Abstract
Cabbage (Brassica oleracea) is a very important vegetable worldwide and biotic stress is a crucial issue for this crop. Enhancement of resistance by exploiting stress resistance-related genes offers the most efficient approach to address this issue. Among the stress resistance-related genes, thaumatin-like proteins (TLPs) play a vital role in enhancement of resistance against stresses. In this study, we identified 12 TLPs from B. oleracea genomic DNA sequencing database, analyzed their sequences and compared with other published pathogenesis-related TLPs, and found a high degree of homology with them. In addition, these genes showed an organ-specific expression, three of which expressed differentially after Pectobacterium carotovorum subsp. carotovorum infection in cabbage plants. Data obtained in this study suggest the probable involvement of TLPs in resistance against soft rot disease of Brassica.
-
Key words: Brassica, Gene expression, Pectobacterium carotovorum subsp. carotovorum, Thaumatin-like protein
INTRODUCTION
Brassica crops are sternly challenged by various fungal, bacterial, and viral diseases and insect pests which led to enormous losses of yield.
Brassica vegetables are highly susceptible to bacterial soft rot disease, which is caused by a gram-negative bacterium,
Pectobacterium carotovorum subsp.
carotovorum (
Pcc). Bacterial soft rot is the most severe and destructive disease
, which causes serious damage and economic losses across the members of
Brassica. Control of this pathogen is difficult due to its ability to infect a wide range of hosts and to survive on plant debris. Effective chemical controls are not available so far. Cultural practices, including raised planting beds, reduced plant density, and delayed planting dates can reduce disease frequency and progression, but may reduce agricultural efficiency (
Fritz and Honma 1987). Genetic tolerance or resistance may represent an ideal alternative approach. But the traditional recurrent selection procedure is labor intensive and time-consuming. Moreover, due to lack of natural resistant genes against soft rot disease, the proceeding of routine raising disease-resistance breeding is restricted (
Li 1995). Therefore, genetic engineering of soft rot disease tolerance in
Brassica is of significant interest to agricultural biotechnology.
Plants respond to biotic stress factors by producing various defense-related proteins such as pathogenesis-related (PR) proteins, hydroxyproline-rich glycoproteins (HRGPs), and several key enzymes involved in the synthesis of phenylpropanoids, phytoalexins and lignins (
Collinge and Slusarenko 1987;
Bowles 1990;
Dixon and Lamb 1990). One promising strategy to develop resistance against biotic stresses is based on the exploitation of PRs and other stress responsive genes in transgenic plants. Accumulation of PRs extends into non-inoculated plant parts that, upon challenge, exhibit acquired resistance (
Van Loon and Van Kammen 1970;
Ryals et al. 1996). PR proteins are classified into 17 families based on sequence similarity and biochemical functions (
Van loon et al. 2006). An important common feature of most PR proteins is their antifungal effect. In addition, some PR proteins exhibit antibacterial, insecticidal, nematicidal, and anti-viral action (
Van Loon and Van Strien 1999;
Selitrennikoff 2001;
Van Loon 2001).
Owing to the sequence homology between PR-5 proteins and thaumatin, an intensely sweet-tasting protein isolated from the fruit of the West African rainforest shrub
Thaumatococcus daniellii (Bennett) Benth (
Iyengar et al. 1979), members of this family of proteins are referred to as thaumatin-like proteins (TLPs). This family also includes the closely related proteins permatin, osmotin and zeamatin. TLPs play a variety of roles including development of seed, fruit and flower tissues, and specific TLPs have been shown to protect plants against osmotic stress, pathogen attack and the deleterious effects of freezing (
Velazhahan et al. 1999;
An lovar and Dermastia 2003). The expression of some TLPs is induced by biotic and abiotic factors, such as microbial infection, osmotic stress, abscisic acid (ABA), ethylene, salicylic acid (SA), methyl jasmonate (MJ) and elicitors (
Kitajima and Sato 1999;
Velazhahan et al. 1999). Purified TLPs from several plant species have
in vitro anti-fungal activity. Several antifungal modes of action have been described such as membrane permeabilization (
Vigers et al. 1991), β-glucan binding and degradation (
Sakamoto et al. 2006), inhibition of enzymes such as xylanases (
Fierens et al. 2007), α-amylase, or trypsin (
Schimoler-O’Rourke et al. 2001), as well as an apoptosis-inducing mechanism reported in yeast (
Narasimhan et al. 2005). A PR5 protein from
Arabidopsis thaliana ATLP3, showed considerable antifungal activity against pathogenic fungi such as
Verticillium alboatrum,
V. dahliae and
Fusarium oxysporum (
Hu and Ready 1997). PR5 proteins from mutant barley grain inhibited growth of
Trichoderma viride and
Candida albicans (
Hejgaard et al. 1991). In addition to inhibiting hyphal growth and spore germination, some TLPs greatly increase the permeability of fungal and oomycete plasma membranes (
Yun et al. 1997;
Kitajima and Sato 1999). Overexpression of TLPs appears to enhance resistance to fungal disease (
Liu et al. 1994;
Zhu et al. 1996;
Chen et al. 1999). Moreover, TLPs in various plant-derived foods (
Hoffmann-Sommergruber 2002) and conifer pollen (
Midoro-Horiuti et al. 2000;
Cortegano et al. 2004) are allergens.
In our previous study, we characterized 20 thaumatin-like genes of
B. rapa and found four genes to respond after
P. carotovorum subsp.
carotovorum infection in Chinese cabbage (
Ahmed et al. 2013). In relation to the previous study, we identified 12 thaumatin-like genes of
B. oleracea from
Brassica oleracea Genome Database (Bolbase) (
http://119.97.203.210/bolbase/index.html). We then analyzed the sequences and studied their homology with pathogenesis-related thaumatin-like proteins of other species. In addition, expression of these genes was analyzed in different organs and after infection with
P. carotovorum subsp.
carotovorum in cabbage plants. We also discussed their association with biotic stress resistance in
Brassica.
MATERIALS AND METHODS
Plant materials
Cabbage (Brassica oleracea ‘ASC621’) plants were grown at the Department of Horticulture, Sunchon National University, Korea. Fresh roots, stems, leaves and flower buds of the Chinese cabbage were harvested, immediately frozen in liquid nitrogen, and then stored at −80°C until subsequent analysis in an organ-specific expression study.
Stress treatments
Chinese cabbage (B. oleracea ‘ASC621’) plants were grown for 6 weeks under culture room conditions with 16 h light and 8 h dark maintaining 25°C temperature prior to treatment. The P. carotovorum subsp. carotovorum stock (10 μl) was cultured in 25 ml of liquid YEP medium until OD600 reading is 1.4 or 1,170, cfu ml−1 and then diluted it to OD600 = 1.19 or 1×106 CFU ml−1 by adding double-distilled water. For pathogen inoculation, 10 μl of P. carotovorum subsp. carotovorum culture solution (1×106 cfu ml−1) was added to the freshly needle-wounded site (at the lower 1/3 position) of the midrib of the upper third leaves, and incubated at 25°C covering with polyvinyl bags to maintain 80–90% humidity. All inoculations were performed three times, and the infection was confirmed by observing disease lesion in the leaves of Chinese cabbage plants. About one-third part from the top of the infected leaves were harvested for RNA extraction at 0, 6, 12, 24, 48 and 72 h after inoculation, frozen in liquid nitrogen, and stored at −80°C.
RNA extraction
Total RNA of previously frozen roots, stems, leaves, flower buds and stress treated samples with Rneasy mini kit (Qiagen, USA) was extracted and treated with RNase-free DNase (Promega, USA) to remove genomic DNA contaminants. The cDNA was subsequently synthesized using a Superscript® III First-Strand synthesis kit (Invitrogen, USA) according to the manufacturer’s instructions.
Sequence analysis
Expression analysis
RT-PCR was conducted using an AMV one step RT-PCR kit (Takara, Japan). Specific primers for all the genes were used for RT-PCR, and actin primers of
Brassica rapa (FJ969844) were used as control (
Table 1). PCR was performed using 50 ng of cDNA from the roots, leaves, stems and flower buds as templates in master mixes composed of 20 pmol of each primer, 150 μM of each dNTP, 1.2 U of
Taq polymerase, 1x
Taq polymerase buffer, and double-distilled H
2O diluted to a total volume of 20 μl in 0.5 ml PCR tubes. The samples were then subjected to the following conditions: pre-denaturing at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s and extension at 72°C for 1 min, with a final extension for 5 min at 72°C. Real-time quantitative PCR was performed using 1 μl of cDNA in a 25 μl reaction volume employing iTaq
TM SYBR
® Green Super-mix with ROX (California, USA). The specific primers used for real-time PCR are listed in
Table 1. The conditions for real-time PCR were as follows: 10 min at 95°C, followed by 40 cycles at 94°C for 30 s, 58°C for 30 s, and 72°C for 45 s. The fluorescence was measured following the last step of each cycle, and three replications were used for each sample. Amplification, detection, and data analysis were conducted using a Rotor-Gene 6000 real-time rotary analyzer (Corbett Life Science, Australia).
RESULTS AND DISCUSSION
Identification and sequence analysis
In this study, 12 thaumatin-like proteins, designated
B. oleracea Thaumatin-like Protein (BoTLP) were identified and their sequences were analyzed and presented in the
Table 2. The size of the 12 BrTLPs varies from 133 to 428 amino acids (~14.66 to 45.33 kDa), and the predicted isoelectric points vary from 4.25 to 9.07. Analysis of the protein domain organization showed that all these proteins contained the characteristic thaumatin (THN) domain in the conserved region located mostly from 1 to 190 amino acid positions of the protein sequences which covers almost 80 to 90% of the entire mature TLPs. Genomic DNA sequences pertaining structural information of all 12 genes were collected from
Brassica oleracea Genome Database (Bolbase) (
http://119.97.203.210/bolbase/index.html). Most typical TLPs described to date have a molecular weight ranging from 20 to 26 kDa, and generally possess 16 conserved cysteine residues that form eight disulfide bonds (
Ghosh and Chakrabarti 2008). Several TLP structures have been solved so far, revealing a strongly conserved 3D organization with a characteristic acidic cleft domain that comprises the five highly conserved amino acids REDDD that are dispersed in the primary sequence (
Koiwa et al. 1999). Despite good conservation of these amino acids in primary sequences, they do not organize into an acidic cleft at the 3D level (
Vandermarliere et al. 2010). The BoTLPs also contained 16 cysteine residues, REDDD and FF hydrophobic motif conserved in an identical position with other published TLPs and sweet tasting thaumatin protein (
Fig. 2A and B). Alongside, it has become evident that several families of cysteine-rich basic antimicrobial polypeptides play an important role in protecting plants against invading phytopathogens (
Garcia-Olmedo et al. 1998). These sequence data revealed that the identified genes are thaumatin-like genes.
For comparison of these
BoTLPs with other published TLPs, an alignment search was carried out using a BLAST search of the NCBI database. The deduced amino acid sequences of 12 BoTLPs shared high homology, primarily with pathogenesis-related TLPs of
A. thaliana and some other homologous species (
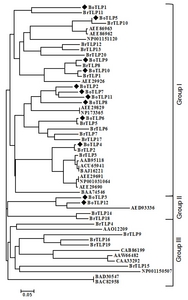
Table 3). We again retrieved 41 pathogenesis-related TLP sequences of different plant species from NCBI and a phylogenetic tree was constructed with the deduced amino acid sequences of 12 TLPs of
B. oleracea using the NJ method (
Fig. 1). In the phylogenetic tree, 12 BoTLPs were divided into two main groups showing close relations with the PR TLP sequences of other plant species. Multiple alignment using PIR also revealed high similarity among the deduced amino acids of the TLPs of the two groups and the thaumatin protein of
T. daniellii (Bennett) Benth (P02884), indicating their relatively conserved evolutionary relationship at the protein level (
Fig. 2A and B). Thus, our results showed that the
BoTLP genes showed high similarity with other PR TLPs and might play roles in defense activities against phytopathogens.
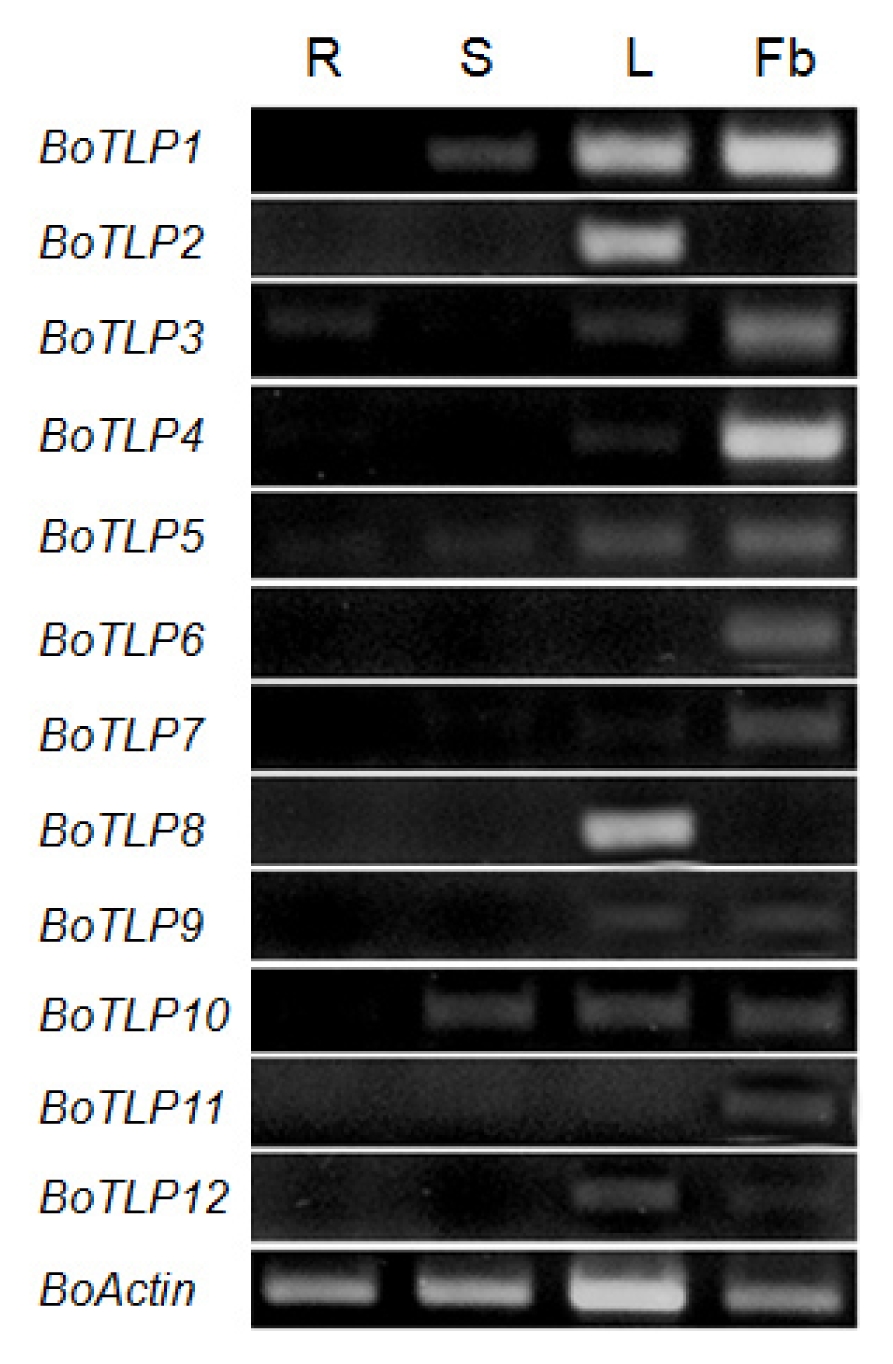
Gene expression in different organ of cabbage was carried out by RT-PCR and varying expression pattern was observed (
Fig. 3). Specifically,
BoTLP1 and
10 showed expressions in all the organs tested except roots.
BoTLP2 and
8 showed high level of expression only in leaves, while
BoTLP3 showed expression in all organs except stem.
BoTLP4, 7, 9 and
12 expressed only in the leaves and flower buds, while only
BoTLP5 expressed in all organs tested.
BoTLP6 and
11 expressed only in flower buds, indicating their probable role as allergen genes. Obviously, all genes described here showed higher level of expressions in leaves and flower buds compared to roots and stems. TLPs are now recognized as a new class of panallergens in food as well as in pollen (
Breiteneder 2004). Moreover, the organs tested in this study share characteristics that make them particularly prone to pathogen attack. Adequate protection of these organs against pathogen attack is critical, and the production of an antifungal molecule could be part of a local defense strategy (
Hamel and Bellemare 1995).
Because EST analysis and previous studies indicated that thaumatin-like genes are biotic stress-related genes, we investigated the responses of the 12
BoTLPs identified in this study to infection by soft rot disease causing necrotroph bacteria,
P. carotovorum subsp.
carotovorum, which is a serious pathogen for
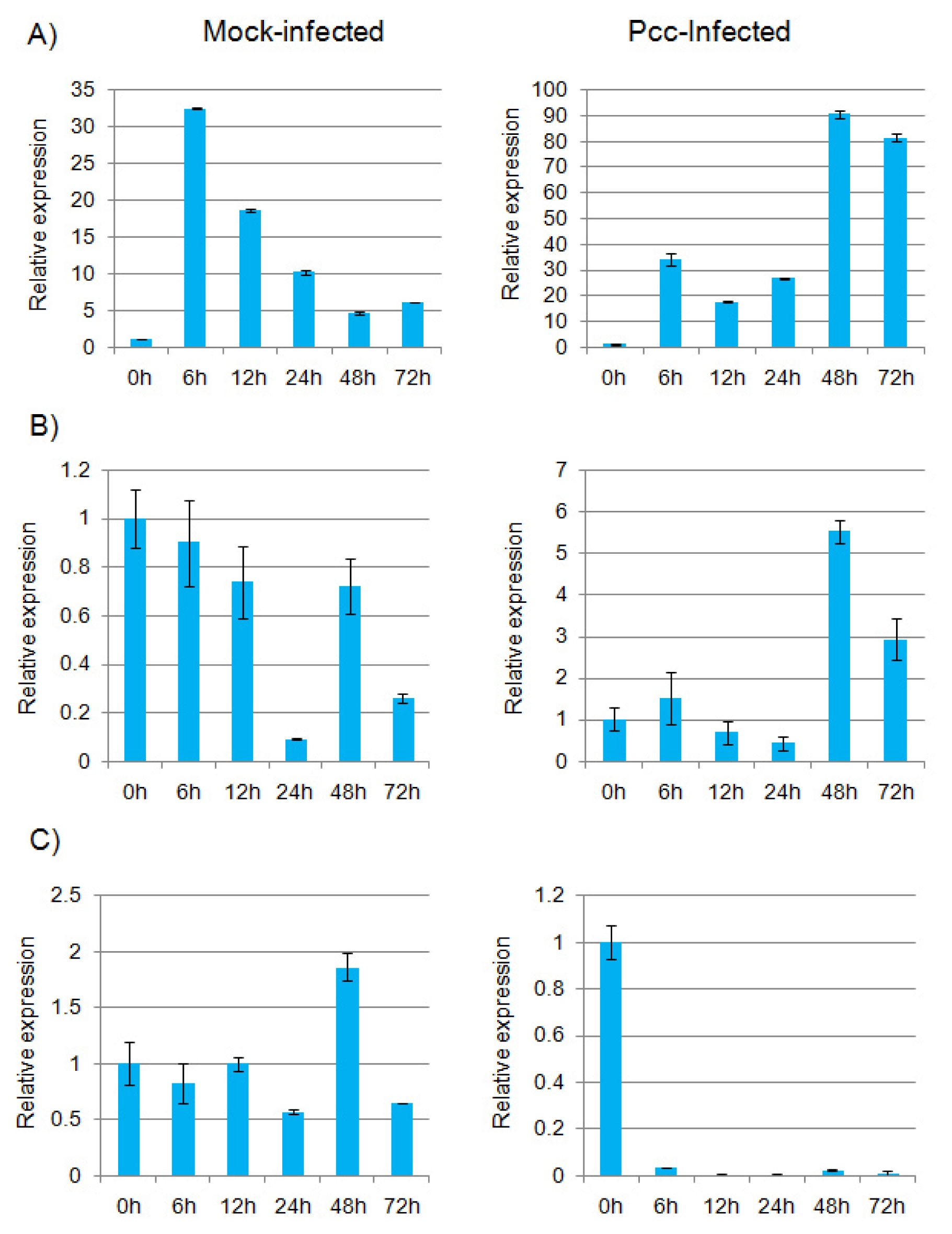
Brassica. In the infected plants, expression of
BoTLP5, BoTLP8 and
BoTLP12 was differentially changed compared to mock-infected plants (
Fig. 4A, B and C).
BoTLP5 and
BoTLP8 showed increased level of transcripts after 48 h of infection but decreased after 72 h (
Fig. 4A and B). Interestingly, the expression of
BoTLP5 and
BoTLP8 in mock-infected plants was higher at the earlier periods of treatment than the later. Accordingly, the expression was 19-fold higher for
BoTLP5 and 5-fold higher for
BoTLP8, in the Pcc-infected plants than in mock-infected plants at 48 h. On the other hand, transcript level of
BoTLP12 was abruptly decreased just after infection and became almost inactive (
Fig. 4C). It seems that only
BoTLP5, 8 and 12 were induced among the
BoTLPs after
P. carotovorum subsp.
carotovorum infection and possibly other genes might be responsive to other biotic stress agents. Thus, the results suggest that
BoTLPs are PR TLPs and have probable involvement with biotic stress resistance.
Previous molecular studies of TLP expression, localization and activity support a role for TLPs in host defense during pathogen infection. TLP up-regulation has been described in many higher plants infected by pathogens such as bacteria, oomycetes and fungi (
Liu et al. 2010;
Mukherjee et al. 2010). It has also been suggested that the expression of PR-5 proteins (TLPs) contributes to resistance to different fungi (
Vleeshouwers et al. 2000;
Monot et al. 2002;
Suo and Leung 2002). A PR5 thaumatin-like protein from
A. thaliana ATLP3 expressed in bacteria showed considerable antifungal activity against pathogenic fungi such as
V. alboatrum,
V. dahlia, F. oxysporum and
A. solani, as well as non-pathogenic fungi including
T. reesei and
C. albicans (
Hu and Reddy 1997). Overexpression of cotton (
Gossypium barbadense L.) thaumatin-like protein gene (
GbTLP1) in tobacco exhibited considerable resistance against
V. dahliae and
F. oxysporum (
Munis et al. 2010). Transgenic tobacco plants with thaumatin gene exhibited enhanced resistance with delayed disease symptoms against fungal diseases caused by
Pythium aphanidermatum and
Rhizoctonia solani (
Rajam et al. 2007). A barley thaumatin-like protein (
Hv-TLP8) in transgenic oilseed rape (
B. napus) exhibited enhanced resistance to the clubroot pathogen (
Plasmodiophora brassicae) (
Reiss et al. 2009). Taken together, the above discussion suggests the probable association of these genes with biotic stress resistance.
In this study BoTLP5, 8 and 12 responded positively after P. carotovorum subsp. carotovorum infection and for that it is mentionable that these genes could be considered as useful resources for biotic stress resistance. Thus, BoTLP5, 8 and 12 might be promising candidate for engineering Brassica resistant to biotic stresses. The remaining genes also might be useful for other biotic stresses resistance in Brassica, thus, investigations involving screening of these genes with other pathogens is recommended.
ACKNOWLEDGEMENTS
This work was supported by a grant from the Next-Generation BioGreen 21 Program (Plant Molecular Breeding Center No. PJ009085022012), Rural Development Administration, Republic of Korea.
Fig. 1Phylogenetic tree showing the relatedness of the deduced full-length amino acid sequences of 12
Brassica oleracea thaumatin-like proteins (BoTLPs) and 41 PR thaumatin-like proteins of other plant species. The phylogenetic tree was generated using MEGA5.05 program. The species names and the GenBank protein accessions numbers are indicated in brackets.
Brassica oleracea (AAO12209),
Brassica rapa (AAB95118),
Brassica rapa subsp.
chinensis (ACU65941 and BAJ16221),
Oryza sativa Japonica Group (BAD30547 and BAC82958),
Arabidopsis thaliana (AEE29691, AEE29690, NP001031064, AEE86963, AEE86962, AEE29829, NP173365, AED93336 and AEE29926),
Solanum lycopersicum (CAB86199),
Nicotiana tabacum (AAW66482, CAA33292 and BAA74546),
Zea mays (NP001150507 and NP001151120) and BrTLP1 to 20 were taken from our previous study (
Ahmed et al. 2013). The scale represents the frequency of amino acid substitution between sequences as determined by poisson evolutionary distance method.
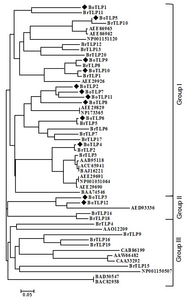
Fig. 2Alignment of deduced amino acid sequences of (A) Group I thaumatin-like proteins; (B) Group II thaumatin-like proteins and sweet-tasting thaumatin protein of
Thaumatococcus daniellii (Bennett) Benth. (P02884) in each group using PIR. Asterisk marks show all 16 conserved cysteine residues, square bullets indicate REDDD motif in the highly conserved region (rectangle) of the domain and circular bullets indicate FF hydrophobic motif described by
Petre et al. (2011) in identical positions among the aligned proteins. Numbers on the right margin indicate the positions of amino acid residues. The identical amino acids are marked in dark background, and more than 50% similarities in light.

Fig. 3RT-PCR expression analysis of 12 BoTLP genes over different organs at Brassica oleracea plants. Lanes 1–4, PCR products of roots (R), stems (S), leaves (L) and flower buds (Fb)
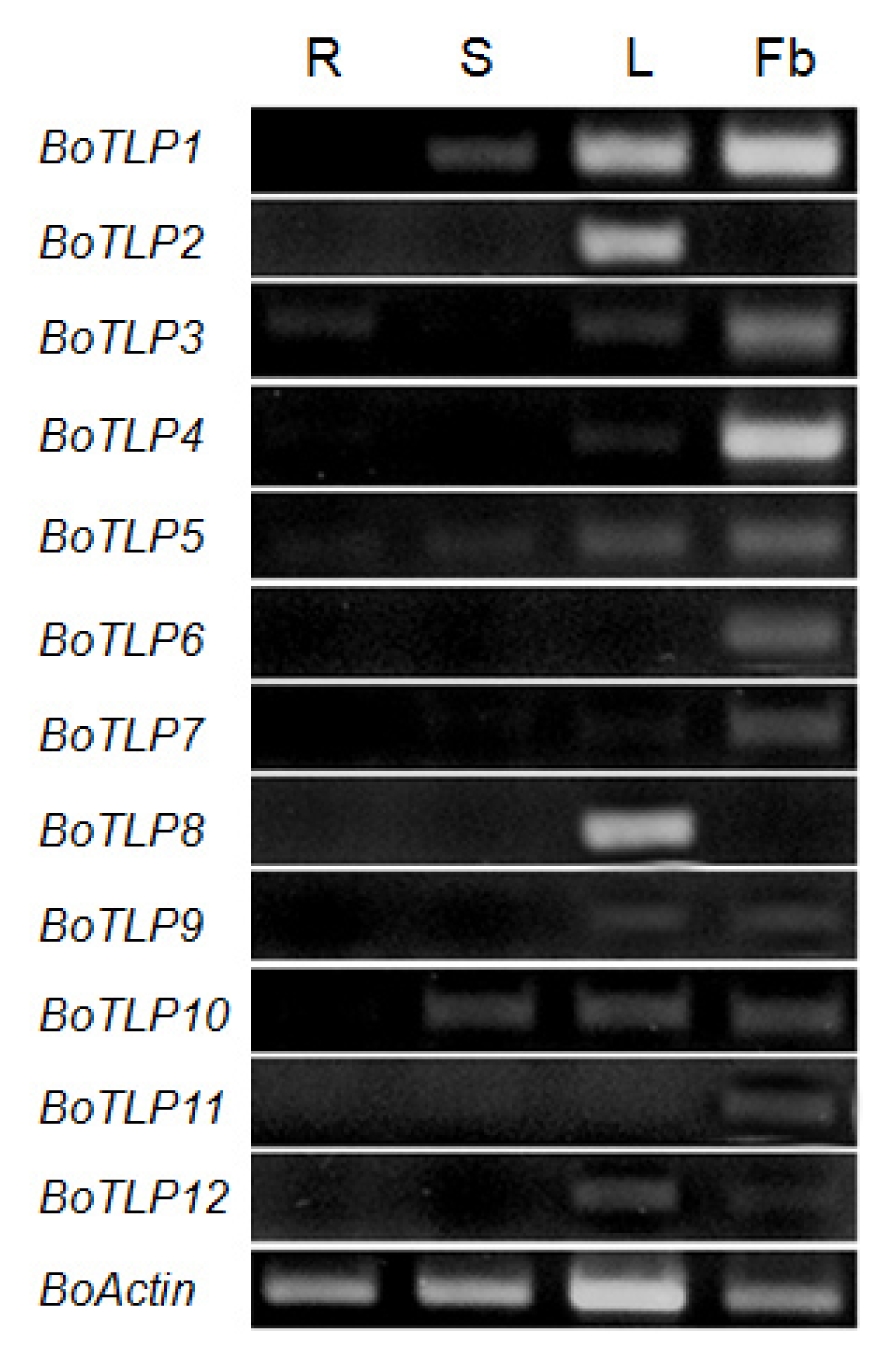
Fig. 4Real-time quantitative PCR expression analysis of (A) BoTLP5, (B) BoTLP8 and (C) BoTLP12 genes in Brassica oleracea after infection with Pectobacterium carotovorum subsp. carotovorum (Pcc). The error bars represent the standard error of the means of three independent replicates.
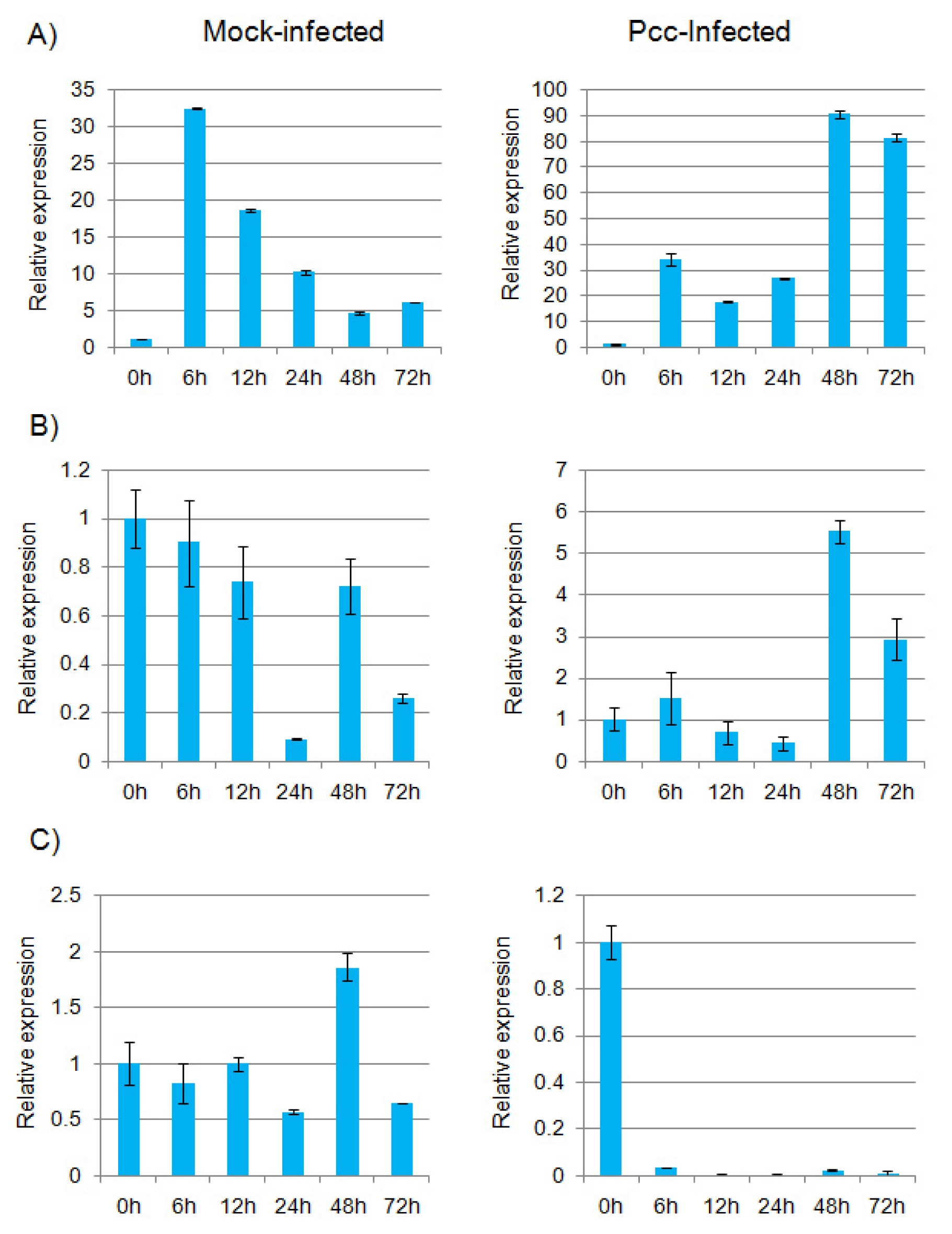
Table 1Primer information of 12 thaumatin-like genes of B. oleracea for RT and Real time PCR.
Table 1
|
Gene name |
Acc. No. of ESTs |
Primer sequence |
|
|
Forward |
Reverse |
|
BoTLP1
|
BOLC06439 |
CAGAAATCAACAAGCTACCC |
GATCCGTCTTGTAACATCGT |
|
BoTLP2
|
BOLC11620 |
TTTTCACCATCCAGAATAGC |
GTCGAAGTTACAGCCTGTTC |
|
BoTLP3
|
BOLC12781 |
TCTACCGTTTTCAAGAGAGC |
CCGTACGGATTCTGAGTTAG |
|
BoTLP4
|
BOLC13543 |
CTACTCTCGCAGAGATCACC |
GAGTTAAAGGCAGAACATGC |
|
BoTLP5
|
BOLC14356 |
CCCTGAAAGAAGAATGAGAA |
ACAAGACGAGCACCTACACT |
|
BoTLP6
|
BOLC17729 |
ACGACTTACTCGATGACGTT |
ATCTTCCGTACATAGCAAGC |
|
BoTLP7
|
BOLC20473 |
CCGCAAAAGCTCTTTATTAC |
TATAGCTACGCATACGACGA |
|
BoTLP8
|
BOLC20492 |
ATCCAGATGAGAATCACGAC |
ACAAGTCTCCGGTTTATTGA |
|
BoTLP9
|
BOLC23111 |
GTAGAGACGGGAACAAAACA |
AAAGTTGCAGTGAAAAGCTC |
|
BoTLP10
|
BOLC23113 |
ACGGGAGGATTCTTAATTGT |
CAACGCAGCATTAGATAACA |
|
BoTLP11
|
BOLC23820 |
CGTCCGGATTAATACTTGAG |
AGTTGTACACCGTTGTCTCC |
|
BoTLP12
|
BOLS14849 |
AGCAAAATCCATATCACAGG |
GACTGTCCCACATCAAGTTT |
|
Br-Actin
|
FJ969844 |
CAACCAATCGTCTGTGACAA |
ATGTCTTGGCCTACCAACAA |
Table 2Overall analysis of thaumatin-like genes collected from the
Brassica oleracea database.
z)
Table 2
|
Name of genes |
Accession Number |
ORF (bp) |
Chromosome number |
Protein |
|
|
Length (aa) |
THN domain Start and end |
Mol.Wt. (KD) |
pI |
|
BoTLP1
|
Bol016062 |
1041 |
C03 |
346 |
1 to 184 |
35.58 |
4.25 |
|
BoTLP2
|
Bol044835 |
738 |
C08 |
245 |
1 to 185 |
25.32 |
5.05 |
|
BoTLP3
|
Bol022411 |
1098 |
C07 |
365 |
1 to 189 |
38.56 |
5.31 |
|
BoTLP4
|
Bol026250 |
738 |
C06 |
245 |
1 to 181 |
26.15 |
9.07 |
|
BoTLP5
|
Bol000789 |
825 |
Scaffold |
274 |
1 to 185 |
29.22 |
8.91 |
|
BoTLP6
|
Bol026131 |
732 |
C06 |
243 |
1 to 180 |
25.63 |
4.62 |
|
BoTLP7
|
Bol026844 |
741 |
C05 |
246 |
1 to 184 |
25.46 |
4.83 |
|
BoTLP8
|
Bol044836 |
402 |
C08 |
133 |
1 to 72 |
14.66 |
8.29 |
|
BoTLP9
|
Bol027694 |
978 |
C06 |
325 |
1 to 185, trnsm 1 |
34.32 |
4.86 |
|
BoTLP10
|
Bol039281 |
963 |
C02 |
320 |
1 to 187 |
33.73 |
4.67 |
|
BoTLP11
|
Bol026846 |
747 |
C05 |
248 |
1 to 186 |
25.88 |
4.63 |
|
BoTLP12
|
Bol016440 |
1287 |
C02 |
428 |
1 to 190 |
45.33 |
4.95 |
Table 3Homology analysis of 12 thaumatin-like genes of
Brassica oleracea z)
References
- Ahmed NU, Park J, Jung H, Kang K, Lim Y, Hur Y, Nou I. 2013. Molecular Characterization of Thaumatin Family Genes Related to Stresses in Brassica rapa. Scientia Horticulturae. 152: 26-34.
- An lovar S, Dermastia M. 2003. The comparative analysis of osmotins and osmotin-like PR5 proteins. Plant Biol. 5: 116-124.
- Bowles DJ. 1990. Defense-related proteins in higher plants. Annu Rev Biochem. 59: 873-907.
- Breiteneder H. 2004. Thaumatin-like proteins-a new family of pollen and fruit allergens. Allergy. 59: 479-481.
- Chen WP, Chen PD, Liu DJ, Kynast R, Friebe B, Velazhahan R, Muthukrishnan S, Gill BS. 1999. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor Appl Genet. 99: 755-760.
- Collinge DB, Slusarenko AJ. 1987. Plant gene expression in response to pathogens. Plant Mol Biol. 9: 389-410.
- Cortegano I, Civantos E, Aceituno E, del Moral A, Lopez E, Lombardero M, del Pozo V, Lahoz C. 2004. Cloning and expression of a major allergen from Cupressus arizonica pollen, Cup a 3, a PR-5 protein expressed under polluted environment. Allergy. 59: 485-490.
- Dixon RA, Lamb CJ. 1990. Molecular communication in interactions between plants and microbial pathogens. Annu Rev Plant Physiol Plant Mol Biol. 41: 339-367.
- Fierens E, Rombouts S, Gebruers K, Goesaert H, Brijs K, Beaugrand J, Volckaert G, Van Campenhout S, Proost P, Courtin CM, Delcour JA. 2007. TLX1, a novel type of xylanase inhibitor from wheat (Triticum aestivum) belonging to the thaumatin family. Biochem J. 403: 583-591.
- Fitt B, Brun H, Barbetti M, Rimmer S. 2006. World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). Eur J Plant Pathol. 114: 3-15.
- Fritz VA, Honma S. 1987. The effect of raised beds, population densities, and planting date on the incidence of bacterial in Chinese cabbage. J Am Soc Hortic Sci. 112: 41-44.
- Futamura N, Tani N, Tsumura Y, Nakajima N, Sakaguchi M, Shinohara K. 2006. Characterization of genes for novel thaumatin-like proteins in Cryptomeria japonica. Tree Physiol. 26: 51-62.
- Gaetan SA. 2005. Occurrence of Fusarium wilt on canola caused by Fusarium oxysporum F.sp. conglutinas in Argentina. Plant Dis. 89: 432
- Garcia-Olmedo F, Molina A, Alamillo JM, Rodriguez-Palenzuela P. 1998. Plant defense peptides. Biopolymers. 47: 479-491.
- Ghosh R, Chakrabarti C. 2008. Crystal structure analysis of NP24-I: a thaumatin-like protein. Planta. 228: 883-890.
- Hamel F, Bellemare G. 1995. Characterization of a class I chitinase gene and of wound-inducible, root and flower-specific chitinase expression in Brassica napus. Biochim Biophys Acta. 1263: 212-220.
- Hejgaard J, Jacobsen S, Svendsen I. 1991. Two antifungal thaumatin-like proteins from barley grain. FEBS Lett. 291: 127-131.
- Hoffmann-Sommergruber K. 2002. Pathogenesis-related (PR)-proteins identified as allergens. Biochem Soc Trans. 30: 930-935.
- Howlett BJ, Indurm A, Pedras MSC. 2001. Leptosphaeria maculans, the causal agent of blackleg disease of Brassica. Fungal Genet Biol. 33: 1-14.
- Hu X, Reddy AS. 1997. Cloning and expression of a PR5-like protein from Arabidopsis: inhibition of fungal growth by bacterially expressed protein. Plant Mol Biol. 34: 949-959.
- Iyengar RB, Smits P, Van der Ouderaa F, Van der Wel H, Van Brouwershaven J, Ravestein P, Richters G, Van Wassenaar PD. 1979. The complete amino-acid sequence of the sweet protein thaumatin. I Eur J Biochem. 96: 193-204.
- Kitajima S, Sato F. 1999. Plant pathogenesis-related proteins: molecular mechanisms of gene expression and protein function. J Biochem. 125: 1-8.
- Koiwa H, Kato H, Nakatsu T, Oda J, Yamada Y, Sato F. 1999. Crystal structure of tobacco PR-5d protein at 1,8 Å resolution reveals a conserved acidic cleft structure in antifungal thaumatin-like proteins. J Mol Biol. 286: 1137-1145.
- Li SD. 1995. Progress in disease resistant breed of main vegetables. Science Press. Beijing: pp. 96-100.
- Lin X, Kaul S, Rounsley S, Shea TP, Benito MI, Town CD, et al. 1999. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature. 402: 761-8.
- Liu D, Raghothama KG, Hasegawa PM, Bressan RA. 1994. Osmotin overexpression in potato delays development of disease symptoms. Proc Natl Acad Sci USA. 91: 1888-1892.
- Liu JJ, Sturrock R, Ekramoddoullah AKM. 2010. The superfamily of thaumatin-like proteins: its origin, evolution, and expression towards biological function. Plant Cell Rep. 29: 419-436.
- Mayer K, Schuller C, Wambutt R, Murphy G, Volckaert G. 1999. Sequence and analysis of chromosome 4 of the plant Arabidopsis thaliana. Nature. 402: 769-777.
- Midoro-Horiuti T, Goldblum RM, Kurosky A, Wood TG, Brooks EG. 2000. Variable expression of pathogenesis-related protein allergen in mountain cedar (Juniperus ashei) pollen. J Immunol. 164: 2188-2192.
- Monot C, Pajot E, Le Corre D, Silue D. 2002. Induction of systemic resistance in broccoli (Brassica oleracea var. botrytis) against downy mildew (Peronospora parasitica) by avirulent isolates. Biol Control. 24: 75-81.
- Mukherjee AK, Carp MJ, Zuchman R, Ziv T, Horwitz BA, Gepstein S. 2010. Proteomics of the response of Arabidopsis thaliana to infection with Alternaria brassicicola. J Proteomics. 73: 709-720.
- Munis M, Hussain F, Tu L, Deng F, Tan J, Xu L, Xu S, Long L, Zhang X. 2010. A thaumatin-like protein gene involved in cotton fiber secondary cell wall development enhances resistance against Verticillium dahliae and other stresses in transgenic tobacco. Biochem Biophys Res Comm. 393: 38-44.
- Narasimhan ML, Coca MA, Jin J, Yamauchi T, Ito Y, Kadowaki T, Kim KK, Pardo JM, Damsz B, Hasegawa PM, Yun DJ, Bressan RA. 2005. Osmotin is a homolog of mammalian adiponectin and controls apoptosis in yeast through a homolog of mammalian adiponectin receptor. Mol Cell. 7: 171-180.
- Ospina-Giraldo MD, Mullins E, Kang S. 2003. Loss of function of the Fusarium oxysporum SNF1 gene reduces virulence on cabbage and Arabidopsis. Curr Genet. 44: 49-57.
- Petre B, Major I, Rouhie N, Duplessis S. 2011. Genome-wide analysis of eukaryote thaumatin-like proteins (TLPs) with an emphasis on poplar. BMC Plant Biol. 11: 33
- Rajam MV, Chandola N, Goud Singh D, Kashyap V, Choudhary ML, Sihachakr D. 2007. Thaumatin gene confers resistance to fungal pathogens as well as tolerance to abiotic stresses in transgenic tobacco plants. Biol Plant. 51: 135-141.
- Reiss E, Schubert J, Scholze P, Kramer R, Sonntag K. 2009. The barley thaumatin-like protein Hv-TLP8 enhances resistance of oilseed rape plants to Plasmodiophora brassicae. Plant Breed. 128: 210-212.
- Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner HY, Hunt MD. 1996. Systemic acquired resistance. Plant Cell. 8: 1809-1819.
- Saitou N, Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 4: 406-425.
- Sakamoto Y, Watnabe H, Nagai M, Nakade K, Takahashi M, Sato T. 2006. Lentinula edodes tlg1 encodes a thaumatin-like protein that is involved in lentinan degradation and fruiting body senescence. Plant Physiol. 141: 793-801.
- Sato S, Kaneko T, Kotani H, Nakamura Y, Asamizu E, Miyajima N, Tabata S. 1998. Structural analysis of Arabidopsis thaliana chromosome 5. IV. Sequence features of the regions of 1,456,315 bp covered by nineteen hysically assigned P1 and TAC clones. DNA Res. 5(1): 41-54.
- Schimoler-O’Rourke R, Richardson M, Selitrennikoff CP. 2001. Zeamatin inhibits trypsin and α-amylase activities. Appl Env Microbiol. 67: 2365-2366.
- Selitrennikoff CP. 2001. Antifungal proteins. Appl Env Microbiol. 67: 2883-2894.
- Sherif S, Paliyath G, Jayasankar S. 2012. Molecular characterization of peach PR genes and their induction kinetics in response to bacterial infection and signaling molecules. Plant Cell Rep. 31(4): 697-711.
- Suo Y, Leung DWM. 2002. Accumulation of extracellular pathogenesis-related proteins in rose leaves following inoculation of in vitro shoots with Diplocarpon rosae. Sci Hortic. 93: 167-178.
- Tabata S, Kaneko T, Nakamura Y, Kotani H, Kato T, et al. 2000. Sequence and analysis of chromosome 5 of the plant Arabidopsis thaliana. Nature. 408(6814): 823-826.
- Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol. 24: 1596-1599.
- Theologis A, Ecker JR, Palm CJ, Federspiel NA. 2000. Sequence and analysis of chromosome 1 of the plant Arabidopsis thaliana. Nature. 408: 816-20.
- Van Loon LC, Rep M, Pieterse CM. 2006. Significance of inducible defense-related proteins in infected plants. Annu Rev Phytopathol. 44: 135-162.
- Van Loon LC, Van Kammen A. 1970. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. ‘Samsun’ and ‘Samsun NN’ II. Changes in protein constitution after infection with tobacco mosaic virus. Virology. 40: 199-211.
- Van Loon LC, Van Strien EA. 1999. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol. 55: 85-97.
- Van Loon LC. 2001. The families of pathogenesis-related proteins. In: 6th International Workshop on PR-proteins; May 20–24; Spa, Belgium. Book of abstracts. pp 9.
- Vandermarliere E, Lammens W, Schoepe J, Rombouts S, Fierens E, Gebruers K, Volckaert G, Rabijns A, Delcours JA, Strelkov SV, Courtin CM. 2010. Crystal structure of the noncompetitive xylanase inhibitor TLX1, member of the small thaumatin-like protein family. Proteins. 78: 2391-2394.
- Velazhahan R, Datta SK, Muthukrishnan S. 1999. The PR-5 family: thaumatin-like proteins. Datta SK, Muthukrishnan S, editors. Pathogenesis-related proteins in plants. CRC Press. Boca Raton, FL: pp. 107-129.
- Vigers AJ, Roberts WK, Selitrennikoff CP. 1991. A new family of plant antifungal proteins. Mol Plant Microbe Interact. 4: 315-23.
- Vleeshouwers VGAA, Van Dooijeweert W, Govers F, Kamoun S, Colon LT. 2000. Does basal PR gene expression in Solanum species contribute to non-specific resistance to Phytophthora infestans? Physiol Mol Plant Pathol. 57: 35-42.
- Yun DJ, Bressan RA, Hasegawa PM. 1997. Plant antifungal proteins. Plant Breed Rev. 14: 39-88.
- Zhu B, Chen THH, Li PH. 1996. Analysis of late-blight disease resistance and freezing tolerance in transgenic potato plants expressing sense and antisense genes for an osmotin-like protein. Planta. 198: 70-77.