Abstract
Microalgae offer a sustainable platform for biotechnology due to their rapid growth and ability to accumulate high-value metabolites such as lipids, proteins, and carbohydrates. This study investigates the biochemical and morphological responses of Chlorella vulgaris and Scenedesmus incrassatulus across different growth phases and under nitrogen (N) and phosphorus (P) deficiency. Molecular identification and phylogenetic analysis confirmed the taxonomic status of both species. Scanning Electron Microscopy (SEM) combined with Energy Dispersive X-ray Spectroscopy (EDS) revealed species-specific surface structures and elemental composition. S. incrassatulus exhibited a compact, granular morphology, while C. vulgaris showed a smoother, colonial structure. EDS analysis demonstrated higher potassium and phosphorus levels in C. vulgaris, whereas S. incrassatulus presented elevated calcium and magnesium contents. Biochemical profiling indicated a metabolic shift during the stationary phase, with reduced protein synthesis and increased lipid and carbohydrate accumulation. Under nitrogen deficiency, C. vulgaris achieved the highest lipid content (53.68%), while S. incrassatulus reached 45.86%. Carbohydrate accumulation was also prominent during the stationary phase, with values of 49.18% for C. vulgaris and 47.99% for S. incrassatulus. Protein content was highest in the exponential phase and declined under nutrient stress. Elemental analysis via Inductively Coupled Plasma-Atomic Emission Spectroscopy (ICP-AES) confirmed species-specific macronutrient and trace element profiles, supporting EDS data. These findings highlight the adaptive biochemical and structural traits of the two microalgae species, reinforcing their potential in biofuel production, sustainable agriculture, bioremediation, and environmental applications. Future work should focus on optimizing cultivation conditions and exploring genetic approaches to maximize metabolite yields for industrial-scale applications.
-
Key words: Microalgae, Chlorella vulgaris, Scenedesmus incrassatulus, Mineralogical composition, Primary metabolites, Growth phase, Nutrient deficiency
Introduction
Microalgae are microscopic, photosynthetic organisms that are widely distributed in aquatic and terrestrial ecosystems. They are considered vital for the global ecosystem due to their ability to capture carbon dioxide, produce oxygen, and synthesize a diverse range of biomolecules, including lipids, proteins, and carbohydrates (
Tripathi et al. 2023;
Udayan et al. 2023). These properties, coupled with their rapid growth rates and ability to adapt to various environmental conditions, make microalgae promising candidates for sustainable biotechnological applications such as biofuel production, wastewater treatment, and high-value biomolecule synthesis (
Dolganyuk et al. 2020;
Khan et al. 2018). Among the vast diversity of microalgae,
Scenedesmus incrassatulus and
Chlorella vulgaris have emerged as two notable species due to their resilience and potential for industrial applications (
Ali et al. 2022;
Fihri et al. 2024).
Chlorella vulgaris, a unicellular green microalga, is renowned for its high lipid content and fast growth, which make it a preferred choice for biofuel production and as a protein supplement in food and feed industries (
Mendes et al. 2024;
Rautenberger et al. 2024). On the other hand,
Scenedesmus incrassatulus, a colonial green microalga, has demonstrated exceptional nutrient uptake capabilities, especially in wastewater treatment, and is known for its ability to mitigate heavy metal toxicity (
Alayi et al. 2021;
Peña-Castro et al. 2004). While the metabolic potential of microalgae species like
Scenedesmus incrassatulus and
Chlorella vulgaris is well-documented, their biochemical responses to specific growth phases and nutrient stress conditions, such as nitrogen (N) and phosphorus (P) deficiencies, remain underexplored. Understanding these responses is critical for optimizing biomass production and metabolite yields in various industrial applications. Recent studies have highlighted the impact of nutrient limitations on microalgal growth and lipid accumulation. For instance,
Ratomski et al. (2021) investigated the effect of limited nutrient access on the growth and lipid content of
Chlorella vulgaris, finding that nutrient stress conditions can significantly influence biomass production and lipid accumulation. Similarly,
Udayan et al. (2023) discussed how nutrient stress, particularly nitrogen limitation, can induce the accumulation of triglycerides (triacylglycerols, TAGs), which serve as direct precursors for biodiesel production through transesterification into fatty acid methyl esters (FAMEs). Due to their high energy density and efficient conversion into liquid biofuels, TAGs represent a key target metabolite in microalgal bioenergy research, thereby enhancing the industrial relevance of nutrient-induced lipid accumulation (
Udayan et al. 2023). Nutrient stress, particularly nitrogen and phosphorus limitation, is known to significantly influence the metabolic pathways of microalgae, driving the accumulation of energy storage compounds such as carbohydrates and lipids while reducing protein synthesis (
Yaakob et al. 2021). Studies on
Chlorella sorokiniana and
Scenedesmus obliquus have shown that nutrient deficiencies enhance lipid production by redirecting carbon fluxes toward triacylglycerol synthesis, often at the expense of protein biosynthesis (
Alcántara et al. 2015;
Trivedi et al. 2022;
Zhu et al. 2022). Similarly, carbohydrate accumulation under nutrient stress has been reported as a survival strategy in various microalgae, including
Arthrospira platensis and
Chlorella salina, where energy storage molecules serve as reserves during unfavorable conditions (
Chong et al. 2019;
Lai et al. 2019). However, there is limited comparative analysis on how
S. incrassatulus and
C. vulgaris metabolically adapt to these stresses, especially in the context of their growth phases. The growth phase of microalgae plays a pivotal role in determining their metabolic activity and biochemical composition. During the exponential growth phase, microalgae prioritize protein synthesis and photosynthetic efficiency to support rapid cell division, whereas the stationary phase is often associated with nutrient depletion and the accumulation of secondary metabolites like lipids and carbohydrates (Da Costa et al. 2017;
Schüler et al. 2020). While growth phase-dependent metabolic shifts have been documented in species such as
Tisochrysis lutea and
Nannochloropsis gaditana, the extent to which these shifts occur in
S. incrassatulus and
C. vulgaris under nutrient-limiting conditions remains unclear (
Fernandes et al. 2016;
Rafay et al. 2020). Addressing this knowledge gap is essential for designing cultivation strategies that maximize the yield of target metabolites. In addition to metabolic characterization, molecular identification is essential for confirming species classification. DNA barcoding, particularly using the internal transcribed spacer (ITS) region, has proven to be a reliable method for identifying microalgal strains. For instance, a study by Zou et al. (2016) demonstrated the effectiveness of ITS sequences in distinguishing between closely related microalgae species, revealing cryptic diversity within the genus
Scenedesmus (Zou et al. 2016). Similarly, a review highlighted the utility of ITS regions in metabarcoding studies of freshwater microalgae, emphasizing their variability and applicability in species-level identification (
Kezlya et al. 2023). Elemental composition analysis provides insights into the macronutrient and trace element profiles of microalgae, which are vital for assessing their nutritional value and industrial applicability. A study analyzed the elemental composition of various microalgae species, providing data on their macronutrient and trace element profiles (
Tibbetts et al. 2015). Techniques such as Inductively Coupled Plasma-Atomic Emission Spectroscopy (ICP-AES) enable precise quantification of essential elements, while Scanning Electron Microscopy coupled with Energy Dispersive X-ray Spectroscopy (SEM-EDS) aids in understanding surface morphology and elemental distribution. Metabolic adaptations of
Scenedesmus incrassatulus and
Chlorella vulgaris under nitrogen and phosphorus limitation were characterized across exponential and stationary growth phases. Integrated biochemical profiling, ITS-based molecular identification, and elemental analysis using Inductively Coupled Plasma-Atomic Emission Spectroscopy (ICP-AES) and Scanning Electron Microscopy coupled with Energy Dispersive X-ray Spectroscopy (SEM-EDS) were performed to evaluate nutrient-driven metabolic shifts and species-specific physiological traits relevant to biotechnological applications. Specifically, we examine how nitrogen and phosphorus limitations affect the accumulation of key biomolecules, carbohydrates, proteins, and lipids during both exponential and stationary phases. These techniques provide insights into their macronutrient and trace element profiles as well as surface morphology, which are crucial for understanding their nutritional value and industrial applicability.
Materials and Methods
Harvest and Isolation
The microalgal species
Scenedesmus incrassatulus and
Chlorella vulgaris were collected from the Sidi Chahed impoundment, located on the Oued Mikkès in Morocco (34°04'30.5"N, 5°20'19.2"W). Sampling was conducted during the spring-summer season using a Nansen-type net with a 40 µm mesh size at multiple points in the reservoir. Isolation of the species was performed using the solid media technique. Sterile Dauta medium
Dauta (1982) was prepared in Petri dishes, and the collected samples were inoculated using a Pasteur pipette. The plates were incubated under suboptimal temperature and light conditions for 10 days. To obtain a monospecific culture (
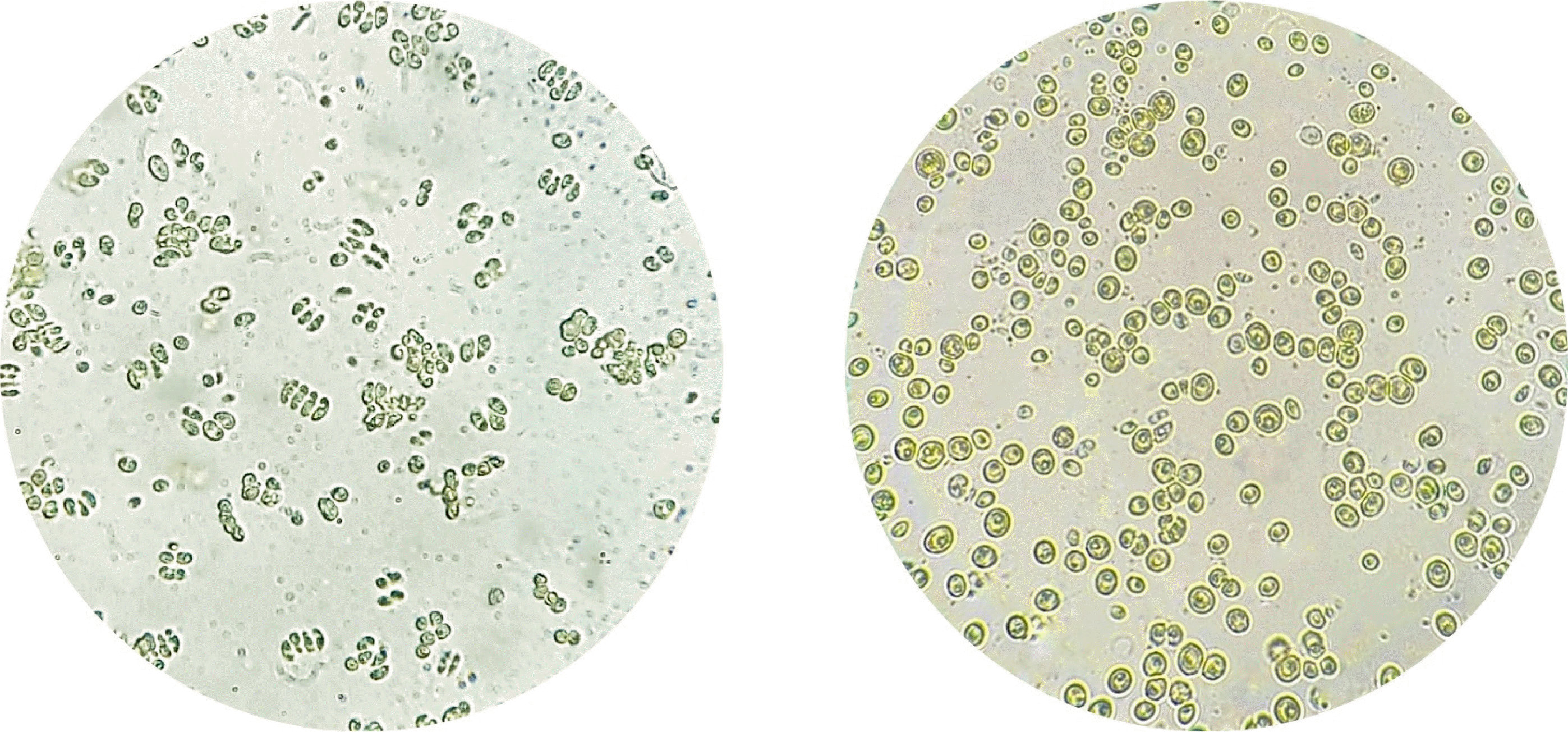
Fig. 1), colonies of microalgae were transferred every three days to fresh medium using a Pasteur pipette until purification was achieved.
The experimental cultures were maintained in the exponential growth stage through regular subculturing every three days in Dauta medium (
Table 1;
Dauta 1982). The cultures were grown in a controlled growth chamber under suboptimal conditions, with a temperature of 25℃ ± 1℃, a light intensity of 37 µmol⋅m
-2⋅s
-1 provided by Philips TL-D 18 W Snow White fluorescent tubes (12,000 K), and a photoperiod of 16:8 hours (Light:Dark). Continuous homogenization was ensured by aerating the cultures with sterile compressed air supplemented with 1% CO
2. The purity of the microalgal colonies was monitored daily using optical microscopy to ensure contamination-free growth.
The morphological characteristics of the isolated microalgae were examined and identified based on the descriptions provided by (
Bourrelly 1988). These morphological observations were further validated through molecular identification methods to ensure accurate taxonomic classification. To confirm the identity of the studied strains,
Scenedesmus incrassatulus and
Chlorella vulgaris, DNA was extracted, PCR amplified, and analyzed using standard molecular protocols. Genomic DNA was extracted using the CTAB-based Doyle and Doyle method (
Doyle et al. 1987), widely applied for plant genomic DNA isolation due to its efficiency in removing polysaccharides and polyphenolic compounds. This protocol has also been successfully adapted for green microalgae, including
Chlorella vulgaris (
Gomaa et al. 2019) and
Scenedesmus spp. (
Sarwa et al. 2017), demonstrating its suitability for obtaining high-quality DNA for downstream ITS amplification and sequencing (
Kunyalung et al. 2021). The internal transcribed spacer (ITS) regions were amplified using ITS1 (TCCGTAGGTGAACCTGCGG) and ITS4 (TCCTCCGCTTATTGATATGC) primers with DreamTaq Green PCR Master Mix (2X) (Thermo Scientific). The PCR reactions followed standard cycling conditions: an initial denaturation at 94℃ for 2 minutes, followed by 35 cycles of denaturation at 94℃ for 30 seconds, annealing at 55℃ for 30 seconds, and extension at 72℃ for 1 minute, with a final extension at 72℃ for 5 minutes. The resulting sequences were compared against the NCBI GenBank database using the BLAST tool to identify homologous sequences. The phylogenetic relationships were inferred using the Maximum Likelihood (ML) method based on the Tamura-Nei model of nucleotide substitution (
Tamura et al. 1993). The tree with the highest log-likelihood value (-986.22) is presented. Branch support values were estimated using bootstrap analysis with 1,000 replicates, and the percentage of replicate trees in which the associated taxa clustered together is indicated next to the branches (
Felsenstein 1985). The initial tree for the heuristic search was obtained by selecting the topology with the highest log-likelihood among Neighbor-Joining (NJ) and Maximum Parsimony (MP) trees (
Saitou et al. 1987). The NJ tree was generated from a matrix of pairwise distances calculated under the Tamura-Nei model, while the MP tree corresponded to the shortest tree obtained from ten random addition sequence searches (
Tamura et al. 1993). The analysis included eight nucleotide sequences. Positions with less than 95% site coverage were eliminated using the partial deletion option, resulting in a final dataset of 383 aligned positions. All evolutionary analyses were conducted using MEGA version 12 with up to four parallel computing threads (
Kumar et al. 2024).
Sample Preparation for ICP-AES
For elemental composition analysis, 3 g of lyophilized microalgal powder was accurately weighed using an analytical balance. The weighed samples were subjected to a high-temperature calcination process in a Nabertherm LHT 02/17 furnace. The temperature ramp was set to increase gradually to 550℃ over 1 hour, followed by a holding period of 4 hours at 550℃ to ensure complete ashing of organic matter. The samples were then allowed to cool to room temperature inside the furnace. The resulting ash (0.1 g) was dissolved in 100 mL of hydrochloric acid (HCl, 37%) and left for 24 hours. The digested samples were filtered through glass fiber membranes (0.2 µm) to remove undissolved particles. Elemental concentrations are expressed as mg g-1 dry weight (DW) of lyophilized biomass.
Dilution and Calibration
The filtered digests were diluted to a final volume of 100 mL (dilution factor of 10) with ultrapure water before analysis. The diluted sample is then ready for ICP-AES analysis. The instrument used for the analysis is an ICP-AES (Varian-Vista) equipped with a Cetac ultrasonic nebuliser. Calibration was performed using multi-element standards in the range of 0.1 to 10 mg L-1. Elemental analysis was performed in triplicate to ensure accuracy and reproducibility and reported as mean ± standard deviation.
The important analysis parameters of the device are as follows:
- Rf power: 0.7 - 1.5 kilowatts (1.2-1.3 kilowatts for axial mode)
- Plasma gas flow rate (Ar): 10.5 - 15 L min-1 (radial mode), 15 L min-1 (axial mode)
- Auxiliary gas flow (Ar): 15 L min-1 (axial mode)
- Viewing size: 5-12 mm
- Copy and read times: 1-5 seconds (maximum 60 seconds)
- Copy time: 3 seconds (maximum 100 seconds)
Morphological Analysis by Scanning Electron Microscopy (SEM)
Sample Preparation for SEM and SEM Imaging
For surface morphology analysis, a small quantity of lyophilized microalgal powder was directly mounted onto conductive black carbon adhesive tapes and fixed to aluminum stubs and then coated with a thin layer of gold. Given the SEM's ability to operate in Variable Pressure (VP) mode, metallization was omitted to prevent artifacts. Microstructural observations were performed using a Philips XL30i Scanning Electron Microscope (SEM) operating in Variable Pressure (VP) mode. The images were taken with an excitation voltage of 16.0 kV. In addition, diffractograms were obtained using an X-ray diffraction apparatus (X'Pert Pro, Kolkata, India), with a voltage of 40 kV and a current of 30 mA as the generator's operating parameters.
Experimental Controls and Precautions
All sample processing and analytical procedures were conducted under sterile conditions to minimize contamination risks. Blank samples (without microalgae) were analyzed alongside experimental samples to account for any background interference from reagents or equipment. To ensure data reliability, all experiments were performed in triplicate, with independent repetitions verifying the robustness of the results.
Monitoring of Growth Kinetics
Growth kinetics were studied using the “batch culture” method, in which all essential nutrients for microalgal growth were added at the start of the experiment (time zero). After inoculation, no additional nutrients or biomass were introduced into the culture. Growth was monitored daily by measuring variations in algal biomass through cell counting and optical density (OD) measurements at 750 nm, alongside Chlorophyll a analysis was conducted following the method described by (
Rai 1980), with considerations for solvent effects and methodology refinements outlined by (
Marker 1994) to ensure accurate quantification. A positive correlation between the optical density of the culture medium and the microalgal biomass has been established (
Birkholz et al. 2022).
The Effect of Growth Phase
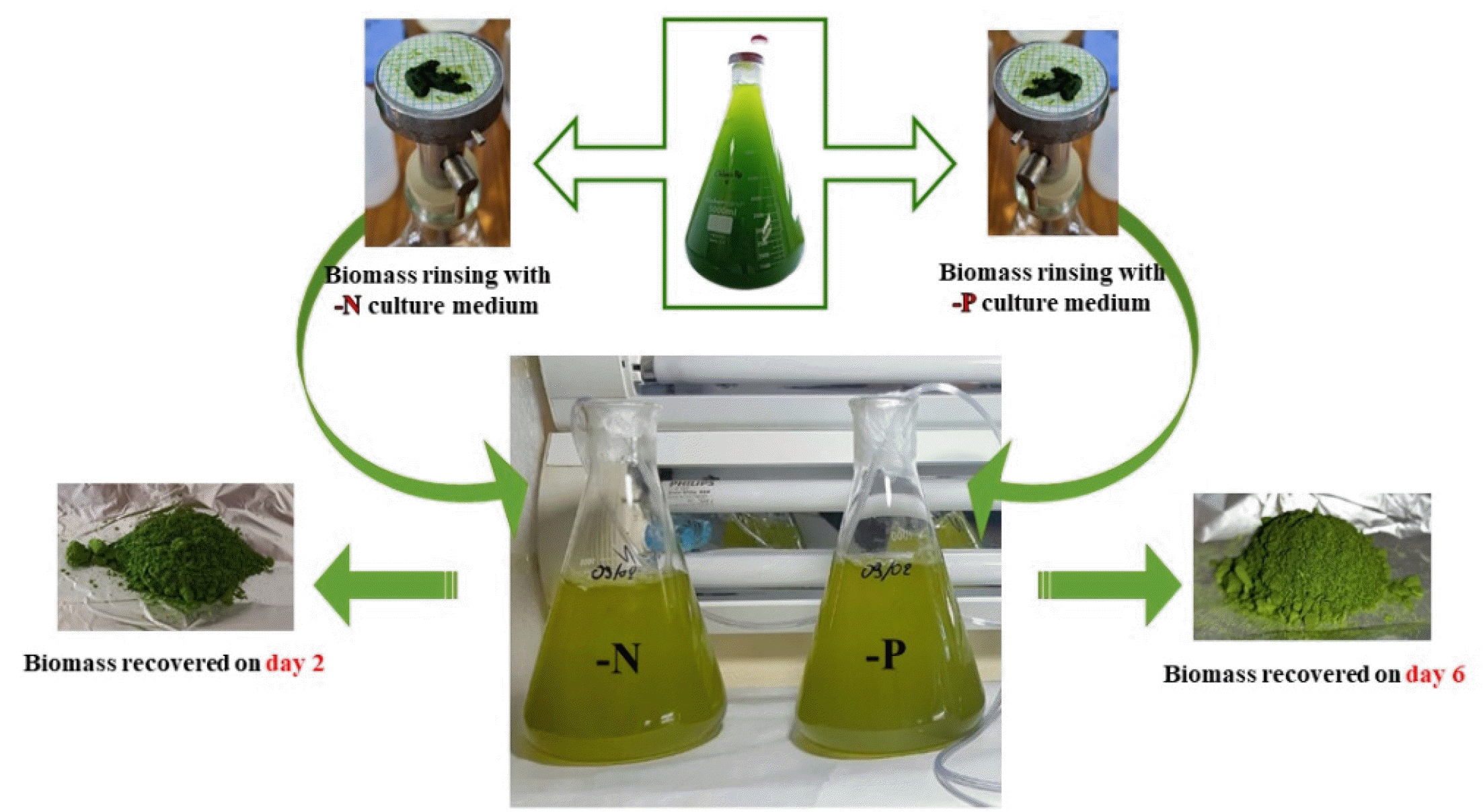
Algal biomass was collected at two distinct growth phases: the exponential phase (day 3) and the stationary phase (day 11). Biomass recovery was performed by gentle filtration using a 0.45 µm Millipore filter. The filtered biomass was freeze-dried at -80℃ and subsequently stored at 4℃ until further analysis.
The Effect of Nutrient Deficiency
The effect of nutrient deficiency on the phytochemical composition of microalgae was studied using an experimental deficiency protocol adapted from several authors (
Belkoura et al. 1994;
Bouchamma et al. 2004;
Dauta 1982). The nutrients investigated were nitrogen (N), supplied as KNO
3, and phosphorus (P), supplied as K
2HPO
4. Cultures were maintained in the exponential growth phase under suboptimal conditions and synchronized by keeping them in the dark for 24 hours. Cells were then harvested by filtration using Whatman GF/C filter paper and rinsed thoroughly with a nutrient-free medium specific to the deficiency being studied (N or P). The cells were subsequently reseeded into the deficient medium and cultured under suboptimal temperature and light conditions. Growth was monitored daily by measuring optical density (OD) at 750 nm and cell counts using a Malassez counting chamber. Cultures were considered deficient when growth ceased (OD remained constant), which occurred after two days for nitrogen deficiency and six days for phosphorus deficiency (
Dauta 1982). Biomass was then harvested, freeze-dried at -80℃, and stored at 4℃ until further analysis (
Fig. 2).
Carbohydrate Assay
The determination of soluble carbohydrates was conducted using the method described by
Dubois et al. (1951). Algal biomass was placed in a test tube containing 5% phenol and left to stand at room temperature for 20 minutes. After vortex mixing, the mixture was transferred to ice for 3 minutes. Subsequently, 5 mL of concentrated sulfuric acid was added, and the mixture was incubated at 100℃ for 20 minutes, followed by rapid cooling on ice. The absorbance was measured at 490 nm after centrifugation at 5000 rpm for 5 minutes. Soluble carbohydrate content was quantified using a glucose standard curve as a reference.
Protein Assay
The protein content was determined using the Lowry assay (
Lowry et al. 1951). An alkaline solution (20 g⋅L
-1 sodium carbonate in 0.1 M sodium hydroxide) was mixed with the microalgal sample, followed by the addition of copper sulfate and potassium sodium tartrate. The mixture was incubated at room temperature for 20 minutes. Subsequently, Folin's reagent (diluted 1:1) was added with rapid stirring. The sample was centrifuged for 5 minutes, and the optical density was measured at 750 nm. Soluble protein content was quantified using a standard curve of Bovine Serum Albumin (BSA) at a concentration of 0.1 g⋅L
-1 as a reference.
Lipid Assay
Lipid determination was performed following the Folch method (
Bligh et al. 1959;
Folch et al. 1957). Lyophilized biomass was ground in 1.5 mL of chloroform and 1.2 mL of distilled water to create a single-phase system, and the piston was rinsed with 3 mL of methanol. The mixture was homogenized by shaking and centrifuged at 5000 rpm for 5 minutes. To form a biphasic system, 1.5 mL of chloroform and 1.2 mL of distilled water were added to the supernatant, resulting in two distinct phases. The upper alcoholic phase was carefully pipetted off, and the lower chloroform phase was transferred to a ground flask. The chloroform extract was dried using a rotary evaporator, and 2 mL of bicarbonate solution (2.5 g⋅L
-1) was added to the residue. The mixture was heated in a water bath at 100℃ for 15 minutes, followed by the addition of 5 mL of distilled water. Lipid content was measured by recording the optical density at 350 nm. Soluble lipid content was quantified using a standard curve based on palmitic acid at a concentration of 0.1 g⋅L
-1 as a reference.
Statistical Analysis
Statistical analysis was performed with R version 4.1.2 for Windows (
R Core Team 2021). Growth curve parameters were obtained using the package “growthcurver” (
Sprouffske et al. 2016) using the logistic equation (Equation 1) with “K”, the carrying capacity, “N0”, the initial population size, “r”, the growth rate, “t” Nt”, the population size at time “t”.
To compare protein, lipid and carbohydrate content depending on the different growth media, least-square linear models were used with ‘medium’ as independent variable. To compare the parameters depending on the growth phase (exponential or stationary), a pairwise T-test was used. Linear model assumptions (normality and homoscedasticity) were verified by a visual check of the residual’s plots. Models were analysed with analysis of variance (ANOVA), Tukey’s range test (Honest Significant Difference, HSD) was used to obtain adjusted P-values to account for multiple comparisons, with level of significance α = 0.05. Plots were made using “ggplot2” (
Wickham 2016,
2009), analysis was performed using the packages “lsmeans” (
Lenth 2016) and “multcomp” (
Hothorn et al. 2008)
Results
Molecular Identification of Chlorella vulgaris and Scenedesmus incrassatulus
Morphological identification of the isolates was first performed using Bourrelly’s taxonomic key for freshwater green algae (
Bourrelly 1988). The observed phenotypic characteristics, including cell shape, size, colony structure, and chloroplast morphology, were consistent with those described for
Chlorella vulgaris and Scenedesmus incrassatulus. Subsequently, molecular identification was performed by amplifying and sequencing the ITS region, followed by BLAST analysis against the GenBank database. The trimmed query sequence (686 bp) showed significant alignment with multiple
C. vulgaris isolates. The highest similarity was observed with
C. vulgaris isolate DRL3 (JX139000.1), displaying 89% identity over the aligned region, with a score of 239 bits and an E-value of 3e-63. In addition to
C. vulgaris DRL3, the query sequence also aligned closely with other
C. vulgaris isolates, including
C. vulgaris liming lake 1,
C. vulgaris CBS 15-2075, and
C. vulgaris strain CCAP 211/11F, all showing high identity, query coverage and significant E-values. Although the percentage identity was moderate, phylogenetic analysis based on ITS sequences clustered the isolate within the
C. vulgaris lineage with > 90% bootstrap support (
Fig. 3). These combined results suggest a close phylogenetic affiliation of the isolate with
C. vulgaris. The moderate percentage identity may be attributed to sequence variability and trimming of low-quality terminal regions.
Similarly, the ITS sequence of the
Scenedesmus isolate exhibited significant alignment with
S. incrassatulus genomic DNA from culture collection CCAP 276/43 (FR865722.1), showing 84% identity across the aligned region with an E-value of 0.0. Phylogenetic reconstruction positioned the isolate within the
S. incrassatulus clade, clustering closely with
S. incrassatulus CCAP 276/43 (FR865722.1). Despite the moderate sequence identity, phylogenetic reconstruction supported its taxonomic placement within this lineage with strong bootstrap support (
Fig. 4).
Macronutrient Analysis
The concentrations of major macronutrients, including calcium (Ca), potassium (K), magnesium (Mg), and phosphorus (P), showed notable differences between
Scenedesmus incrassatulus (SI) and
Chlorella vulgaris (CV). SI exhibited a higher calcium concentration (20.31 mg g
-1) compared to CV (16.824 mg g
-1). Similarly, magnesium levels were higher in SI (5.1887 mg g
-1) compared to CV (4.848 mg g
-1). However, potassium levels were substantially greater in CV (4.510 mg g
-1) relative to SI (1.468 mg g
-1). Phosphorus content was also higher in CV (2.855 mg g
-1) than in SI (2.055 mg g
-1), highlighting species-specific differences in nutrient uptake and storage (
Table 2).
Trace Element Distribution
Both microalgae species demonstrated distinct profiles for trace metals. Iron (Fe) concentrations were higher in CV (0.132 mg g
-1) compared to SI (0.103 mg g
-1). Additionally, zinc (Zn) levels were slightly greater in CV (0.0817 mg g
-1) than in SI (0.0777 mg g
-1). Other elements, such as boron (Bo) and barium (Ba), were present in trace amounts, with similar concentrations across both species. Silicon (Si), a structural component for cell walls, was more abundant in SI (0.382 mg g
-1) compared to CV (0.365 mg g
-1) (
Table 2).
Surface Morphology and Elemental Analysis (SEM-EDS)
The surface morphology of
Scenedesmus incrassatulus (SI) and
Chlorella vulgaris (CV) was analyzed using Scanning Electron Microscopy (SEM), revealing distinct structural characteristics. SEM images demonstrated that
S. incrassatulus exhibited a compact and granular surface morphology with small, densely packed cells, reflecting its cenobial organization. This granular appearance may indicate its adaptation to efficient nutrient absorption and storage. In contrast,
Chlorella vulgaris displayed a relatively smooth and interconnected surface morphology with a unicellular structure, consistent with its adaptation to a collective growth form. Energy-Dispersive X-ray Spectroscopy (EDS) provided complementary, semi-quantitative information on the surface elemental composition of both microalgae. Surface EDS spectra indicated that
S. incrassatulus was primarily composed of carbon (58.49%) and oxygen (37.47%), with minor elements including potassium (1.78%) and phosphorus (1.22%). Similarly,
C. vulgaris exhibited dominant carbon (57.75%) and oxygen (35.64%) signals, with a relatively higher potassium content (4.03%) compared to
S. incrassatulus (1.78%). Phosphorus signals in
C. vulgaris were also slightly higher (1.50%) relative to
S. incrassatulus (1.22%). These observations reflect relative surface elemental distribution and are consistent with the bulk elemental trends obtained by ICP-AES (
Table 3).
These findings suggest that while both species share similar elemental compositions, the higher potassium and phosphorus content in
C. vulgaris may reflect species-specific physiological adaptations, such as enhanced nutrient uptake and storage capabilities. The interconnected morphology of
S. incrassatulus, on the other hand, may contribute to its resilience and efficiency in environmental stress conditions or nutrient acquisition in colonial growth forms (
Fig. 5).
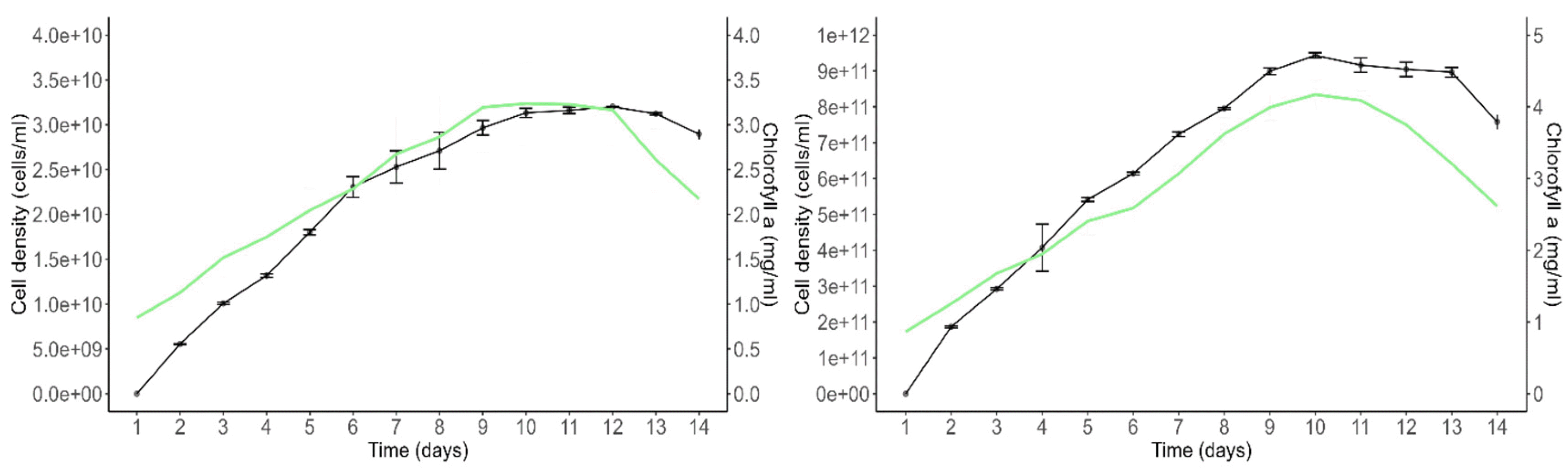
Growth and Chlorophyll a Content
The growth curves of
Scenedesmus incrassatulus and
Chlorella vulgaris under non-limiting nutrient and suboptimal heliothermic conditions are presented (
Fig. 6). The growth of
S. incrassatulus, measured by cell density (cells per milliliter), exhibited an exponential growth phase during which both cell density and chlorophyll a content increased concurrently. On day 9, the stationary phase was reached, marked by growth arrest and a subsequent decline in chlorophyll a content. Beyond day 14, the culture entered a decline phase characterized by further deterioration of chlorophyll a level, leading to the chlorosis of the cultures. Similarly, the growth of
C. vulgaris followed a pattern with an exponential growth phase during which cell density and chlorophyll a content increased simultaneously. On day 9, the stationary phase was observed, and by day 13, the culture entered a decline phase due to a reduction in chlorophyll a level.
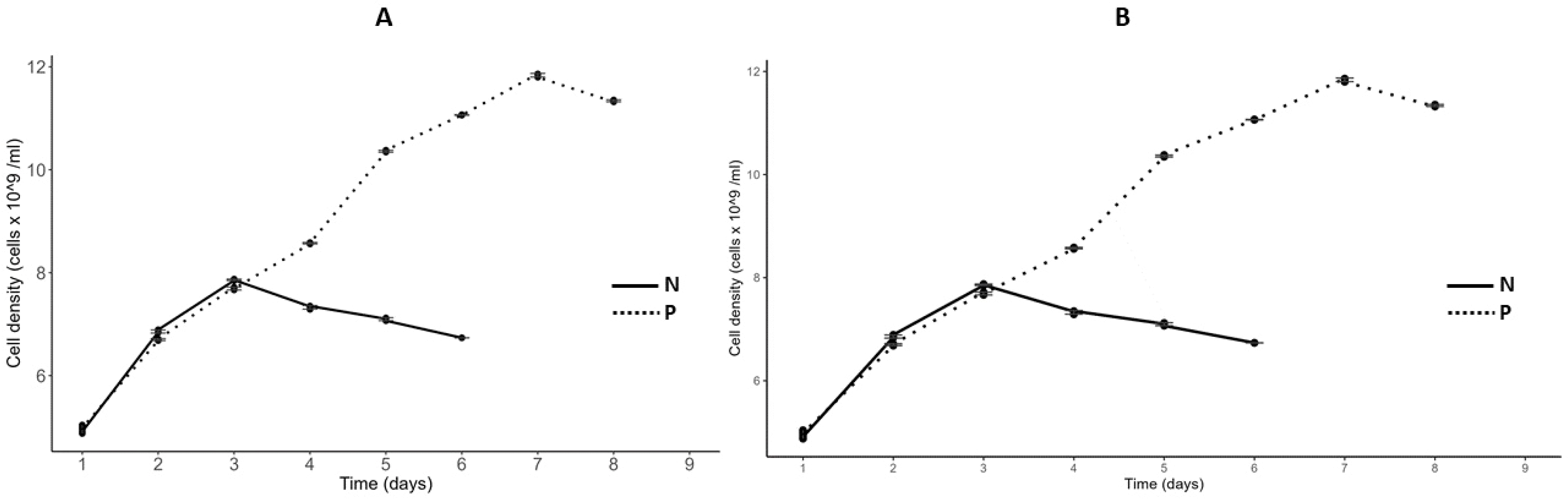
The growth curves of
C. vulgaris and
S. incrassatulus in nitrogen- (N) and phosphorus- (P) depleted media are shown (
Fig. 7). Daily measurements of optical density (OD) at 750 nm and cell density using a Malassez counting chamber were conducted until growth ceased (OD = constant). Under nitrogen-deficient conditions, growth halted after two days, while both cultures continued to grow until the sixth day under phosphorus-deficient conditions. At this point, the cultures were confirmed to be deficient in the respective nutrient (N or P). The growth parameters of these cultures, including carrying capacity, initial population size, and growth rate, are summarized in
Table 4.
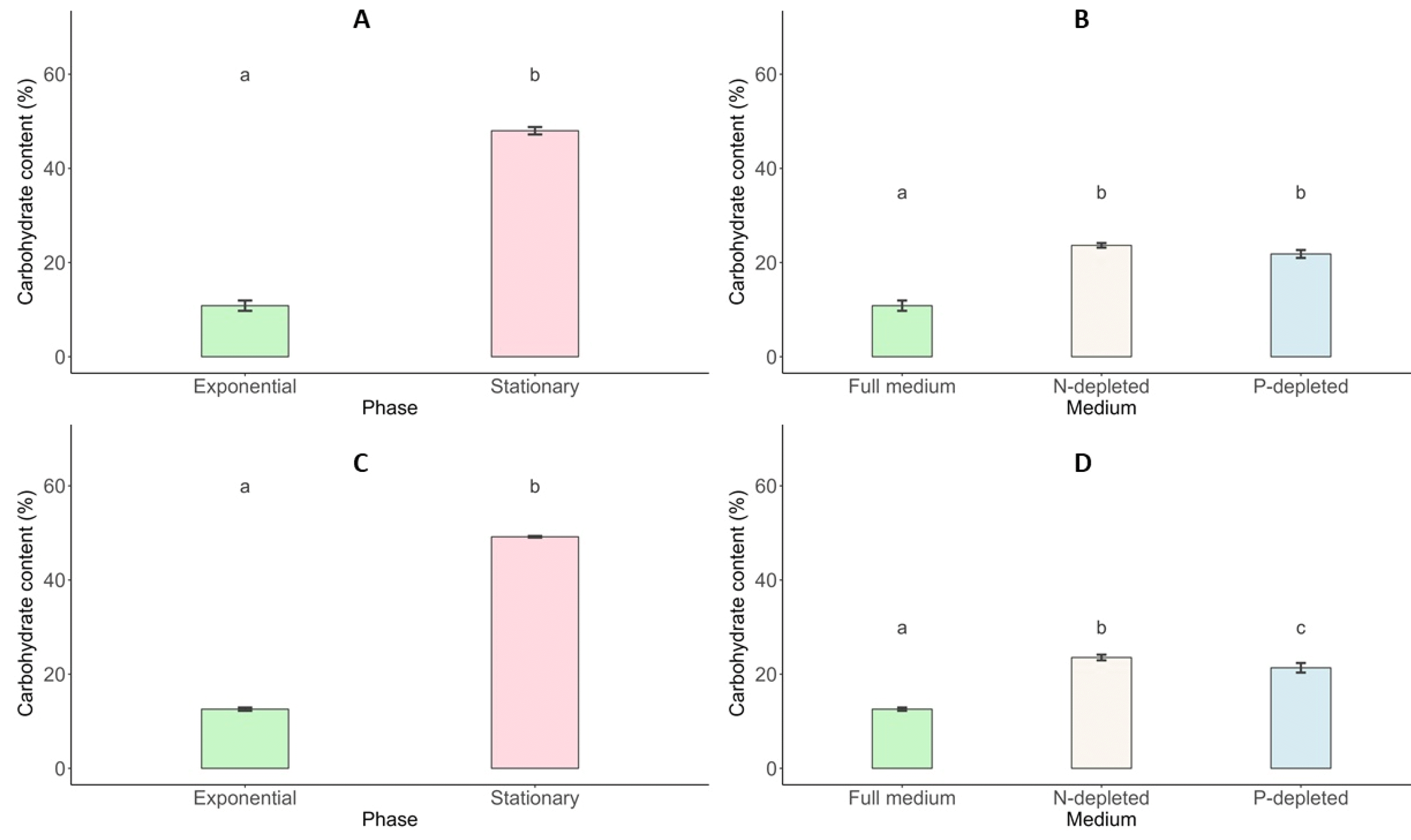
Total Carbohydrate Content
The changes in carbohydrate content for
Scenedesmus incrassatulus and
Chlorella vulgaris under different conditions are presented (
Fig. 8). The data highlight the effects of growth phases and nutrient deficiencies, particularly nitrogen (N-NO
3) and phosphorus (P-PO
4) limitations. During the stationary growth phase,
S. incrassatulus achieved its highest total carbohydrate content of 47.99% on day 11, while
C. vulgaris reached a maximum of 49.18%. In contrast, during the exponential phase, the carbohydrate content was significantly lower, with
S. incrassatulus at 10.84% and
C. vulgaris at 12.57%. Both microalgae species demonstrated an enhanced capacity for carbohydrate production when subjected to nutrient deficiencies compared to conditions with sufficient nutrients. Under nitrogen and phosphorus deficiency,
S. incrassatulus produced total carbohydrate contents of 23.65% and 21.82%, respectively. Similarly,
C. vulgaris exhibited total carbohydrate contents of 23.54% and 21.35% under nitrogen- and phosphorus-limited conditions, respectively.
These results suggest that nutrient deficiencies, especially of nitrogen and phosphorus, stimulate carbohydrate synthesis in both S. incrassatulus and C. vulgaris, highlighting their ability to adapt to nutrient stress by modulating their metabolic responses. Overall, this analysis underscores the variations in carbohydrate content based on growth phases and nutrient conditions, providing valuable insights into the metabolic adaptations of these microalgae and their potential applications in biotechnological processes.
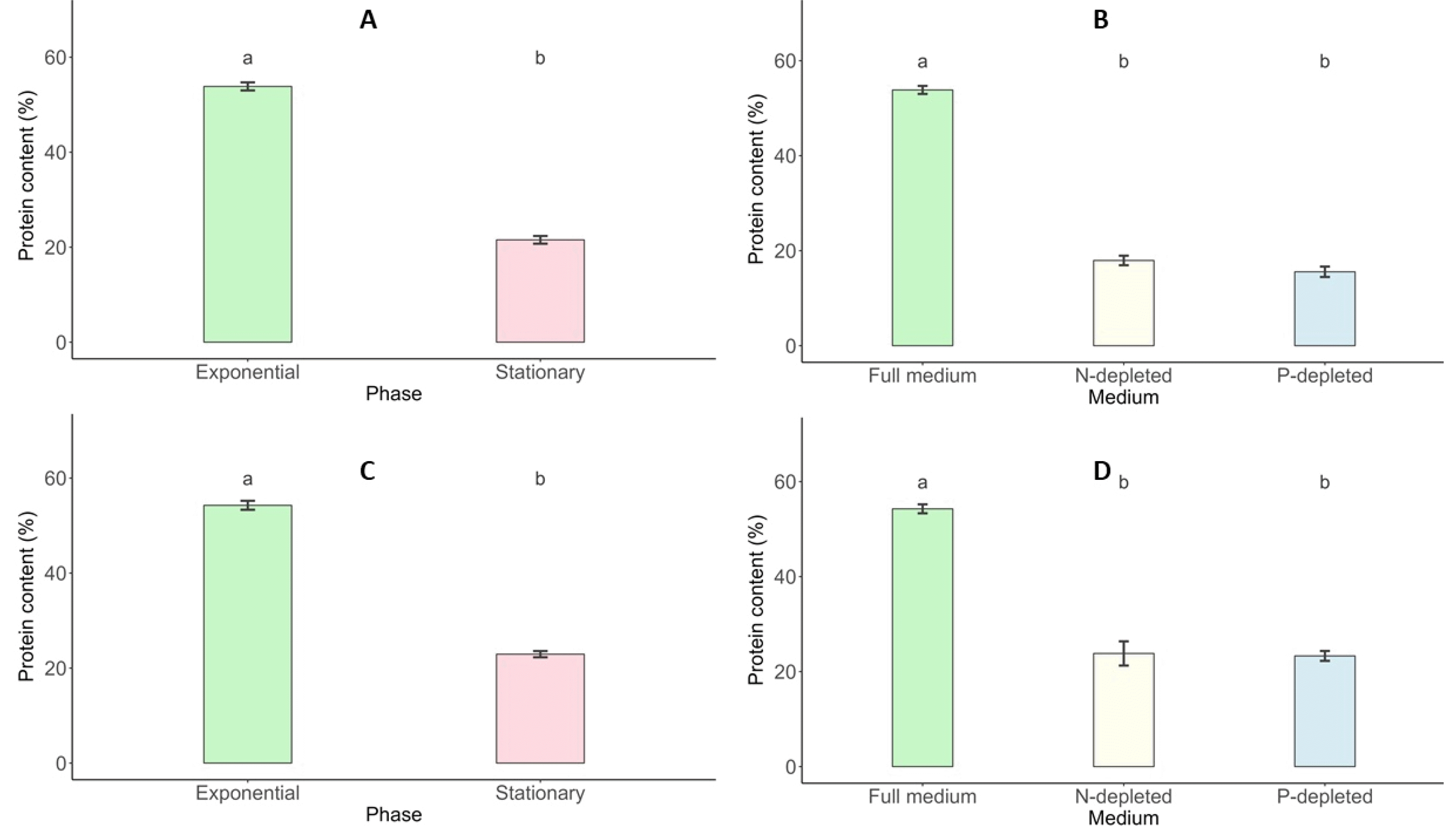
Total Protein Content
The intracellular protein content of
Scenedesmus incrassatulus and
Chlorella vulgaris varied significantly across growth stages and nutrient conditions. During the exponential growth phase,
S. incrassatulus exhibited a peak protein content of 53.85%, which declined sharply to 21.54% in the stationary phase. Similarly,
C. vulgaris reached its maximum protein content of 54.28% during the exponential phase, decreasing to 22.93% during the stationary phase (
Fig. 9). Nutrient deficiencies further impacted protein synthesis in both species. Under nitrogen (N) and phosphorus (P) deficiency, the protein content of
S. incrassatulus decreased from 53.85% in optimal conditions to 17.94% and 15.56%, respectively.
C. vulgaris displayed a similar trend, with protein content dropping from 54.28% in non-limiting conditions to 23.82% and 23.30% under N- and P-deficient conditions, respectively (
Fig. 9).
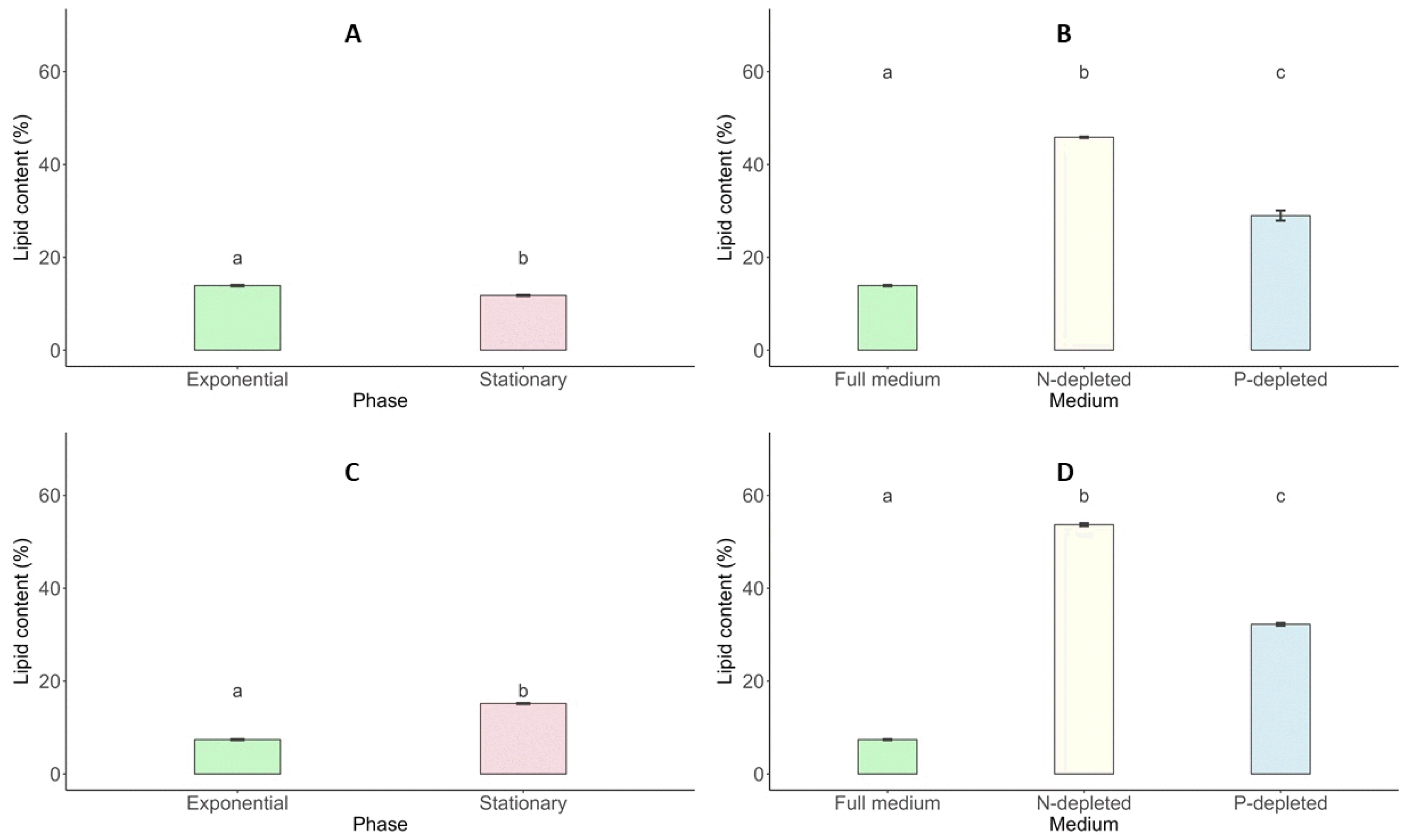
Total Lipid Content
Lipid synthesis in
Scenedesmus incrassatulus and
Chlorella vulgaris showed distinct patterns across growth stages and nutrient conditions. During the exponential and stationary phases,
S. incrassatulus exhibited lipid contents of 13.92% and 11.79%, respectively, while
C. vulgaris had 7.39% and 15.16% during the same phases, indicating species-specific lipid dynamics (
Fig. 10). Nutrient stress, particularly nitrogen deficiency, significantly enhanced lipid production in both microalgae. Under nitrogen-deficient conditions, the lipid content of
S. incrassatulus increased from 13.92% in optimal conditions to 45.86%, while phosphorus deficiency resulted in a lipid content of 28.99%.
C. vulgaris displayed even greater lipid accumulation under nitrogen and phosphorus limitations, reaching 53.68% and 32.23%, respectively, compared to 13.92% under non-limiting conditions (
Fig. 10).
The variations in protein, carbohydrate, and lipid content under different growth stages and nutrient conditions are summarized in
Table 5. These results provide a comprehensive understanding of the biochemical shifts occurring in
S. incrassatulus and
C. vulgaris, offering valuable insights into their metabolic responses to nutrient stress and growth phase-dependent changes (
Table 5).
Discussion
This study presents a comprehensive investigation of two green microalgal species, Scenedesmus incrassatulus and Chlorella vulgaris, isolated from the waters of the Sidi Chahed impoundment in Morocco. Molecular and elemental analyses were conducted to confirm their taxonomic identities and assess their macro- and micronutrient compositions. In addition, their growth and biochemical responses under varying nitrogen and phosphorus levels and different growth phases were evaluated, providing a holistic view of the metabolic adaptations of these native strains. The findings reveal the adaptive potential of both strains and support their relevance for various biotechnological applications, including biofuel production and environmental sustainability.
Molecular Identification and Phylogenetic Analysis
The use of the internal transcribed spacer (ITS) region for molecular identification and taxonomic classification of microalgae is well-established in scientific research. The ITS region, located between the small and large subunit ribosomal RNA genes, is highly variable among species, making it a valuable marker for distinguishing closely related taxa. For instance, a study focusing on the molecular characterization and strain identification of
Tetraselmis and
Chlorella species utilized the ITS barcode approach. By analyzing the full nuclear ITS region through Sanger sequencing, researchers obtained ITS barcodes that were subsequently compared with other ITS sequences of various
Tetraselmis and
Chlorella species, facilitating accurate taxonomic classification (
Fathy et al. 2023). Additionally, a comprehensive DNA barcoding study on 118
Scenedesmus strains employed multiple genetic markers, including the ITS region, to assess species boundaries and relationships. This research demonstrated the effectiveness of the ITS region in resolving taxonomic identities within the
Scenedesmus genus (Zou et al. 2016). These studies, among others, support the application of ITS region sequencing and phylogenetic analysis as reliable methods for the molecular identification of microalgae, aligning with the findings of current research on
Chlorella vulgaris and
Scenedesmus incrassatulus. The phylogenetic trees constructed in this study placed both species within their respective clades, further validating their molecular identities. The molecular confirmation of these species is critical for ensuring the reliability of subsequent biochemical and physiology ical analyses. Misidentification of microalgae strains is a common issue in algal biotechnology, leading to inconsistent results across studies. For example,
C. vulgaris is often confused with other
Chlorella species due to their morphological similarities. By using molecular techniques, this study eliminates such ambiguities, providing a robust foundation for comparative analyses.
Elemental analysis revealed significant differences in macronutrient and trace element profiles between
C. vulgaris and
S. incrassatulus.
C. vulgaris exhibited higher potassium (4.510 mg g
-1) and phosphorus (2.855 mg g
-1) content compared to
S. incrassatulus (1.468 mg g
-1 and 2.055 mg g
-1, respectively). These findings suggest that
C. vulgaris has a greater capacity for nutrient uptake and storage, which may contribute to its higher lipid and carbohydrate accumulation under nutrient stress. In contrast,
S. incrassatulus showed higher calcium (20.31 mg g
-1) and magnesium (5.188 mg g
-1) content, which may reflect its structural adaptations and resilience to environmental stress. The trace element profiles also revealed species-specific differences, with
C. vulgaris exhibiting higher iron (0.132 mg g
-1) and zinc (0.081 mg g-1) content compared to
S. incrassatulus. These elements are essential for enzymatic activities and metabolic processes, and their higher concentration in
C. vulgaris may contribute to its enhanced metabolic flexibility under stress conditions. These findings are consistent with studies by
Peña-Castro et al. (2004), who reported that
S. incrassatulus has a high affinity for heavy metals, making it suitable for wastewater bioremediation (
Peña-Castro et al. 2004). Comparative studies on related microalgae species support these observations. For instance, research on
Chlorella vulgaris has demonstrated its efficiency in nutrient uptake and accumulation of valuable metabolites, which is attributed to its elemental composition and metabolic flexibility (
Mendes et al. 2024). Similarly, studies on
Scenedesmus obliquus, a species closely related to
S. incrassatulus, have shown its potential in heavy metal removal and tolerance to environmental stress, linked to its specific elemental makeup (
Goswami et al. 2022). A study evaluated the growth, contaminant removal, and biochemical component accumulation of
Chlorella vulgaris,
Scenedesmus obliquus, and
Chlamydomonas reinhardtii. The findings indicated that
C. vulgaris exhibited higher biomass productivity and nutrient removal efficiency, which may be attributed to its superior nutrient uptake and storage capabilities. In contrast,
S. obliquus demonstrated a different nutrient assimilation pattern, potentially due to variations in elemental composition (
Ajala et al. 2020). Research focusing on the influence of media composition on the growth rates of
Chlorella vulgaris and
Scenedesmus actus revealed that media components significantly affect the elemental composition and growth dynamics of these microalgae. The study suggested that optimizing nutrient availability in the growth medium could enhance the accumulation of desired macronutrients and trace elements, thereby improving biomass yield and biochemical profiles (
Crofcheck et al. 2013). An investigation into the cell wall structures of
Chlorella vulgaris and
Scenedesmus sp. highlighted differences in their cell surface compositions and morphologies. These structural variations may influence their nutrient uptake efficiency and resilience to environmental stressors, correlating with differences in their elemental profiles (
Spain et al. 2022). These studies along with the current study collectively suggest that
C. vulgaris tends to have a higher capacity for nutrient uptake and storage, which may contribute to its enhanced lipid and carbohydrate accumulation under nutrient stress conditions. In contrast,
Scenedesmus species exhibit structural adaptations and elemental compositions that may confer resilience to environmental stressors, aligning with their observed higher calcium and magnesium contents.
The structural differences highlight species-specific adaptations to their respective growth strategies and environmental conditions. EDS analysis further revealed variations in elemental composition, with
C. vulgaris showing higher potassium and phosphorus content compared to
S. incrassatulus. These elevated levels of potassium and phosphorus in
C. vulgaris may indicate enhanced nutrient uptake and storage capabilities, which contribute to its metabolic versatility and potential for industrial applications. Several studies have utilized SEM-EDS to analyze the morphological and elemental characteristics of microalgae, providing insights into their structural adaptations and potential applications. Such as, research on
Vaucheria sessilis employed SEM-EDS to observe significant morphological changes and elemental composition after bioaccumulation of metal ions. The study revealed alterations in cell wall structure and elemental distribution, highlighting the alga's capacity for metal uptake and potential use in bioremediation (
Michalak et al. 2014). An investigation into the brown macroalga
Dictyota menstrualis used SEM-EDS to assess morphological and chemical changes before and after extraction processes. The findings indicated significant structural modifications and variations in elemental composition, emphasizing the importance of such analyses in understanding the impact of extraction methods on algal biomass (
Lima et al. 2019). While direct SEM-EDS comparisons between
Chlorella vulgaris and
Scenedesmus incrassatulus are limited, studies on related species provide valuable context. For instance, SEM-EDS analysis of
C. vulgaris has demonstrated a rigid cell structure with specific elemental ratios, such as an O/C ratio of 1.03 under certain light conditions. These structural and elemental characteristics are crucial for understanding the alga's metabolic processes and optimizing conditions for bioenergy production (
Dhanasekar et al. 2023). Another study investigated the effects of nanomaterials on lipid production in
S. obliquus. SEM was used to observe morphological changes in the microalgae when exposed to different nanomaterials, providing insights into their potential for enhanced biofuel production (
He et al. 2017).
The growth patterns of
C. vulgaris and
S. incrassatulus followed a typical trajectory, characterized by an exponential growth phase, followed by a stationary phase and a decline phase. The decline phase occurs as nutrients are depleted and metabolic byproducts accumulate, underscoring the importance of nutrient management in microalgae cultivation (
Korozi et al. 2023). A strong correlation was observed between chlorophyll a content and cell density in both species, suggesting that chlorophyll a can serve as an indicator of metabolic activity and physiological state (
Salmi et al. 2021). Similar findings were reported by
Piasecka et al. (2022), who examined growth phase-dependent metabolic shifts in
Chlorella sorokiniana, highlighting the influence of growth stages on metabolic adaptations. The observed decline in chlorophyll a content during the stationary phase further supports the idea that nutrient depletion triggers a shift from primary metabolism (e.g., photosynthesis and protein synthesis) to secondary metabolism (e.g., lipid and carbohydrate accumulation). This decline in photosynthetic efficiency under nutrient stress is a well-documented phenomenon in microalgae (
Benavente-Valdés et al. 2016;
Khoo et al. 2023).
Microalgae exhibit significant metabolic changes depending on their growth phase and nutrient availability. In this study,
Scenedesmus incrassatulus and
Chlorella vulgaris were more affected by nitrogen deficiency than phosphorus deficiency. Growth halted after two days in nitrogen-depleted medium, while both species continued to grow until the sixth day under phosphorus-depleted conditions. A marked increase in carbohydrate content during the stationary phase suggests that microalgae accumulate carbohydrates as an energy reserve in response to slowed growth and nutrient stress. This aligns with previous research emphasizing carbohydrate storage as a survival strategy under nutrient limitation (de Carvalho Silvello et al. 2022;
Debnath et al. 2021;
Olguín et al. 2022). Studies by
Chen et al. (2022) on
C. vulgaris and
Scenedesmus obliquus under sludge extract culture conditions demonstrated distinct autoregulatory mechanisms, particularly in sugar metabolism, which play a crucial role in energy synthesis and distribution.
Lai et al. (2019) found that nitrogen limitation resulted in the highest carbohydrate concentrations in Arthrospira platensis, while phosphorus limitation also significantly influenced carbohydrate accumulation. Similarly,
Chong et al. (2019) reported that
Chlorella salina exhibited the highest carbohydrate accumulation under nutrient-limiting conditions. The accumulation of carbohydrates under nitrogen-limited conditions has been linked to the degradation of nitrogen-containing macromolecules such as proteins (
González-Fernández et al. 2012). Nitrogen is essential for the synthesis of nucleic acids, amino acids, proteins, and chlorophyll. When nitrogen is scarce, microalgal growth slows, photosynthetic efficiency declines, and protein synthesis decreases, leading to the redirection of carbon fixation toward carbohydrate accumulation (
Shah et al. 2022). This phenomenon was further emphasized by
Ratomski et al. (2021), who examined the effects of nutrient stress on
C. vulgaris, demonstrating its relevance for biotechnological applications. In contrast to carbohydrates, protein synthesis was more active during the exponential phase. The highest protein contents were observed in the exponential biomass of
S. incrassatulus (53.85%) and
C. vulgaris (54.28%), but significantly declined during the stationary phase. These findings are in agreement with da Costa et al. (2017), who reported that protein content was considerably higher in the exponential phase of
Tisochrysis lutea, regardless of nitrogen availability. Similarly,
Sánchez-Bayo et al. (2020) observed that protein accumulation in
C. vulgaris peaked during exponential growth but decreased significantly in the stationary phase. Nutrient availability, particularly nitrogen and phosphorus, plays a crucial role in regulating protein synthesis. The substantial reduction in protein content observed in
S. incrassatulus (17.94% under nitrogen deficiency and 15.56% under phosphorus deficiency) and
C. vulgaris (23.83% and 23.30% under nitrogen and phosphorus deficiency, respectively) highlights this dependence. Previous research has demonstrated that nitrogen and phosphorus are essential macronutrients required for protein synthesis and overall microalgal growth (
Yang et al. 2018;
Zarrinmehr et al. 2019). Studies by
Danesh et al. (2017) showed that nitrogen starvation led to halted cell division, reduced protein production, and diminished chlorophyll synthesis. Additionally,
Gao et al. (2023) reported a decrease in protein synthesis in
S.
obliquus under nitrogen deficiency due to reduced amino acid biosynthesis and lower ribosomal protein expression. Lipid biosynthesis in both
S. incrassatulus and
C. vulgaris appeared to be largely independent of the growth phase. No significant differences were observed between the exponential and stationary phases, with
S. incrassatulus showing lipid contents of 13.92% and 11.79% in these phases, respectively, while
C. vulgaris exhibited 7.39% and 15.16%. These results align with the findings of
Schwenk et al. (2013), who reported that lipid content remained stable in brackish-water microalgae but varied significantly in marine species across different growth phases. Similarly,
You et al. (2020) found that
Dunaliella primolecta exhibited only slight increases in neutral lipid content from the latent to the exponential phase. However, under nutrient stress, particularly nitrogen deficiency, lipid accumulation increased substantially. The lipid content of
S. incrassatulus reached 45.86% under nitrogen deprivation, while
C. vulgaris exhibited a higher lipid content of 53.68%. These findings are consistent with those of
Kokabi et al. (2019), who reported that
Lobosphaera incisa accumulated triacylglycerols enriched in long-chain polyunsaturated fatty acids in response to nitrogen and phosphorus deprivation.
Huang et al. (2019) further demonstrated that nutrient limitations induced neutral lipid accumulation and membrane lipid remodeling in
Tisochrysis lutea, where the degradation and recycling of nitrogen-containing cellular compounds contributed to triacylglycerol accumulation.
Zhu et al. (2022) also observed that lipid content in S.
obliquus increased under nutrient-limited conditions, underscoring the potential of microalgae as biofuel feedstocks. The metabolic responses of
S. incrassatulus and
C. vulgaris to nutrient limitation revealed significant differences between the two species.
C. vulgaris exhibited a higher lipid accumulation potential under nitrogen stress (53.68%) compared to
S. incrassatulus (45.86%), making it a more suitable candidate for biofuel production under nitrogen-limiting conditions. Conversely,
S. incrassatulus demonstrated greater carbohydrate accumulation during the stationary phase (47.99%), suggesting its potential use in bioethanol production. These findings align with previous research highlighting
C. vulgaris as a high-lipid-producing microalga with strong industrial potential (
Mujtaba et al. 2012;
Rautenberger et al. 2024). Meanwhile,
Scenedesmus species, including
S. incrassatulus, have been recognized for their robust nutrient uptake capabilities and resilience under stress conditions, making them ideal candidates for wastewater treatment and heavy metal bioremediation (
Jácome-Pilco et al. 2009;
Mahat et al. 2024).
This study provides crucial insights into the metabolic adaptations of Chlorella vulgaris and Scenedesmus incrassatulus under varying nutrient conditions. The observed shifts in carbohydrate, protein, and lipid synthesis in response to nitrogen and phosphorus availability highlight their potential for industrial biotechnology applications, including biofuel production, bioethanol generation, and wastewater remediation. To fully harness the biotechnological potential of these microalgae, future research should focus on elucidating the genetic and molecular mechanisms driving these metabolic adaptations. Advanced omics approaches, such as transcriptomics and metabolomics, could provide a deeper understanding of how regulatory pathways govern nutrient stress responses. Additionally, genetic engineering strategies could be employed to enhance lipid accumulation or carbohydrate storage in a targeted manner, improving their suitability for specific industrial applications. Furthermore, optimizing cultivation strategies tailored to these species’ metabolic profiles could maximize their productivity under controlled conditions. Adaptive laboratory evolution and metabolic engineering approaches may enhance their resilience to fluctuating environmental factors, improving biomass yields and metabolite production efficiency. By integrating these insights with large-scale bioprocessing techniques, these microalgae could serve as sustainable resources for next-generation bio-based industries. Overall, this study underscores the metabolic plasticity of C. vulgaris and S. incrassatulus in response to nutrient stress and highlights their significance in industrial biotechnology. Future research should continue to refine cultivation and genetic modification strategies to enhance their productivity, ensuring their effective utilization in sustainable resource management and bio-based economy applications.
Conclusion
This study provides a comprehensive assessment of the mineralogical traits, metabolic responses, and adaptive strategies of two green microalgae, Chlorella vulgaris and Scenedesmus incrassatulus, isolated from Moroccan freshwater. Under varying nutrient conditions and growth phases, biochemical profiling was complemented by elemental and morphological analyses, revealing species-specific traits with strong biotechnological relevance. Molecular identification confirmed their taxonomic classification, while elemental analysis (ICP-AES) revealed clear differences in elemental composition: S. incrassatulus exhibited higher calcium and magnesium levels, whereas C. vulgaris showed increased potassium and phosphorus accumulation. These findings, supported by SEM-EDS imaging, also highlighted distinct surface morphologies. C. vulgaris presenting a smoother, interconnected structure rich in K and P, and S. incrassatulus displaying a compact, granular form enriched in Ca and Mg. Biochemical analyses revealed species-specific metabolic shifts under nutrient stress. C. vulgaris demonstrated significant lipid accumulation (53.68%) under nitrogen deficiency, supporting its potential for biodiesel production. Conversely, S. incrassatulus accumulated more carbohydrates (47.99%) during the stationary phase, making it a promising candidate for bioethanol applications. Both species showed reduced protein synthesis under stress, indicating a metabolic trade-off favoring energy storage. Altogether, these differences in biochemical and elemental traits underline the adaptive flexibility of these native strains in response to environmental stressors. The findings highlight the potential of C. vulgaris and S. incrassatulus for biofuel production, wastewater treatment, and sustainable resource management. Further research should focus on optimizing cultivation strategies and investigating the role of trace elements in metabolic regulation to enhance their industrial viabilit
Fig. 1Microscopic observation of Scenedesmus incrassatulus (left) and Chlorella vulgaris (right). (Inverted microscope Olympus IMT2) (× 400).
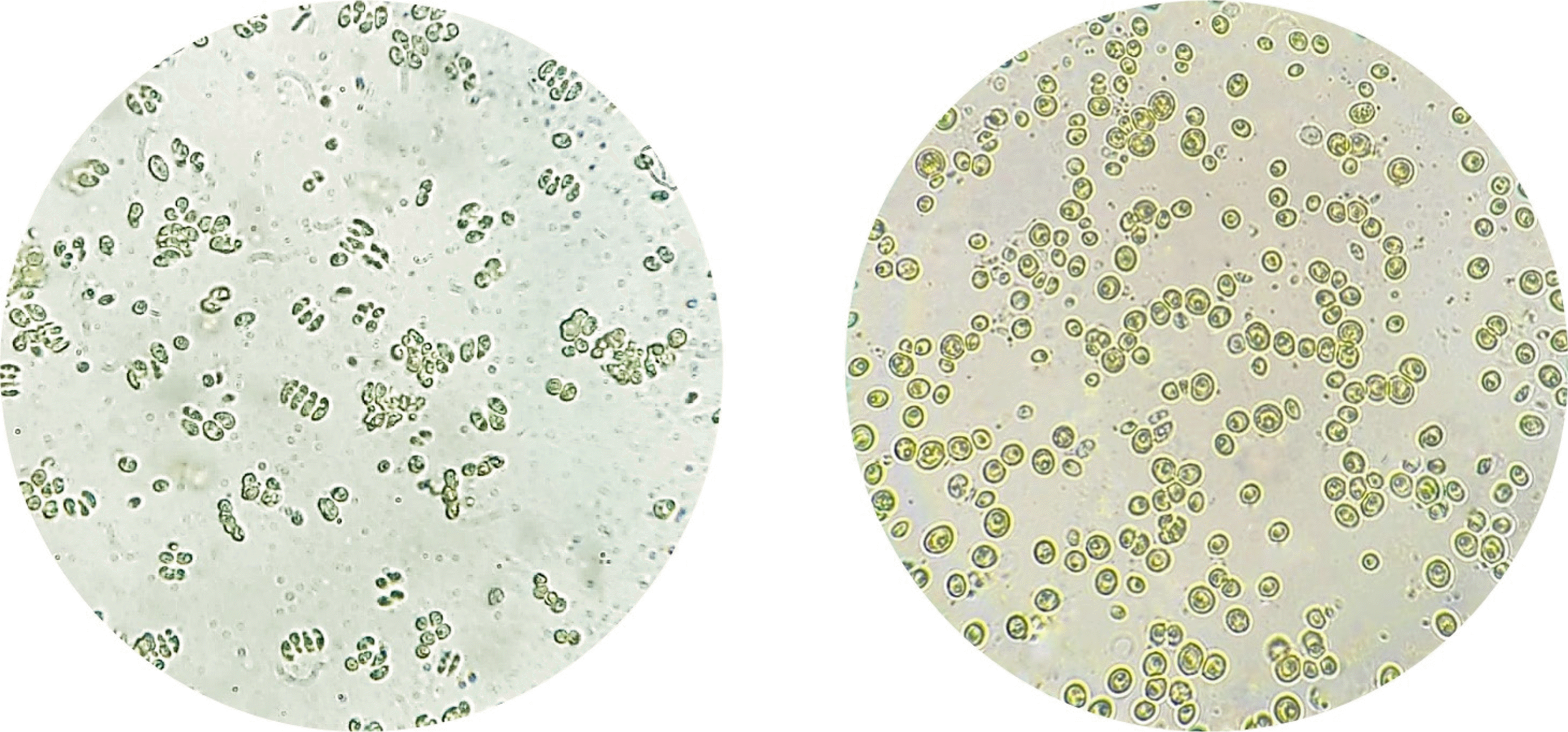
Fig. 2Experimental protocol of biomass recovery.
Fig. 3Phylogenetic tree of Chlorella vulgaris isolate based on ITS sequences, constructed using MEGA software. Bootstrap values (%) are shown at the nodes.

Fig. 4Phylogenetic tree of Scenedesmus incrassatulus isolate based on ITS sequences, constructed using MEGA software. Bootstrap values (%) are shown at the nodes.

Fig. 5SEM images illustrating the distinct surface morphologies of (a) S. incrassatulus and (b) C. vulgaris at ×1000 magnification.

Fig. 6Growth curve of S. incrassatulus (A) and of C. vulgaris (B) in a complete medium. The black line shows cell density (cells mL-1; mean values), the green line shows the chlorophyll “a” concentration (mg mL-1; mean values). Error bars show the standard deviation (n = 2).
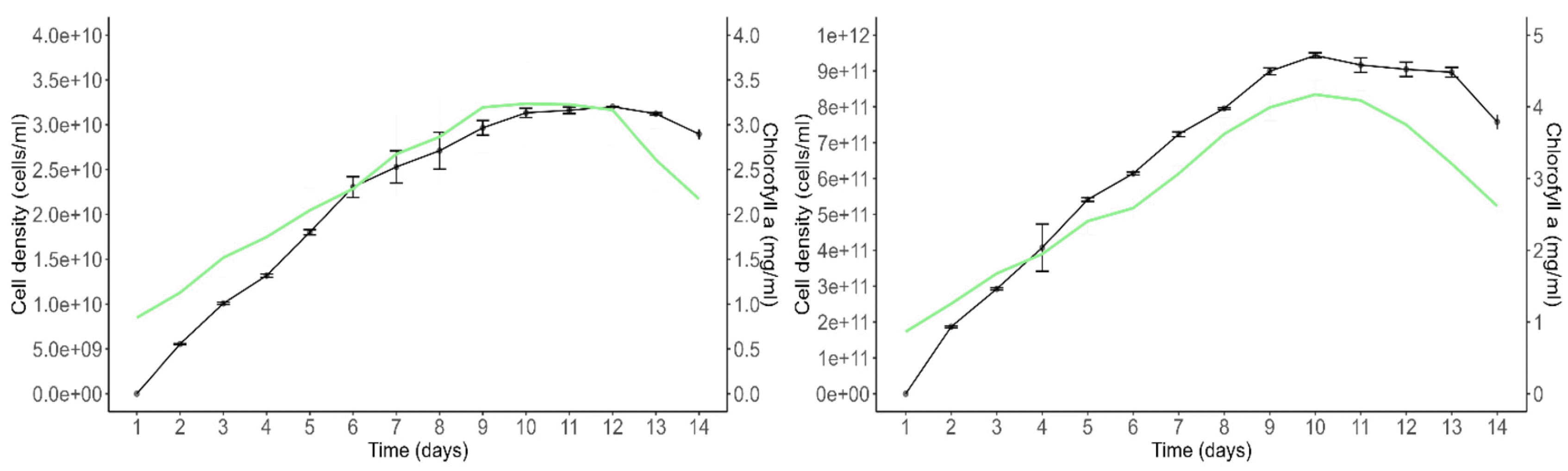
Fig. 7Growth curve of S. incrassatulus (A) and C. vulgaris (B) in N and P depleted medium.
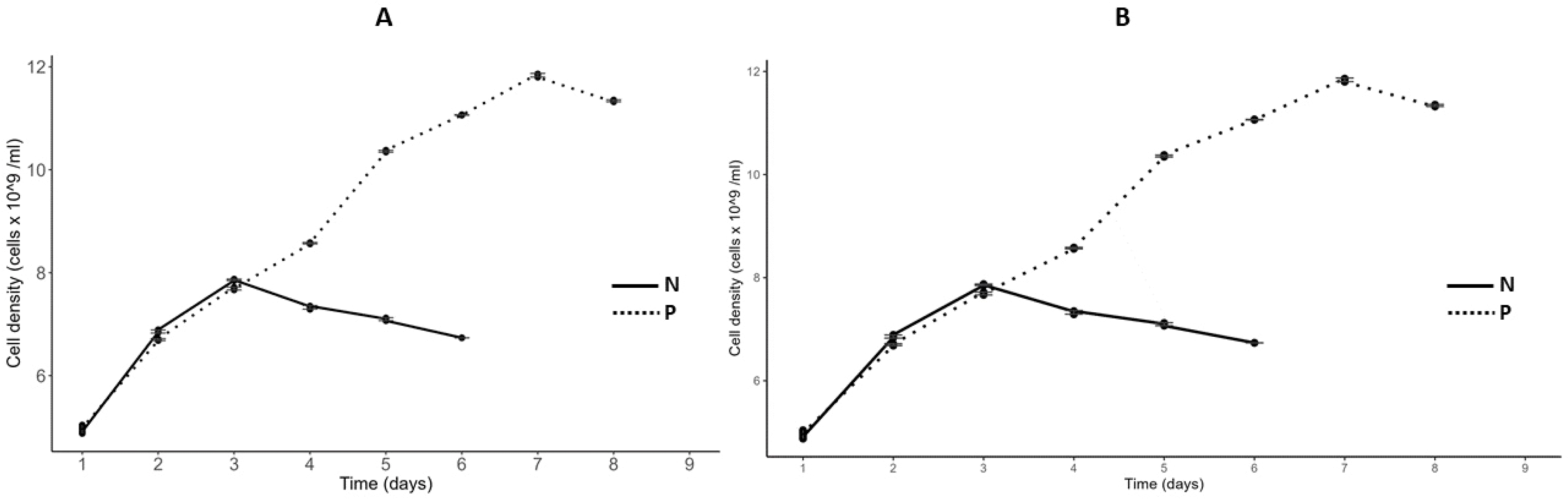
Fig. 8Carbohydrate content (%; mean value) of S. incrassatulus (A&B) and C. vulgaris (C&D) in harvested in the exponential phase and stationary phase (A&C) and in N-depleted medium and P-depleted biomass medium (B&D). Error bars show the standard deviation. Different letters (a&b) show statistically significant differences. T-test (p<0.001; A&C), one-way ANOVA (p<0.001), Tukey’s Range Test, n = 3, α = 0.05 (B&D).
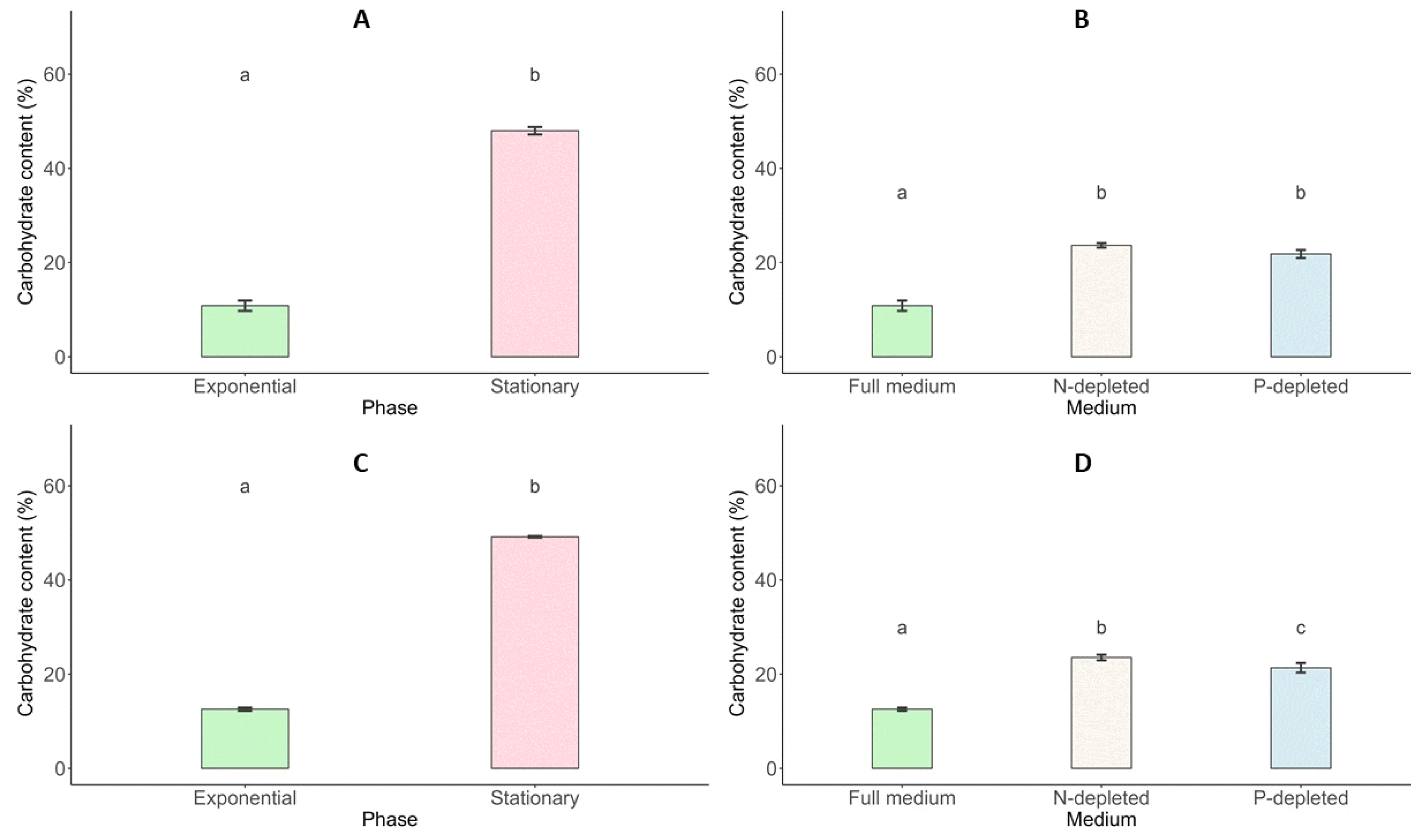
Fig. 9Protein content (%; mean value) of S. incrassatulus (A&B) and C. vulgaris (C&D) in biomass harvested in the exponential phase and stationary phase (A&C) and in N-depleted medium and P-depleted medium (B&D). Error bars show the standard deviation. Different letters (a&b) show statistically significant differences. T-test (p<0.001; A&C), one-way ANOVA (p<0.001), Tukey’s Range Test, n = 3, α = 0.05 (B&D).
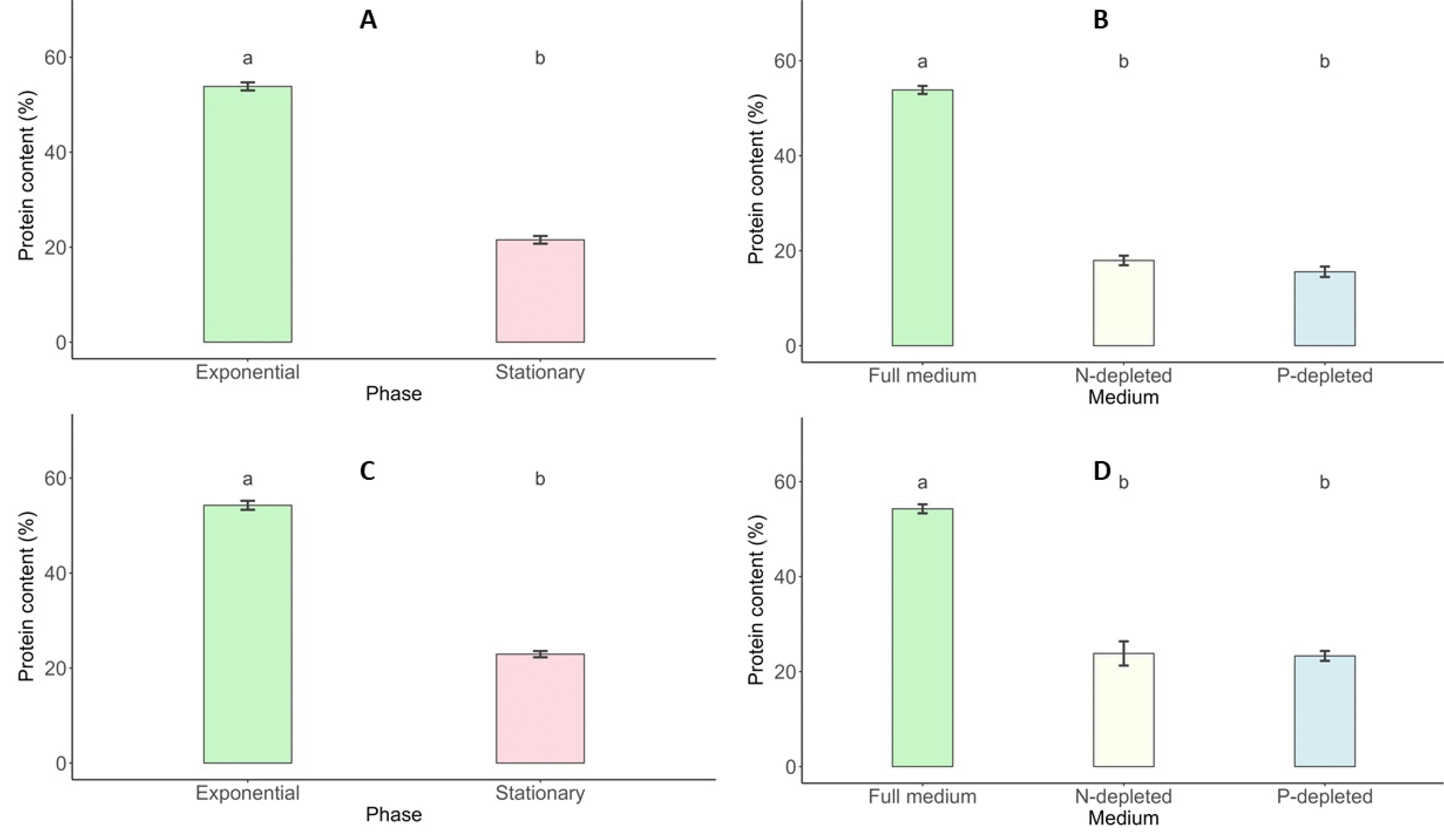
Fig. 10Lipid content (%; mean value) of S. incrassatulus (A&B) and C. vulgaris (C&D) in biomass harvested in the exponential phase and stationary phase (A&C) and in N-depleted medium and P-depleted medium (B&D). Error bars show the standard deviation. Different letters (a&b) show statistically significant differences. T-test (p<0.001; A&C), one-way ANOVA (p<0.001), Tukey’s Range Test, n = 3, α = 0.05 (B&D).
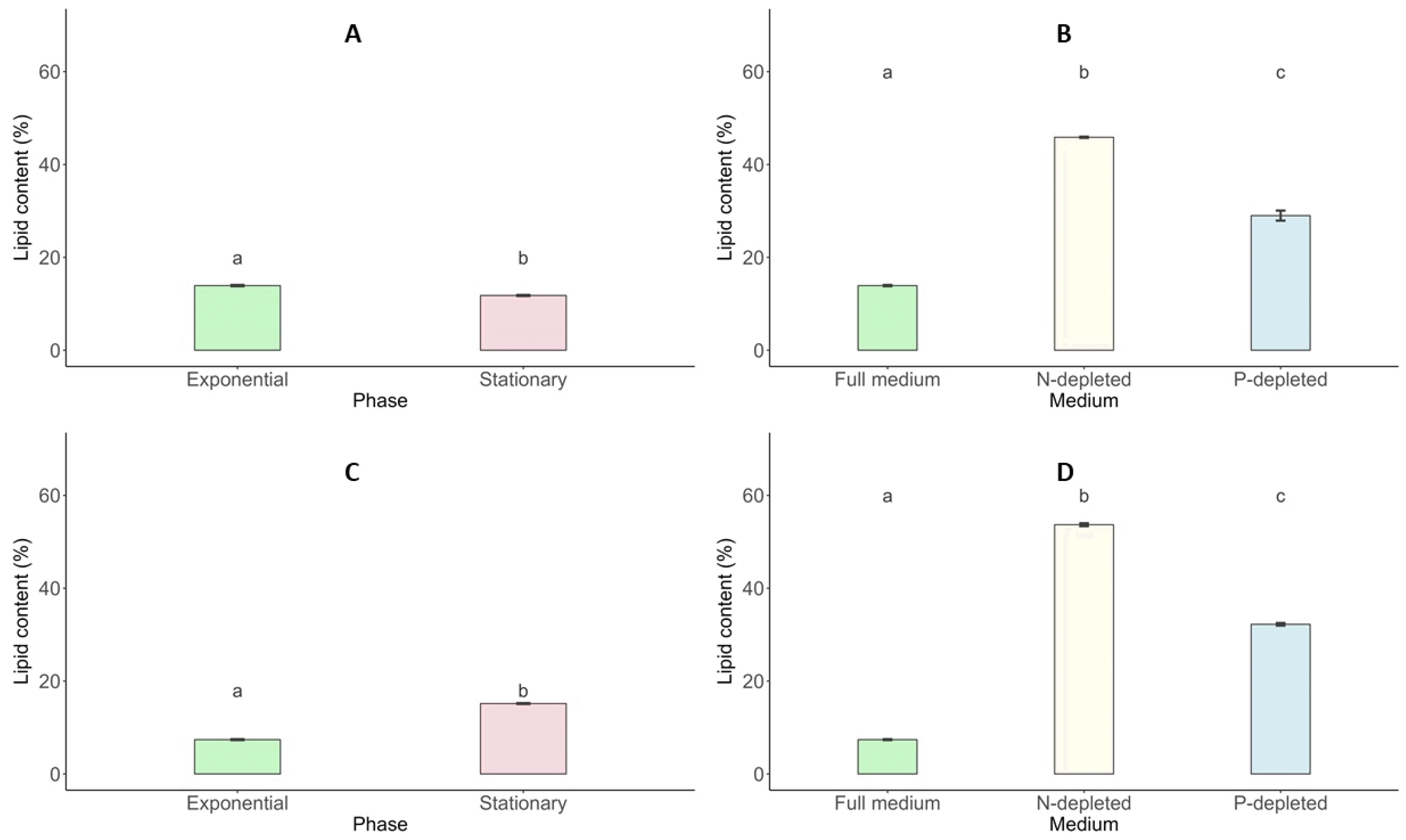
Table 1Culture medium’s chemical composition (
Dauta 1982).
Table 1
|
Chemical Composition |
Concentration (mg. L-1) |
|
Base medium |
MgSO4, 7H2O
FeSO4, 7H2O
CaCl2, 2H2O
Na2EDTA
NaHCO3
Na2CO3
|
25
1
25
1
50
5 |
|
|
Trace elements |
ZnSO4, 7H2O
CuCl2, 2H2O
MnCl2, 4H2O
CoCl2, 6H2O
BO3H3
Na2MoO4
|
0.020
0.020
0.400
0.01
0
0.001
0.035 |
|
|
Nutrients |
KNO3
K2HPO4
|
200
25
|
Table 2Elemental Composition of Microalgae by ICP-AES.
Table 2
|
Element |
Unit |
Chlorella vulgaris (CV) |
Scenedesmus incrassatulus (SI) |
|
Macronutrient |
Calcium (Ca) |
mg g-1
|
16.824 |
20.31 |
|
Magnesium (Mg) |
mg g-1
|
4.848 |
5.188 |
|
Potassium (K) |
mg g-1
|
4.510 |
1.468 |
|
Phosphorus (P) |
mg g-1
|
2.855 |
2.055 |
|
|
Trace Element |
Aluminum (Al) |
mg g-1
|
0.057 |
0.083 |
|
Boron (Bo) |
mg g-1
|
0.00787 |
0.0035 |
|
Iron (Fe) |
mg g-1
|
0.132 |
0.103 |
|
Silicon (Si) |
mg g-1
|
0.365 |
0.382 |
|
Zinc (Zn) |
mg g-1
|
0.081 |
0.0777 |
Table 3Elemental Composition of Scenedesmus incrassatulus and Chlorella vulgaris. Mean values ± standard deviations.
Table 3
|
Element |
Line |
Mass% (SI) |
Atom% (SI) |
Mass% (CV) |
Atom% (CV) |
Measurement Conditions |
Detector |
|
C |
K |
58.49 ± 0.12 |
66.39 ± 0.13 |
57.75 ± 0.15 |
66.55 ± 0.17 |
20.0 kV, x1000 |
First Detector |
|
O |
K |
37.47 ± 0.26 |
31.93 ± 0.22 |
35.64 ± 0.32 |
30.83 ± 0.28 |
30 seconds live time |
High Vacuum Detector |
|
K |
K |
1.78 ± 0.03 |
0.62 ± 0.01 |
4.03 ± 0.05 |
1.43 ± 0.02 |
T2 Process |
Standardless ZAF |
|
P |
K |
1.22 ± 0.02 |
1.06 ± 0.02 |
1.50 ± 0.03 |
1.19 ± 0.03 |
ZAF Quantification Method |
High Vacuum Detector |
Table 4Growth parameters, obtained from the fitted logistic growth model.
Table 4
|
S. incrassatulus
|
C. vulgaris
|
|
Carrying capacity (K) |
3.11⋅1010 |
8.91⋅1011 |
|
Standard error |
6.53⋅108 |
2.89⋅1010 |
|
p-value |
<0.001 |
<0.001 |
|
Initial population size (N0) |
1.57⋅109 |
4.88⋅1010 |
|
Standard error |
4.20⋅108 |
1.97⋅1013 |
|
p-value |
0.003 |
0.03 |
|
Growth rate (r) |
0.655 |
0.650 |
|
Standard error |
0.063 |
0.098 |
|
p-value |
<0.001 |
<0.001 |
|
Residual standard error a |
1.37⋅109 |
6.10⋅1010 |
|
Time at inflection point b |
4.47 |
4.38 |
Table 5Protein, carbohydrate, and lipid content of C. vulgaris and S. incrassatulus. Mean values ± standard deviations are shown. T-test (p<0.001) to compare growth phase (test 1), one-way ANOVA (p<0.001), Tukey’s Range Test to compare medium (test 2), n = 3, α = 0.05.
Table 5
|
Biomass |
Chlorella vulgaris
|
|
Scenedesmus incrassatulus |
|
|
|
Protein |
Carbohydrate |
Lipid |
Protein |
Carbohydrate |
Lipid |
|
Exponential |
54.28±0.94 |
54.28 ± 0.94 |
12.57 ± 0.35 |
|
7.39 ± 0.12 |
53.85 ± 0.85 |
10.84 ± 1.09 |
|
Stationary |
22.93±0.66 |
22.93 ± 0.66 |
49.18 ± 0.17 |
|
15.16 ± 0.11 |
21.54 ± 0.81 |
48.00 ± 0.79 |
|
N-depleted |
23.82±2.55 |
23.82 ± 2.55 |
23.54 ± 0.60 |
|
53.68 ± 0.33 |
17.94 ± 1.01 |
23.65 ± 0.49 |
|
P-depleted |
23.30±1.04 |
23.30 ± 1.04 |
21.35 ± 1.02 |
|
32.23 ± 0.28 |
15.56 ± 1.10 |
21.82 ± 0.84 |
|
p-value (test 1 and 2) |
<0.001 |
<0.001 |
<0.001 |
|
<0.001 |
<0.001 |
<0.001 |
References
- Ajala SO, Alexander ML. 2020. Assessment of chlorella vulgaris, scenedesmus obliquus, and oocystis minuta for removal of sulfate, nitrate, and phosphate in wastewater. Int. J. Energy Environ. Eng. 11: 311-326.
- Alayi M, Mohammadi A, Mashhadi H, Mahmoudnia F. 2021. Investigating the capability of chromium heavy metal removal and biodiesel production by three species of algae: Scenedesmus acutus, scenedesmus incrass atulus, scenedesmus obliquus. Ann. Rom. Soc. Cell Biol. 25(4): 21127-21138.
- Alcántara C, Fernández C, García-Encina PA, Muñoz R. 2015. Mixotrophic metabolism of chlorella sorokiniana and algal-bacterial consortia under extended dark-light periods and nutrient starvation. Appl. Microbiol. Biotechnol. 99: 2393-2404.
- Ali SS, El-Sheekh M, Manni A, Ruiz HA, Elsamahy T, Sun J, et al. 2022. Microalgae-mediated wastewater treatment for biofuels production: A comprehensive review. Microbiol. Res. 265: 127187
- Belkoura M, Dauta A. 1994. Stockage intracellulaire des nutriments chez l'algue verte Chlorella sorokiniana Shihira & Krauss. Ann. Limnol. EDP Sciences. 313-321..
- Benavente-Valdés JR, Aguilar C, Contreras-Esquivel JC, Méndez-Zavala A, Montañez J. 2016. Strategies to enhance the production of photosynthetic pigments and lipids in chlorophycae species. Biotechnol. Rep. 10: 117-125.
- Birkholz M, Malti DE, Hartmann S, Neubauer P. 2022. Separation of heterotrophic microalgae crypthecodinium cohnii by dielectrophoresis. Front. Bioeng. Biotechnol. 10: 855035
- Bligh EG, Dyer WJ. 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37: 911-917.
- Bouchamma EO, Derraz M, Naji B, Dauta A. 2004. Influence des conditions nutritionnelles sur la croissance et les capacités de stockage intracellulaire (azote et phosphore) chez Planktothrix agardhii isolée des eaux de la retenue eutrophe El Kansera (Maroc). Acta. Bot. Gall. 151: 381-392.
- Bourrelly P. 1988. Les algues d'eau douce: Compléments tome 1: Algues vertes, Faunes et flores actuelles. Société nouvelle des éditions Boubée. Paris.
- Chen X, Zhao J, Zhang X, Song M, Ye X. 2022. Self-regulation mechanism difference of Chlorella vulgaris and Scenedesmus obliquus in toxic sludge extract caused by hydroquinone biodegradation. Environ. Res. 214: 114107
- Chong JF, Fadhullah W, Lim V, Lee CK. 2019. Two-stage cultivation of the marine microalga Chlorella salina for starch and carbohydrate production. Aquacult. Int. 27: 1269-1288.
- Crofcheck CL, E X, Shea A, Montross MD, Crocker M. 2013. Influence of media composition on the growth rate of chlorella vulgaris and scenedesmus acutus utilized for CO2 mitigation. Biosystems and Agricultural Engineering Faculty Publications. 48.
- da Costa F, Le Grand F, Quéré C, Bougaran G, Cadoret JP, Robert R, et al. 2017. Effects of growth phase and nitrogen limitation on biochemical composition of two strains of Tisochrysis lutea. Algal Res. 27: 177-189.
- Danesh AF, Ebrahimi S, Salehi A, Parsa A. 2017. Impact of nutrient starvation on intracellular biochemicals and calorific value of mixed microalgae. Biochem. Eng. J. 125: 56-64.
- Dauta A. 1982. Conditions de développement du phytoplancton. Etude comparative du comportement de huit espèces en culture. I. Détermination des paramètres de croissance en fonction de la lumière et de la température. Ann. Limnol. 18: 217-262.
- de Carvalho Silvello MA, Gonçalves IS, Azambuja SPH, Costa SS, Silva PGP, Santos LO, et al. 2022. Microalgae-based carbohydrates: A green innovative source of bioenergy. Bioresour. Technol. 344: 126304
- Debnath C, Bandyopadhyay TK, Bhunia B, Mishra U, Narayanasamy S, Muthuraj M. 2021. Microalgae: Sustainable resource of carbohydrates in third-generation biofuel production. Renew. Sustain. Energy Rev. 150: 111464
- Dhanasekar S, Sathyanathan R. 2023. Bioenergy potential of Chlorella vulgaris under the influence of different light conditions in a bubble column photobioreactor. Glob. J. Environ. Sci. Manag. 9: 789-804.
- Dolganyuk V, Belova D, Babich O, Prosekov A, Ivanova S, Katserov D, et al. 2020. Microalgae: A promising source of valuable bioproducts. Biomolecules. 10: 1153
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19(1): 11-15.
- Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. 1951. A colorimetric method for the determination of sugars. Nature. 168: 167-167.
- Fathy WA, Techen N, Elsayed KNM, Essawy EA, Tawfik E, Alwutayd KM, et al. 2023. Applying an internal transcribed spacer as a single molecular marker to differentiate between Tetraselmis and Chlorella species. Front. Microbiol. 14: 1228869
- Felsenstein J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 39: 783-791.
- Fernandes T, Fernandes I, Andrade CAP, Cordeiro N. 2016. Changes in fatty acid biosynthesis in marine microalgae as a response to medium nutrient availability. Algal Res. 18: 314-320.
- Fihri RF, Ez-Zoubi A, Mbarkiou L, Amar A, Farah A, Bouchamma EO. 2024. Antibacterial and antioxidant activities of Chlorella vulgaris and Scenedesmus incrassatulus using natural deep eutectic solvent under microwave assisted by ultrasound. Heliyon. 10(15): e35071
- Folch J, Lees M, Sloane Stanley GH. 1957. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226: 497-509.
- Gao L, Ding W, Xi J, Gao S, Zhou X, Chen Y, et al. 2023. Effects of different nitrogen/phosphorus ratios on the growth and metabolism of microalgae Scenedesmus obliquus cultured in the mixed wastewater from primary settling tank and sludge thickener. Process Saf. Environ. Prot. 170: 824-833.
- Gomaa MA, Refaat MH, Salim TM, El-Sayed AE-KB, Bekhit MM. 2019. Identification of green alga chlorella vulgaris isolated from freshwater and improvement biodiesel productivity via uv irradiation. Microbiol. Biotechnol. Lett. 47: 381-389.
- González-Fernández C, Ballesteros M. 2012. Linking microalgae and cyanobacteria culture conditions and key-enzymes for carbohydrate accumulation. Biotechnol. Adv. 30: 1655-1661.
- Goswami RK, Agrawal K, Shah MP, Verma P. 2022. Bioremediation of heavy metals from wastewater: A current perspective on microalgae-based future. Lett. Appl. Microbiol. 75: 701-717.
- He M, Yan Y, Pei F, Wu M, Gebreluel T, Zou S, et al. 2017. Improvement on lipid production by Scenedesmus obliquus triggered by low dose exposure to nanoparticles. Sci. Rep. 7: 15526
- Hothorn T, Bretz F, Westfall P. 2008. Simultaneous inference in general parametric models. Biom. J. 50: 346-363.
- Huang B, Marchand J, Thiriet-Rupert S, Carrier G, Saint-Jean B, Lukomska E, et al. 2019. Betaine lipid and neutral lipid production under nitrogen or phosphorus limitation in the marine microalga Tisochrysis lutea (Haptophyta). Algal Res. 40: 101506
- Jácome-Pilco CR, Cristiani-Urbina E, Flores-Cotera LB, Velasco-García R, Ponce-Noyola T, Cañizares-Villanueva RO. 2009. Continuous Cr(VI) removal by Scenedesmus incrassatulus in an airlift photobioreactor. Bioresour. Technol. 100: 2388-2391.
- Kezlya E, Tseplik N, Kulikovskiy M. 2023. Genetic markers for metabarcoding of freshwater microalgae: Review. Biology. 12: 1038
- Khan MI, Shin JH, Kim JD. 2018. The promising future of microalgae: current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Factories. 17: 36
- Khoo KS, Ahmad I, Chew KW, Iwamoto K, Bhatnagar A, Show PL. 2023. Enhanced microalgal lipid production for biofuel using different strategies including genetic modification of microalgae: A review. Prog. Energy Combust. Sci. 96: 101071
- Kokabi K, Gorelova O, Ismagulova T, Itkin M, Malitsky S, Boussiba S, et al. 2019. Metabolomic foundation for differential responses of lipid metabolism to nitrogen and phosphorus deprivation in an arachidonic acid-producing green microalga. Plant Sci. 283: 95-115.
- Korozi E, Kefalogianni I, Tsagou V, Chatzipavlidis I, Markou G, Karnaouri A. 2023. Evaluation of growth and production of high-value-added metabolites in scenedesmus quadricauda and chlorella vulgaris grown on crude glycerol under heterotrophic and mixotrophic conditions using monochromatic light-emitting diodes (LEDs). Foods. 12: 3068
- Kumar S, Stecher G, Suleski M, Sanderford M, Sharma S, Tamura K. 2024. MEGA12: Molecular evolutionary genetic analysis version 12 for adaptive and green computing. Mol. Biol. Evol. 41: msae263
- Kunyalung W, Putanyawiwat P, Chaipanya C, Hongtrakul V. 2021. An an efficiency of DNA extraction methods for green microalgae. Recent Sci. Technol. 13: 742-753.
- Lai YH, Puspanadan S, Lee CK. 2019. Nutritional optimization of Arthrospira platensis for starch and Total carbohydrates production. Biotechnol. Prog. 35: e2798
- Lenth RV. 2016. Least-squares means: The R package lsmeans. J. Stat. Softw. 69: 1-33.
- Lima I, Martinez S, Teixeira V, Gonzalez W. 2019. Morphological analysis by scanning electron microscopy of dictyota menstrualis in natura and following an extraction process. Chem. Eng. Trans. 75: 571-576.
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265-275.
- Mahat SB, Abobaker MSA, Chun CNW, Wibisono Y, Ahmad AL, Omar WMW, et al. 2024. Scenedesmus sp. as a phycoremediation agent for heavy metal removal from landfill leachate in a comparative study: Batch, continuous, and membrane bioreactor (MBR). Environ. Sci. Pollut. Res. 31: 50443-50463.
- Marker AFH. 1994. Chlorophyll-a-SCA mehtod revision.
- Mendes AR, Spínola MP, Lordelo M, Prates JAM. 2024. Chemical compounds, bioactivities, and applications of chlorella vulgaris in food, feed and medicine. Appl. Sci. 14: 10810
- Michalak I, Marycz K, Basińska K, Chojnacka K. 2014. Using SEM-EDX and ICP-OES to investigate the elemental composition of green macroalga vaucheria sessilis. Sci. World J. 2014: 891928
- Mujtaba G, Choi W, Lee C-G, Lee K. 2012. Lipid production by Chlorella vulgaris after a shift from nutrient-rich to nitrogen starvation conditions. Bioresour. Technol. 123: 279-283.
- Olguín EJ, Sánchez-Galván G, Arias-Olguín II, Melo FJ, González-Portela RE, Cruz L, et al. 2022. Microalgae-based biorefineries: Challenges and future trends to produce carbohydrate enriched biomass, High-added value products and bioactive compounds. Biology. 11: 1146
- Peña-Castro JM, Martínez-Jerónimo F, Esparza-García F, Cañizares-Villanueva RO. 2004. Heavy metals removal by the microalga Scenedesmus incrassatulus in continuous cultures. Bioresour. Technol. 94: 219-222.
- Piasecka A, Baier A. 2022. Metabolic and proteomic analysis of Chlorella sorokiniana, Chloroidium saccharofilum, and Chlorella vulgaris cells cultured in autotrophic, photoheterotrophic, and mixotrophic cultivation modes. Molecules. 27: 4817
- R Core Team.2021. R: A language and environment for statistical computing.
- Rafay R, Uratani JM, Hernandez HH, Rodríguez J. 2020. Growth and nitrate uptake in nannochloropsis gaditana and tetraselmis chuii cultures grown in sequential batch reactors. Front. Mar. Sci. 7: 77
- Rai H. 1980. The measurement of photosynthetic pigments in freshwater and standardization of methods. Schweizerbart'sche Verlagsbuchhandlung..
- Ratomski P, Hawrot-Paw M. 2021. Influence of nutrient-stress conditions on chlorella vulgaris biomass production and lipid content. Catalysts. 11: 573
- Rautenberger R, Détain A, Skjånes K, Schulze PSC, Kiron V, Morales-Sánchez D. 2024. Growth strategies of Chlorella vulgaris in seawater for a high production of biomass and lipids suitable for biodiesel. Algal Res. 77: 103360
- Saitou N, Nei M. 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4: 406-425.
- Salmi P, Eskelinen MA, Leppänen MT, Pölönen I. 2021. Rapid quantification of microalgae growth with hyperspectral camera and vegetation indices. Plants. 10: 341
- Sánchez-Bayo A, Morales V, Rodríguez R, Vicente G, Bautista LF. 2020. Cultivation of microalgae and cyanobacteria: Effect of operating conditions on growth and biomass composition. Molecules. 25: 2834
- Sarwa P, Verma SK. 2017. Identification and characterization of green microalgae, scenedesmus sp. MCC26 and acutodesmus obliquus MCC33 isolated from industrial polluted site using morphological and molecular markers. Int. J. Appl. Sci. Biotechnol. 5: 415-422.
- Schüler L, Greque de Morais E, Trovão M, Machado A, Carvalho B, Carneiro M, et al. 2020. Isolation and characterization of novel chlorella vulgaris mutants with low chlorophyll and improved protein contents for food applications. Front. Bioeng. Biotechnol. 8: 469
- Schwenk D, Seppälä J, Spilling K, Virkki A, Tamminen T, Oksman-Caldentey K-M, et al. 2013. Lipid content in 19 brackish and marine microalgae: influence of growth phase, salinity and temperature. Aquat. Ecol. 47: 415-424.
- Shah S, Li X, Jiang Z, Fahad S, Hassan S. 2022. Exploration of the phytohormone regulation of energy storage compound accumulation in microalgae. Food Energy Secur. 11: e418
- Spain O, Funk C. 2022. Detailed characterization of the cell wall structure and composition of nordic green microalgae. J. Agric. Food Chem. 70: 9711-9721.
- Sprouffske K, Wagner A. 2016. Growthcurver: An R package for obtaining interpretable metrics from microbial growth curves. BMC Bioinformatics. 17: 1-4.
- Tamura K, Nei M. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10: 512-526.
- Tibbetts SM, Milley JE, Lall SP. 2015. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 27: 1109-1119.
- Tripathi S, Choudhary S, Meena A, Poluri KM. 2023. Carbon capture, storage, and usage with microalgae: A review. Environ. Chem. Lett. 21: 2085-2128.
- Trivedi J, Agrawal D, Atray N, Ray A. 2022. Enhanced lipid production in Scenedesmus obliquus via nitrogen starvation in a two-stage cultivation process and evaluation for biodiesel production. Fuel. 316: 123418
- Udayan A, Pandey AK, Sirohi R, Sreekumar N, Sang B-I, Sim SJ, et al. 2023. Production of microalgae with high lipid content and their potential as sources of nutraceuticals. Phytochem. Rev. 22: 833-860.
- Wickham H. 2009. ggplot2: Elegant graphics for data analysis. Springer-Verlag. New York:
- Wickham H. 2016. ggplot2, Use R!. Springer International Publishing. Cham:
- Yaakob MA, Mohamed RMSR, Al-Gheethi A, Aswathnarayana Gokare R, Ambati RR. 2021. Influence of nitrogen and phosphorus on microalgal growth, biomass, lipid, and fatty acid production: An overview. Cells. 10: 393
- Yang F, Xiang W, Li T, Long L. 2018. Transcriptome analysis for phosphorus starvation-induced lipid accumulation in Scenedesmus sp. Sci. Rep. 8: 16420
- You J, Mallery K, Mashek DG, Sanders M, Hong J, Hondzo M. 2020. Microalgal swimming signatures and neutral lipids production across growth phases. Biotechnol. Bioeng. 117: 970-980.
- Zarrinmehr M, Farhadian O, Paykan-Heyrati F, Keramat J, Koutra E, Kornaros M, et al. 2019. Effect of nitrogen concentration on the growth rate and biochemical composition of the microalga, Isochrysis galbana. Egypt. J. Aquat. Res. 46(2): 153-158.
- Zhu Z, Sun J, Fa Y, Liu X, Lindblad P. 2022. Enhancing microalgal lipid accumulation for biofuel production. Front. Microbiol. 13: 1024441