Abstract
This study aimed to enhance the Inpari 42 rice variety by integrating the aromatic trait through Marker-Assisted Backcrossing (MAB) while improving key agronomic characteristics in the BC1F1 generation. The aromatic gene from Merah Wangi was successfully introgressed into BC1F1 plants, confirmed through molecular analysis using the Bradbury marker, which detects mutations in the BADH2 gene responsible for fragrance in rice. Thirty BC1F1 plants with a heterozygous mutation in BADH2 were identified, demonstrating the reliability of MAB in maintaining the aromatic trait across generations. Agronomic evaluations revealed that BC1F1 plants exhibited intermediate flowering time, increased plant height, and longer panicles compared to the parental lines. Additionally, BC1F1 plants showed enhanced tillering capacity, more productive tillers, and higher grain yield per plant, indicating the successful integration of desirable traits from both parents. The next step in this research is to conduct further backcrosses with Inpari 42 to develop a stable aromatic variety, combining the high-yielding characteristics of Inpari 42 with the aromatic traits of Merah Wangi. These findings highlight the potential of BC1F1 lines for developing high-yielding aromatic rice varieties suitable for diverse agricultural settings.
-
Key words: Aromatic rice breeding, BADH2 gene, Marker-Assisted Backcrossing, BC1F1, Agronomic traits
Introduction
Aromatic rice, which is renowned for its distinctive fragrance, holds a significant position in the global rice market. The characteristic aroma of aromatic rice is primarily attributed to 2-acetyl-1-pyrroline (2AP), which is popular among consumers and, leads to higher market prices than those of non-aromatic varieties (
Buttery et al. 1982;
Calingacion et al. 2014;
Okpala et al. 2019). This growing consumer demand has incentivized the production and breeding of fragrant rice varieties (
Imran et al. 2023a). Advances in genetic technologies, including genome editing, have played a pivotal role in meeting this demand by enhancing the expression of fragrant genes, thereby converting nonfragrant cultivars into aromatic cultivars (
Ashokkumar et al. 2020;
Imran et al. 2023b). Furthermore, the integration of fragrance genes with other desirable traits has resulted in the development of elite rice varieties that possess an enhanced aroma and exhibit superior agronomic traits (
Singh et al. 2012;
Vanavichit et al. 2018;
Wang et al. 2023). These varieties not only possess enhanced fragrance, but also exhibit other superior traits, making them attractive to both farmers and consumers. This combination of attributes ensures that aromatic rice remains a viable and profitable crop, further driving global demand.
The fragrance trait in rice is primarily controlled by the BADH2 gene on chromosome 8, which encodes the enzyme betaine aldehyde dehydrogenase. A specific and well-known mutation an 8-base pair deletion in exon 7 disrupts the enzyme’s function, resulting in the accumulation of 2AP (
Bradbury et al. 2005a;
Chen et al. 2008). Although BADH2 is also associated with certain stress tolerance functions, its loss of activity does not appear to negatively affect plant growth. A functional molecular marker developed to detect this mutation has been widely used due to its high accuracy, cost-effectiveness, and perfect co-segregation with the aroma phenotype in segregating rice populations (
Bradbury et al. 2005b).
Marker-Assisted Backcrossing (MAB) has significantly improved the efficiency of incorporating aromatic traits into near-isogenic lines (NILs) of aromatic rice. By utilizing molecular markers, MAB accelerates the breeding process and ensures the precise introgression of target traits. In aromatic rice breeding, MAB enables rapid and accurate selection of plants carrying the desired aromatic trait. For instance, in a study by Ishak and Fedora, marker-assisted selection (MAS) was used to identify the fragrance gene in rice, resulting in 75% of the F2 generation exhibiting the aroma trait, which remained consistent in subsequent generations (
Ishak et al. 2024). This consistency demonstrates MAB's effectiveness in maintaining trait homogeneity and stability across generations. Additionally, the successful introgression of bacterial blight resistance genes into aromatic rice cultivars highlights MAB's dual advantage in enhancing both aroma and disease resistance (
Kumar et al. 2023;
Sagar et al. 2020). These findings emphasize the pivotal role of MAB in the development of robust aromatic rice varieties with superior agronomic performance.
In this study, Inpari 42 was selected as the recurrent parent because it is a high-yielding, early-maturing rice variety that is resistant to Rice Ragged Stunt Virus (RRSV) and widely cultivated across Indonesia; however, it does not belong to the aromatic rice group (
Kurniawati et al. 2022). In contrast, Merah Wangi is a local red rice landrace from East Java Province, renowned for its strong fragrance (
Fanata et al. 2021;
Ramadhan et al. 2020). However, it has relatively low yield performance, limited agronomic stability under intensive cultivation, and produces grains that are prone to breakage during post-harvest processing, which reduces its milling and market quality.
The objective of this research was to introgress the aromatic trait from Merah Wangi into the high-yielding genetic background of Inpari 42, with the aim of developing a novel rice line that combines strong aroma with superior agronomic performance to meet both market demands and consumer preferences. To achieve this, F1 hybrids were initially generated from a cross between Inpari 42 and Merah Wangi. In the present study, the first backcross generation (BC1F1) was developed by backcrossing Inpari 42 with F1 plants carrying the BADH2 mutation, which controls fragrance. This stage represents the initial step in a series of planned backcrosses toward the development of a genetically stable aromatic rice variety based on Inpari 42. Agronomic and molecular characterizations were performed to assess the performance of BC1F1 individuals and to identify promising lines for further backcrossing. This study contributes to the early phase of a breeding program aimed at producing a stable, high-yielding aromatic rice variety that is adaptable to diverse agricultural environments and aligned with national rice improvement goals.
Materials and Methods
Plant Materials and Growth Conditions
The seeds of the Inpari 42, Merah Wangi, and BC
1F
1 lines were stored for 7 d at 50℃ to break dormancy (
Wan et al. 1997). The seeds were germinated on moist straw paper for 3 days at 30℃ under dark conditions, and the germinated seeds were subsequently cultivated on cubed rockwool for 2 weeks and irrigated with half-strength Yoshida nutrition (pH 5.0). Two-week-old seedlings were then grown in natural soil in buckets in an open field protected with a 0.75-inch net to prevent seed-eating bird predation and ensure optimal light exposure for the plants. Regular irrigation was provided, and fertilization was performed using urea, monocalcium phosphate, and potassium chloride at rates equivalent to 200, 50, and 50 kg/ha, respectively. Comprehensive pest and disease management protocols were rigorously implemented throughout the cultivation period to maintain optimal plant health and minimize potential yield loss.
Genomic DNA was extracted from the leaf tissue using a buffer containing 200 mM Tris-Cl, 250 mM NaCl, 25 mM EDTA, and 0.5% SDS. DNA was extracted using an equal volume of phenol-chloroform-isoamyl alcohol (HiMedia), and DNA precipitation was performed using 0.8 volume of isopropanol. The precipitated DNA was washed with 70% ethanol, and the dried DNA was solubilized in 50 µL TE buffer (pH 8.0) and stored at -20℃. For genotyping, 2 µL of genomic DNA was used as the template in polymerase chain reaction (PCR) using GoTaq® Green Master Mix (Promega) in combination with Bradbury markers. PCR was performed using a T100 Thermal Cycler (Bio-Rad). The employed Bradbury markers consist of ESP (External Sense Primer) 5‵-TTGTTTGGAGCTTGCTGATG-3‵, EAP (External Antisense Primer) 5‵-AGTGCTTTACAAAGTCCCGC-3‵, INSP (Internal Non Sense Primer) 5‵-CTGGTAAAAAGATTATGGCTTCA-3‵, and IFAP (Internal Fragrant Antisense Primer) 5‵-CATAGGAGCAGCTGAAATATATACC-3‵ (
Bradbury et al. 2005b). The PCR program consisted of one cycle at 95℃ for 5 minutes (denaturation), followed by 30 cycles at 95℃ for 30 seconds (denaturation), 55℃ for 30 seconds (annealing), and 72℃ for 1 minutes (elongation), and was terminated with 1 cycle at 72℃ for 5 minutes. The PCR products were visualized using 1.5% agarose gel electrophoresis and DNA band pattern images were captured using a gel documentation system (Major Science).
Plant height at maturity was determined by measuring the distance from the base of the plant at the soil surface to the apex of the tallest plant organ immediately prior to harvest, with the results expressed in centimeters. Heading dates were recorded for each plant and the number of days from sowing until at least five panicles emerged from the leaf sheath. The total number of tillers per plant was counted during the late vegetative stage, including all tillers, to evaluate the overall tillering capacity. During the late generative stage, the number of productive tillers, those forming panicles, was documented. Panicle length was measured from the base to the apex of each panicle and recorded in centimeters. The grain number per panicle was calculated by enumerating the total number of grains from the selected panicles on each plant, and the average grain number was derived by dividing the total grain count by the number of sampled panicles. The percentage of empty grains was determined by dividing the number of empty grains by the total number of grains in the sampled panicles and multiplying it by 100. The 1000-grain weight was determined by measuring the mass of 1000 grains that had been oven-dried to a constant weight, and the results were expressed in grams. Grain weight per plant was calculated by weighing all fully developed grains harvested from each plant, and the values were recorded in grams. Kernel length and width were measured using a digital caliper.
Statistical analysis
The collected data were analyzed using descriptive statistics, including means, standard deviations, and coefficients of variation (CV). The degree of variability for each trait was categorized as “broad” or “narrow” based on CV values, following the threshold classification described by
Chaniago et al. (2022). The collected data were subjected to an analysis of variance, standard deviation, coefficient of variation, and variability. Specifically, variability was considered broad when the coefficient of variation was greater than or equal to twice the genotypic standard deviation (CVg ≥ 2 σg²), and narrow when it was less than or equal to this threshold (CVg < 2 σg²). This classification facilitates the interpretation of trait stability and helps identify characters with high genetic heterogeneity within the BC
1F
1 population. All data visualizations were generated using Microsoft Excel 2021 MSO.
Results
Molecular Analysis of BC1F1 Plants
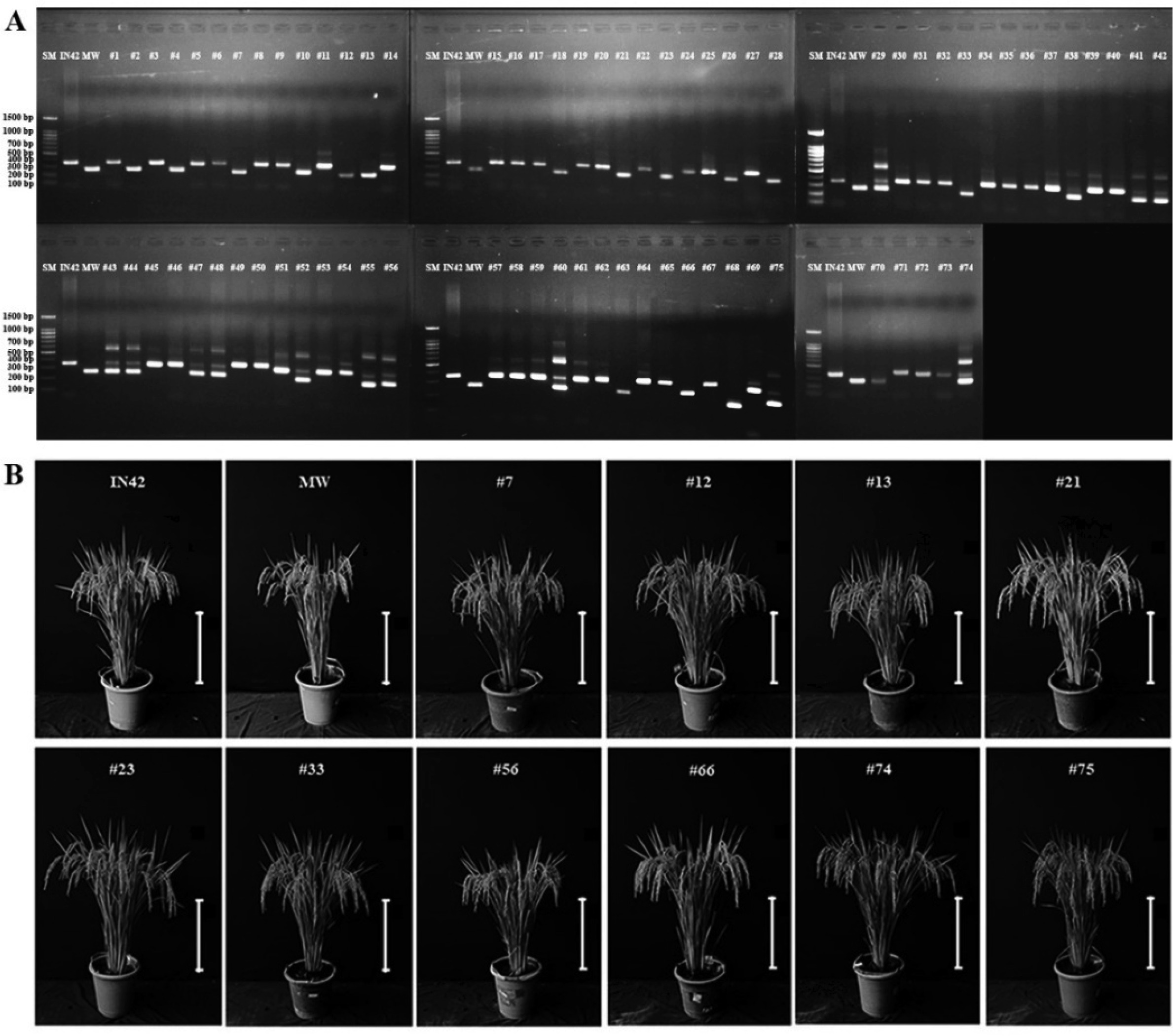
Molecular analysis was conducted using the Bradbury marker to detect the presence of aromatic traits in BC
1F
1 rice plants derived from a backcross between the F1 hybrids of Inpari 42 (IN42) and Merah Wangi (MW) with Inpari 42. This marker specifically targets a deletion mutation in
BADH2, which causes aromatic and non-aromatic rice varieties to produce DNA fragments of different sizes. The analysis revealed that Inpari 42 produced 355 bp DNA fragments, indicating the absence of the mutation in
BADH2, while Merah Wangi produced 257 bp fragments, confirming that a homozygous mutation in BADH2 is responsible for the aromatic trait. In the BC
1F
1 population, 30 out of 75 plants exhibited both 257 bp and 355 bp DNA fragments, indicating a heterozygous mutation in
BADH2 (
Fig. 1A). This result confirmed the successful introgression of the aromatic gene from the F1 generation into BC
1F
1 plants.
To ascertain whether the observed segregation of aromatic traits adhered to Mendelian inheritance patterns, a chi-square analysis was conducted. In accordance with Mendelian genetics, a 1:1 segregation ratio is anticipated in a backcross generation for a single gene trait, wherein half of the progeny should exhibit the aromatic trait (heterozygous; BADH2+/−) and the other half should not (BADH2+/+). The Chi-square test was calculated with 1 deg of freedom (df = 1), and the critical value at the 0.05 significance level was 3.841. The calculated chi-square value of 3.0, which is less than the critical value, indicates that the observed segregation ratio did not deviate significantly from the expected 1:1 Mendelian ratio.
Chi-square analysis confirmed that the segregation of the aromatic trait in the BC
1F
1 population is consistent with Mendelian inheritance. This finding further corroborates the genetic results of the present study, demonstrating that the breeding process effectively maintained the expected genetic ratios. Subsequently, 30 plants containing the heterozygous mutation were selected for further evaluation of agronomic performance. Ten representative BC
1F
1 plants were photographed to illustrate that the BC
1F
1 population exhibited aromatic traits (
Fig. 1B).
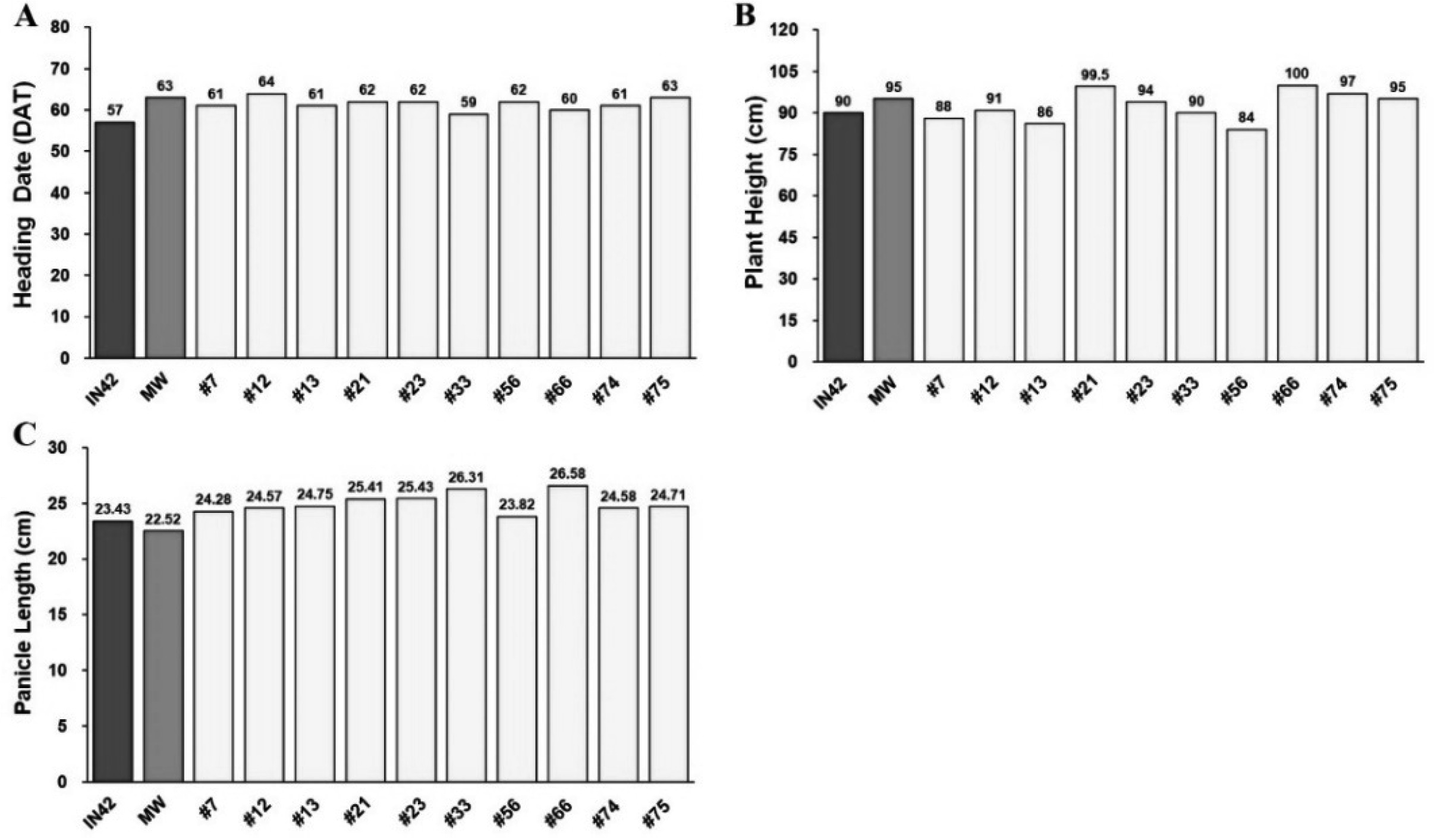
Heading date is a critical trait that influences the adaptability of rice varieties to specific growing conditions. In this study, the mean heading date for BC
1F
1 plants was recorded at 60.33 days after transplanting (DAT), which is intermediate between that of the recurrent parent, Inpari 42 (58.25 DAT), and the aromatic donor parent, Merah Wangi (62.75 DAT) (
Fig. 2A,
Table 1). This intermediate heading date in BC
1F
1 plants suggests the inheritance of flowering traits from both parents, potentially conferring an adaptive advantage in environments in which early or late heading may be beneficial. Furthermore, the low coefficient of variation (4.55%) indicated that heading date is a relatively stable trait within the BC
1F
1 population, which is advantageous for breeding programs aimed at developing varieties with predictable and uniform flowering periods.
Plant height is another critical trait that influences not only the plant’s resistance to lodging, but also its overall growth architecture. The BC
1F
1 plants had an average height of 95.15 cm, which is slightly taller than Inpari 42 (89 cm) and closely matches the height of Merah Wangi (95.50 cm) (
Fig. 2B,
Table 1). This suggests that the height trait from Merah Wangi was effectively introgressed into BC
1F
1 plants, potentially offering enhanced lodging resistance. The relatively narrow variability in plant height (6.19%) within the BC
1F
1 population indicated that this trait is consistently expressed, providing a uniform plant structure that is advantageous for mechanical harvesting and overall crop management.
Panicle length, which directly correlates with the grain number per panicle, is a critical component of yield. The BC
1F
1 plants demonstrated an average panicle length of 24.97 cm, longer than both Inpari 42 (23.10 cm) and Merah Wangi (22.43 cm) (
Fig. 2C,
Table 1). The increased panicle length in BC
1F
1 plants suggests a higher potential for grain production, as longer panicles can support more spikelets, and consequently, more grains. However, the broad variability in panicle length (3.73%) indicated that this trait may benefit from further refinement through selective breeding or additional backcrossing to achieve more uniform panicle lengths across the population.
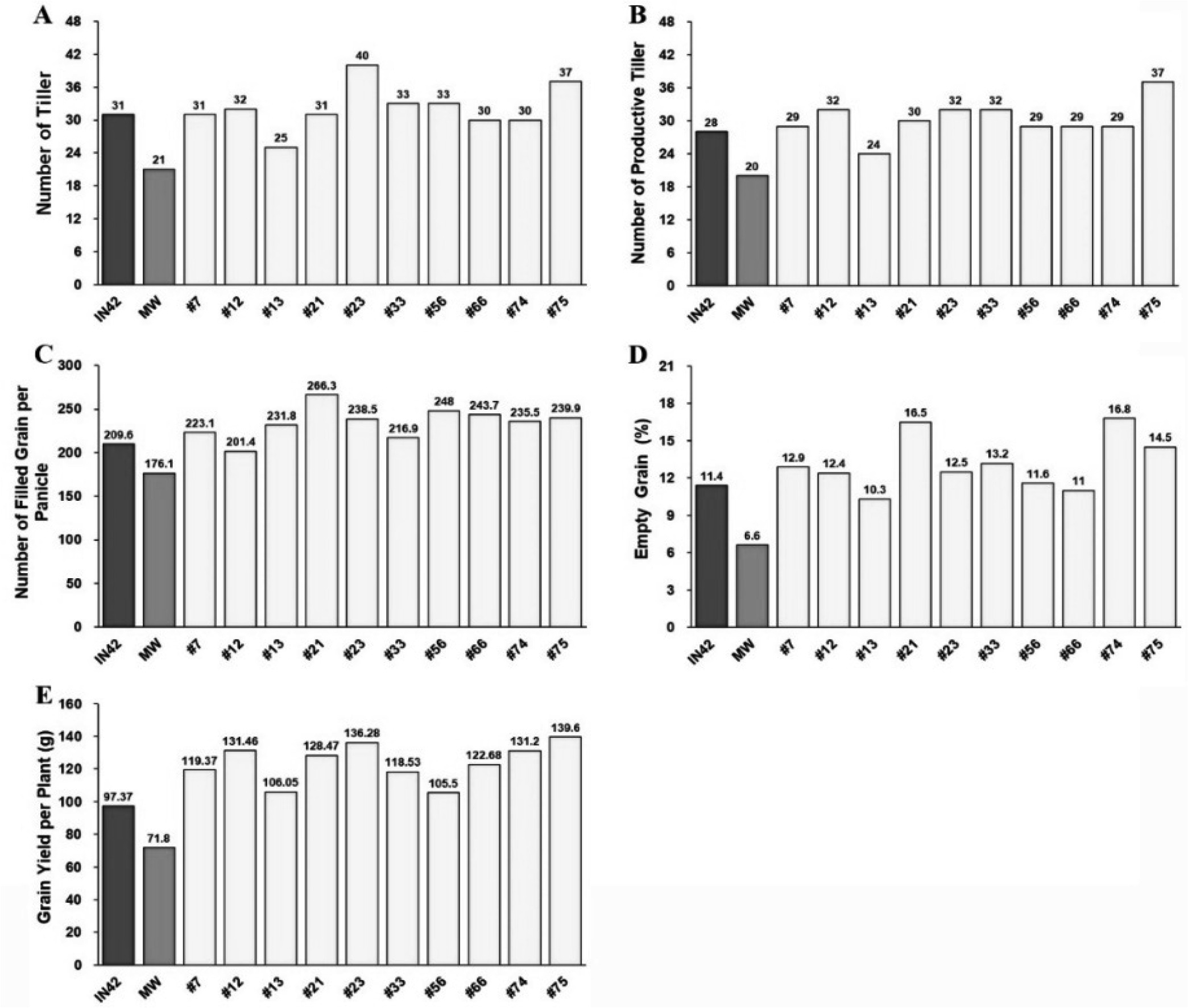
Tillering is a vital trait for rice plants, as each tiller has the potential to produce panicles and contribute to overall grain yield. BC
1F
1 plants demonstrated enhanced tillering capacity, with an average of 32.17 tillers per plant, compared with 31.25 in Inpari 42 and 21 in Merah Wangi (
Fig. 3A,
Table 2). The high tillering capacity observed in BC
1F
1 plants, combined with their broad variability (13.93%), indicated a significant potential for selecting lines with optimal tiller numbers. In rice breeding, the goal is often to balance the number of tillers with the production of productive tillers, which directly contributes to the yield.
Productive tillers, or those that form panicles, are directly related to the yield. BC
1F
1 plants had an average of 30.07 productive tillers per plant, surpassing those of both Inpari 42 (28) and Merah Wangi (20.25) (
Fig. 3B,
Table 2). The higher number of productive tillers in BC
1F
1 plants is a positive indication of their yield potential. The broad variability in this trait (11.11%) suggests that there is room for further selection of lines that not only have high tillering capacity but also convert a high percentage of tillers into productive ones. This is crucial for ensuring that the resources invested in vegetative growth translate into grain production.
The number of filled grains per panicle is a critical yield component because it directly affects the total grain output. BC
1F
1 plants averaged 224.88 filled grains per panicle, which was higher than both Inpari 42 (215.4) and Merah Wangi (171.23) (
Fig. 3C,
Table 2). This increase in the number of filled grains suggests that the BC
1F
1 plants successfully inherited the grain-filling capacity of both parents. However, BC
1F
1 plants also exhibited a higher number of empty grains per panicle, averaging 12.08, compared to Inpari 42 (11.05) and Merah Wangi (6.73) (
Fig. 3D,
Table 2). The broad variability in the number of empty grains per panicle (37.11%) suggests that, while BC
1F
1 plants have a high potential for grain production, there is variability in grain filling, which could be improved through further selection to minimize the number of empty grains.
The grain yield per plant is the most critical trait for evaluating the overall productivity of a rice line. The BC
1F
1 plants exhibited a significantly higher yield, with an average of 118.04 g per plant, compared to 104.95 g in Inpari 42 and 72.79 g in Merah Wangi (
Fig. 3E,
Table 2). The narrow variability in yield per plant (10.48%) indicated that this trait was consistently expressed across the BC
1F
1 population. The higher yield of BC
1F
1 plants can be attributed to the combined effect of increased tillering capacity, higher productive tillers, and a greater number of filled grains per panicle.
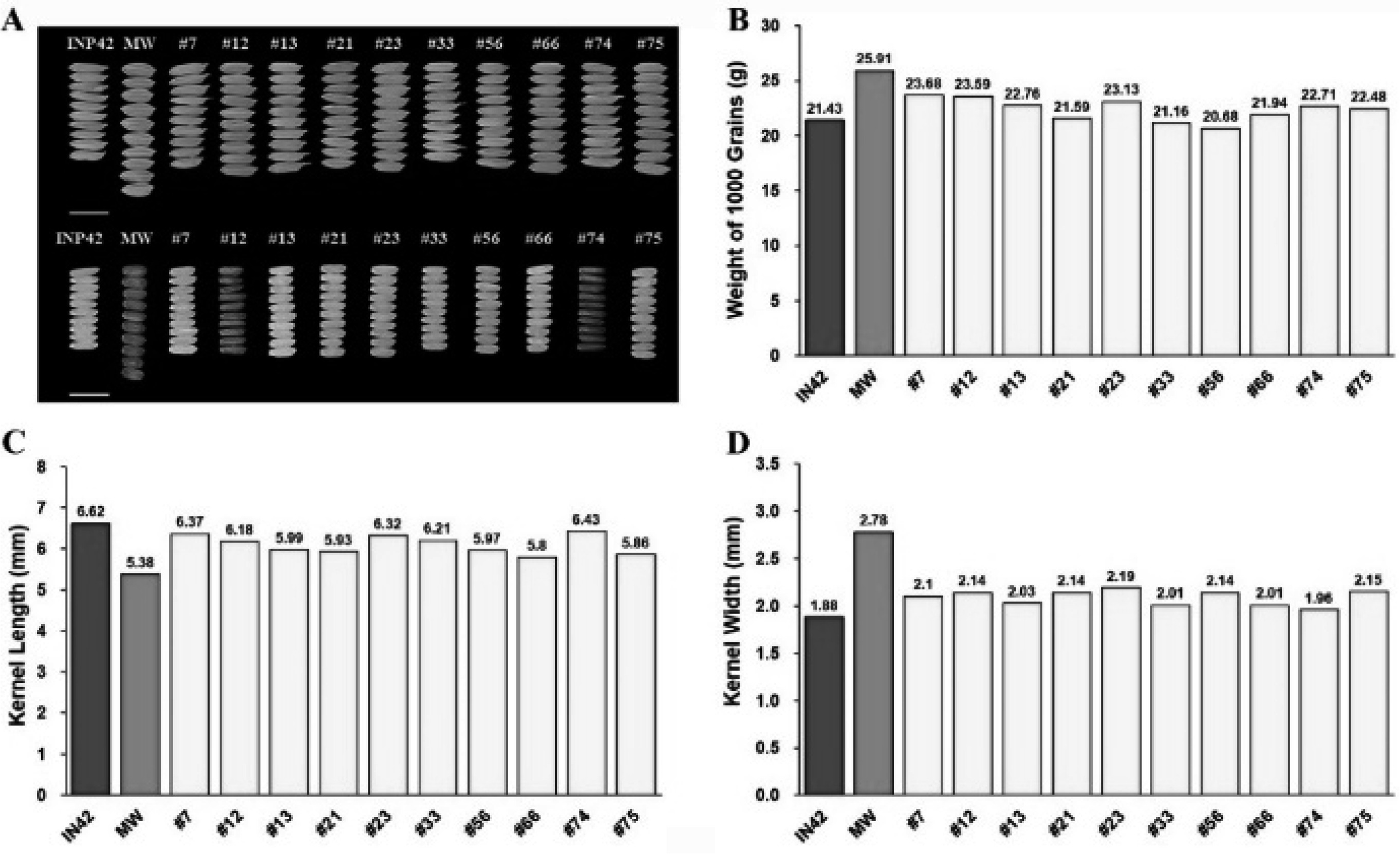
The 1000-grain weight is an important indicator of grain size and density, and is critical for both yield and market value. The BC
1F
1 plants had an average 1000-grain weight of 22.34 g, which was higher than that of Inpari 42 (21.46 g) but lower than that of Merah Wangi (25.84 g) (
Fig. 4B,
Table 3). The broad variability in this trait (4.35%) suggests that there is potential for selecting lines with heavier grains, which could further increase the yield and market appeal of rice. Kernel dimensions are important in determining grain quality, which affects consumer preferences and marketability. The average kernel length in BC
1F
1 plants was 6.13 mm, which was intermediate between that of Merah Wangi (5.37 mm) and Inpari 42 (6.60 mm) (
Fig. 4C,
Table 3). The kernel width in BC
1F
1 plants averaged 2.07 mm, which was wider than that of Inpari 42 (1.86 mm), but narrower than that of Merah Wangi (2.75 mm) (
Fig. 4D,
Table 3). The broad variability in both kernel length (4.26%) and width (4.72%) indicated significant genetic diversity within the BC
1F
1 population. This diversity provides opportunities for further selection to refine the grain size and shape to meet specific market demands.
Discussion
The findings from this study demonstrate the effectiveness of Marker-Assisted Backcrossing (MAB) in integrating aromatic traits into the Inpari 42 rice variety, while simultaneously enhancing key agronomic characteristics in the BC
1F
1 generation. The successful introgression of the aromatic gene from Merah Wangi into the BC
1F
1 plants was confirmed through molecular analysis using the Bradbury marker, which specifically detects a mutation in the
BADH2 gene responsible for fragrance in rice (
Fig. 1A). The resulting 30 BC
1F
1 plants carrying a heterozygous mutation in
BADH2 underscored the reliability of MAB in maintaining aromatic traits across generations. Rice is a recessive trait, meaning that aroma is expressed only when both copies of the
BADH2 gene carry a loss-of-function mutation, leading to the accumulation of 2-acetyl-1-pyrroline, the compound responsible for its characteristic fragrance (
Behera et al. 2023;
Bradbury et al. 2005a). Chi-square analysis confirmed that the observed segregation ratio adheres to Mendelian inheritance, further validating the genetic stability achieved through MAB.
The BC
1F
1 plants had an intermediate heading date of 60.33 days after transplanting, falling between the flowering times of the recurrent parent, Inpari 42, and the aromatic donor parent, Merah Wangi (
Fig. 2A,
Table 1). This intermediate flowering period offers valuable flexibility, enabling these plants to adapt to various growing environments and potentially increasing their suitability across a range of climatic conditions. The low coefficient of variation observed for flowering time within the BC
1F
1 population indicated that this trait is stable, making it a desirable characteristic for breeding programs aimed at developing rice varieties with consistent and predictable heading dates. Heading date is a critical agronomic trait influencing rice adaptability to different latitudes and environmental conditions, directly affecting the length of the growing season and harvest timing, both of which are essential for yield optimization (
Sun et al. 2023). Early maturing rice varieties, which typically flower around 60 days after transplanting, are especially promising for increasing cropping frequency in tropical regions such as Indonesia. These varieties not only enhance yield potential and reduce production risks but also contribute to climate change mitigation by lowering greenhouse gas emissions (
Adhi et al. 2024;
Liang et al. 2024).
The BC
1F
1 plants also exhibited a mean height of 95.15 cm, marginally exceeding that of Inpari 42 and closely approximating Merah Wangi, indicating successful transmission of this trait from the donor parent (
Fig. 2B,
Table 1). This moderate increase in stature coupled with low variability suggests that the BC
1F
1 population demonstrates stable plant height, which is advantageous for breeding programs. Plant height is a crucial trait in rice cultivation because it significantly influences lodging resistance and stem bends or breaks, and root lodging can lead to substantial yield losses and reduced grain quality (
Liu et al. 2017;
Shah et al. 2019; Zhang et al. 2024). The intermediate height of BC
1F
1 plants provides a good balance between yield potential and resistance to lodging, enhancing their suitability for diverse production systems.
The observed variability in tiller number (
Fig. 2A,
Table 2) and panicle length (
Fig. 2C,
Table 1) among BC
1F
1 individuals warrants further discussion. The variability in panicle length may be due to ongoing segregation of quantitative trait loci (QTLs), recombination events disrupting favorable gene linkages, or epistatic interactions between alleles inherited from both parents. This increase in panicle length is significant because longer panicles are positively correlated with grain yield by supporting more spikelets, which leads to an increase in grain number per panicle (
Bai et al. 2016;
Shang et al. 2020). These genetic factors can result in phenotypic inconsistency, especially in early backcross generations where the donor genome is not yet fully diluted. Similarly, the variability in tiller number may reflect transgressive segregation, genotype × environment interaction, or the influence of linked genes from Merah Wangi that affect hormonal regulation and sink–source dynamics (
Li et al. 2023;
Rieseberg et al. 1999;
Won et al. 2022). This heterogeneity is particularly concerning in traits that directly impact yield stability and crop management.
Grain quality traits also exhibited intermediate values in BC
1F
1 plants. The 1000-grain weight was higher than Inpari 42 but lower than Merah Wangi, while kernel length and width reflected a similar intermediate trend (
Figs. 3B–3D,
Table 3). The broad variability observed in 1000-grain weight within the BC
1F
1 population indicates substantial genetic diversity, providing an opportunity for selective breeding to enhance yield, quality, and commercial value (
Li et al. 2018;
Xuedan et al. 2023). Additionally, the kernel length and width in BC
1F
1 plants were intermediate between those of the two parental lines, with a kernel length of 6.13 mm and kernel width of 2.07 mm (
Figs. 3C–3D,
Table 3). The broad variability in both kernel length (4.26%) and width (4.72%) further highlight the genetic diversity within this population, offering scope for breeders to refine grain size and shape, which is essential for improving both marketability and consumer preferences (
Saidon et al. 2020). The broad variation observed in these traits confirms that additional cycles of backcrossing and selection (BC2F1, BC3F1) will be essential to fix desired traits and eliminate undesirable variation. This stepwise breeding strategy is crucial for minimizing genetic drag, enhancing aroma stability, and restoring the elite agronomic profile of Inpari 42 while incorporating the fragrance of Merah Wangi.
To address the observed inconsistencies and variability in key traits, the next phase of this breeding program will involve advancing selected BC
1F
1 lines through successive backcross generations up to BC6F1in accordance with the release requirements set by the Indonesian Ministry of Agriculture for backcross-derived varieties. Each backcross cycle will be accompanied by foreground selection for the mutant
BADH2 allele to ensure the retention of the aromatic trait. By the BC6F1 generation, the resulting lines are expected to possess a theoretical genetic similarity of approximately 99.21% to the recurrent parent, Inpari 42 (
Hasan et al. 2015). Early-generation phenotypic screening will be combined with marker-assisted selection to identify promising lines with stable trait expression. In addition, QTL mapping or genomic selection approaches may be incorporated to enhance the precision and efficiency of trait fixation. This stepwise breeding strategy is essential for minimizing genetic drag, stabilizing both aromatic and agronomic traits, and ultimately developing a uniform, high-yielding aromatic rice variety based on Inpari 42 that meets national standards for varietal release.
Conclussion
This study demonstrated the successful introgression of the mutant BADH2 allele responsible for fragrance from Merah Wangi into the high-yielding genetic background of Inpari 42 through Marker-Assisted Backcrossing. The resulting BC1F1 plants exhibited improved agronomic traits such as enhanced tillering, longer panicles, and higher yield alongside stable expression of key characteristics like heading date and plant height. The main contribution of this research is the establishment of early-generation lines that combine fragrance quality with desirable agronomic performance, forming a basis for developing a new Inpari 42 aromatic variety. However, the study is limited to the BC1F1 generation, which still shows trait variability and lacks multi-location evaluation. Future work will focus on advancing the population to BC6F1 to meet varietal release standards, alongside phenotypic selection, molecular screening, and field trials to stabilize traits and ensure commercial viability.
Acknowledgment
We would like to express our gratitude tothe Indonesia Endowment Fund for Education (LPDP) and the National Research and Innovation Agency (BRIN) for their support in funding this research through the Riset dan Inovasi untuk Indonesia Maju (RIIM) program under research contract number 146/IV/KS/11/2023 and 10875/UN25.3.1/LT/2023. We also extend our appreciation to the Center for Development of Advanced Science and Technology (CDAST) and Agrotechnopark Universitas Jember for their invaluable support in providing the facilities required to conduct this study.
Fig. 1Detection of the Aromatic Trait in BC1F1 Rice Plants using the Bradbury Marker. A. Visualization of the aromatic trait in BC1F1 plants via agarose gel electrophoresis. B. Morphological characteristics of a selected BC1F1 plant exhibiting the aromatic trait. The scale bar represents 50 cm.
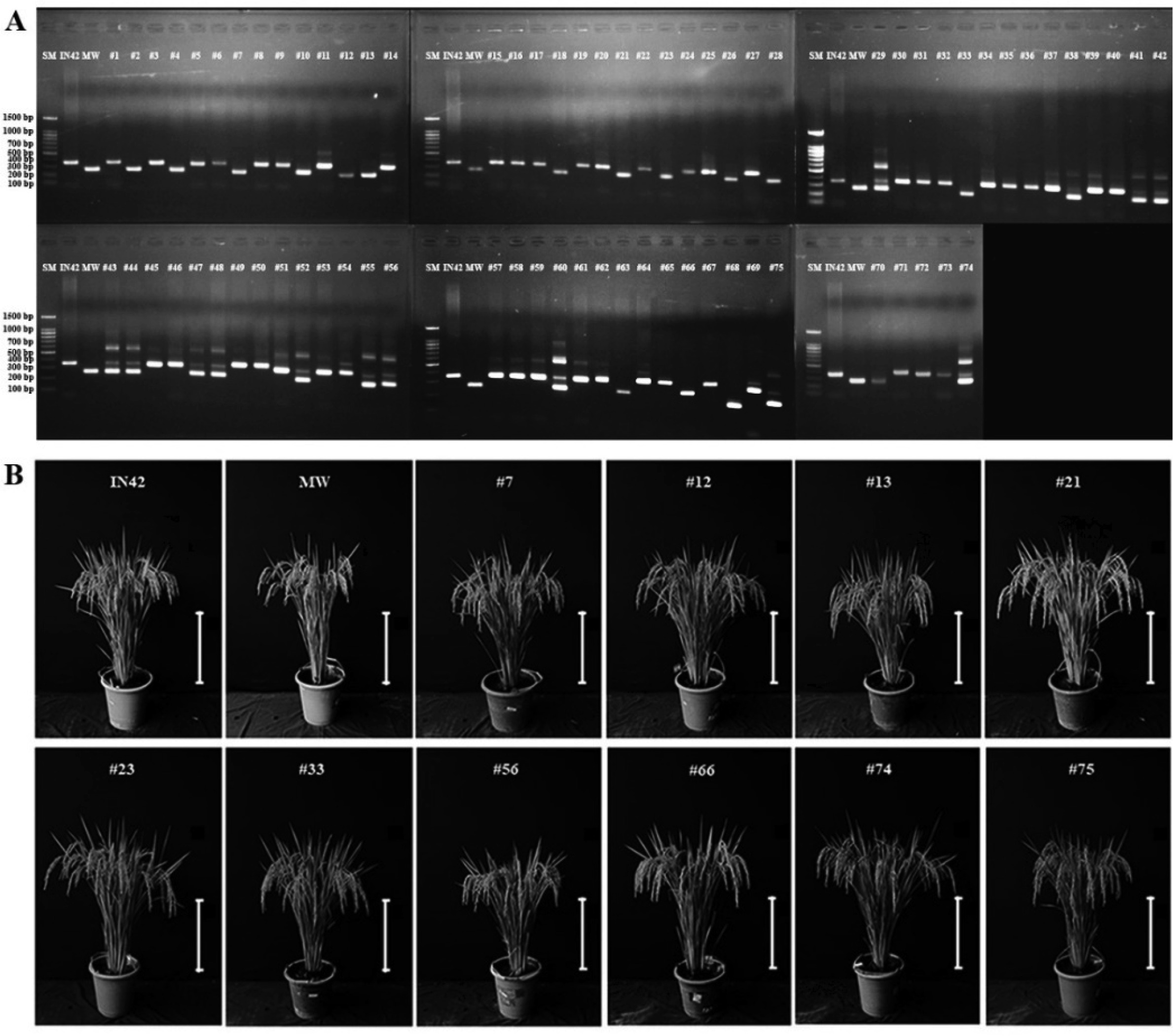
Fig. 2Phenological and Morphological Traits of BC1F1 Rice Plants Compared to Parental Lines. (A) Flowering Time, (B) Plant Height, and (C) Panicle Length.
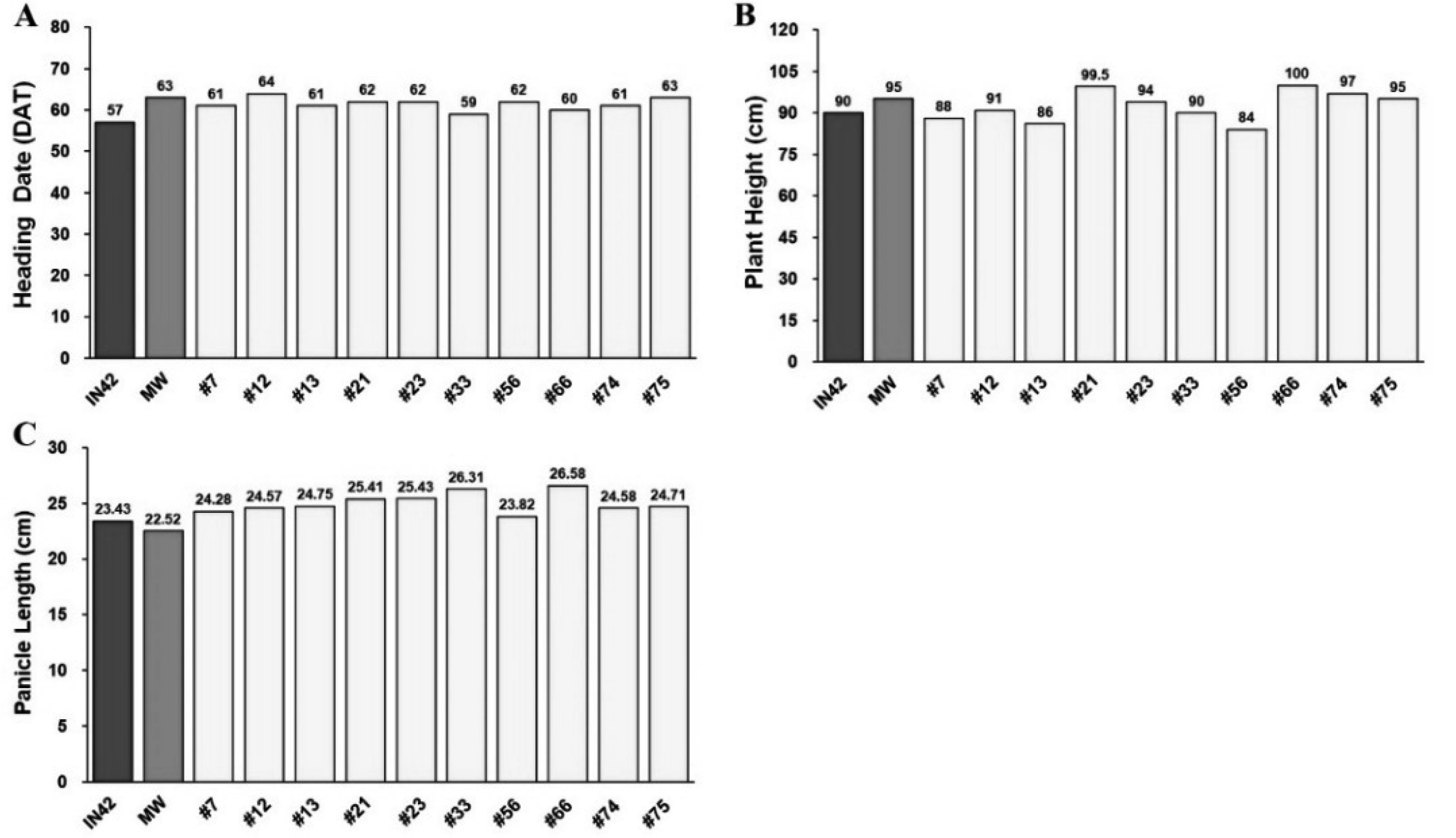
Fig. 3Assessment of Tillering Capacity and Grain Yield Components in BC1F1 Rice Plants Compared to Parental Lines: (A) Total Number of Tillers, (B) Number of Productive Tillers, (C) Number of Filled Grains per Panicle, (D) Percentage of Empty Grains, and (E) Grain Yield per Plant
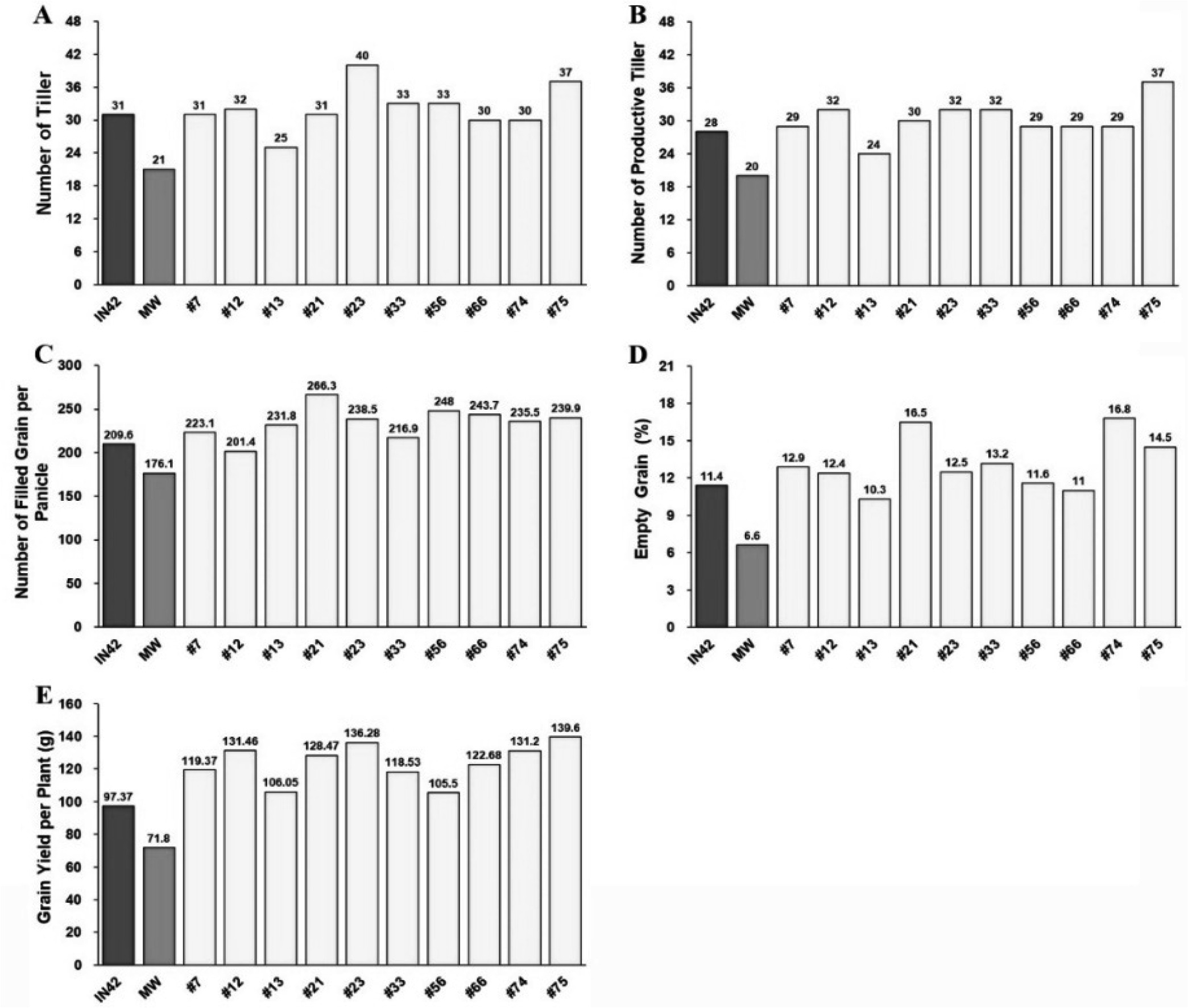
Fig. 4Grain Quality Assessment of BC1F1 Rice Plants Compared to Parental Lines: (A) Grain and Kernel Morphology, with scale bar representing 1 cm, (B) 1000-Grain Weight, (C) Kernel Length, and (D) Kernel Width.
Table 1Comparison of Phenological and Morphological Traits of BC1F1 Rice Plants with Recurrent Parent Inpari 42 and Donor Parent Merah Wangi
Table 1
|
Parameters |
Plant Line |
Sample Number |
Average |
Range |
Standar Deviation |
Coefficient of Variation |
Variability |
|
Heading Date |
MW |
4 |
62.75 |
61–64 |
1.26 |
2.01 |
Narrow |
|
IN42 |
4 |
58.25 |
55–61 |
2.75 |
4.73 |
Narrow |
|
BC1F1 |
30 |
60.33 |
55–67 |
2.75 |
4.55 |
Narrow |
|
Plant Height (cm) |
MW |
4 |
95.50 |
92–100 |
3.32 |
3.47 |
Narrow |
|
Inpari 42 |
4 |
89 |
87–92 |
2.45 |
2.75 |
Narrow |
|
BC1F1 |
30 |
95.15 |
84–107.5 |
5.89 |
6.19 |
Narrow |
|
Panicle Length (cm) |
MW |
4 |
22.43 |
21.0–23.9 |
1.16 |
5.19 |
Broad |
|
Inpari 42 |
4 |
23.10 |
21.6–23.8 |
0.99 |
4.27 |
Broad |
|
BC1F1 |
30 |
24.97 |
23.3–27.4 |
0.93 |
3.73 |
Broad |
Table 2Tillering Capacity and Grain Yield Components of BC1F1 Rice Plants Compared to Inpari 42 and Merah Wangi
Table 2
|
Parameters |
Plant Line |
Sample Number |
Average |
Range |
Standar Deviation |
Coefficient of Variation |
Variability |
|
Tiller Number |
MW |
4 |
21 |
17–25 |
3.27 |
15.55 |
Broad |
|
IN42 |
4 |
31.25 |
22–39 |
7.04 |
22.53 |
Broad |
|
BC1F1
|
30 |
32.17 |
25–42 |
4.48 |
13.93 |
Broad |
|
Productive Tiller |
MW |
4 |
20.25 |
16–24 |
3.30 |
16.32 |
Broad |
|
Number |
IN42 |
4 |
28 |
20–38 |
7.48 |
26.73 |
Broad |
|
BC1F1
|
30 |
30.07 |
24–38 |
3.34 |
11.11 |
Broad |
|
Number of Filled |
MW |
4 |
171.23 |
159–184 |
11.10 |
6.48 |
Narrow |
|
Grain per Panicle |
IN42 |
4 |
215.4 |
187–233 |
21.80 |
10.12 |
Narrow |
|
BC1F1
|
30 |
224.88 |
187–266 |
20.01 |
8.90 |
Narrow |
|
Empty Grain (%) |
MW |
4 |
6.73 |
5.4–8.8 |
1.47 |
21.83 |
Broad |
|
IN42 |
4 |
11.05 |
9.6–11.7 |
0.97 |
8.82 |
Broad |
|
BC1F1
|
30 |
12.08 |
6.0–24.1 |
4.48 |
37.11 |
Broad |
|
Grain Yield per Plant |
Merah Wangi |
4 |
72.79 |
57.0–90.8 |
13.86 |
19.04 |
Narrow |
|
(g) |
IN42 |
4 |
104.95 |
65.7–143.6 |
32.45 |
30.92 |
Narrow |
|
BC1F1
|
30 |
118.04 |
88.9–139.6 |
12.37 |
10.48 |
Narrow |
Table 3Grain Quality Characteristics: 1000-Grain Weight and Kernel Dimensions in BC1F1 Rice Plants Compared to Inpari 42 and Merah Wangi
Table 3
|
Parameters |
Plant Line |
Sample Number |
Average |
Range |
Standar Deviation |
Coefficient of Variation |
Variability |
|
Weight of 1000 seed |
MW |
4 |
25.8 |
24.9–26.9 |
0.83 |
3.22 |
Broad |
|
(g) |
IN42 |
4 |
21.5 |
20.7–22.4 |
0.69 |
3.21 |
Broad |
|
BC1F1 |
30 |
22.3 |
20.7–25.1 |
0.97 |
4.35 |
Broad |
|
Kernel Length (mm) |
MW |
4 |
5.4 |
5.3–5.5 |
0.10 |
1.85 |
Broad |
|
IN42 |
4 |
6.6 |
6.4–6.7 |
0.12 |
1.84 |
Broad |
|
BC1F1 |
30 |
6.1 |
5.8–6.8 |
0.26 |
4.26 |
Broad |
|
Kernel Width (mm) |
MW |
4 |
2.8 |
2.7–2.8 |
0.03 |
0.91 |
Broad |
|
IN42 |
4 |
1.9 |
1.7–1.9 |
0.09 |
4.61 |
Broad |
|
BC1F1 |
30 |
2.1 |
1.8–2.2 |
0.10 |
4.72 |
Broad |
References
- Adhi A, Aryanto G, Kusumaningrum N. 2024. Policy pathway to resilience: Shifting to high-yielding rice seeds to reduce emissions and strengthen rice production in Indonesia. BIO Web Conf..
- Ashokkumar S, Jaganathan D, Ramanathan V, Rahman H, Palaniswamy R, Kambale R, et al. 2020. Creation of novel alleles of fragrance gene OsBADH2 in rice through CRISPR/Cas9 mediated gene editing. PLoS One. 15(8): e0237018
- Bai X, Zhao H, Huang Y, Xie W, Han Z, Zhang B, et al. 2016. Genome-Wide Association Analysis Reveals Different Genetic Control in Panicle Architecture Between Indica and Japonica Rice. T. P. G. 9(2): 1-10.
- Behera PK, Panda D. 2023. Germplasm Resources, Genes and Perspective for Aromatic Rice. Rice Sci. 30(4): 294-305.
- Bradbury LMT, Fitzgerald TL, Henry RJ, Jin Q, Waters DLE. 2005a. The gene for fragrance in rice. P. B. J. 3(3): 363-370.
- Bradbury LMT, Henry RJ, Jin Q, Reinke RF, Waters DLE. 2005b. A perfect marker for fragrance genotyping in rice. M. B. 16(4): 279-283.
- Buttery RG, Ling LC, Juliano BO. 1982. 2-Acetyl-1-pyrroline: An important aroma component of cooked rice. Chem. Ind. 12: 958-959.
- Calingacion M, Laborte A, Nelson A, Resurreccion A, Concepcion JC, Daygon VD, et al. 2014. Diversity of Global Rice Markets and the Science Required for Consumer-Targeted Rice Breeding. PLOS One. 9(1): e85106
- Chaniago N, Suliansyah I, Chaniago I, Rozen N. 2022. Morphological characteristics of local rice in Deli Serdang District, North Sumatra, Indonesia. Biodiv. 23(2): 883-894.
- Chen S, Yang Y, Shi W, Ji Q, He F, Zhang Z, et al. 2008. Badh2, Encoding Betaine Aldehyde Dehydrogenase, Inhibits the Biosynthesis of 2-Acetyl-1-Pyrroline, a Major Component in Rice Fragrance. Plant Cell. 20(7): 1850-1861.
- Fanata WID, Husna SF. 2021. The Determination of Aromatic Character of Several Local Rice Varieties using Phenotypic Analysis and Molecular DNA. J. I. D. 22(2): 111-118.
- Hasan MM, Rafii MY, Ismail MR, Mahmood M, Rahim HA, Alam MA, et al. 2015. Marker-assisted backcrossing: a useful method for rice improvement. Biotechnol. Biotechnol. Equip. 29(2): 237-254.
- Imran M, Shafiq S, Tang X. 2023b. CRISPR-Cas9-mediated editing of BADH2 gene triggered fragrance revolution in rice. Physiol. Plant. 175(1): e13871
- Imran M, Shafiq S, Ashraf U, Qi J, Mo Z, Tang X. 2023a. Biosynthesis of 2-Acetyl-1-pyrroline in Fragrant Rice: Recent Insights into Agro-management, Environmental Factors, and Functional Genomics. J. Agric. Food Chem. 71(10): 4201-4215.
- Ishak I, Fedora F. 2024. Selection of rice aroma from crossing aromatic rice x T250.7 mutant genotype using DNA marker. Biodiv. 25(5): 2191-2197.
- Kumar M, Singh RP, Jena D, Singh V, Rout D, Arsode PB, et al. 2023. Marker-Assisted Improvement for Durable Bacterial Blight Resistance in Aromatic Rice Cultivar HUR 917 Popular in Eastern Parts of India. Plants. 12(6): 1363
- Kurniawati YT, Febrianti WN, Novidiarsih CI, Jumantoro G, Mukhtar K. 2022. Loss of Rice Yields Due to Rice Ragged Stunt Virus (RRSV) on Several Varieties (Inpari 32, Inpari 42, Inpari 16, and Ciherang) in Madiun. at S.N.M.A. 2022. Surabaya Presented..
- Li N, Xu R, Duan P, Li Y. 2018. Control of grain size in rice. Plant Reprod. 31(3): 237-251.
- Li X, Zhou Y, Shuai P, Wang X, Peng S, Wang F. 2023. Source-Sink Balance Optimization Depends on Soil Nitrogen Condition So as to Increase Rice Yield and N Use Efficiency. Agron. 13(3): 907
- Liang Z, Ruiz-Menjivar J, Zhang L, Zhang J, Shen X. 2024. Examining the effects of adopting early maturing crop varieties on agricultural productivity, climate change adaptation, and mitigation. I. J. L. C. T. 19: 1256-1274.
- Liu F, Wang P, Zhang X, Li X, Yan X, Fu D, et al. 2017. The genetic and molecular basis of crop height based on a rice model. Planta. 247(1): 1-26.
- Okpala NE, Mo Z, Duan M, Tang X. 2019. The genetics and biosynthesis of 2-acetyl-1-pyrroline in fragrant rice. Plant Physiol. Biochem. 135: 272-276.
- Ramadhan GR, Fanata WID. Usmadi2020. Effect of Potassium Fertilization on Growth and Head Rice Yield of rice (Oryza sativa L.) Merah Wangi Variety. J. I. D. 21(1): 61-66.
- Rieseberg LH, Archer MA, Wayne RK. 1999. Transgressive segregation, adaptation and speciation. Hered. 83(4): 363-372.
- Sagar V, Dhawan G, Gopala Krishnan S, Vinod KK, Ellur RK, Mondal KK, et al. 2020. Marker assisted introgression of genes governing resistance to bacterial blight and blast diseases into an elite Basmati rice variety, 'Pusa Basmati 1509'. Euphytica. 216(1): 1-18.
- Saidon SA, Kamaruzaman R, Razak MSFA, Ramli A, Sarif HM, Zuki ZM, et al. 2020. Studies on heritability and genetic variability for grain physical properties in Malaysian rice germplasm. IOP Conf. Ser.: E. E. S. 482(1): 012022
- Shah L, Yahya M, Shah SMA, Nadeem M, Ali A, Ali A, et al. 2019. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. Int. J. Mol. Sci. 20(17): 4211
- Shang F, Chen L, Meng X, Yang K, Wang J. 2020. Fine mapping and grain yield analysis of a major QTL controlling primary branch number in rice (Oryza sativa L.). Genet. Resour. Crop. Evol. 67(2): 421-431.
- Singh A, Singh VK, Singh SP, Pandian RTP, Ellur RK, Singh D, et al. 2012. Molecular breeding for the development of multiple disease resistance in Basmati rice. A. o. B. P. 2012: pls029
- Sun K, Zong W, Xiao D, Wu Z, Guo X, Li F, et al. 2023. Effects of the core heading date genes Hd1, Ghd7, DTH8, and PRR37 on yield-related traits in rice. Theor. Appl. Genet. 136(11): 1-14.
- Vanavichit A, Kamolsukyeunyong W, Siangliw M, Siangliw JL, Traprab S, Ruengphayak S, et al. 2018. Thai Hom Mali Rice: Origin and Breeding for Subsistence Rainfed Lowland Rice System. Rice. 11(1): 1-12.
- Wan J, Nakazaki T, Kawaura K, Ikehashi H. 1997. Identification of Marker Loci for Seed Dormancy in Rice (Oryza sativa L.). Crop Sci. 37(6): 1759-1763.
- Wang Y, Tang S, Guo N, An R, Ren Z, Hu S, et al. 2023. Pyramiding Rice Blast Resistance Gene Pi2 and Fragrance Gene badh2. Agron. 13(2): 589
- Won PLP, Kanno N, Banayo NPM, Bueno CS, Sta. Cruz P, Kato Y. 2022. Source-sink relationships in short-duration and hybrid rice cultivars in tropical Asia. Field Crops Res. 282: 108485.
- Xuedan L, Fan L, Yunhua X, Feng W, Guilian Z, Huabing D, et al. 2023. Grain Shape Genes: Shaping the Future of Rice Breeding. Rice Sci. 30(5): 379-404.