Abstract
Cotton is a globally important fiber crop, but many elite cultivars are recalcitrant to in vitro regeneration. We developed a callus-mediated plant regeneration protocol for the local G. barbadense L. cultivar Surkhan-106. Sterilized seeds were germinated in vitro, and hypocotyl and cotyledon explants were cultured on Murashige and Skoog (MS) medium with various auxin/cytokinin combinations. Robust callus induction was achieved from hypocotyl explants within 2–3 weeks. Somatic embryogenesis was induced from callus after approximately 12–14 weeks on medium supplemented with indole-3-butyric acid (IBA), 2,4-dichlorophenoxyacetic acid (2,4-D), kinetin, and 6-benzylaminopurine (BAP). Embryogenic callus was periodically subcultured to prevent browning, and torpedo-stage somatic embryos were matured on hormone-free medium. These embryos germinated into plantlets within 10–15 days on MS medium. The entire regeneration cycle, from seed to plantlet, was completed in about 4–5 months. Hypocotyl explants showed significantly higher callus formation frequency (85%) and regeneration efficiency than cotyledon explants (60%) (ANOVA, p < 0.05). This study establishes the first efficient callus-based regeneration system for Surkhan-106, an important Uzbek cotton variety. The optimized protocol can facilitate rapid clonal propagation and genetic transformation of Surkhan-106 and other recalcitrant cotton cultivars, accelerating breeding programs for improved stress tolerance and yield.
-
Key words: Cotton; Gossypium barbadense, Somatic embryogenesis, Callus induction, Tissue culture, Plant regeneration, Plant growth regulators
Introduction
Cotton (
Gossypium spp.) is one of the world’s most economically important fiber crops, widely cultivated for its valuable fiber, oil, and protein-rich seed (
Abdurakhmonov et al. 2014). Globally, cotton is grown across approximately 35 million hectares, providing livelihood to millions of people and significantly contributing to the economies of cotton-producing countries (
Fang et al. 2017). Uzbekistan historically has been among the world’s leading cotton producers, with cotton remaining a vital component of its agricultural economy (
Babadjanov et al. 2025).
Despite its importance, cotton cultivation faces numerous biotic and abiotic stresses that severely limit yield and fiber quality (
Saud et al. 2022). Traditional breeding has significantly contributed to enhancing these traits; however, conventional methods are generally slow, labor-intensive, and limited by the increasingly narrow genetic base of modern cultivars (
Shahzad et al. 2022). Consequently, breeders are increasingly integrating advanced biotechnological methods, such as genetic transformation and genome editing, into cotton breeding programs to accelerate the development of improved varieties (
Zhang et al. 2024).
Somatic embryogenesis (SE) is a fundamental plant tissue culture technique that facilitates rapid clonal propagation and serves as a critical tool for genetic transformation studies (
Sakhanokho et al. 2016;
Wilkins et al. 2004). SE enables the regeneration of genetically uniform plants from non-reproductive tissues, bypassing limitations associated with sexual reproduction (
Ramírez-Mosqueda et al. 2025;
Trolinder et al. 1987). Despite these advantages, cotton is known to be recalcitrant to regeneration through somatic embryogenesis due to strong genotype dependency, high rates of somaclonal variation, and specific nutritional and hormonal requirements (
Guo et al. 2020;
Jin et al. 2005).
Various studies have identified the critical role of explant source and culture media composition in influencing cotton somatic embryogenesis. Hypocotyl explants, in particular, have consistently demonstrated higher embryogenic potential compared to other tissues such as cotyledons and leaves (
Kumar et al. 2013a;
Trolinder et al. 1988;
Wilkins et al. 2004). The hormonal balance between auxins and cytokinins, particularly the use of 2,4-dichlorophenoxyacetic acid (2,4-D) combined with kinetin or benzylaminopurine (BAP), has been reported to significantly enhance callus induction and embryo formation (
Davidonis et al. 1983;
Kumar et al. 2004;
Trolinder et al. 1987).
Moreover, issues such as callus browning due to phenolic oxidation often hinder the regeneration efficiency in cotton (
Chaudhary et al. 2015). Frequent subculturing and careful medium management have been suggested as effective strategies to mitigate these problems, thus maintaining callus viability and embryogenic capacity (
Ahsan et al. 2014;
Cheng et al. 1995).
Recent advancements, including refined culture media, optimized hormone combinations, and improved sterilization protocols, have facilitated regeneration in various recalcitrant cotton genotypes (
Pathi et al. 2013;
Sakhanokho et al. 2016). Nonetheless, efficient regeneration systems for numerous commercially significant local varieties remain underdeveloped or entirely lacking, underscoring the need for genotype-specific optimization (
Zhang et al. 2024).
Somatic embryogenesis (SE), the formation of embryos from somatic (non-reproductive) cells, is a fundamental technique in plant biotechnology. In cotton, SE allows for rapid clonal propagation and provides a route to recover fertile transgenic plants after in vitro gene transfer or mutagenesis. Despite its potential, cotton is considered a recalcitrant species for tissue culture: regeneration via somatic embryogenesis is complex, time-consuming, and highly genotype-dependent. Several factors make cotton SE challenging, including (1) strong genotype dependence, (2) somaclonal variation among regenerants, (3) limited understanding of the genetic and molecular control of embryogenesis, and (4) genotype-specific requirements for culture media (
Kumar et al. 2013b). Consequently, many cotton varieties do not readily respond to standard regeneration protocols.
Notably, most successful cotton regeneration reports have been restricted to a few responsive genotypes. The American upland cotton cultivar Coker-312 (and related Coker lines) has been widely used as a model for developing regeneration and transformation protocols, with numerous reports of plant regeneration via somatic embryogenesis (
Kumar et al. 2013b). In contrast, many elite commercial cultivars (especially outside the Coker lineage) show low or no regenerative response, limiting the broad application of cotton biotechnology in breeding programs (
Kumar et al. 2013a). There have been some successes with other genotypes: for instance, Chinese cultivar Simian-3 was shown to undergo high-frequency somatic embryogenesis, as were Turkish cultivars Nazilli 143 and Nazilli M-503. Several Indian cotton cultivars (e.g., MCU-5, Khandwa-2, CEMB-66) have also demonstrated moderate embryogenic potential (
Kumar et al. 2013b). Nonetheless, the regeneration efficiencies in these non-Coker varieties are often much lower, and protocols may need substantial genotype-specific optimization (
Kumar et al. 2013a). Recalcitrance to regeneration remains a major hurdle in cotton improvement efforts (
Shazia Yasin et al. 2018).
Uzbekistan’s local cotton varieties are important for regional agriculture but have not been extensively studied for in vitro culture responses. Surkhan-106 is an agronomically valuable G. barbadense cultivar in Uzbekistan, prized for its yield and fiber quality under local growing conditions. However, like many local cultivars, it has been recalcitrant to plant regeneration in vitro, which impedes the application of advanced breeding techniques (e.g., gene editing or transformation) to improve this cultivar. Developing an efficient regeneration system for Surkhan-106 would therefore significantly enhance the toolkit for cotton breeders, enabling faster incorporation of novel traits such as stress resistance and improved fiber quality.
Materials and Methods
Plant Material and Explant Preparation
Seeds of the local upland cotton (Gossypium barbadense L.) cultivar Surkhan-106 were used as the starting plant material. The seeds were obtained from the cotton germplasm collection of the Cotton Breeding, Seed Production and Agrotechnologies research Institute. To remove the linters (seed fuzz) and facilitate sterilization, seeds were first treated with concentrated H₂SO₄ for 2–3 minutes, then thoroughly rinsed with water to eliminate all acid and fiber residues.
For surface sterilization, we adopted an efficient and cost-effective protocol modified from (
Bakhsh et al. 2016). The sterilization steps were carried out under aseptic conditions as follows:
Detergent wash: Soak seeds in a lab detergent (liquid soap) solution and agitate for 10 minutes to remove debris and microbial contaminants.
Water rinse: Rinse seeds under running tap water for 5 minutes to wash off all soap residues.
Ethanol disinfection: Immerse seeds in 70% ethanol for 1 minute, then decant. (This brief alcohol treatment sterilizes the seed surface.)
Sterile water rinse: Rinse seeds five times with sterile distilled water to remove any remaining ethanol.
Hydrogen peroxide treatment: Soak seeds in 15% H₂O₂ (hydrogen peroxide) for 10 minutes. This strong oxidizing agent kills persistent spores and bacteria.
Sterile water rinse: Rinse the seeds five times with sterile distilled water.
Bleach treatment: Immerse seeds in 20% (v/v) sodium hypochlorite (commercial bleach) for 5 minutes for further disinfection.
Final rinses: Rinse seeds at least five times with sterile distilled water to remove all traces of bleach.
After the final rinse, the sterilized seeds were aseptically transferred to Petri dishes containing germination medium. The germination medium consisted of half-strength Murashige and Skoog (MS) basal salts (
Kumar et al. 2013b), 3% (w/v) sucrose, and 0.7% (w/v) agar, with the pH adjusted to 5.8 prior to autoclaving. Petri dishes were incubated in a growth room at 28 ± 2℃ in the dark for the first 2 days to facilitate imbibition, then moved to a lighted condition (16 h photoperiod at ~60 µmol provided by cool-white fluorescent lamps) to promote seed germination. Under these conditions, Surkhan-106 seeds began germinating within 3–5 days.
When the seedlings reached ~10 days old (with well-developed hypocotyls and emergent cotyledons), explants were prepared for callus induction. Hypocotyl segments (~0.5–1.0 cm length) and cotyledon segments (~0.5 cm² pieces) were excised from the sterile seedlings. The explants were taken from similar positions for each seedling to maintain consistency: hypocotyl segments were cut from the mid-hypocotyl region, and cotyledon pieces were taken from the central area of each cotyledon. All cuts were made with a sterile scalpel. Explants were immediately transferred onto callus induction medium with the cut surface in contact with the medium. At least 20 explants of each type (hypocotyl and cotyledon) were cultured for each experimental replicate.
Culture Media and Conditions
We optimized several culture media for the sequential stages of cotton callus culture and plant regeneration, drawing on protocols reported for other cotton genotypes (
Kumar et al. 2013b). All media were based on MS basal salts supplemented with Gamborg’s B5 vitamins, 3% carbon source, and 0.7% agar. The compositions of each medium and their intended use are as follows:
Callus Induction Medium (CIM): MS salts (full strength) + B5 vitamins; 15 g/L maltose + 15 g/L glucose; 0.05 mg/L indole-3-butyric acid (IBA); 1.0 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D); 0.5 mg/L 6-benzylaminopurine (BAP); 0.5 mg/L kinetin; 7 g/L agar; pH 5.8. This medium (analogous to SSh-1 in the original protocol) was used to induce callus from the explants. The combination of a high auxin (2,4-D) with low concentrations of IBA and cytokinins was chosen to promote vigorous callus proliferation.
Embryo Induction Medium (EIM): MS + B5; 15 g/L maltose + 15 g/L glucose; 0.1 mg/L IBA; 1.0 mg/L 2,4-D; 0.5 mg/L BAP; 2.0 mg/L kinetin; 7 g/L agar; pH 5.8. (Corresponding to SSh-2.) This medium contains a slightly higher cytokinin (kinetin) level to stimulate the differentiation of embryogenic structures while still maintaining auxin presence. It supports the transition of proliferating callus to somatic embryogenesis.
Embryo Maturation Medium (EMM): MS + B5; 15 g/L maltose + 15 g/L glucose; 5 mg/L α-naphthaleneacetic acid (NAA); 0.1 mg/L BAP; 7 g/L agar; pH 5.8. (Corresponding to SSh-3.) In this medium, a moderate concentration of NAA (a less potent auxin) and a low level of cytokinin help maturing somatic embryos develop. The relatively high auxin may assist in root meristem formation during embryo maturation. This medium lacks 2,4-D and kinetin, as high auxin/cytokinin levels are no longer needed once embryos are formed.
Germination Medium (GM): MS + B5; 15 g/L maltose + 15 g/L glucose; no hormones; 7 g/L agar; pH 5.8. (Corresponding to SSh-4.) Hormone-free MS medium facilitates the conversion of mature somatic embryos into plantlets (shoot and root development) without further callus proliferation. Additionally, following Davidonis & Hamilton’s findings, the total inorganic nitrogen content was optimized by adding extra KNO₃ and adjusting NH₄⁺ levels in this medium to support late-stage embryo development. These modifications (higher nitrate availability) have been reported to be critical for maturation and germination of cotton somatic embryos.
All media were autoclaved at 121℃ for 20 minutes. Vitamins and growth regulators were filter-sterilized and added to cooled media to avoid degradation. The pH of each medium was adjusted to 5.8 before gelling. Cultures were incubated in a controlled growth room at 28 ± 2℃. For callus induction and somatic embryo induction stages, explants and calli were kept under a 16 hours light/8 hours dark photoperiod (cool-white fluorescent lights). However, during the initial 10 days of somatic embryo germination on GM, plates were kept in darkness to improve root emergence, after which they were returned to light to encourage chlorophyll development in the new plantlets.
Explants and callus cultures were subcultured regularly to fresh media to ensure a continuous supply of nutrients and hormones. Specifically, callus cultures were transferred to fresh EIM or EMM every 3 weeks. Frequent subculturing also helped to minimize browning of callus tissue by discarding necrotic regions and providing fresh medium to dilute accumulated phenolic compounds. Cultures were observed weekly, and any contamination was recorded; contaminated cultures were discarded immediately.
Experimental Design and Data Analysis
Each treatment was conducted in three independent biological experiments, performed on different days using newly germinated seed batches and freshly prepared media. In each replicate, at least 20 explants of each type (hypocotyl and cotyledon) were cultured, ensuring reproducibility and reliability of the results. In total, over 100 explants of each type (hypocotyl, cotyledon) were assessed for callus formation. Callus induction frequency was calculated as the percentage of explants that produced callus tissue after 4 weeks. For somatic embryogenesis, we recorded the percentage of callus cultures that formed visible somatic embryos after 12–14 weeks on EIM/EMM, and the number of embryos per callus in each responding culture.
Data were analyzed using GraphPad Prism 9 (GraphPad Software, USA). One-way analysis of variance (ANOVA) was used to compare means of quantitative data (e.g. callus induction rates) between explant types. Significance was evaluated at the 5% level, and post hoc Tukey tests were applied if applicable. The callus induction frequencies for hypocotyl vs. cotyledon explants were compared using a two-sample t-test (as there were two groups), which was equivalent to a one-way ANOVA with 1 df. Results are reported as mean ± standard error (SE). We also documented qualitative observations (e.g., callus texture, color changes, time to embryo appearance).
Results
Callus Induction and Explant Response
Sterilized seeds of Surkhan-106 germinated readily on half-strength MS medium, producing healthy seedlings suitable for explanting. Hypocotyl and cotyledon explants from these seedlings showed markedly different responses in terms of callus induction. Within 15–20 days of culture on Callus Induction Medium (CIM), a majority of hypocotyl explants began to swell and develop callus at the cut edges. In contrast, cotyledon explants showed slower and less prolific callusing, with some explants only producing small nodular callus or none at all by the 3-week mark. Initially, the newly formed callus on hypocotyl segments was white, semi-translucent, and friable (soft). As the callus grew, its color gradually shifted: patches of the callus became cream or pale green after ~4 weeks. By 6–8 weeks, actively growing callus from hypocotyls often had a yellow-green to light green hue. Cotyledon-derived calli, when they formed, tended to remain smaller and sometimes developed a brownish color at the edges, indicating slower growth or onset of browning.
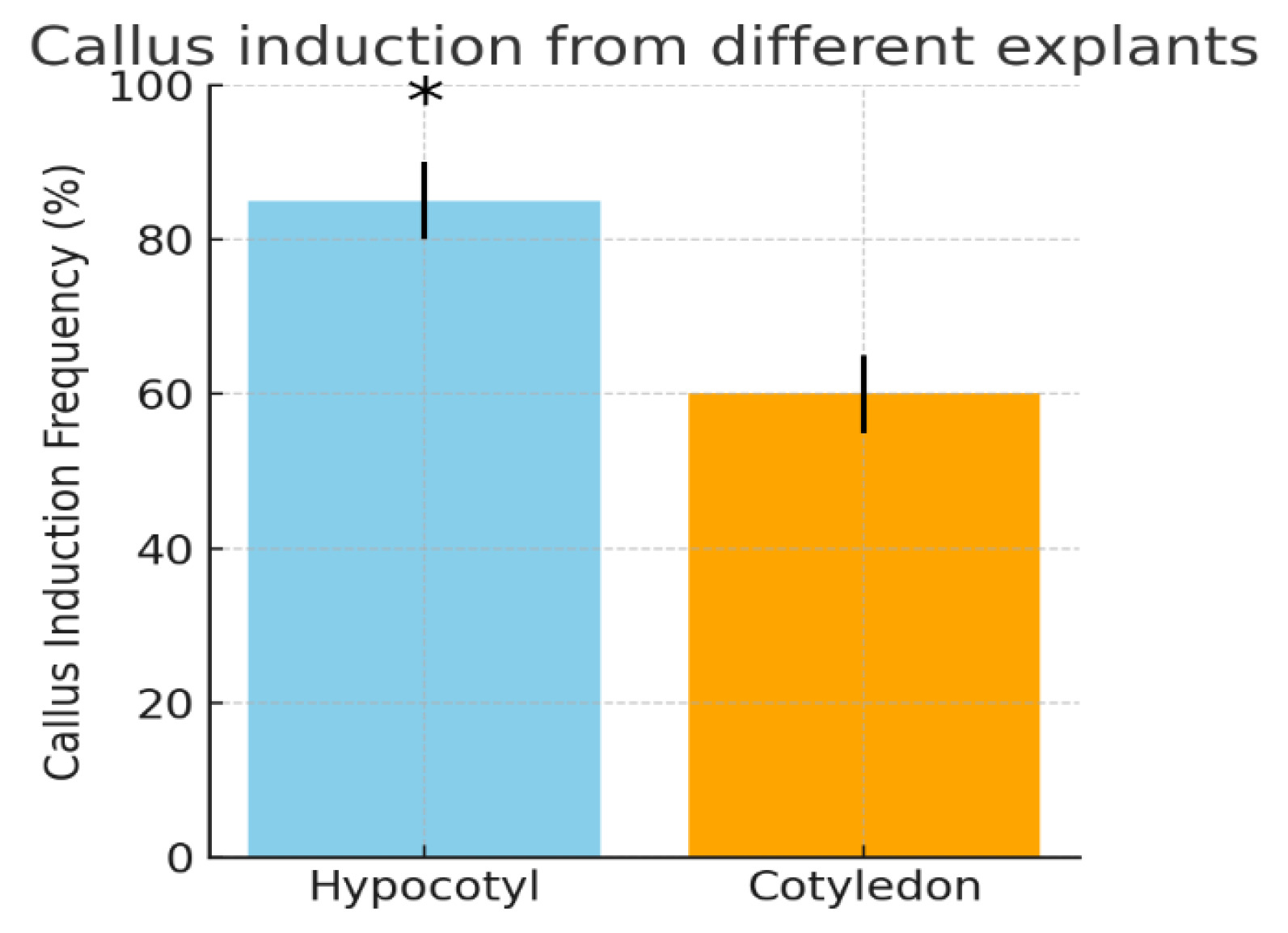
Importantly, hypocotyl explants yielded a significantly higher frequency of callus formation compared to cotyledon explants (
Fig. 1). Quantitatively, about 80–90% of hypocotyl segments produced callus, whereas only ~50–60% of cotyledon pieces did so. Statistical analysis confirmed the superiority of the hypocotyl as an explant: the callus induction rate from hypocotyls was significantly greater (
p < 0.05) than that from cotyledons. This finding aligns with previous reports that the choice of explant tissue affects cotton callus culture success. For example,
Wilkins et al. (2004) noted that hypocotyl sections of cotton seedlings were more responsive to callus induction than cotyledon or leaf tissues. Hypocotyls may contain more meristematic cells or a more favorable endogenous hormone balance for dedifferentiation (
Ahsan et al. 2014), explaining their better performance. Additionally, hypocotyl segments have a larger wounding surface area (from both cut ends) which can stimulate callus via wound-induced hormones. Cotyledons, being more differentiated and containing tannins, often exhibit lower totipotency and can brown more quickly.
All callus cultures (from both explant types) were maintained by regular subculturing onto fresh CIM to encourage further proliferation. Through continuous subculture every 2–3 weeks, we obtained a substantial amount of Surkhan-106 callus. We observed that hypocotyl-derived callus remained generally more embryogenic-looking (light green and actively growing), whereas cotyledon-derived callus, when present, was often prone to faster browning and necrosis. Because of the much higher yield and quality of callus from hypocotyls, subsequent experiments on somatic embryogenesis were focused primarily on callus derived from hypocotyl explants.
Somatic Embryogenesis and Callus Proliferation
After 8 weeks of initial growth on CIM, the actively proliferating callus masses (especially those from hypocotyls) were transferred to the Embryo Induction Medium (EIM) to stimulate somatic embryo formation. The callus at this stage was chunky and nodular, with a mixture of creamy-white and green sectors. Upon transfer to EIM (which had a slightly higher cytokinin:auxin ratio), subtle morphological changes were noted in the following weeks. By around 12–14 weeks from the start of culture (i.e., roughly 4–6 weeks on EIM), some callus pieces began showing structures characteristic of somatic embryogenesis. Small, globular to heart-shaped green structures became visible on the surface of the callus; these were the developing somatic embryos. We define an embryogenic callus as one that produces somatic embryos (embryoid structures) rather than just undifferentiated cells. In Surkhan-106, only a portion of the callus tissue turned embryogenic – typically, green, compact sectors of callus gave rise to embryoids, while other parts remained non-embryogenic.
Notably, the color and texture of the callus were indicative of its embryogenic potential. Embryogenic callus regions were usually firm and green, sometimes with a meristematic, organogenic appearance. Non-embryogenic portions were looser, yellowish, or began to brown. We observed that the initial embryogenic nodules were green with a smooth surface; over time, these developed into recognizable somatic embryo stages (globular, heart, torpedo shapes). The primary somatic embryos appeared after ~12 weeks in culture as green protuberances. With continued incubation (up to 16 weeks), some embryos reached the torpedo stage and became bipolar (distinguishable shoot and root poles).
During this period, a challenge encountered was callus browning. Some callus, especially older portions, turned brown due to the oxidation of phenolic compounds – a common issue in cotton cultures. Browning can be detrimental as it is often associated with tissue necrosis and loss of regeneration potential. In our cultures, browning was particularly observed in callus that had not been subcultured promptly or in regions where growth had slowed. We addressed this by implementing a strict subculture regime: every 3 weeks, all callus cultures were transferred to fresh medium, and any brown, necrotic tissue was carefully trimmed away prior to transfer. This practice kept the callus actively growing and reduced the accumulation of growth-inhibiting phenolic exudates. The effectiveness of frequent subculturing for maintaining embryogenic competence has been noted by other researchers as well.
Chaudhary et al. (2015) report that regularly removing browned sections of callus and refreshing the medium can rejuvenate cultures and support continuous somatic embryogenesis. In our case, the callus pieces that remained creamy-white or green continued to produce new somatic embryos upon subsequent subcultures.
By around 14–16 weeks, Surkhan-106 embryogenic callus had generated numerous somatic embryos at different stages. The embryos typically emerged in clusters on the callus surface. We transferred these embryogenic calli with developing embryos to the Embryo Maturation Medium (EMM) for further development. On EMM (which contained 5 mg/L NAA but no 2,4-D or kinetin), embryos enlarged and matured. Many reached torpedo stage (5–7 mm in length) with visible cotyledonary structures. The inclusion of a moderate auxin (NAA) in the maturation medium likely helped in rooting capacity, while the reduction of cytokinin minimized callus overgrowth around the embryos.
In addition, we found that adjusting the nitrogen composition of the medium was beneficial at the maturation stage. Following the approach of
Davidonis et al. (1983), extra potassium nitrate (KNO₃) was added and the ammonium levels were balanced in the hormone-free Germination Medium (GM). Cotton somatic embryos are known to have improved maturation when provided with higher nitrate:ammonium ratios. The rationale is that developing embryos consume a lot of nitrogen, and additional nitrate can promote their growth and conversion to plantlets. In our protocol, embryogenic calli were moved from EMM to the hormone-free GM (with optimized nitrates) for the final phase of development and germination.
Once the somatic embryos of Surkhan-106 had matured (becoming torpedo-shaped and greening up), they began to germinate on the hormone-free GM. We initially incubated the plates in the dark for ~10 days to stimulate root emergence from the embryos – this mimics the natural germination of zygotic embryos which often root in darkness underground. Indeed, within a week on GM, many somatic embryos broke dormancy: the radical end (future root) elongated, and the hypocotyl axis extended. After 10–15 days, the germinated somatic embryos were producing small roots and shoots, effectively becoming somatic seedlings (tiny plantlets). These regenerating plantlets were then brought into light conditions to develop chlorophyll and expand their cotyledons (embryonic leaves). Over the next 2 weeks, the plantlets turned green and grew ~3–4 cm tall. We successfully recovered a number of whole cotton plantlets from somatic embryos of Surkhan-106 using this procedure.
Somatic embryos originating from callus cultures successfully regenerated into plantlets at a conversion rate of approximately 65 ± 3.5%, based on three independent biological replicates. Although some embryos exhibited morphological abnormalities or failed to germinate—a phenomenon commonly observed during cotton somatic embryogenesis (
Kumar et al. 2013b)—the majority developed into healthy plantlets. Each embryogenic callus produced an average of 5–6 mature somatic embryos, of which around 65% germinated and developed into complete plantlets. During the acclimatization stage, about 80 ± 4% of regenerated plantlets survived under ex vitro conditions. Statistical analysis (t-test,
p < 0.05) confirmed that the overall regeneration efficiency was significantly higher in hypocotyl-derived callus (85 ± 2%) compared with cotyledon-derived callus (60 ± 3%), consistent with previous findings on the superior morphogenic competence of hypocotyl tissue in cotton. The regenerated plantlets exhibited normal morphology, including a well-developed primary root, hypocotyl, and two cotyledonary leaves, resembling typical seedlings of the Surkhan-106 cultivar (
Table 1.). After a gradual acclimatization, the plantlets were transferred to soil pots in a greenhouse to confirm their viability. They continued to grow, eventually forming true leaves, indicating that they were fully regenerated cotton plants.
Table 1 summarizes the quantitative parameters of somatic embryogenesis and regeneration in Surkhan-106. Hypocotyl explants showed significantly higher callus induction and conversion rates compared to cotyledon explants (
p < 0.05).
Through this regeneration scheme, we established a complete plant regeneration cycle for Surkhan-106: seed → explant → callus → somatic embryo → plantlet. To our knowledge, this is the first report of successful plant regeneration via somatic embryogenesis in an Uzbek local cotton variety. The process took about 4–5 months from seed inoculation to recovered plantlets, which is comparable to timelines reported for other cotton cultivars in tissue culture.
Fig. 2 illustrates a key stage in the process, showing the embryogenic callus with multiple somatic embryos. The presence of both callus and embryo stages together highlights the indirect nature (callus-mediated) of this regeneration.
Our results highlight several important considerations for cotton tissue culture. Firstly, genotype is crucial – Surkhan-106 required specific conditions to regenerate, underlining that protocols must often be tailored to each cotton variety. Even with an optimized protocol, some somatic embryos did not convert to plants, reflecting an inherent limitation in cotton SE where maturation and germination remain less efficient than embryo induction. Secondly, the combination and concentration of plant growth regulators need to be finely tuned. We found that an initial high auxin (2,4-D) environment was essential for callus induction, but a reduction or removal of auxin was necessary for embryo differentiation and maturation. This is consistent with general observations that 2,4-D is embryogenic at callus initiation but must be withdrawn to allow embryo development, as high auxin can inhibit the later stages of embryo morphogenesis (
Kumar et al. 2013b).
Kumar et al. (2004) similarly noted that a temporary auxin starvation or reduction synchronized embryo development in cotton cultures.
Discussion
The present study establishes, for the first time, a reproducible and efficient somatic embryogenesis protocol for the Uzbek cotton cultivar Surkhan-106 (
Gossypium barbadense L.), which has been considered recalcitrant to in vitro regeneration. The results confirm that genotype plays a crucial role in determining regeneration capacity, as previously emphasized by
Trolinder et al. (1987). Among the two explant types tested, hypocotyl segments exhibited a significantly higher callus induction frequency (85 ± 2%) compared with cotyledon explants (60 ± 3%), consistent with earlier findings that hypocotyl tissues possess greater morphogenic potential (
Ahsan et al. 2014;
Wilkins et al. 2004).
In terms of efficiency, the callus induction frequency obtained for Surkhan-106 (85%) is comparable with that reported for the highly responsive model genotype Coker-312 (80–90%, (
Kumar et al. 2013a)). Similarly, Turkish cultivars such as Nazilli 143 and Nazilli M-503 showed moderate callus induction rates of 70–75% (
Aydin et al. 2004), while Chinese cultivar Simian-3 demonstrated a higher response (
Kumar et al. 2013b). These comparisons indicate that Surkhan-106 performs within the upper range of previously reported efficiencies, confirming the suitability of the optimized medium composition and subculture regime.
The embryo-to-plantlet conversion rate in Surkhan-106 reached 65 ± 3.5%, which is comparable to those reported for Coker-312 (60–70%, (
Trolinder et al. 1987)), Simian-3 (60–68%, (
Kumar et al. 2013a), and Nazilli 143 (62%, (
Aydin et al. 2004)). Although some somatic embryos exhibited morphological abnormalities or failed to germinate—a common phenomenon in cotton somatic embryogenesis—the majority developed into morphologically normal plantlets. Furthermore, approximately 80 ± 4% of regenerated plantlets survived during the acclimatization stage, indicating strong viability and physiological stability under ex vitro conditions.
The complete regeneration cycle from seed inoculation to acclimatized plantlets was accomplished within 4–5 months, which is comparable to the 4–6 months reported for Coker-312 (
Trolinder et al. 1987) and Nazilli 143 (
Aydin et al. 2004), and shorter than the 6–7 months required for Khandwa-2 (
Kumar et al. 2013b). Frequent subculturing every 3 weeks proved essential for maintaining callus viability and reducing browning—a major limiting factor in cotton tissue culture (
Chaudhary et al. 2015). This approach ensured a continuous supply of fresh nutrients and reduced the accumulation of phenolic compounds, thereby enhancing embryogenic competence and embryo quality.
The hormonal regime used in this study—initial high auxin concentration (2,4-D) during callus induction followed by a gradual shift to moderate NAA and cytokinin levels during maturation—proved highly effective. This pattern aligns with previous reports indicating that 2,4-D promotes embryogenic dedifferentiation at the early stage, while its subsequent reduction is required for embryo differentiation and germination (
Kumar et al. 2004). Additionally, the use of maltose as a carbon source likely contributed to the reduced phenolic oxidation and improved callus texture, supporting the findings of
Sakhanokho et al. (2016). Optimizing the nitrate:ammonium ratio in the germination medium further enhanced embryo maturation and conversion, consistent with the observations of
Davidonis et al. (1983).
Despite its effectiveness, the developed protocol has certain limitations. The approach is genotype-dependent and may require optimization for different cultivars. The embryo-to-plant conversion rate (~65%) indicates that some embryos fail to develop into normal plantlets. Frequent subculturing is labor-intensive and increases the risk of contamination, while long-term culture may cause somaclonal variation. Such variation may lead to genetic and phenotypic instability among regenerated plants, which should be carefully monitored when applying the protocol in breeding or transformation programs. However, controlled induction and screening of somaclonal variants can also provide useful genetic diversity for cotton improvement. Nevertheless, the proposed system provides a practical and time-efficient alternative to conventional breeding. Unlike standard breeding methods requiring several years of selection cycles, this protocol allows regenerated plants to be obtained within 4–5 months, significantly accelerating the introduction of new genetic constructs into elite cotton varieties.
Acknowledgments
The authors express their gratitude to the staff of the Cotton Breeding, Seed Production and Agrotechnologies research Institute, Uzbekistan, for providing the Surkhan-106 seed material and supportive facilities. This research was carried out in the “Cotton Biotechnology” Laboratory of the institute, as part of the national program on cotton crop improvement. We thank colleagues for technical assistance in tissue culture and the Ministry of Innovative Development of Uzbekistan for funding support.
Fig. 1Callus induction from different explants of cotton cv. Surkhan-106. Hypocotyl explants (left bar) showed a significantly higher callus induction frequency (approximately 85%) than cotyledon explants (right bar, ~60%). Error bars indicate ±SE for three replicates. Asterisks denote a statistically significant difference between the two explant types (p < 0.05, t-test). The superior callusing response of hypocotyl tissue is consistent with earlier findings in cotton tissue culture.
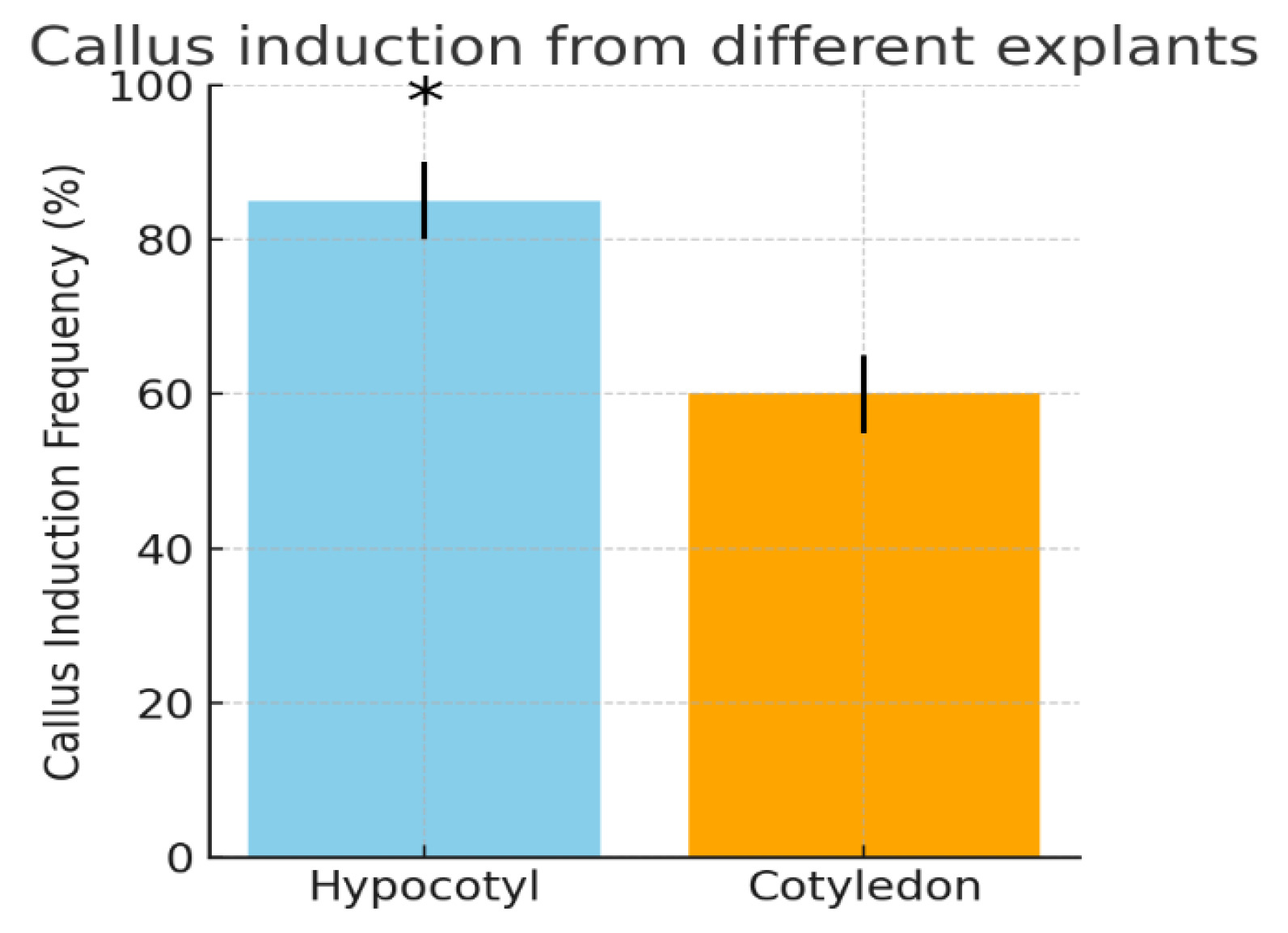
Fig. 2Somatic embryogenesis in Gossypium barbadense cultivar Surkhan-106.

Table 1Somatic embryogenesis and regeneration efficiency in cotton cultivar Surkhan-106.
Table 1
|
Parameter |
Explant type |
Mean ± SE |
n |
Statistical significance
(t-test, p < 0.05) |
|
Callus induction frequency (%) |
Hypocotyl |
85 ± 2.0 |
3 |
a |
|
Cotyledon |
60 ± 3.0 |
3 |
b |
|
SE induction rate (%) |
Hypocotyl |
72 ± 3.2 |
3 |
a |
|
Cotyledon |
48 ± 2.8 |
3 |
b |
|
Average embryos per callus |
Hypocotyl |
5.8 ± 0.4 |
3 |
a |
|
Cotyledon |
3.6 ± 0.3 |
3 |
b |
|
Embryo-to-plant conversion rate (%) |
Hypocotyl |
65 ± 3.5 |
3 |
A |
|
Cotyledon |
45 ± 2.5 |
3 |
B |
|
Survival rate after acclimatization (%) |
Combined |
80 ± 4.0 |
3 |
– |
References
- Abdurakhmonov IY, Buriev ZT, Saha S, Jenkins JN, Abdukarimov A, Pepper AE. 2014. Phytochrome RNAi enhances major fibre quality and agronomic traits of the cotton Gossypium hirsutum L. Nat. Commun. 5(1): 3062
- Ahsan MZ, Majidano MS, Channa AR, Panhwar FH, Soomro AW, Khaskheli FI, et al. 2014. Regeneration of cotton (Gossypium hirsutum L.) through asexual methods, a review. Am. Eurasian. J. Agric. Environ. Sci. 14(12): 1478-1486.
- Aydin Y, Ipekci Z, Talas-Oğraş T, Zehir A, Bajrovic K, Gozukirmizi N. 2004. High frequency somatic embryogenesis in cotton. Biol. Plant. 48(4): 491-495.
- Babadjanov J, Petrick M. 2025. Uzbekistan's cotton clusters in the context of the industrial policy debate. Eurasian Geogr. Econ. 66(3): 354-383.
- Bakhsh A, Anayol E, Sancak C, Özcan S. 2016. An efficient and cost effective sterilizing method with least microbial contamination and maximum germination ratio for in vitro cotton (Gossypium hirsutum L.) culture. J. Anim. Plant Sci. 26(3): 47-58.
- Chaudhary G, Dantu PK. 2015. Evaluation of callus browning and develop a strategically callus culturing of Boerhaavia diffusa L. J. Plant Dev. 22: 868-873.
- Cheng GW, Crisosto CH. 1995. Browning potential, phenolic composition, and polyphenoloxidase activity of buffer extracts of peach and nectarine skin tissue. J. Am. Soc. Hortic. Sci. 120(5): 835-838.
- Davidonis GH, Hamilton RH. 1983. Plant regeneration from callus tissue of Gossypium hirsutum L. Plant Sci. Lett. 32(1-2): 89-93..
- Fang L, Gong H, Hu Y, Liu C, Zhou B, Huang T, et al. 2017. Genomic insights into divergence and dual domestication of cultivated allotetraploid cottons. Genome Biol. 18(1): 33
- Guo H, Guo H, Zhang L, Fan Y, Wu J, Tang Z, et al. 2020. Dynamic Transcriptome Analysis Reveals Uncharacterized Complex Regulatory Pathway Underlying Genotype-Recalcitrant Somatic Embryogenesis Transdifferentiation in Cotton. Genes. 11(5): 519
- Jin S, Zhang X, Liang S, Nie Y, Guo X, Huang C. 2005. Factors affecting transformation efficiency of embryogenic callus of Upland cotton (Gossypium hirsutum) with Agrobacterium tumefaciens. Plant Cell Tiss. Org. 81(2): 229-237.
- Kumar M, Tuli R. 2004. Plant regeneration in cotton: A short-term inositol starvation promotes developmental synchrony in somatic embryogenesis. In Vitro Cell. Dev. Biol. Plant. 40(3): 294-298.
- Kumar M, Shukla A, Singh H, Verma P, Pradhyumna , Singh K. 2013a. A genotype-independent Agrobacterium mediated transformation of germinated embryo of cotton (Gossypium hirsutum L.). Int. Res. J. Biotechnol. 3: 81-90.
- Kumar M, Singh H, Shukla AK, Verma PC, Singh PK. 2013b. Induction and establishment of somatic embryogenesis in elite Indian cotton cultivar (Gossypium hirsutum L. cv Khandwa-2). Plant Signal. Behav. 8(10): e26762
- Pathi KM, Tuteja N. 2013. High-frequency regeneration via multiple shoot induction of an elite recalcitrant cotton (Gossypium hirsutum L. cv Narashima) by using embryo apex. Plant Signal. Behav. 8(1): e22763
- Ramírez-Mosqueda MA, Cadena-Zamudio JD, Cruz-Cruz CA, Aguirre-Noyola JL, Barbón R, Gómez-Kosky R, et al. 2025. Somatic Embryogenesis: A Biotechnological Route in the Production of Recombinant Proteins. BioTech. 14(4): 93
- Sakhanokho HF, Rajasekaran K. 2016. Cotton regeneration in vitro. pp.87-110. In: KG. Ramawat, MR. Ahuja (eds.). Fiber Plants: Biology, Biotechnology and Applications. Cham: Springer.
- Saud S, Wang L. 2022. Mechanism of cotton resistance to abiotic stress, and recent research advances in the osmoregulation related genes. Front. Plant Sci. 13: 2022
- Shahzad K, Mubeen I, Zhang M, Zhang X, Wu J, Xing C. 2022. Progress and perspective on cotton breeding in Pakistan. J. Cotton Res. 5(1): 29
- Shazia Yasin SY, Aneela Yasmin AY. 2018. Optimisation of in vitro propagation of Gossypium arboreum L. Pure Appl. Biol. 7(2): 419-426.
- Trolinder NL, Goodin JR. 1987. Somatic embryogenesis and plant regeneration in cotton (Gossypium hirsutum L.). Plant Cell Rep. 6(3): 231-234.
- Trolinder NL, Goodin JR. 1988. Somatic embryogenesis in cotton (Gossypium) I. Effects of source of explant and hormone regime. Plant Cell Tissue Organ. Cult. 12(1): 31-42.
- Wilkins TA, Mishra R, Trolinder NL. 2004. Agrobacterium-mediated transformation and regeneration of cotton. J. Food Agric. Environ. 2(1): 179-187.
- Zhang BH, Feng R, Liu F, Wang Q. 2001. High frequency somatic embryogenesis and plant regeneration of an elite Chinese cotton variety. Bot. Bull. Acad. Sin. 42: 9-16.
- Zhang T, Wang S, Chen J, Zhu S, Zhu Q, Zhao T. 2024. Advances in Cotton Genomics, Genetics and Breeding. Plants. 13: 2579