Abstract
This research was conducted to evaluate the contribution of four candidate genes — CsABA2 (CsGy5G015450), CsLRR-RPK2 (CsGy5G015660), CsPM5.2 (CsGy5G015960), and CsaMLO8 (CsGy5G026660) — to powdery mildew (PM) resistance in cucumber germplasms. For this purpose, fifty-four germplasms, consisting of thirty-three breeding lines and twenty-one landraces from fourteen diverse origins, were evaluated for PM-resistance at the adult plant stage and genotyped using molecular markers associated with the four PM-resistance-related genes. According to the phenotypic assessment, sixteen germplasms were susceptible, nineteen germplasms exhibited intermediate-resistant, and nineteen germplasms demonstrated resistant to PM. The results from phenotypic evaluation and genotyping analysis indicated that among the PM-susceptible germplasms, 50% of CsLRR-RPK2, 31% of CsABA2, 25% of CsaMLO8, and 18% of CsPM5.2 had inconsistent genotypes. Genotype inconsistencies among PM-resistant germplasms were observed in 16% of CsLRR-RPK2, 16% of CsABA2, 10% of CsaMLO8, and 10% of CsPM5.2. This study showed that the combination of CsPM5.2 and CsaMLO8 alleles is more effective and beneficial for PM-resistance breeding programs in cucumber. And also, this suggests there is a relationship between PM-resistance and the genotype of a combination of PM-resistance candidate genes. The findings of the genotyping and PM-phenotypic evaluation of the current study may improve selection accuracy in breeding for PM-resistance in cucumber.
-
Key words: Cucumis sativus L., CsABA2, CsLRR-RPK2, CsPM5.2, CsaMLO8, PM-resistance marker-assisted breeding
Introduction
Cucumber (
Cucumis sativus L., 2n = 2x = 14) is one of the most widely grown and consumed vegetable crops globally, with an annual production exceeding sixty-five million tons. It ranks among the top four most cultivated vegetables in the world, following tomatoes, brassicas, and onions (FAOSTAT, www.fao.org/faostat/en/#data/QCL, 2022). It also is a valuable health food source due to its high-water content, low-calorie profile, contribution to hydration, and weight management (
Badri Anarjan et al. 2024;
Badri Anarjan et al. 2021;
Kishor et al. 2021;
Samba et al. 2024) Considering this, improving the cucumber quality and yield is critical.
Cucumber production has been challenged by the occurrence of both pre- and post-harvest stresses. Reductions in cucumber quality and yield can occur at any plant growth stage (
Badri Anarjan et al. 2024). Among the most severe biotic stresses impacting cucumber both in open-field and in protected cultivations is powdery mildew (PM), which leads to declines in both quality and yield (
Badri Anarjan et al. 2024;
Zhang et al. 2021). Infection by
Podosphaera xanthii (P. xanthii) causes chlorosis, premature drying, and affects photosynthesis, leading to yield losses. Additionally, it impacts taste, quality, and market value of the fruit causing sunscald (
Alavilli et al. 2022). To manage PM in cucumbers, protective fungicides are often used. However, over the years, the fungus
P. xanthii has developed resistance to these fungicides, and also many consumers have become increasingly concerned about food safety issues caused by pesticide overuse. Breeding cucumbers by conventional methods is labor-intensive and time-consuming. Environmental factors play a major role in the occurrence of PM. The PM is an obligate parasite, and it is difficult to screen resistance at the right time through artificial inoculation. In cucumber, it has been reported that PM-resistance is linked to major quantitative trait loci (QTL) or genes in a number of studies. It is a complex trait influenced by various factors including physiological races, genetic background, environmental factors such as temperature and humidity, cultivation season, and inoculation method. Consequently, breeding PM-resistant cucumber varieties is more complex and difficult to develop (
Badri Anarjan et al. 2021;
Hosoya et al. 1999;
Innark et al. 2014;
Nie et al. 2015a;
Nie et al. 2015b;
Nie et al. 2023;
Sun et al. 2024;
Zhang et al. 2021).
Marker-assisted breeding (MAB) of PM-resistance, on the other hand, has several advantages, including a reduction in environmental impact, a reduction in labor requirements, and a substantial shortening of the breeding period. For this, the selection of proper markers is crucial for accurate selection of resistant genotypes. The application of marker-assisted selection (MAS) to cucumber is lagging behind other vegetable crops, especially in Korea. Therefore, to breed PM-resistant cucumber varieties, MAB remains the most effective approach (
Badri Anarjan et al. 2021;
Nie et al. 2023;
Sun et al. 2024;
Zhang et al. 2021). Several QTLs have been linked to PM-resistance in cucumbers, but their effects on resistance of different germplasms with different genetic backgrounds have not been examined. Additionally, there are few reports on selection accuracy between the PM-resistance-related genes in cucumbers. Combining multiple genetic markers can indeed improve selection accuracy in PM-breeding programs. A study conducted by
Badri Anarjan et al. (2021) focused on the application of PM-resistance molecular markers derived from two candidate genes,
CsaMLO8 and
CsLRR-RPK2. In this study, 100 inbred lines were genotyped for PM-resistance, and the types of resistance sources were assessed. The results indicated that the
CsLRR-RPK2-CAPS marker demonstrated a strong correlation with the PM-resistant phenotype compared to the
CsaMLO8-InDel marker.
Recent studies have identified key putative candidate genes associated with resistance to cucumber PM, including
CsaMLO8 (
CsGy5G026660), a mildew resistance locus O (MLO)-like gene identified in PI 197088 and the variety ‘Anaxo’. Loss-of-function mutations in
CsaMLO8 impair fungal entry at the penetration stage and thereby confer resistance, consistent with its role as a susceptibility gene regulating cell wall apposition and pathogen accommodation (
Berg et al. 2017;
Berg et al. 2015;
Nie et al. 2015a;
Nie et al. 2015b).
CsABA2 (
CsGy5G015450) was identified through a genome-wide association study (GWAS) in a panel of 264 accessions and encodes xanthoxin dehydrogenase, a key enzyme in abscisic acid (ABA) biosynthesis (
Lee et al. 2020).
CsLRR-RPK2 (
CsGy5G015660) was discovered in a Korean PM-resistant inbred line and encodes a leucine-rich repeat receptor-like kinase (LRR-RPK) that functions in pattern-recognition receptor (PRR)-mediated immunity by initiating defense signaling cascades upon pathogen detection (
Zhang et al. 2021).
CsPM5.2 (
CsGy5G015960) was identified in the North Chinese-type inbred line C-8-6 and encodes a phosphate transporter-like protein PHO1; H3, whose expression is associated with enhanced resistance and potential involvement in salicylic acid (SA)-dependent defense signaling (
Sun et al. 2024).
CsaMLO8,
CsABA2,
CsLRR-RPK2, and
CsPM5.2 have been implicated in cucumber PM-resistance and are involved in distinct defense-related pathways: cell wall-associated susceptibility, ABA biosynthesis, receptor-like kinase-mediated signaling, and phosphate transport/SA signaling, respectively. These genes are all located on chromosome 5, which has been identified as a major hotspot for cucumber PM-resistance. In this study, four PM-resistance candidate genes (
CsABA2,
CsLRR-RPK2,
CsPM5.2, and
CsaMLO8) were employed as molecular indicators to evaluate the effectiveness of MAS in fifty-four cucumber accessions and to understand the origin of their resistance to PM. The results showed that combining
CsPM5.2 and
CsaMLO8 in cucumber would be more effective and advantageous for breeding PM-resistance. Combinations of PM-resistance candidate genes appear to be responsible for the different levels of resistance observed. This study is expected to contribute to the development of varieties with improved PM-resistance by increasing selection accuracy in molecular breeding.
Materials and Methods
Plant material and growth conditions
To conduct this study, we used fifty-four cucumber germplasms, including twenty-one landraces and thirty-three breeding lines, representative of typical variety groups from various origins, including Brazil, China, France, India, Iran, Japan, Kenya, Korea, Mauritius (East-Africa), the Netherlands, the Philippines, Taiwan, Thailand, and the United States. Detailed information about the germplasms is presented in
Table 1. In the spring of 2024, the germplasms’ seeds were germinated indoors on March 4
th and transplanted in the greenhouse in the experimental farm of Sejong University in Anseong, Korea, on April 25
th. The germplasms were evaluated for PM-resistance at the adult plant stage (25~30 nodes) and genotyped with markers developed from the genes associated with PM-resistance.
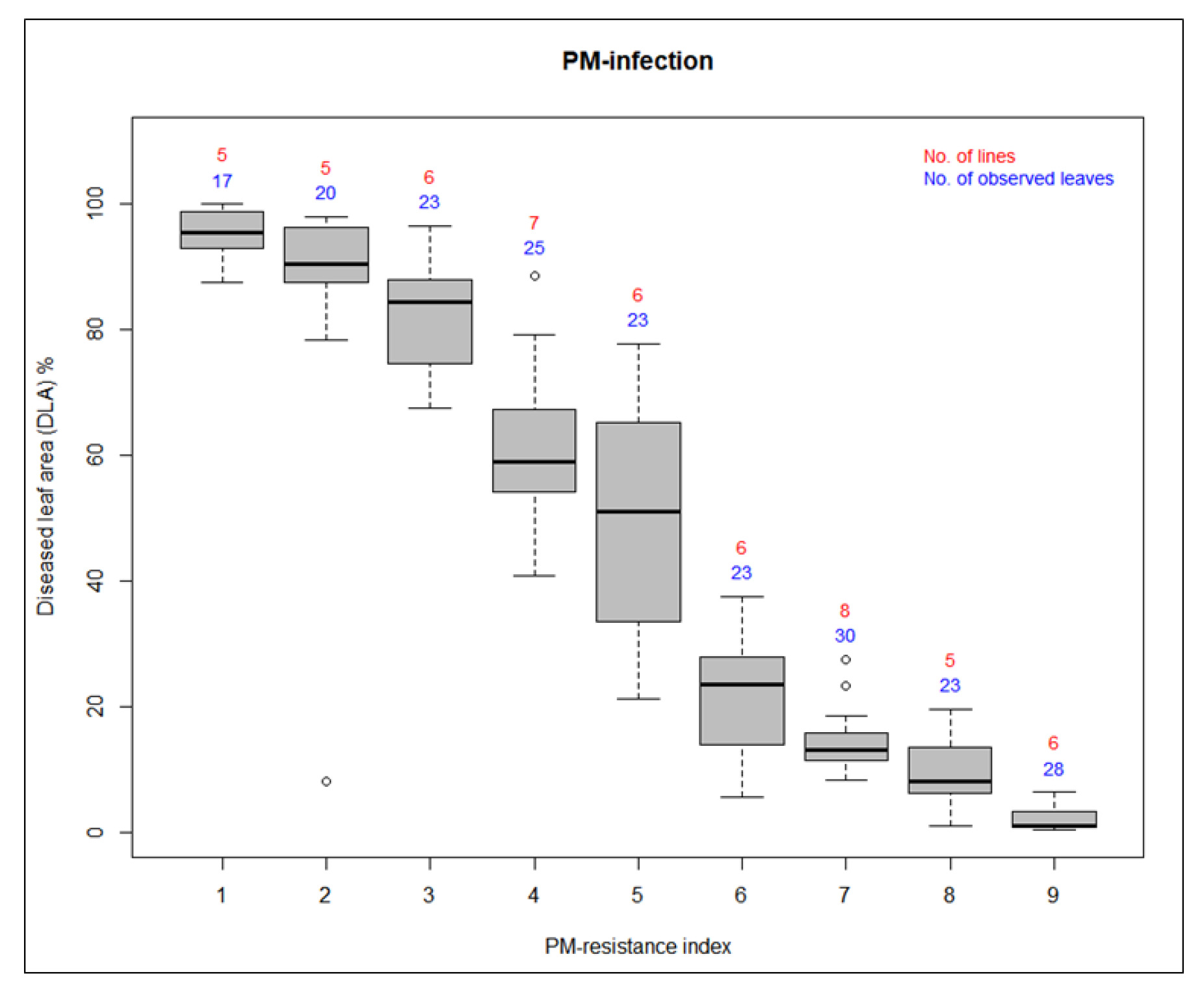
The evaluation of PM-resistance at the adult plant stage of the fifty-four germplasms was performed by visual scoring and quantification of the percentage of diseased leaf area (DLA). Sixty days post-transplantation, visible disease symptoms developed on plants naturally infected with P. xanthii in the greenhouse environment. Phenotypic evaluation of PM-resistance was conducted at two-week intervals beginning when disease symptoms were conspicuously evident. This assessment continued until susceptible plants exhibited complete disease coverage. The PM response of each cucumber germplasm was determined by averaging disease scores from five individual plants. The scores were calculated using data collected from three separate rating sessions conducted on June 25th, July 9th, and July 23rd.
A visual disease resistance scale ranging from 1 (susceptible) and 9 (resistant) was used to evaluate the PM-resistance of the germplasms. Ratings were categorized as susceptible (1~3), intermediate resistant (4~6), and resistant (7~9). We analyzed diseased leaf area (DLA) % of each germplasm to quantify the disease resistance. For this, the image data were analyzed with the online software ImageJ (www.imagej.nih.gov/ij). An image of the leaf samples was taken using a Samsung Galaxy S105G equipped with a 12-megapixel rear camera, a 6 mm wide-angle lens, and an aperture of F2.4 (telephoto). For accuracy, we used the same color threshold value for all the samples since we imported the processed images into ImageJ. Based on the “diseased area / whole leaf area × 100”, diseased leaf area (DLA) % was calculated (
Alavilli et al. 2022). Finally, by performing visual scoring and quantification of the percentage of DLA, a clear classification of the cucumber germplasms into PM-susceptible, PM-intermediate resistant, and PM-resistant groups was established. Detailed analysis of PM-phenotypic evaluation is presented in
Figs. 1,
2 and
Table 1.
Genomic DNA (gDNA) was isolated from the fresh young leaves of fifty-four germplasms by employing the PureHelix™ Genomic DNA Prep Kit, Ver. 3.0 (NanoHelix Co., Ltd., Daejeon, Korea). Subsequent assessment of DNA quality and quantity was performed using a Thermo Scientific™ NanoDrop™ OneC according to the manufacturer's protocol.
The objective of this study was to assess the involvement of alleles from four candidate genes (
CsABA2,
CsLRR-RPK2,
CsPM5.2, and
CsaMLO8) in conferring PM-resistance. To accomplish this, molecular markers associated with these PM-resistance genes were utilized to evaluate the effectiveness of MAS in a set of germplasms. Cleaved amplified polymorphic sequence (CAPS) and insertion-deletion (InDel) markers, based on the methodology outlined by
Zhang et al. (2021) and
Badri Anarjan et al. (2021), were utilized for the analysis of
CsLRR-RPK2 and
CsaMLO8 genes, respectively. Derived cleaved amplified polymorphic sequences (dCAPS) markers were developed using the online dCAPS finder 2.0 (http://helix.wustl.edu/dcaps/) for the analysis of
CsABA2 and
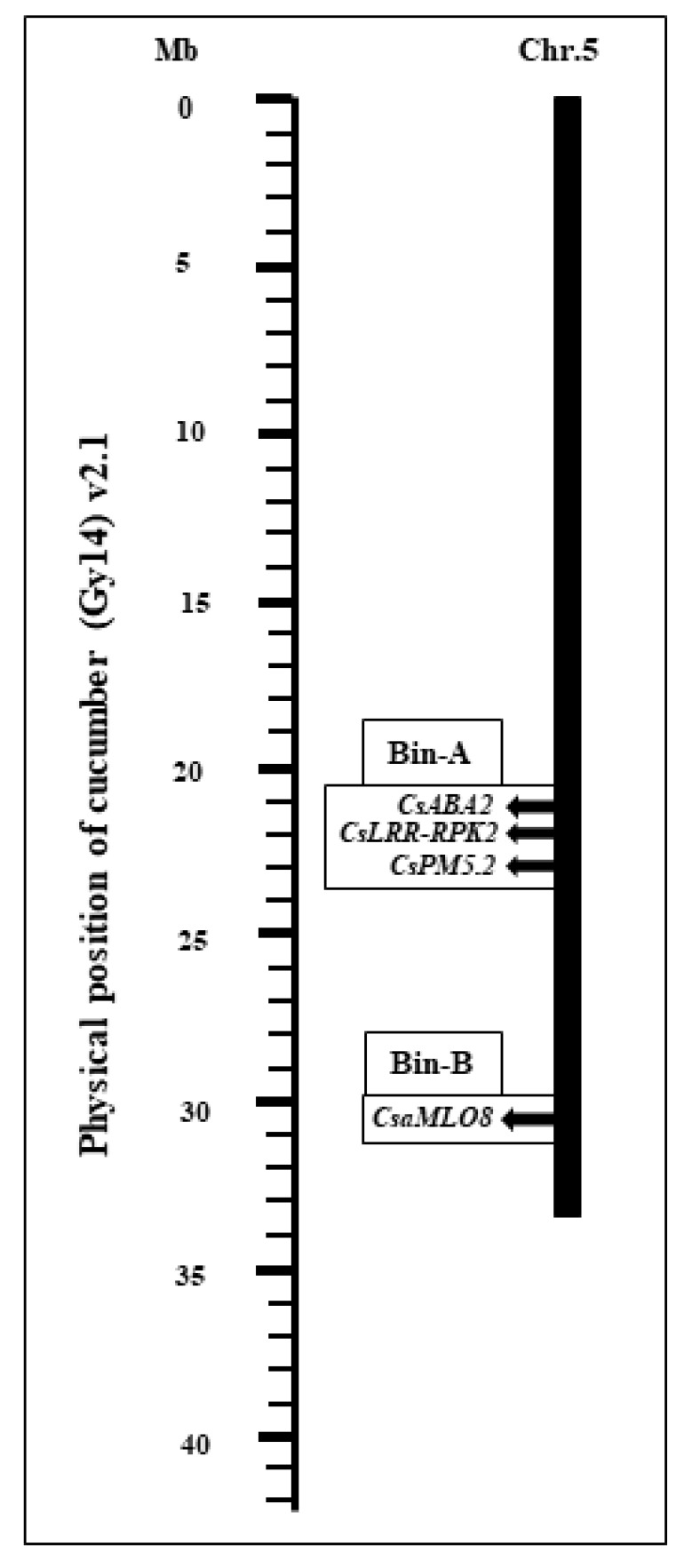
CsPM5.2 genes. The chromosomal positions of four candidate genes associated with PM-resistance on chromosome 5 along with their corresponding marker information are presented in
Fig. 3 and
Table 2.
The DNA amplification of CsABA2, CsLRR-RPK2, and CsPM5.2 was performed through the polymerase chain reaction (PCR) protocol consisting of an initial denaturation step at 94℃ for 5 minutes, followed by 35 cycles of denaturation at 94℃ for 20 seconds, annealing at 57℃, 60℃, and 60℃ for 10 seconds, and extension at 72℃ for 20 seconds. A final extension step at 72℃ for 5 minutes was included. The DNA amplification of CsaMLO8 was performed through PCR, which consists of an initial denaturation step at 94℃ for 5 minutes, followed by 30 cycles of denaturation at 94℃ for 20 seconds, annealing at 65℃ for 20 seconds, and extension at 72℃ for 90 seconds. A final extension step at 72℃ for 5 minutes was included. The PCR amplification was performed using Maxime PCR PreMix (iNtRON Biotechnology, Seoul, Korea) in a TaKaRa PCR Thermal Cycler Dice Touch TP350 (Japan). Restriction enzymes NcoI, HinfI, and Mn1I (New England BioLabs, Ipswich, MA, USA) were added to the PCR amplicons for CsABA2, CsLRR-RPK2, and CsPM5.2, respectively. The digestion reaction was incubated at 37℃ for 1 hour. The final PCR products were subjected to electrophoresis on 2% (CAPS and InDel markers) and 3% (dCAPS markers) agarose gels using 0.5x Tris-borate-EDTA (TBE) buffer. The DNA bands were visualized by staining with RedSafe nucleic acid stain (iNtRON Biotechnology, Seoul, Korea).
Results
The assessment of PM-resistance and genotyping analysis using four alleles (
CsABA2,
CsLRR-RPK2,
CsPM5.2, and
CsaMLO8) were illustrated in
Figs. 1,
2,
3,
4 and
Table 1. According to the phenotypic assessment of the germplasms, sixteen were susceptible to PM, whereas nineteen were intermediate-resistant. Disease resistance was demonstrated in the remaining nineteen germplasms. For
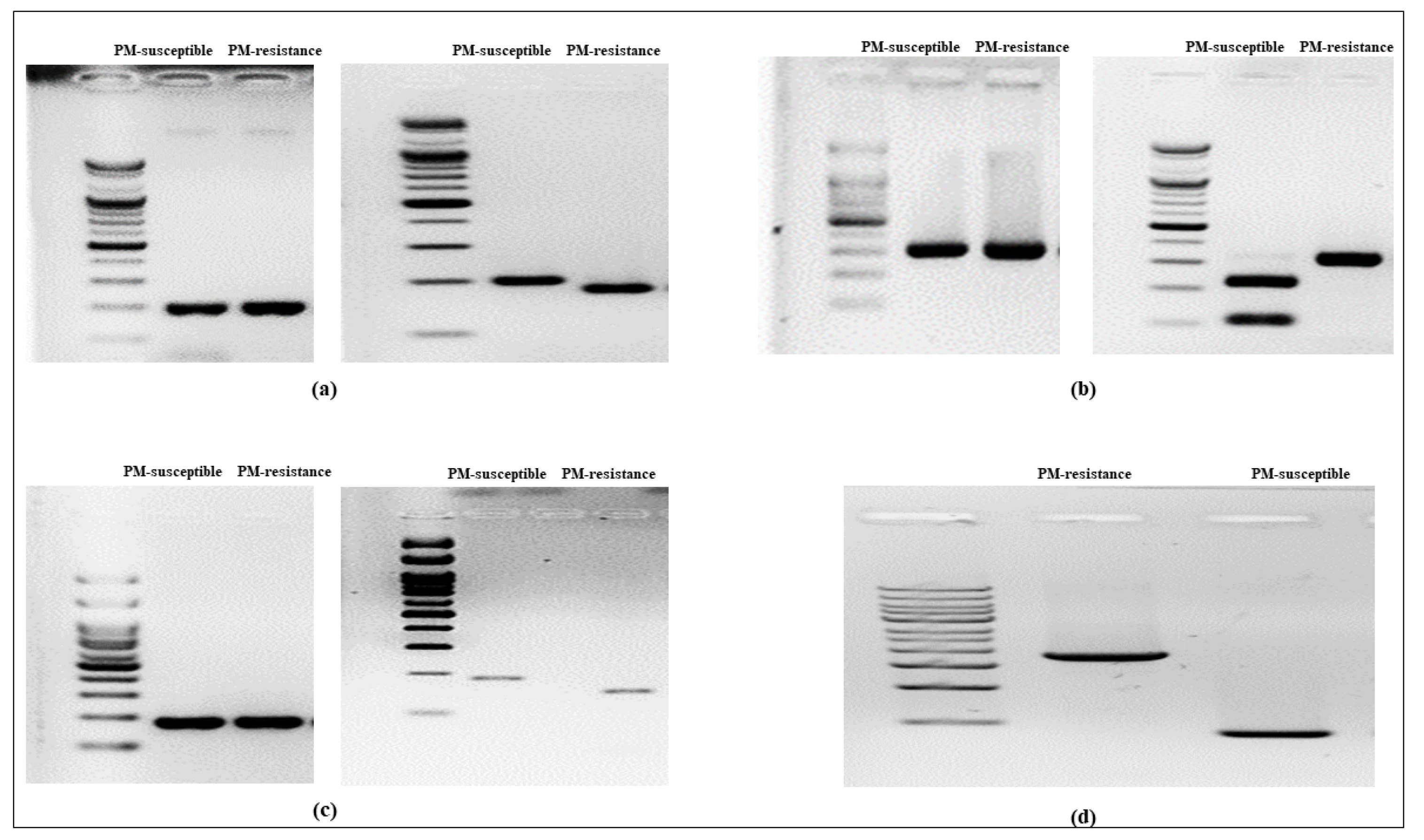
CsABA2, the dCAPS primer set (
CsABA2-dCAPS) amplified both flanking regions of the single nucleotide polymorphism (SNP) within its coding region. A double band (175 bp and 25 bp) was observed after NcoI digestion of the SNP of the PM-resistant line, whereas a restriction site was absent from the PM-susceptible line (
Fig. 4a). In the case of
CsLRR-RPK2, the CAPS primer set (
CsLRR-RPK2-CAPS) amplified the flanking region of the SNP located in the coding region. After HinfI digestion of the SNP of the PM-susceptible line, double bands (224 bp and 100 bp) were observed, whereas the PM-resistant line showed an absence of restriction sites (
Fig. 4b). Regarding
CsPM5.2, the flanking areas of the SNP within its coding region were amplified by the dCAPS primer set (
CsPM5.2-dCAPS). Mn1I digestion of the PM-resistant line's SNP produced a double band (152 bp and 31 bp), while the PM-susceptible line's SNP lacked a restriction site (
Fig. 4c). Regarding
CsaMLO8, both flanking areas of the 1,449 bp insertion inside the 11
th coding region of the PM-resistant line were amplified by InDel primers (
CsaMLO8-InDel). The PM-resistant line showed a band of 1,854 bp, while the PM-susceptible line showed a band of 405 bp (
Fig. 4d).
Sixteen germplasms displayed phenotypic susceptibility to PM. Of these, seven were breeding lines and nine were landraces. Genotyping analyses revealed 50% (8/16) of
CsLRR-RPK2, 31% (5/16) of
CsABA2, 25% (4/16) of
CsaMLO8, and 18% (3/16) of
CsPM5.2 had inconsistent genotypes. Therefore, in the course of developing PM-resistance breeding programs, analysis of the
CsPM5.2 allele could be more useful than the other three genes to eliminate PM-susceptible germplasms (
Table 1).
Nineteen germplasms displayed phenotypic resistance to PM. Of these, fourteen were breeding lines and five were landraces. Genotyping analyses revealed that 16% [(3/19); SJC2309, SJC2324, and “11758”] of
CsABA2, 16% [(3/19); SJC2309, SJC2324, and “Concombre blanc; ABD 14”] of
CsLRR-RPK2, 10% [(2/19); “Concombre blanc; ABD 14” and “11758”] of
CsPM5.2, and 10% [(2/19); “Concombre blanc; ABD 14” and “11758”] of
CsaMLO8 exhibited inconsistent genotypes (
Table 1). Among the five landraces with a PM-resistant phenotype, the resistance in “Hei Pei Ta Kua”, “Seoul Madi”, and “UPL-Cu-2 (PI 426170)” is derived from both bins A and B, whereas “Concombre blanc; ABD 14” and “11758” carry resistance derived only from bin A. In addition, all fourteen breeding lines with a PM-resistant phenotype harbor resistance sources derived from both bins A and B (
Table 1), reflecting the pyramiding of bins A and B resistance alleles in our breeding program. Therefore, in PM-resistant breeding programs, analyzing alleles at the two PM-resistance candidate genes (
CsPM5.2 and
CsaMLO8) appears to be effective for identifying PM-resistant plants.
Nineteen of the germplasms showed intermediate-resistant to PM in their phenotype. Twelve of them are breeding lines, while seven are landraces. Genotyping analyses revealed that SJC2314, SJC2316, SJC2317, SJC2328, SJC2333, “UPL-Cu-1”, “PI 419017”, and “San dong si gua 22b” exhibited a PM-resistance genotype for all four candidate genes. Despite this, their phenotype is intermediate-resistant. SJC2318 and SJC2331exhibited a PM-susceptible genotype for all four candidate genes. Despite this, their phenotype is intermediate-resistant. This fact suggests that their resistance may be controlled by other factors or that they possess a different genetic background influencing PM-resistance. Among the remaining nine PM-intermediate resistant germplasms, PM-resistance in SJC2330, “Yomaki-asahi Aonaga;73”, and “TCMG” is derived from the bin-A source. The PM-resistance in SJC2315 and “Tu-Pien-Chiao-San-Chi” originates from the bin-B source. The PM-resistance in SJC2319, SJC2329, SJC2332, and “Cool and Crisp” partially derives from both bins A and B (
Fig. 3;
Table 1).
Discussion
Cucumbers are among the world's most economically significant vegetable crops and are gaining attention for breeding new value-added varieties. However, their production is threatened by PM. The development of effective MAS applications is crucial for accelerating PM-resistance breeding in cucumbers, thereby enabling breeders to develop more PM-resilient varieties. The basic idea behind molecular breeding for cucumber PM-resistance is to identify markers linked to susceptible (S) and/or resistance (R) genes and introduce resistance alleles into high-quality cucumber varieties (
Badri Anarjan et al. 2021;
Nie et al. 2023;
Sun et al. 2024;
Zhang et al. 2021). MAS for PM-resistance in cucumbers has indeed been less advanced in Korea than in other countries, where substantial progress has been made in developing PM-resistant varieties. The findings of this study represent a promising step forward through the identification of four PM-resistance alleles (
CsABA2,
CsLRR-RPK2,
CsPM5.2, and
CsaMLO8) that serve as efficient markers for PM-resistance in cucumbers. Leveraging these markers can significantly enhance breeding efficiency for PM-resistance, providing a focused approach to developing resilient cucumber varieties through molecular breeding.
Studies indicate that PM-resistance in cucumbers is a highly complex trait. Despite variations in the results of different genetic analyses, there are notable similarities in the findings across various research efforts. Thus far, it has been demonstrated in several previous studies that PM-resistance in cucumber is linked to the loss-of-function of the
CsaMLO8 gene (
Berg et al. 2017;
Berg et al. 2015;
Liu et al. 2017;
Nie et al. 2015a;
Nie et al. 2015b). Nonetheless,
MLO-mediated resistance still has several major problems. First, other genes must be involved to achieve high levels of PM-resistance through
MLO-based mechanisms; however, there is little research on this topic to date (
Li et al. 2022). Second, leaf senescence and PM-resistance are tightly linked and are strongly influenced by ethylene signaling in cucumber (
Zhang et al. 2022). This implies that the expression and agronomic consequences of
CsaMLO8-mediated resistance depend on the ethylene-related genetic background, so introgressing a
CsaMLO8 loss-of-function allele into different elite cultivars may lead to variable levels of resistance and potential trade-offs, such as altered senescence dynamics or yield penalties. Third, cucumber plants deploy multiple layers of defense against PM pathogens, including SA-dependent signaling pathways (
Sun et al. 2024;
Wang et al. 2009;
Xiao et al. 2001), and PM-resistance is further modulated by environmental factors such as temperature (
Sakata et al. 2006), light quality (
Wang et al. 2010), and hydrogen peroxide (H₂O₂) accumulation (
Wang et al. 2010). Thus, the
CsaMLO8-based resistance operates within a broader, SA- and environment-dependent defense network, and a single
CsaMLO8 loss-of-function allele may not provide consistently stable resistance under diverse growing conditions. Fourth, studies in Korean cucumber lines have shown that PM-resistance is associated with specific combinations of alleles at
CsLRR-RPK2 and
CsaMLO8 (
Badri Anarjan et al. 2021;
Nie et al. 2023;
Zhang et al. 2021), indicating that
CsaMLO8 does not act in isolation but functions together with other resistance-related genes. This supports the view that effective utilization of
CsaMLO8 in breeding programs will require pyramiding it with additional PM-resistance loci rather than relying on
CsaMLO8 loss-of-function alone. The examination of four cucumber PM-resistance genes (
CsABA2,
CsLRR-RPK2,
CsPM5.2, and
CsaMLO8) in the present study confirmed that there is a relationship between PM-resistance and the genotypic combinations at these candidate loci.
Conclusions
This study revealed that the correspondence between genes involved in PM-resistance and phenotypes showed more accurate selection is possible by applying a combination of two genes, CsPM5.2 and CsaMLO8. Additionally, the results indicate that the PM-resistance of adult plants increases through the combination of resistance-related genes. Future research on PM-resistance in cucumber could focus on several key areas to deepen understanding of the functional genomics of resistance genes, the identification of new resistance genes, and development of high-throughput markers for new genes and phenotyping for durable PM-resistance under field conditions.
Fig. 1The phenotypic response of cucumber to PM-infection. (a) shows visual scoring of PM-resistance index using a 1~9 scale. Ratings of 1~3 indicates PM-susceptibility, 4~6 represents PM-intermediate resistance, and 7~9 denotes PM-resistance. (b) shows percentage of quantified PM-disease index [diseased leaf area (DLA %)] by ImageJ for the same leaves of various disease severities of PM. Red-colored regions indicate PM-lesions detected using the ImageJ thresholding function. The threshold was set based on color and brightness contrast to distinguish infected tissues from healthy areas. ImageJ was configured to detect both the lesion area and the total leaf area at the pixel level, and these measurements were used to calculate the percentage of diseased area relative to the whole leaf.

Fig. 2Distribution of diseased leaf area (DLA) % according to the PM-resistance index
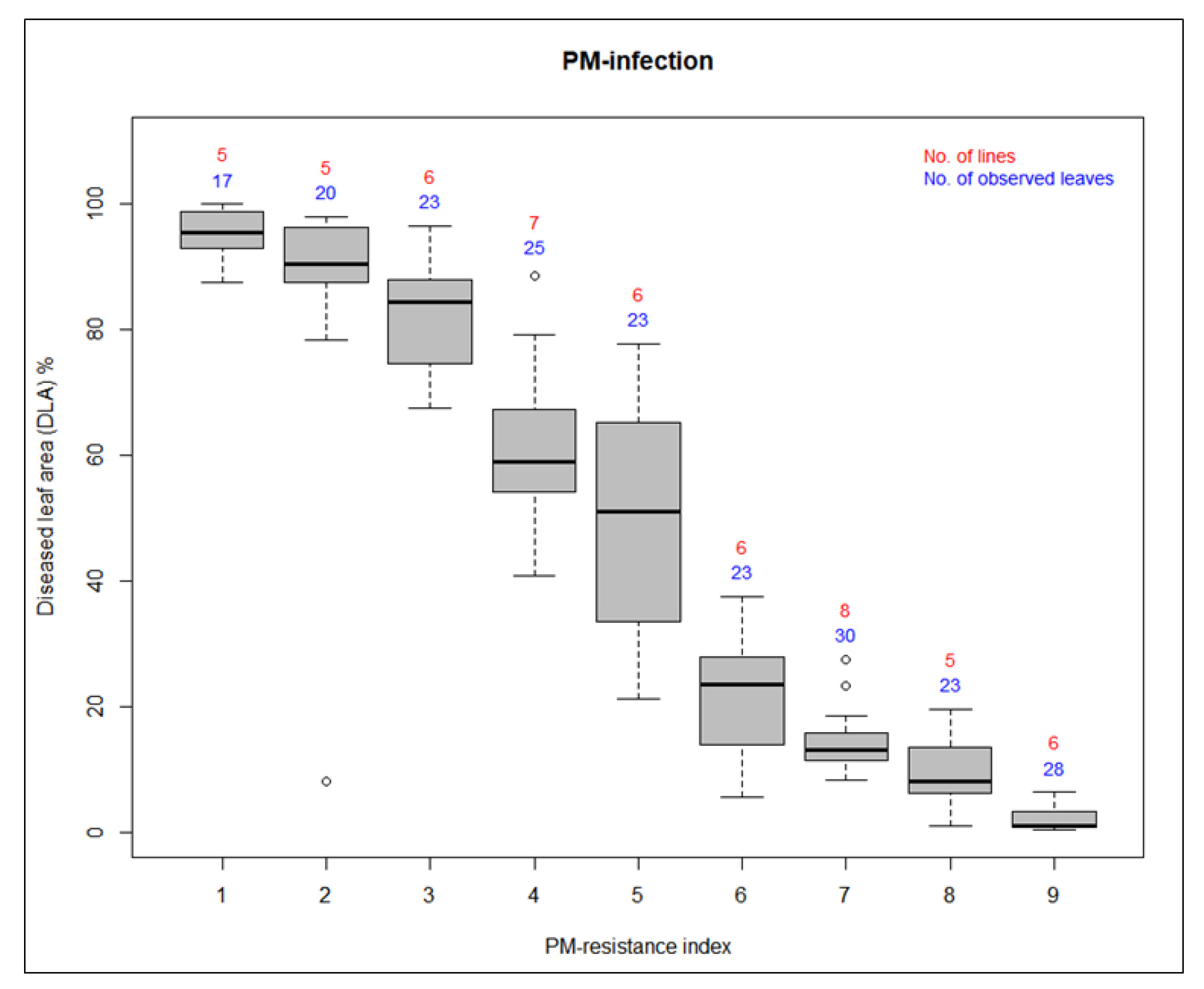
Fig. 3Chromosomal locations of cucumber candidate genes for PM-resistance. The genetic loci for CsABA2, CsLRR-RPK2, and CsPM5.2 are located within bin-A, while CsaMLO8 is found within bin-B.
Fig. 4Genotyping analysis using markers for cucumber PM-resistance candidate genes. (a) shows the bands amplified by the
CsABA2 gene primer (left) and the band patterns after NcoI enzyme application (right); (b) shows the bands amplified by the
CsLRR-RPK2 gene primer (left) and the band patterns after HinfI enzyme application (right); (c) shows the bands amplified by the
CsPM5.2 gene primer (left) and the band patterns after Mn1I enzyme application (right); and (d) shows the bands amplified by the
CsaMLO8 gene primer. Details on the markers for cucumber PM-resistance candidate genes are provided in
Table 2.
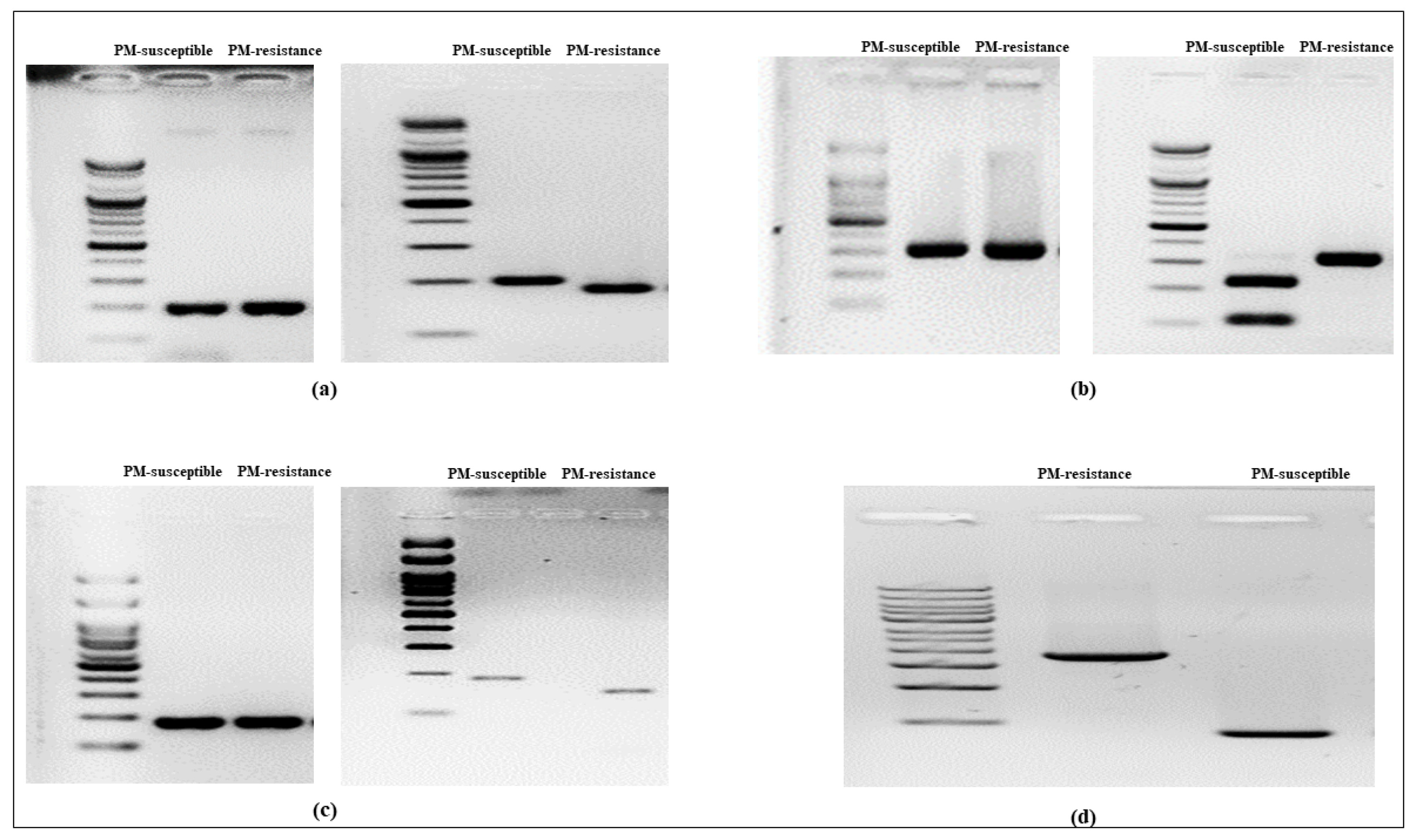
Table 1Results of phenotypic evaluation and genotyping analysis for fifty-four cucumber germplasms
Table 1
|
# |
Germplasm type |
Variety groupz)
|
Line name |
Origin |
Diseased leaf area (DLA) % |
Resistance index |
Phenotypic evaluationy)
|
PM-resistance sourcex)
|
CsABA2
|
CsLRR-RPK2
|
CsPM5.2
|
CsaMLO8
|
|
1 |
Breeding Line |
KSL |
SJC2301 |
Korea |
92.3±3.8 |
2.3±1.2 |
S |
- |
S |
S |
S |
S |
|
2 |
KSL |
SJC2302 |
97.5±2.2 |
1.1±0.3 |
S |
- |
S |
S |
S |
S |
|
3 |
KSL |
SJC2303 |
89.9±4.3 |
3.3±0.9 |
S |
- |
S |
S |
S |
S |
|
4 |
CHS |
SJC2304 |
72.7±4.3 |
2.9±1.3 |
S |
A & B |
R |
S |
S |
R |
|
5 |
KSL |
SJC2305 |
17.8±1.1 |
7.3±0.5 |
R |
A & B |
R |
R |
R |
R |
|
6 |
KSL |
SJC2306 |
11.9±1.4 |
6.9±0.9 |
R |
A & B |
R |
R |
R |
R |
|
7 |
KSL |
SJC2307 |
0.7±0.2 |
9±0 |
R |
A & B |
R |
R |
R |
R |
|
8 |
KSL |
SJC2308 |
8.1±0.8 |
7.5±1.1 |
R |
A & B |
R |
R |
R |
R |
|
9 |
KSL |
SJC2309 |
17.2±6.8 |
6.9±0.7 |
R |
A & B |
S |
S |
R |
R |
|
10 |
KSL |
SJC2310 |
6.3±0.5 |
8.2±1 |
R |
A & B |
R |
R |
R |
R |
|
11 |
KSL |
SJC2311 |
5.7±0.6 |
8.7±0.5 |
R |
A & B |
R |
R |
R |
R |
|
12 |
KSL |
SJC2312 |
14.1±1.7 |
6.9±1.1 |
R |
A & B |
R |
R |
R |
R |
|
|
13 |
Breeding Line |
KSL |
SJC2313 |
Korea |
10.7±2.7 |
6.9±0.5 |
R |
A & B |
R |
R |
R |
R |
|
14 |
KSL |
SJC2314 |
26.3±3.2 |
5.1±2.2 |
MR |
A & B |
R |
R |
R |
R |
|
15 |
KSL |
SJC2315 |
38.7±3.5 |
4.6±0.6 |
MR |
B |
S |
S |
S |
R |
|
16 |
KSL |
SJC2316 |
48.1±4.2 |
3.9±0.9 |
MR |
A & B |
R |
R |
R |
R |
|
17 |
KSL |
SJC2317 |
30.7±1.1 |
4.7±0.9 |
MR |
A & B |
R |
R |
R |
R |
|
18 |
KCU |
SJC2318 |
68.4±4.1 |
4.9±0.9 |
MR |
- |
S |
S |
S |
S |
|
19 |
CHS |
SJC2319 |
33±3.9 |
6.3±1.6 |
MR |
A & B |
S |
R |
R |
R |
|
|
1 |
Breeding Line |
EUR |
SJC2320 |
Netherlands |
84.5±5.5 |
2.7±1.1 |
S |
A |
S |
R |
S |
S |
|
2 |
ASL |
SJC2321 |
USA |
97.6±2.8 |
1.2±0.5 |
S |
A |
R |
R |
R |
S |
|
3 |
THS |
SJC2322 |
Thailand |
73.6±3.9 |
3.3±0.7 |
S |
- |
S |
S |
S |
S |
|
4 |
PSL |
SJC2323 |
France |
15.6±0 |
7±0 |
R |
A & B |
R |
R |
R |
R |
|
5 |
THS |
SJC2324 |
Thailand |
11.8±1 |
7.5±0.5 |
R |
A & B |
S |
S |
R |
R |
|
6 |
PSL |
SJC2325 |
France |
13.6±5 |
7.9±0.4 |
R |
A & B |
R |
R |
R |
R |
|
7 |
TAS |
SJC2326 |
Taiwan |
14±1 |
7.8±0.7 |
R |
A & B |
R |
R |
R |
R |
|
8 |
TAS |
SJC2327 |
Taiwan |
0.8±0.2 |
8.9±0.3 |
R |
A & B |
R |
R |
R |
R |
|
9 |
CHS |
SJC2328 |
China |
48.3±4.7 |
6.4±1.6 |
MR |
A & B |
R |
R |
R |
R |
|
10 |
API |
SJC2329 |
USA |
63.2±3.5 |
5.6±1.4 |
MR |
A & B |
S |
S |
R |
R |
|
11 |
THS |
SJC2330 |
Thailand |
69.7±4.1 |
5.9±0.6 |
MR |
A |
R |
R |
R |
S |
|
12 |
THS |
SJC2331 |
Thailand |
63.2±3.7 |
4.1±1 |
MR |
- |
S |
S |
S |
S |
|
13 |
THS |
SJC2332 |
Thailand |
59±3.9 |
4.2±1.5 |
MR |
A & B |
S |
R |
S |
R |
|
14 |
ASL |
SJC2333 |
USA |
55.8±1.4 |
4.4±1 |
MR |
A & B |
R |
R |
R |
R |
|
|
1 |
Landrace |
Z |
Khira Balam |
India |
94.4±4.4 |
1.8±0.6 |
S |
- |
S |
S |
S |
S |
|
2 |
Z |
Abad Golfaigar; 266 |
Iran |
75±33.5 |
2.4±0.6 |
S |
A & B |
R |
S |
S |
R |
|
3 |
Z |
45 |
Iran |
92.8±3.4 |
2±0.5 |
S |
A & B |
S |
R |
S |
R |
|
4 |
Z |
KHIRA/salmon (PI 163217) |
India |
88±7 |
2.8±0.4 |
S |
A |
S |
R |
S |
S |
|
5 |
Z |
Kakri |
India |
83±3.5 |
3.2±1.9 |
S |
A |
R |
R |
R |
S |
|
6 |
Z |
11757 |
India |
80.8±2 |
2.6±1 |
S |
A |
S |
R |
S |
S |
|
7 |
CHS |
Kuai Huang Kua |
China |
92.6±4.7 |
1.8±1.1 |
S |
A & B |
R |
R |
R |
R |
|
8 |
Z |
Kuan chi chiao pai hasia chiu No. 1: 448 |
Japan |
91.2±4.2 |
1.8±0.8 |
S |
- |
S |
S |
S |
S |
|
9 |
Z |
Pepino verde comprido; 3868 |
Brazil |
94.7±1.9 |
1.3±0.5 |
S |
A |
S |
R |
S |
S |
|
10 |
KSL |
Seoul Madi |
Korea |
1.6±0.4 |
8.9±0.3 |
R |
A & B |
R |
R |
R |
R |
|
11 |
TAS |
Hei Pei Ta Kua |
Taiwan |
15.6±1.8 |
7.3±1.2 |
R |
A & B |
R |
R |
R |
R |
|
12 |
Z |
Concombre blanc; ABD 14 |
Mauritius (East Africa) |
6.6±0.8 |
6.8±1.2 |
R |
A |
R |
S |
S |
S |
|
|
13 |
Landrace |
Z |
11758 |
India |
27.1±2.9 |
6.6±1 |
R |
A |
S |
R |
S |
S |
|
14 |
Z |
UPL-Cu-2 (PI 426170) |
Philippines |
1.5±0.4 |
8.2±0.6 |
R |
A & B |
R |
R |
R |
R |
|
15 |
Z |
Tu pien chiao san chi |
Japan |
69.1±7.9 |
3.6±1 |
MR |
B |
S |
S |
S |
R |
|
16 |
Z |
Cool and Crisp |
Kenya |
39.1±8.5 |
5.1±1.3 |
MR |
A & B |
S |
R |
R |
R |
|
17 |
Z |
UPL-Cu-1 |
Philippines |
54.8±5.1 |
5.2±1.7 |
MR |
A & B |
R |
R |
R |
R |
|
18 |
Z |
PI 419017 |
China |
44.9±3.7 |
5.6±1.8 |
MR |
A & B |
R |
R |
R |
R |
|
19 |
CHS |
San dong si gua 22 |
China |
68.4±7.3 |
3.9±1.3 |
MR |
A & B |
R |
R |
R |
R |
|
20 |
Z |
Yomaki-asahi Aonaga;73 |
Japan |
58.3±11.3 |
4.4±1 |
MR |
A |
R |
R |
R |
S |
|
21 |
TAS |
TCMG |
China |
51±5.4 |
5.7±1.6 |
MR |
A |
R |
R |
R |
S |
Table 2Details on the markers associated with cucumber PM-resistance candidate genes
Table 2
|
Name of gene |
Gene ID
[Gy14 (v2.1)] |
Marker type |
Restriction enzyme |
Forward sequence
(5'~3') |
Reverse sequence
(5'~3') |
Susceptible vs. Resistance (bp) |
Reference |
|
CsABA2
|
CsGy5G015450
|
dCAPS |
NcoI |
GGTTATCGTCTCCAAGAAAGCCATG |
TCCGATACATCGACGTTGCG |
200 vs. 175 and 25 |
, Lee et al. (2020)
|
|
CsLRR-RPK2
|
CsGy5G015660
|
CAPS |
HinfI |
GCAACAAGTTCAATGGACCAC |
GAATCTCTCCAGTCAAATTGTTTCC |
224 and 100 vs. 324. |
, Zhang et al. (2021)
|
|
CsPM5.2
|
CsGy5G015960
|
dCAPS |
MnlI |
GGATTGAAATACTCGTTTGCGATCGCTCCT |
CCATCGGTTCTTTGAATGTCG |
183 vs. 152 and 31 |
, Sun et al. (2024)
|
|
CsaMLO8
|
CsGy5G026660
|
InDel |
- |
TATGGCTGCCTTTCATCTCCT |
TCCAAGCAAAGAAGGCAAGC |
405 vs. 1854 |
, Nie et al. (2015a)
|
References
- Alavilli H, Lee HY, Lee JJ, Pamidimarri DS, Reddy CK, Song K. 2024. Identification of a gene causing yellow endocarp in Cucumis sativus through high-throughput sequencing. Sci. Hortic. 323: 112515
- Alavilli H, Lee JJ, You CR, Poli Y, Kim HJ, Jain A, et al. 2022. GWAS reveals a novel candidate gene CmoAP2/ERF in pumpkin (Cucurbita moschata) involved in resistance to powdery mildew. Int. J. Mol. Sci. 23(12): 6524
- Badri Anarjan M, Bae I, Lee S. 2021. Marker-assisted evaluation of two powdery mildew resistance candidate genes in Korean cucumber inbred lines. Agronomy. 11(11): 2191
- Badri Anarjan M, Li H, Lee S. 2024. Current Status of Viral Resistance Molecular Breeding in Cucumbers (Cucumis sativus L.). In Allele Mining for Genomic Designing of Vegetable Crops. 1st ed. Taylor & Francis Group. CRC Press, England & Wales: pp. 105-131.
- Berg JA, Appiano M, Bijsterbosch G, Visser RG, Schouten HJ, Bai Y. 2017. Functional characterization of cucumber (Cucumis sativus L.) Clade V MLO genes. BMC Plant Biol. 17(1): 80
- Berg JA, Appiano M, Santillán Martínez M, Hermans FW, Vriezen WH, Visser RG, et al. 2015. A transposable element insertion in the susceptibility gene CsaMLO8 results in hypocotyl resistance to powdery mildew in cucumber. BMC Plant Biol. 15(1): 243
- Hosoya K, Narisawa K, Pitrat M, Ezura H. 1999. Race identification in powdery mildew (Sphaerotheca fuliginea) on melon (Cucumis melo) in Japan. Plant Breed. 118(3): 259-262.
- Innark P, Ratanachan T, Khanobdee C, Samipak S, Jantasuriyarat C. 2014. Downy mildew resistant/susceptible cucumber germplasm (Cucumis sativus L.) genetic diversity assessment using ISSR markers. Crop Prot. 60: 56-61.
- Kishor D, Lee HY, Alavilli H, You CR, Kim JG, Lee SY, et al. 2021. Identification of an allelic variant of the CsOr gene controlling fruit endocarp color in cucumber (Cucumis sativus L.) using genotyping-by-sequencing (GBS) and whole-genome sequencing. Front. Plant Sci. 12: 802864
- Lee HY, Kim JG, Kang BC, Song K. 2020. Assessment of the genetic diversity of the breeding lines and a genome wide association study of three horticultural traits using worldwide cucumber (Cucumis spp.) germplasm collection. Agronomy. 10(11): 1736
- Li S, Lin D, Zhang Y, Deng M, Chen Y, Lv B, et al. 2022. Genome-edited powdery mildew resistance in wheat without growth penalties. Nature. 602(7897): 455-460.
- Liu PN, Miao H, Lu WH, Cui JY, Tian GL, Wehner TC, et al. 2017. Molecular mapping and candidate gene analysis for resistance to powdery mildew in Cucumis sativus stem. Genet. Mol. Res. 16(3): 10-4238.
- Nie J, He H, Peng J, Yang X, Bie B, Zhao J, et al. 2015a. Identification and fine mapping of pm5.1: a recessive gene for powdery mildew resistance in cucumber (Cucumis sativus L. ). Mol. Breed. 35(1): 7
- Nie J, Wang Y, He H, Guo C, Zhu W, Pan J, et al. 2015b. Loss-of-function mutations in CsMLO1 confer durable powdery mildew resistance in cucumber (Cucumis sativus L.). Front. Plant Sci. 6: 1155
- Nie J, Yuan Q, Zhang W, Pan J. 2023. Genetics, resistance mechanism, and breeding of powdery mildew resistance in cucumbers (Cucumis sativus L.). Hortic. Plant J. 9(4): 603-615.
- Sakata Y, Kubo N, Morishita M, Kitadani E, Sugiyama M, Hirai M. 2006. QTL analysis of powdery mildew resistance in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 112(2): 243-250.
- Samba N, Nunomura O, Lu N, Johkan M, Nakano A, Tsukagoshi S. 2024. Cucumber (Cucumis sativus L.) growth and productivity under solar radiation-based quantitative nutrient management in hydroponic system. Agronomy. 14(2): 296
- Sun J, Nie J, Xiao T, Guo C, Lv D, Zhang K, et al. 2024. CsPM5.2, a phosphate transporter protein‐like gene, promotes powdery mildew resistance in cucumber. Plant J. 117(5): 1487-1502.
- Wang H, Jiang YP, Yu HJ, Xia XJ, Shi K, Zhou YH, et al. 2010. Light quality affects incidence of powdery mildew, expression of defence-related genes and associated metabolism in cucumber plants. Eur. J. Plant Pathol. 127(1): 125-135.
- Wang W, Wen Y, Berkey R, Xiao S. 2009. Specific targeting of the Arabidopsis resistance protein RPW8.2 to the interfacial membrane encasing the fungal haustorium renders broad-spectrum resistance to powdery mildew. Plant Cell. 21(9): 2898-2913.
- Xiao S, Ellwood S, Calis O, Patrick E, Li T, Coleman M, et al. 2001. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science. 291(5501): 118-120.
- Zhang C, Badri Anarjan M, Win KT, Begum S, Lee S. 2021. QTL-seq analysis of powdery mildew resistance in a Korean cucumber inbred line. Theor. Appl. Genet. 134(2): 435-451.
- Zhang D, Wu S, Li N, Gao J, Liu S, Zhu S, et al. 2022. Chemical induction of leaf senescence and powdery mildew resistance involves ethylene-mediated chlorophyll degradation and ROS metabolism in cucumber. Hortic. Res. 2(9): uhac101