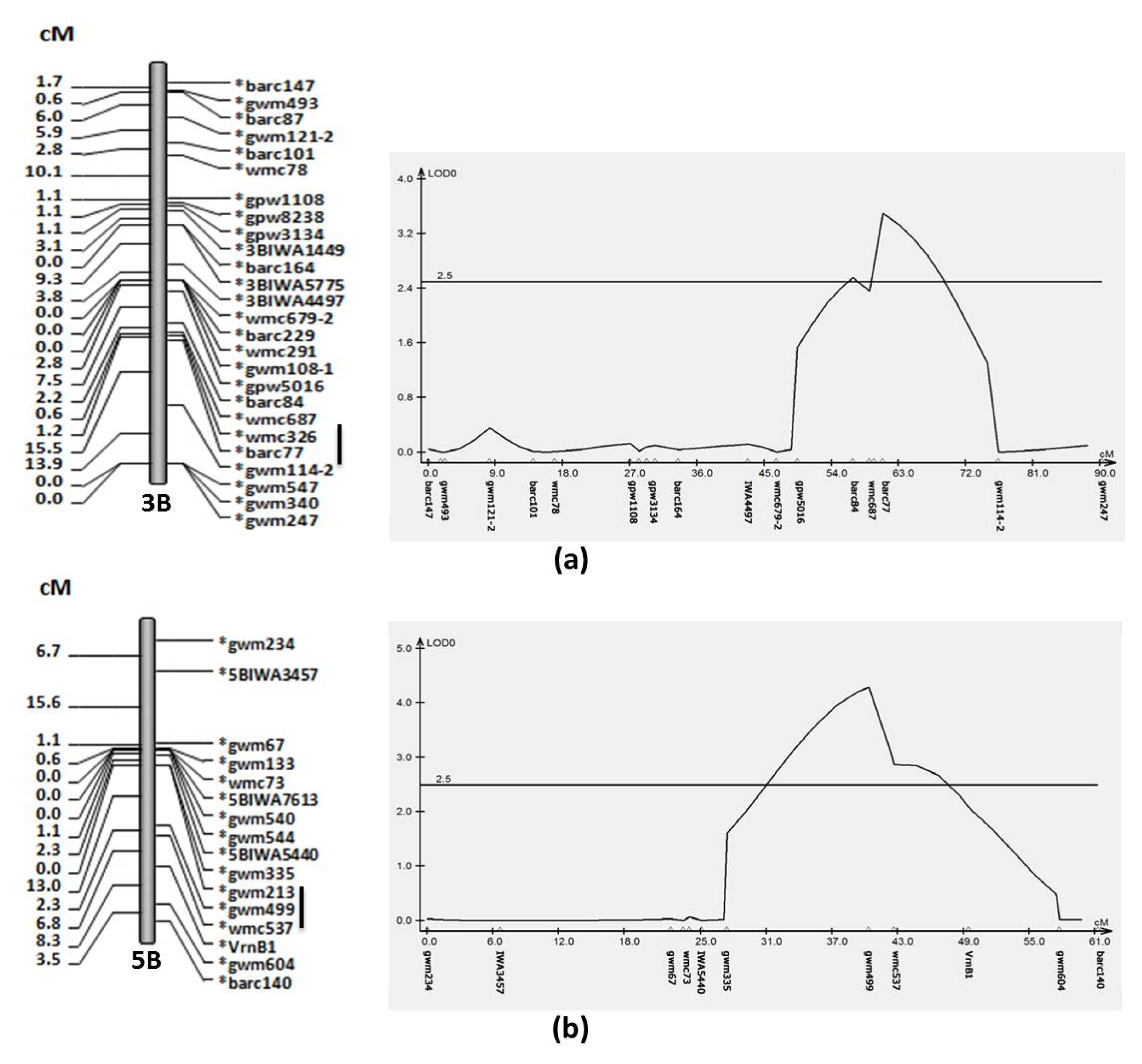
Table 3. There were two QTLs for GPC on chromosome 3B and 5B in the average of four circumstances. The ‘Choteau’ allele for the 3B QTL flanked by barc77 had additive effect of 0.17 and the other QTL identified by gwm499 on chromosome 5B had additive effect of 0.19 (
Fig. 1). Both QTLs with the high protein content allele came from ‘Choteau’.
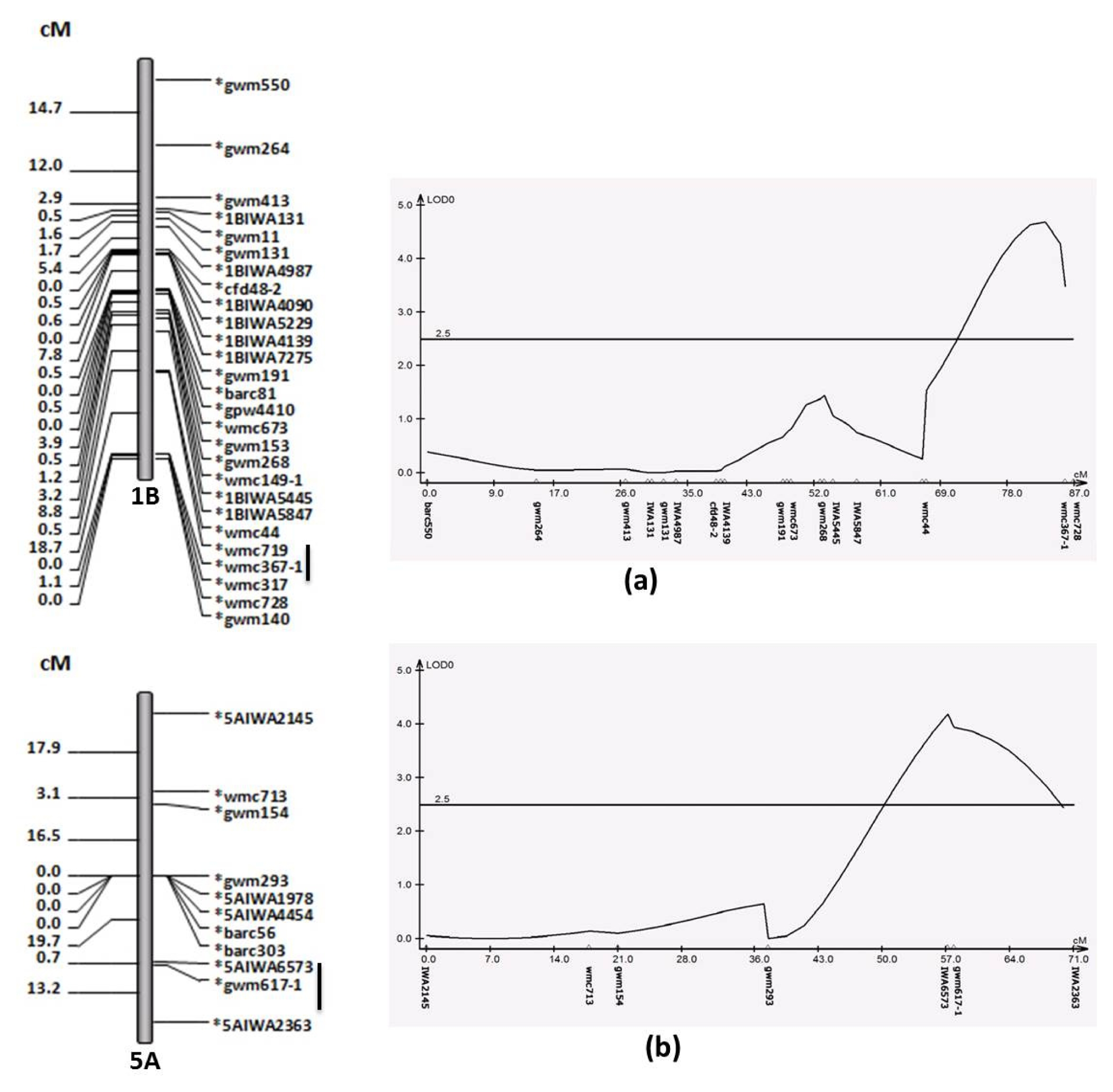
There were two major QTLs for GH identified on Chromosome 1B and chromosome 5A (
Table 3). The QTL on 1B was localized within 18.7cM region flanked by wmc719 and wmc367-1 with 1.75 of additive effect (
Fig. 2). The QTL on chromosome 5A flanked by SNP markers, IWA6573 and IWA2363, had additive effect of 1.44 (
Fig. 2). The alleles of spring version of ‘Yellowstone’ caused higher GH.
We estimated the effects of individual alleles by dividing the RILs into two groups. The closest marker linked to each QTL was used in the analysis. The contribution of each allele to the GPC and GH and standard errors are presented in
Table 4. The two alleles on chromosome 3B and 5B had same effect on GPC with increasing 0.3% of GPC and two alleles on 1B and 5A also similar effect on GH with increasing about 2.5% of GH.
DISCUSSION
Across the four environments, ‘Choteau’ had a significantly higher GPC (15.2%) than spring version of ‘Yellowstone’ (13.4%) while spring version of ‘Yellowstone’ had higher GH score than that of ‘Choteau’ by 9.2%. For the RIL population, some lines showed phenotypic values greater than the higher parent and smaller than the lower parent (i.e. transgressive segregants) in both traits (
Table 2) and this suggests that favorable alleles are present in both parents (
Table 2).
Groos et al. (2004) reported that the population showed both positive and negative transgression, which implicates that positive alleles may be found in both parental lines.
For GPC, the ‘Choteau’ allele for the 3B QTL flanked by barc77 had additive effect of 0.17 and the other QTL identified by gwm499 on chromosome 5B had additive effect of 0.19 (
Fig. 1). These results suggested that the marker gwm499 may either be tightly linked to QTL with a small effect or loosely linked to a QTL with a large effect. Both QTLs with high protein content allele came from ‘Choteau’. In bread wheat, genes for protein content have been located on chromosome 5A (
Morris et al. 1973), 5D (
Law et al. 1978;
Morris et al. 1973), and on chromosomes 1A, 1B and 7A (
Stein et al. 1992). The QTLs detected for GPC appeared differently from those detected in previous studies on wheat (
Turner et al. 2004;
Prasad et al. 1999;
Groos et al. 2003). These GPC related QTLs might be new QTLs from winter wheat or minor QTLs. To confirm the effect of QTLs associated with GPC, verification of the detected QTLs will be carried out with different populations.
For the GH, there were two major QTLs located on Chromosome 1B and chromosome 5A (
Table 3) and the alleles of spring version of ‘Yellowstone’ caused higher GH. These QTL regions have been previously reported to influence hardness (
Turner et al. 2004).
The QTL analysis conducted in this study would provide preliminary information of genetic regions that may be important for GPC and GH. However, further mapping and validation of the QTLs should be carried out before applying in marker assisted breeding.
Fig. 1A linkage map (left) and QTL analysis (right) for grain protein content on chromosome 3B (a) and 5B (b), flanking markers are underlined.
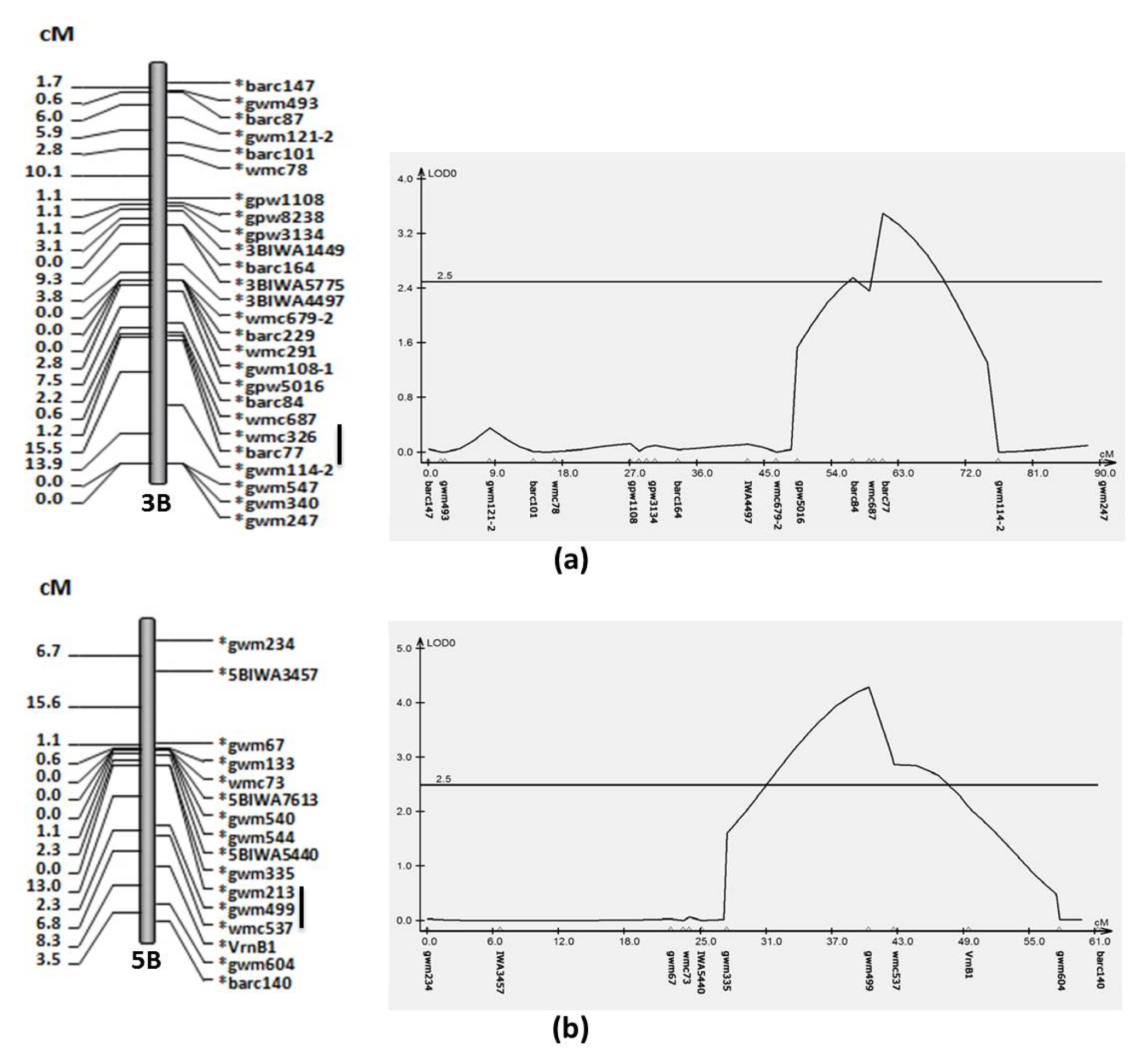
Fig. 2A linkage map (left) and QTL analysis (right) for grain hardness on chromosome 1B (a) and 5A (b), flanking markers are underlined.
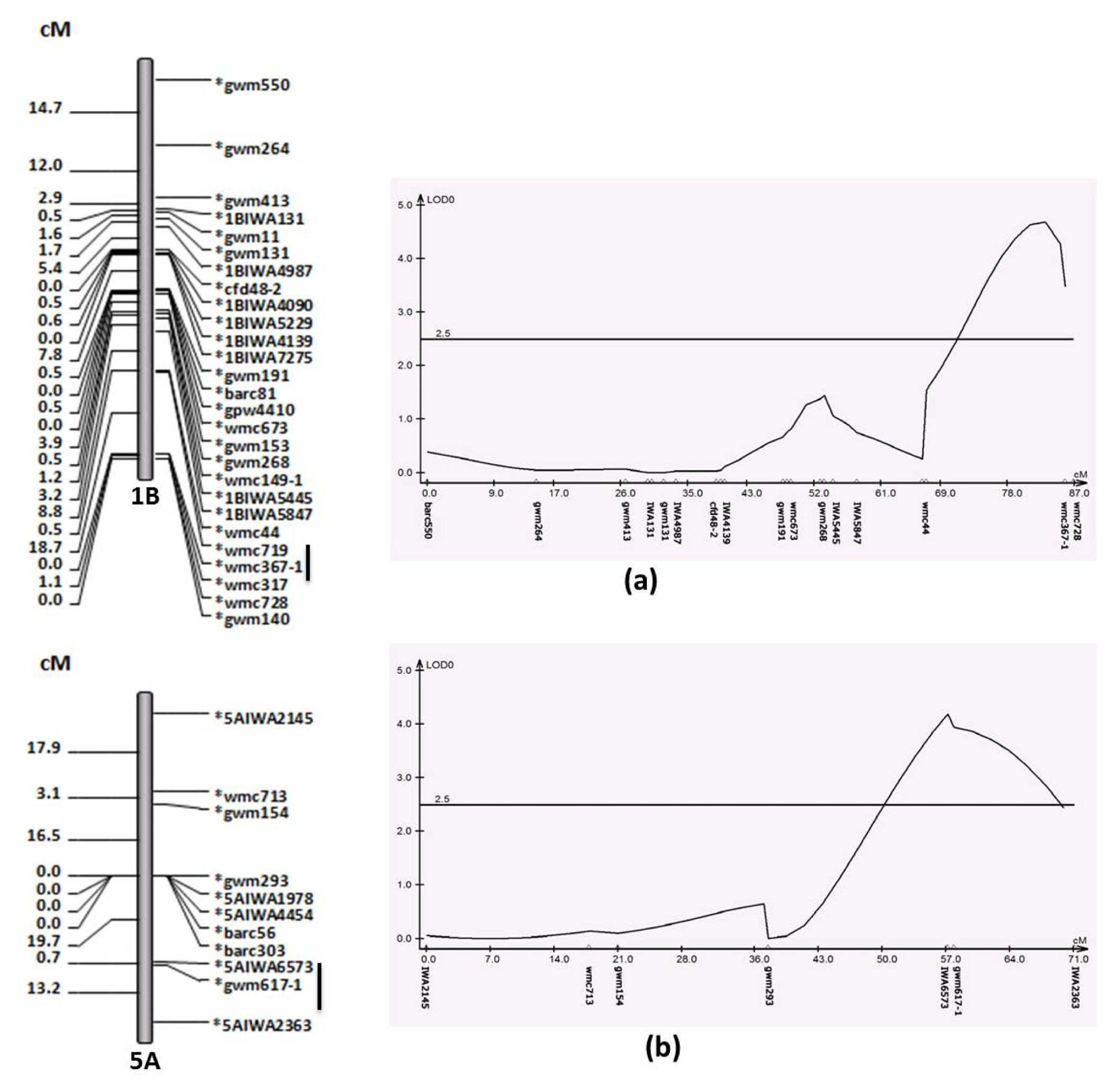
Table 1Population and environments used for genetic analysis of grain protein and hardness.
Table 1
|
Parents |
Number of RILsa
|
Year of experiment |
Irrigation regime |
|
Choteau/Spring version of Yellowstone |
97 |
2011 |
Rainfed, Irrigated |
|
2012 |
Rainfed, Irrigated |
Table 2Mean performance and range of grain protein content and grain hardness of the parental lines and mapping population in four environments.
Table 2
|
Line |
Grain protein content (%) |
Grain hardness (%) |
|
Choteau |
15.2*
|
68.14**
|
|
S-Yellowstonea
|
13.4 |
74.39 |
|
Mean of population |
14.3 |
70.39 |
|
Range of population |
13.0 – 15.9 |
50.03 – 83.29 |
Table 3Significant QTLs for grain protein content (GPC) and grain hardness (GH) detected through composite interval mapping of the Choteau/S-Yellowstone RILs in four environments.
Table 3
|
Trait |
Chromosome |
Marker or interval |
LODa
|
Additive effectb
|
|
GPC (%) |
3B |
Barc77 |
3.5 |
−0.17 |
|
5B |
Gwm499 |
4.3 |
−0.19 |
|
|
GH (%) |
1B |
WMC719 – WMC367-1 |
4.6 |
+1.75 |
|
5A |
IWA6573 – IWA2363 |
4.2 |
+1.44 |
Table 4Relative contribution mean of alleles to the grain protein content (GPC) and grain hardness (GH) in four environments.
Table 4
|
Trait |
Chromosome |
Marker |
Allele Aa
|
SEb of the allele A |
Allele B |
SE of the allele B |
|
GPC (%) |
3B |
Barc77 |
14.5 |
0.1 |
14.2 |
0.1 |
|
5B |
Gwm499 |
14.5 |
0.1 |
14.2 |
0.1 |
|
|
GH (%) |
1B |
WMC367-1 |
68.9 |
0.5 |
71.5 |
0.7 |
|
5A |
IWA6573 |
69.0 |
0.6 |
71.5 |
0.6 |
References
- Araki E, Miura H, Sawada S. 1999. Identification of genetic loci affecting amylase content and agronomic traits on chromosome 4A of wheat. Theor Appl Genet. 98: 977-984.
- Campbell KG, Bergman CJ, Gualberto DG, Anderson JA, Giroux MJ, Hareland G, Fulcher RG, Sorrells ME, Finney PL. 1999. Quantitative trait loci associated with kernel traits in a soft X hard wheat cross. Crop Sci. 39: 1184-1195.
- D’Ovidio R, Anderson OD. 1994. PCR analysis to distinguish between alleles of a member of a multigene family correlated with wheat bread-making quality. Theor Appl Genet. 88: 759-763.
- Dubcovsky J, Lijavetzky D, Appendino L, Tranquilli G. 1998. Comparative RFLP mapping of Triticum monococcum genes controlling vernalization requirement. Theor Appl Genet. 97: 968-975.
- Groos C, Robert N, Bervas E, Charmet G. 2003. Genetic analysis of grain protein-content, grain yield and thousand-kernel weight in bread wheat. Theor Appl Genet. 106: 1032-1040.
- Groos C, Bervas E, Charmet G. 2004. Genetic analysis of grain protein content, grain hardness and dough rheology in a hard X hard bread wheat progeny. J Cereal Sci. 40: 93-100.
- Korzun V, Röder MS, Worland AJ, Börner A. 1997. Intrachromosomal mapping of genes for dwarfing (Rht12) and vernalization response (Vrn1) in wheat by using RFLP and microsatellite markers. Plant Breed. 116: 227-232.
- Law CN, Young CF, Brown JW, Snape JW, Worland AJ. 1978. The study of grain protein control in wheat using whole chromosome substitution lines. Seed protein improvement by nuclear techniques. International Atomic Energy Agency. Vienna, Austria: pp. 483-502.
- Luo MC, Dvorak J. 1996. Molecular mapping of aluminium tolerance locus on chromosome 4D of Chinese Spring wheat. Euphytica. 91: 31-35.
- Morris R, Schmidt JW, Mattern PJ, Johnson VA. 1973. Chromosomal locations of genes for high protein in the wheat cultivar & Atlas 66′. Sears ER, Sears LMS, editors. Proc 4th Int Wheat Genet Symp. University of Missouri. Columbia, Mo: pp. 715-718.
- Morrison WR, Law CN, Wylie LJ, Coventry AM, Seekings J. 1989. The effect of group 5 chromosomes on the free polar lipids and breadmaking quality of wheat. J Cereal Sci. 9: 41-51.
- Nelson JC, Van-deynze AE, Autrique E, Sorrells ME, Lu YH. 1995. Molecular mapping of wheat: Homoeologous group 2. Genome. 38: 516-524.
- Parker G, Ken C, Anthony R, Peter L. 1997. Identification of molecular markers linked to flour colour and milling yield in wheat (Abstr. 246). In: 5th Int. Congr. Plant Mol. Biol; 21-27 Sept. 1997; Singapore.
- Prasad M, Varshney RK, Kumar A, Balyan HS, Sharma PC, Edwards KJ, Singh H, Dhaliwal HS, Roy JK, Gupta PK. 1999. A microsatellite marker associated with a QTL for grain protein content on chromosome are 2DL of bread wheat. Theor Appl Genet. 99: 341-345.
- Rogers WJ, Sayers EJ, Ru KL. 2001. Deficiency of individual high molecular weight glutenin subunits affords flexibility in breeding strategies for bread-making quality in wheat Triticum aestivum L. Euphytica. 117: 99-109.
- Shewry PR, Tatham AS, Lazzeri P. 1997. Biotechnology of wheat quality. J Sci Food Agric. 73: 397-406.
- Snape J, Semikhodskii WA, Sarma R, Korzun V, Fish L, Quarrie SA, Gill BS, Sasaki T, Galiba G, Sutka J. 1998. Mapping vernalization loci in wheat and comparitive mapping with other cereals. Slinkard AE, editor. Proc 9th Int Wheat Genet Symp. 3: pp. 156-158. Univ. Extension Press. Univ. of Saskatchewan, Saskatoon. 38.
- Stein Ira S, Sears RG, Gill BS, Hoseney RC, Cox TS. 1992. Heterogeneity of the ‘Wichita’ wheat monosomic set for grain quality and agronomic traits. Crop Sci. 32: 581-584.
- Sourdille P, Perretant MR, Charmet G, Leroy P, Gautier MF, Joudrier P, Nelson JC, Sorrells ME, Bernard M. 1996. Linkage between RFLP markers and genes affecting kernel hardness in wheat. Theor Appl Genet. 93: 580-586.
- Symes KJ. 1965. The inheritance of grain hardness in wheat as measured by the particle size index. Aust J Agric Res. 16: 113-123.
- Turner AS, Bradburne RP, Fish L, Snape JW. 2004. New quantitative trait loci influencing grain texture and protein content in bread wheat. J Cereal Sci. 40: 51-60.
- Uauy C, Distelfeld A, Fahima T, Blechl A, Dudcovsky J. 2006. A NAC gene regulation senescence improves grain protein, zinc, and iron content in wheat. Science. 314: 1298-1301.
- Zeng ZB. 1993. The theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci. Proc Natl Acad Sci USA. 90: 10972-10976.
- Zeng ZB. 1994. Precision mapping of quantitative trait loci. Genetics. 136: 14-1457.