Abstract
Salicylic acid (SA) is a plant hormone that has a significant role in many biochemical processes involved in cotton plant resistance against biotic and abiotic stress factors. Exogenous SA has been shown to have effects on plant growth and development, resistance to fungi and insects, and mitigation of abiotic stress factors. Treating cotton seedlings or plants with SA in a culture medium or spraying them with SA has enabled scientists to identify genes responsible for this chemical, associated with several biological functions. SA has been established as part of the defense system in cotton plants: antifungal resistance and insect resistance. Besides, it is a part of plant growth and development as well as fiber development. Identifying SA-responsive genes and understanding their roles in plant resistance enables the development of stress-resistant genotypes. This paper reviews scientific data resulting from the treatment of cotton plants with exogenous SA. In the first section, we discussed antifungal resistance-related data linked to SA treatment, which makes up the highest content of the review. We highlighted its significant role in cotton plant antifungal resistance. The second section deals with SA-responsive genes and metabolites linked with insect resistance. In the third section, we reviewed SA-responsive genes and enzymes associated with cotton plant growth and development. The scientific data regarding SA-linked cotton fiber development have been discussed in the last section. In summary, we discussed SA-responsive genes, proteins, and metabolites that could be used to develop genotypes with enhanced traits.
-
Key words: Salicylic acid, cotton plant, Gossypium hirsutum L, V. dahliae, antifungal resistance, salt stress
Introduction
The increasing global population and consumption of food resources necessitate the development of new approaches to enhance crop yields. However, water scarcity and soil degradation, caused by rising salt and alkaline levels, pose significant challenges to agriculture. Using exogenous chemicals like salicylic acid (SA), which is involved in diverse biological processes, enables the identification of genes that could be responsive to stress processes. Treating plants with exogenous substances is an efficient way to mitigate various biotic and abiotic stress factors. Phytohormones, such as SA, are promising candidates for this approach due to their involvement in plant biological processes at very low concentrations. SA, also termed
ortho-hydroxybenzoic acid, has been shown to alleviate the effects of various stresses, including salinity, temperature extremes, metal ions (
Koche 2021), fungi, and bacteria (
Hayat et al. 2010). SA was reported to regulate various stress responses by activating processes involved in oxidation-reduction balance, secondary metabolite modulation, and nutrient optimization (
Koche 2021). Studying these processes enables us to identify genes that could be effectively used to develop and breed cotton genotypes with enhanced traits.
SA is a plant hormone biosynthesized from cinnamate, a phenylalanine derivative (
Chen et al. 2009). It is one of the most investigated plant hormones with significant practical implications for economically important crops. It is accumulated in plant cells following the pathogen attack (
Kim et al. 2023) or abiotic stress factors (
Koche 2021).
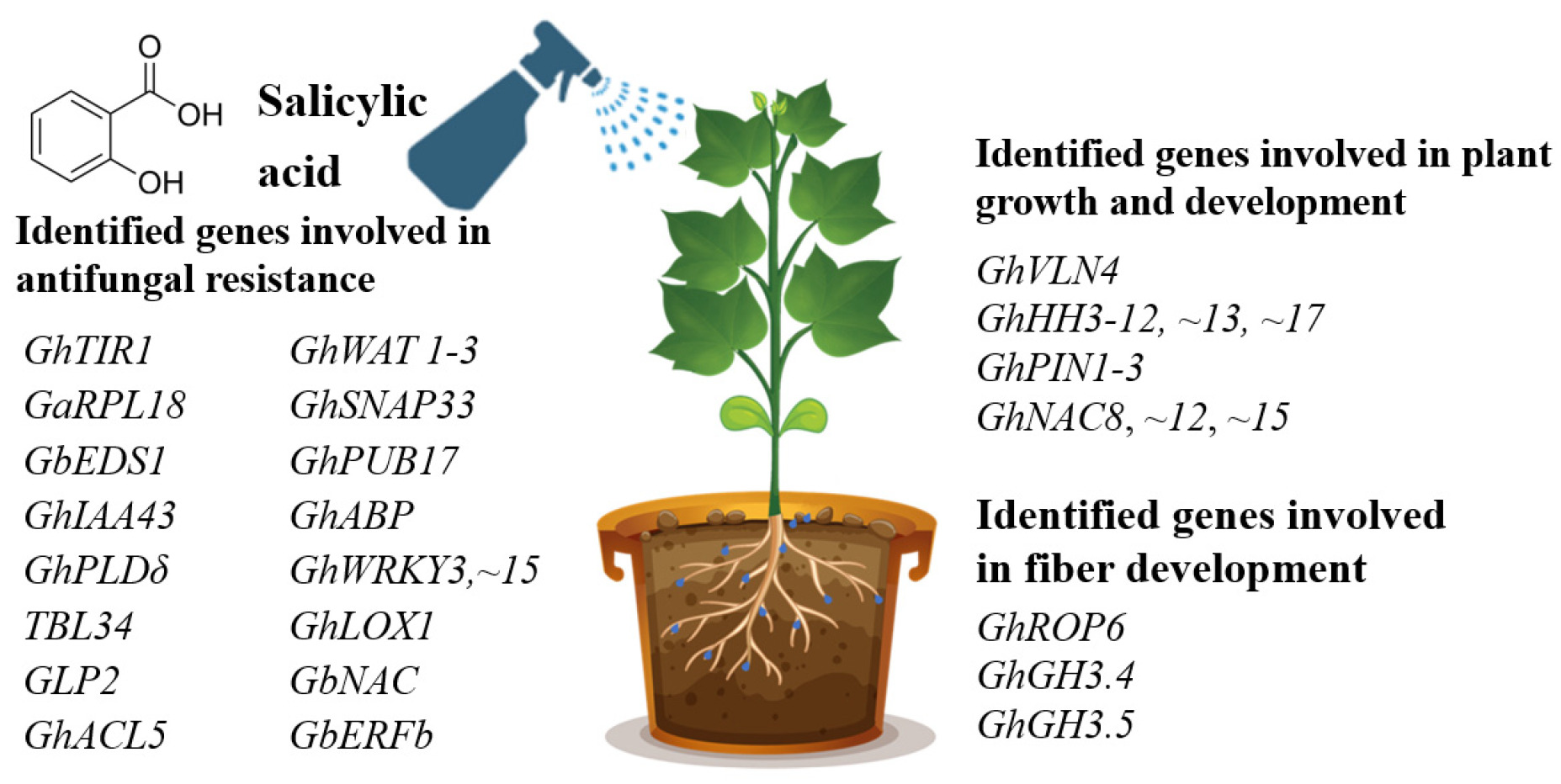
Our search on the effects of exogenous SA on cotton plant genes and proteins allowed finding papers related to three biological processes: antifungal resistance, plant growth and development, and fiber development. In most cases, studying the effects of SA on cotton plants enabled the authors to identify genes involved in antifungal resistance against both
V. dahliae and
F. oxysporum. More than fifteen SA-dependent genes were identified in the cotton plant that were linked with the antifungal resistance. Besides, eight genes were found to be involved in plant growth and development. Moreover, three genes linked with fiber development were also identified by SA treatment (
Fig. 1).
Below, we highlight the scientific data on the effects of exogenous SA on cotton plant genes, enzymes, and metabolites involved in plant resistance. We review the effects of exogenous SA on plant antifungal resistance, growth and development, and fiber development in separate sections.
Antifungal resistance
Cotton crop loss because of fungal diseases is a serious problem that needs to be mitigated. Developing genetically engineered resistant crops is a practical approach, the efficiency of which can be proven by studying the impact of various exogenous chemicals that play pivotal roles in plants. Utilizing exogenous chemicals to combat different stress factors can help us more deeply understand biological processes. SA is an economically feasible approach that could be used for research purposes to determine mechanisms to combat fungal diseases in cotton plants. In this section, we review the efficacy of exogenous SA for identifying cotton plant genes associated with antifungal resistance.
SA-related genes play essential roles in cotton plant resistance to
Verticillium dahliae wilt. The knockdown of
GhTIR1, a gene involved in auxin signaling, significantly reduced SA-related genes and enhanced susceptibility to
V. dahliae, demonstrating the significant role of SA in cotton resistance (
Shi et al. 2022). In addition, SA was found to upregulate ribosomal protein L18 in
G. arboretum (
GaRPL18), which mediated resistance to Verticillium wilt. The transformation of
GaRPL18 into Arabidopsis enhanced resistance to
V. dahliae, suggesting that this effect is mediated by the SA pathway (
Gong et al. 2017).
GbEDS1, a gene homologous to the Arabidopsis defense regulator EDS1 (Enhanced Defense Regulator), was also reported to be involved in cotton plant responses to Verticillium wilt and SA. Silencing of
GbEDS1, which is expressed in both the cytoplasm and nucleus, led to reduced SA and H
2O
2 levels and increased susceptibility to
V. dahliae in
G. barbadense. In contrast, overexpressing this gene in Arabidopsis led to induced susceptibility to
V. dahliae. Treatment with 10 mM SA significantly increased
GbEDS1 transcript levels in cotton, confirming that
GbEDS1 is SA-responsive (
Yan et al. 2016).
The essential role of SA-related defense in cotton resistance to
V. dahliae was proven in
GhIAA43-silenced cotton plants, as
GhIAA43 is suggested to be a negative regulator of this defense. The enhanced antifungal resistance in
GhIAA43-silenced cotton plants was explained by monitoring SA levels and SA marker gene expression (
Su et al. 2022). In
G. hirsutum, treating cotton plants with 1-aminocyclopropane-1-carboxylic acid (ACC) activated SA-dependent resistance, leading to enhanced resistance to
V. dahliae (
Jia et al. 2022). In another work, the authors associated the mechanistic action of SA in wilt resistance with an enhanced level of beta-1,3-glucanase localized in the callus. They demonstrated the significance of SA treatment in resistant and susceptible plants after treatment with
V. dahliae toxin + SA. Exogenous SA alleviated the damage resulting from
V. dahliae toxin (
Zhen et al. 2004). SA signaling is suggested to be aligned with JA signaling and reactive oxygen species (ROS) to generate cotton plant resistance to Verticillium infection (
Xu et al. 2014).
GhPLDδ, a phospholipase D (PLP) gene in
G. hirsutum, confers resistance to biotic and abiotic stresses, including Verticillium wilt resistance. It is highly induced by plant hormones, including SA. The accumulation of SA and JA is associated with genes involved in MAPK cascades (Zhu et al. 2022). One of the mechanisms regulated by SA to improve wilt resistance in
G. hirsutum is acetylation of the cell wall polysaccharides mediated by
TBL34 genes. These genes are upregulated by
V. dahliae and by exogenous plant phytohormones – SA, JA, and ethylene (
Zhao et al. 2021a).
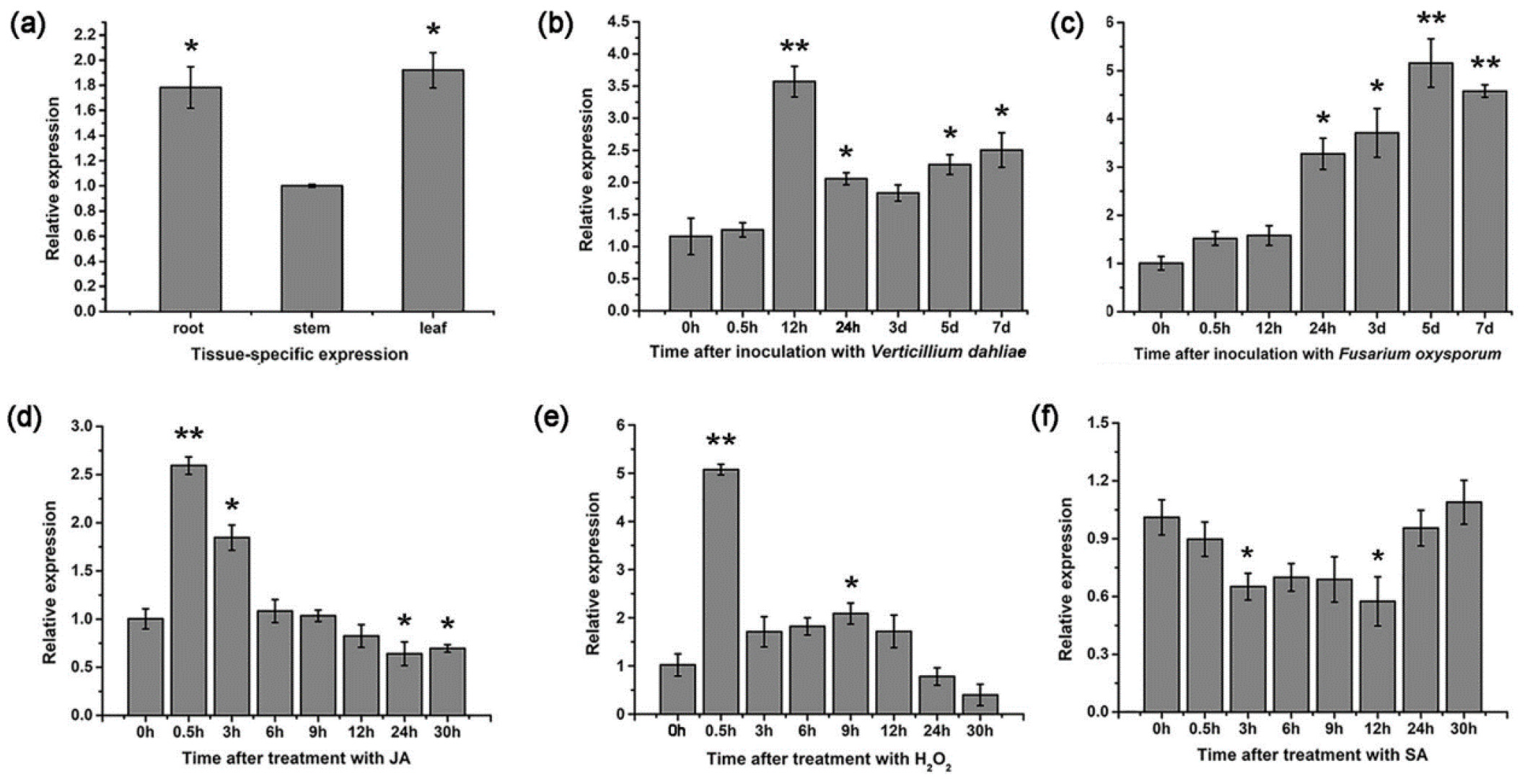
Germin-like proteins (GLPs) are glycoproteins that play a pivotal role in the antifungal resistance of cotton plant. These proteins were found to be efficient against both
V. dahliae and
F. oxysporum. Silencing the
GLP2 gene in cotton enhanced its susceptibility to these pathogens. Its expression was found to trigger several stress-related genes.
GLP2 expression was enhanced several times by these fungi in different time intervals. It was also found to increase several-fold by JA and H
2O
2 in 30 minutes. However, no significant increases were found by SA treatment for 30 hours. Significant differences were observed after 3 and 12 hours of the treatment, but not in their intermediates (
Pei et al. 2020) (
Fig. 2). Thus, the expression of the
GLP2 gene, found to be responsible for antifungal resistance resulting from treatment with
V. dahliae and
F. oxysporum, was established to be SA-dependent in a time-dependent manner.
GhACL5 is another gene linked with
V. dahliae resistance. Silencing
GhACL5 in cotton resulted in a dwarf phenotype. The expression of
GhACL5 in Pima 90-53, a
G. barbadense variety resistant to Verticillium wilt, was similar in response to
V. dahliae and SA treatments for 48 hours. Foliar application of 1 mM SA to two-week-old resistant plants resulted in several-fold enhancements in
GhACL5 gene expression levels for two days. Methyl jasmonate (MeJA) and ethylene treatments caused fluctuating
GhACL5 gene expression levels for the next 48 hours (
Mo et al. 2015).
So-called Walls Are Thin (WAT) genes identified in
G. hirsutum (
GhWAT 1-3) were found to be responsible for
V. dahliae resistance. Silencing all three of these genes, which are highly expressed in roots and internodes, upregulated SA-related genes. The expression of lignification-related genes was enhanced following the silencing of
GhWAT123 genes. Treatment with 2 mM SA in a culture solution reduced the expression of the
GhWAT1-3 for the next six days. Thus, 2 mM SA was found to reduce
GhWAT 1-3 genes and increase lignification-related genes (
Tang et al. 2019). SNAP25-type protein is a 25 kDa synaptosome-associated protein molecule that plays a role in exocytosis. It was suggested that exogenous SA controls SNAP25-type protein, leading to enhanced resistance to
V. dahliae. The
GhSNAP33 gene encodes a SNAP25-type protein and is induced by
V. dahliae and SA. Loss of
GhSNAP33 led to increased levels of ROS in leaves and cell death, while its overexpression improved resistance to
V. dahliae. Two-week-old seedlings kept in 1 mM SA solution showed several-fold increases in
GhSNAP33 gene expression levels. Thus, the role of SA in antifungal resistance was linked with
GhSNAP33 genes (
Wang et al. 2018).
Another mechanism of SA-mediated antifungal resistance is associated with the ubiquitin ligase GhPUB17, which is involved in the response to
V. dahliae.
GhPUB17 is significantly upregulated in cotton roots following exogenous SA treatment (
Qin et al. 2019). RING domain-containing E3 ubiquitin ligase genes were identified in four cotton species as responsive to
V. dahliae and to stress responses resulting from JA, SA, and ethylene. However, no correlation was found between these two traits. Some genes were responsive to
V. dahliae, while others responded to phytohormone treatments (
Zhao et al. 2021b).
GhABP, a novel germin-like protein gene in
G. hirsutum, is upregulated in response to
V. dahliae or JA treatment. However, SA treatment suppresses
GhABP expression. A 1 mM dose of SA added to the Hoagland medium reduced
GhABP expression levels several-fold in both resistant and susceptible cultivars. The opposite effect was observed with JA treatment (
Pei et al. 2019).
WRKY transcription factors play a role in plant defense, stress adaptation, and growth and development. The
GhWRKY3 gene encodes a 507-amino-acid protein that plays a significant role in plant defense.
GhWRKY3 gene expression was also enhanced when plants were infected with
Fusarium oxysporum f. sp.
vasinfectum,
Rhizoctonia solani, and
Colletotrichum gossypii. The authors suggested that
GhWRKY3 is responsible for SA-associated antifungal resistance in cotton plants (
Guo et al. 2011). SA involvement in cotton fungal resistance is associated with other defense-related genes. The nuclear gene
GhWRKY15 was reported as another gene responsive to fungal infection and SA treatment. Its expression is higher in roots and stems than in cotyledons. Higher fungal resistance resulting from
GhWRKY15 expression was associated with increased activity of peroxidase and ascorbate peroxidase, suggesting that ROS regulation is a key pathway in this resistance (
Yu et al. 2012).
Another study showed that treating plants with the
V. dahliae protein VdCP1 induced antifungal resistance associated with increased SA, ROS, and ion electrolyte leakage levels in infected plants (
Zhang et al. 2017). The
CG05 gene is also responsive to
V. dahliae and SA treatments in cotton. Endogenous SA influences
CG05 gene expression in Arabidopsis. The mechanism of action is thought to be controlled by GST, which regulates ROS using glutathione, which in turn affects SA content (
Gong et al. 2018). SA involvement in alleviating the effects of bacterial infection through ROS was also established. Hypersensitive reaction cell death in cotton caused by
Xanthomonas campestris pv.
malvacearum is associated with the activity of 9S-lipoxygenase, which is encoded by the
GhLOX1 gene. Sequence analysis revealed that the
GhLOX1 gene contains SA- and JA-responsive elements. Treatment of cotton cotyledons with these phytohormones induced both
GhLOX1 gene expression and enzyme activity (
Marmey et al. 2007).
NAC transcription factor genes in
G. barbadense (
GbNAC) were reported as another family of genes sensitive to treatments with SA as well as MeJA. After treatment with 1 mM SA, 6 of the 15 studied genes were upregulated. In four cases, the upregulation was observed at one hour, but the expression levels then decreased to their original levels. The transcriptomic analysis showed that these family genes are expressed in response to Verticillium wilt. The authors concluded that these candidate genes could be useful for improving antifungal resistance via the SA pathway (
Liu et al. 2019). A novel
AP2/ERF (ethylene-responsive factors) transcription factor gene, termed
GbERFb, was identified in
G. barbadense as responsive to several phytohormones, including SA, MeJA, ethylene, as well as treatment with
V. dahliae, H
2O
2, and wounding. Early responses in the gene expression levels by SA, MeJA, ethylene, and
V. dahliae were observed. The maximum expression levels for ethylene and MeJA were observed at 6 and 12 hours, respectively. SA and
V. dahliae treatments caused maximum responses at 3 hours (
Liu et al. 2017). These results enable concluding earlier responses triggered by SA pathways compared to JA.
Insect resistance
Crop loss resulting from insect attacks is another serious problem for cotton plants. Several studies have investigated the role of SA as an approach to mitigate this negative impact. In this section, we discuss scientific evidence on the role of exogenous SA in cotton plant insect resistance. In this section, we discuss scientific data devoted to the role of exogenous SA in cotton plant insect resistance.
One study found that treating Egyptian cotton with exogenous SA reduced levels of jasmonic acid (JA), which is expected to trigger resistance to Egyptian cotton worms (
Stotz et al. 2002). Another study concluded that SA was found not to promote controlling this chewing pest with that approach. However, the
cry1EC gene was found to be effective in controlling Spodoptera. SA treatment also induced the expression of the synthetic
cry1EC gene in cotton.
Spodoptera litura larvae fed on CaMV35S(r)PR-1a
cry1EC-expressing transgenic cotton could not survive. Induction of transcription by SA (2 mM) and wound showed similar results for 20 hours, with several-fold increases in transgenic cotton leaves (
Kumar et al. 2009). In another work, the involvement of SA in resistance against the
Spodoptera exigua beet armyworm was investigated, and changes in SA levels in local and systemic leaves grown in normal and nitrogen-fertilized soils in response to beet armyworm feeding were measured. However, in most cases, no significant differences were determined in SA levels between nitrogen-fertilized and nitrogen-unfertilized samples following larval feeding (
Chen et al. 2008).
SA was also associated with plant resistance to cotton bollworms. Field experiments revealed that treatment with SA, cotton bollworm, or cotton bollworm plus larval injury enhanced levels of volatiles, which are known antifeedants. This establishes the involvement of SA in plant resistance against this insect (
Yu et al. 2007). Another economically significant pest of cotton plants is the solenopsis mealybug
Phenacoccus solenopsis. Research on controlling this pest with exogenous SA or JA demonstrated that females were repelled by leaves treated with JA but not SA. This phenomenon was explained by the JA-regulated production of gossypol. Thus, treatment with exogenous SA was found to be ineffective in controlling the solenopsis mealybug (
Zhang et al. 2011). The contents of gossypol precursors and metabolites vary widely in cotton plants. In general, these compounds, which are biosynthesized from ocimene and myrcene, are expected to be major antifeedants. However, their effectiveness against various insects requires further analysis.
Another study found that treating cotton plants with 5 mM SA enhanced plant resistance to cotton aphids. Both aphid and SA treatments upregulated the callose synthase
GhCalS5 genes, whose overexpression increased resistance to this sucking pest. The expression of
GhCalS5 was enhanced almost 5-fold following treatment with SA at 24 hours, but no significant changes were observed at 48 and 72 hours after treatment (
Mbiza et al. 2022).
Overall, exogenous SA has been found to be effective against both chewing and sucking pests, but some research has shown that SA is ineffective against certain chewing pests. More research is needed to understand the efficacy of SA against chewing insects more properly.
Plant Growth and Development
Plant growth and development is a complex process characterized by many interrelated functions. The role of SA in cotton plant growth and development has been well-established, particularly under abiotic stress conditions. In this section, we review the role of SA in cotton plant growth and development in association with genes and enzymes.
SA can improve plant growth and development under salt stress by optimizing photosynthesis and the metabolism of antioxidative compounds. This is due to the enhanced activity of the enzymes peroxidase, catalase, and superoxide dismutase (SOD) (
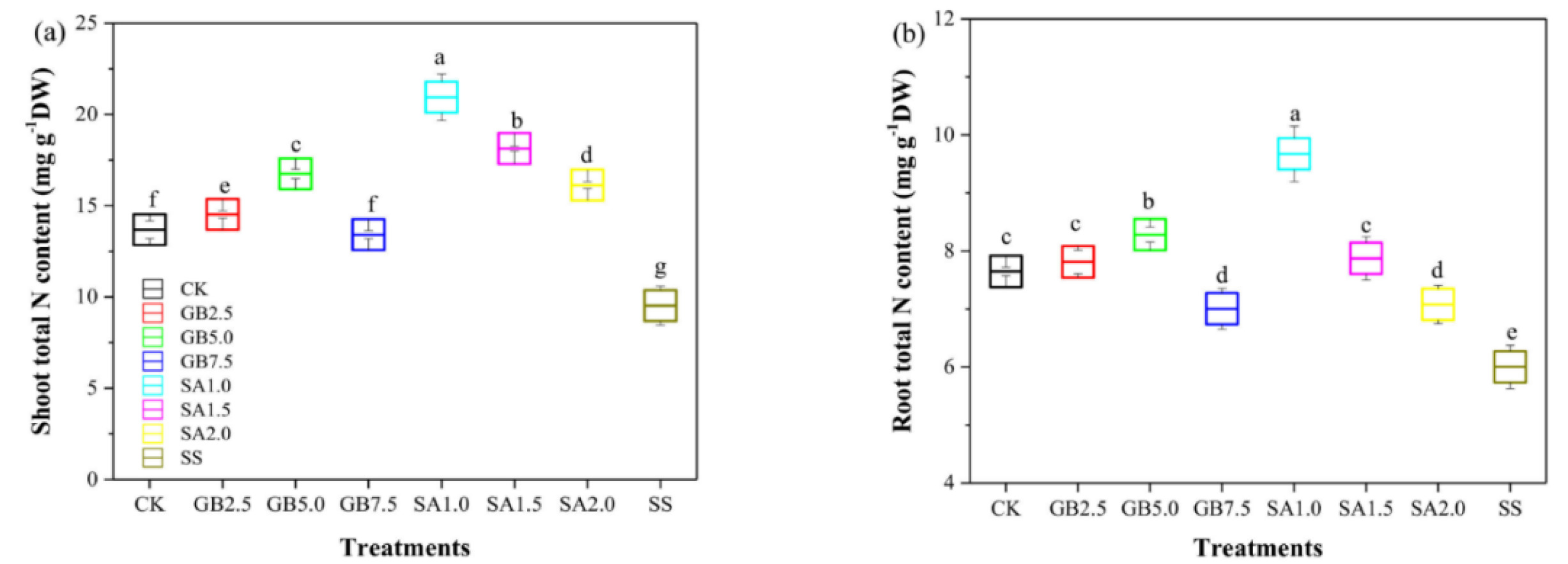
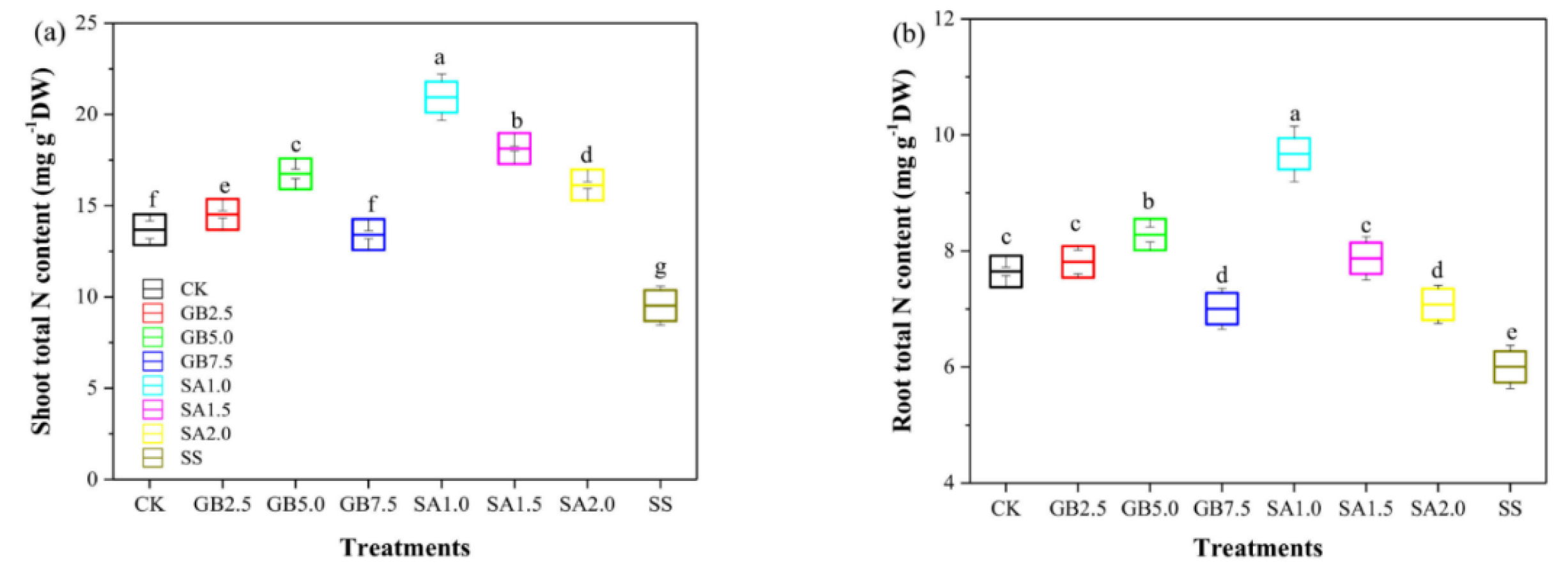
Wang et al. 2022). In one work, cotton seedlings exposed to salt stress and treated with 1 mM SA or 5 mM glycine betaine revealed improvements. The biomass accumulation was enhanced with the highest nitrogen content in roots and shoots following the 1 mM SA treatment. The treatments with 1.5 mM and 2 mM SA caused significantly lower nitrogen content in both root and shoots. The authors also reported a negative correlation between SA and sodium content in plant biomass (
Hamani et al. 2021) (
Fig. 3). These results imply finding correlations between SA metabolites involved in plant growth and development.
Exogenous SA application was found to promote cotton plant growth. In one work, treating cotton seedlings under salt stress with 1 mM SA enhanced chlorophyll content and promoted gas exchange. A positive correlation was found between photosynthetic rate and chlorophyll b content after treatment with SA (
Hamani et al. 2020). In another work, SA promoted root growth in cotton plants. Adding 0.010 mM SA to culture substrates significantly increased root growth in two of three cotton varieties. Three doses of SA (0.01, 0.05, and 0.1 mM) significantly enhanced SOD activity in roots in two cotton varieties. Two doses (0.1 and 0.05 mM) were established as optimal for the three cotton varieties. These results imply identifying SA-responsive genes using this approach for various cotton varieties (
Cao et al. 2022).
The
GhVLN4 gene, which encodes an actin-binding protein, was found to be responsible for microfilament bundling, which is important for plant growth. The expression of this gene was upregulated in response to treatment with
V. dahliae, as well as with exogenous plant hormones SA, JA, ethylene, and abscisic acid. A 2.5 mM dose of SA significantly enhanced gene expression levels for 3 hours but caused no significant changes at 6, 12, and 48 hours after treatment. The highest responses were observed with 0.1 mM MeJA (
Ge et al. 2021).
Three
GhHH3 genes, including
GHH3-12,
GHH3-13, and
GHH3-17, were identified as responsive to SA treatment (10 µM) in cotton plants. Compared to the effects of other phytohormones, including gibberellic acid (GA), IAA, and MeJA, several-fold increases in gene expression levels were observed within the first 3 hours, indicating a strong effect of this hormone. All of these genes lack introns and possess highly conserved protein motifs, suggesting that they are involved in growth and development, as well as stress responses (
Qanmber et al. 2019).
GhPIN1-3 and
GhPIN2 are two PIN-FORMED efflux carrier gene family members that are responsive to SA treatment and involved in plant growth. Treatment with a 0.5 mM SA spray caused the upregulation of most
PIN genes in
G. hirsutum (
He et al. 2017). Transgenic cotton plants in which endochitinase from
Trichoderma virens was expressed were reported to be conditionally associated with lesion mimic phenotype (LMP) in cotton seedlings when grown in a nutrient-deficient medium. SA treatment at 0.1 and 0.5 mM doses induced the expression of LMP and was concluded as the causative agent of LMP (
Kumar et al. 2012).
NAC family genes and proteins play important roles in diverse aspects of plant development as well as stress regulation. The expression of stress-responsive
GhNAC8-
GhNAC17 genes was linked with leaf senescence. The maximum expression of eight of the ten
GhNAC genes was observed in senescent leaves. Among these eight genes,
GhNAC8,
GhNAC12, and
GhNAC15 showed higher expression in cotton seedlings following treatment with 2 mM SA. In other cases, no significant changes were detected (
Shah et al. 2013). The SA signaling pathway, which was established in Arabidopsis (
Rivas-San Vicente et al. 2011), was also confirmed in cotton plants (
Shah et al. 2013).
Fiber development
A few studies have investigated the role of SA in fiber development. One study reports the relevance of
Rho-related GTPase6 (
GhROP6) expression in fiber development. The authors examined the effects of SA (0.5, 5, and 50 mM), JA (0.001, 0.01, and 0.1 mM), and GA (0.5, 5, and 50 mM) added to ovule culture. All three hormones caused slightly lower β-glucuronidase activity
in vitro at their highest doses (
Li et al. 2022). In another work, a genome-wide analysis of
GH3 gene family members identified auxin-induced genes that could be involved in fiber development. Adding 0.01 mM SA to a hydroponic culture solution caused various changes in the expression of
GhGH3 subfamily II genes. In total, 6 of 11 genes studied were suppressed in the stem following SA treatment, while more genes were upregulated in the root. Two genes,
GhGH3.4 and
GhGH3.5, were upregulated in the root and stem following treatment with all three plant hormones (SA, JA, and GA). The authors suggested that these genes are candidates for developing cotton fiber (
Yu et al. 2018).
Conclusion
The scientific data discussed above on the role of exogenous SA in cotton plant resistance and growth/development proves its involvement in a variety of biological processes. Its pivotal role can be linked with antifungal resistance in cotton plants. The involvement of SA in insect resistance of cotton plants deserves further investigation. Some studies claim the correlation between SA and insect resistance against chewing insects, while others conclude that SA has no role. SA has been involved in cotton plant growth and development through several approaches. It has been found to attenuate the toxic effects of salt stress or V. dahliae. Recent data suggest that SA may also play a role in fiber development, but further analysis is needed to confirm this claim. Both foliar spray of SA and its addition to growth culture media have been shown to be effective for plant growth and development, and could be efficiently used to identify SA-responsive genes and proteins. Increased nitrogen uptake, reduced sodium uptake, and enhanced photosynthetic parameters all demonstrate the effectiveness of this chemical as a plant growth regulator.
Acknowledgments
We acknowledge the financial support from the Ministry of Innovative Development of the Republic of Uzbekistan. We thank the research team from the Center of Genomics and Bioinformatics.
Fig. 1Genes identified in cotton plant by the treatment with exogenous SA
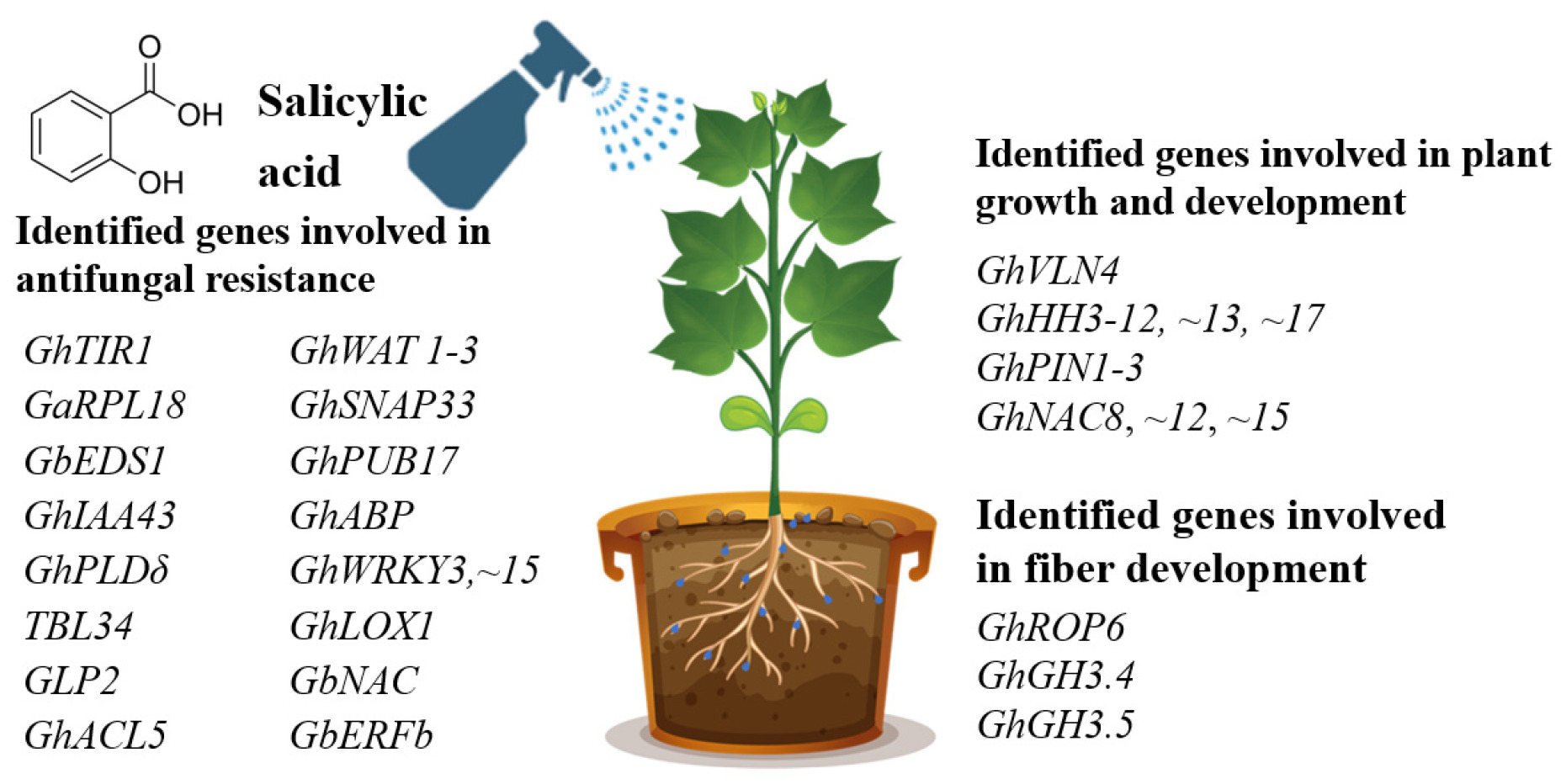
Fig. 2The expression levels of
GLP2 genes in three different cotton tissues (a), and following the treatments with
V. dahliae (b) and
F. oxysporum (c), with JA (d), H
2O
2 (e), and SA (f). The figure has been re-used from (
Pei et al. 2020) with the permission of Springer Nature.
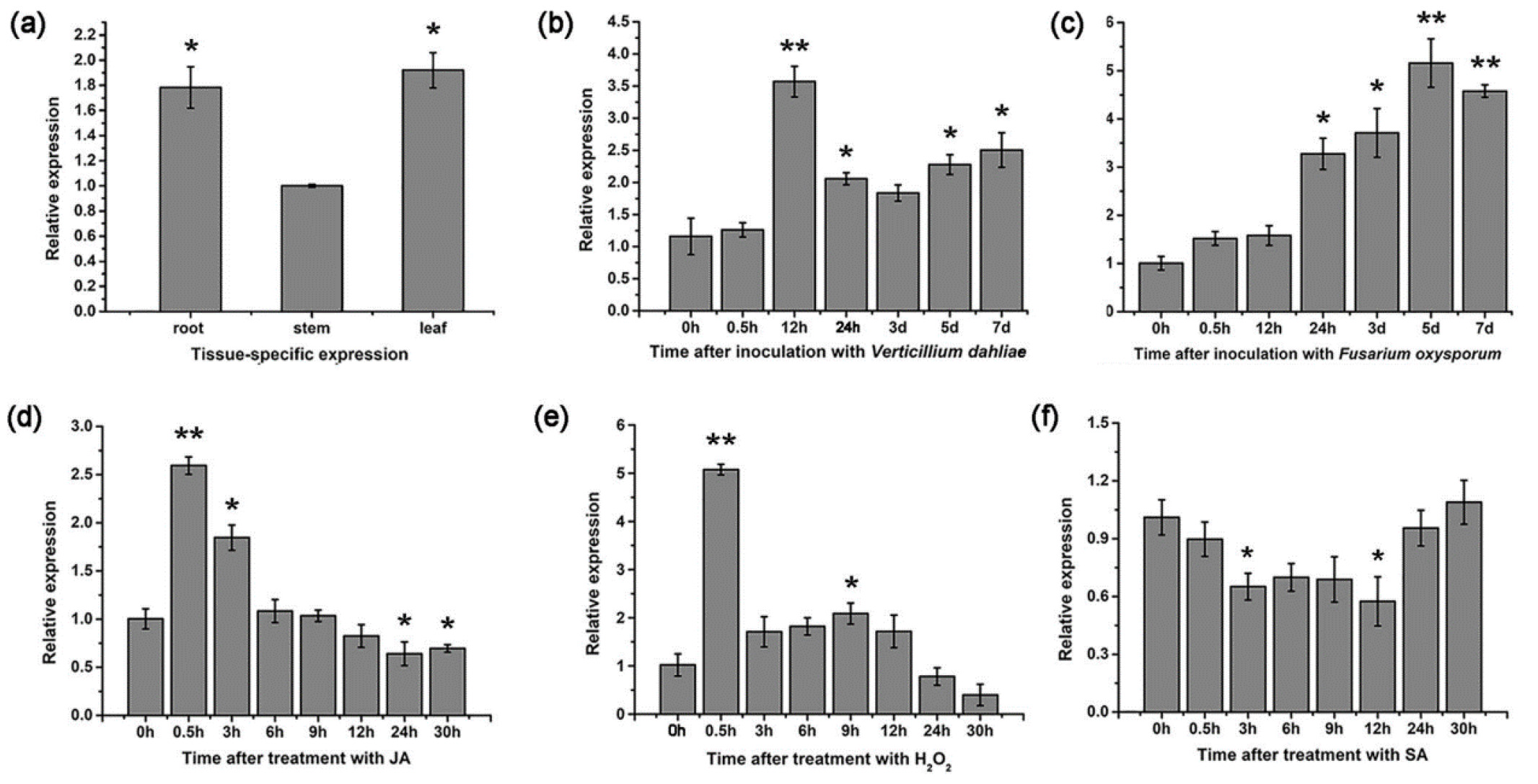
Fig. 3Effects of various doses of SA and glycine betaine (GB) on the nitrogen content of roots and shoots. a) Effects of various treatments on shoot nitrogen content. b) Effects of various treatments on root nitrogen content. CK was used for control. GB2.5, GB5.0, and GB7.5 show treatments with respective GB doses. Similarly, SA1.0, SA1.5, and SA2.0 stand for treatments with respective doses of SA. SS means salt stress (150 mM) without supplementation. The figure has been re-used from (
Hamani et al. 2021) with the permission of MDPI.
References
- Cao Z, Wang X, Gao Y. 2022. Effect of Plant Growth Regulators on Cotton Seedling Root Growth Parameters and Enzyme Activity. Plants (Basel, Switzerland). 11(21): 2964
- Chen Y, Schmelz EA, Wäckers F, Ruberson JR. 2008. Cotton plant, Gossypium hirsutum L., defense in response to nitrogen fertilization. J. Chem. Ecol. 34(12): 1553-64.
- Chen Z, Zheng Z, Huang J, Lai Z, Fan B. 2009. Biosynthesis of salicylic acid in plants. Plant Signal. Behav. 4(6): 493-6.
- Ge D, Pan T, Zhang P, Wang L, Zhang J, Zhang Z, et al. 2021. GhVLN4 is involved in multiple stress responses and required for resistance to Verticillium wilt. Plant Sci. 302: 110629
- Gong Q, Yang Z, Chen E, Sun G, He S, Butt HI, et al. 2018. A Phi-Class Glutathione S-Transferase Gene for Verticillium Wilt Resistance in Gossypium arboreum Identified in a Genome-Wide Association Study. P. C. P. 59(2): 275-289.
- Gong Q, Yang Z, Wang X, Butt HI, Chen E, He S, et al. 2017. Salicylic acid-related cotton (Gossypium arboreum) ribosomal protein GaRPL18 contributes to resistance to Verticillium dahliae. BMC Plant Biol. 17(1): 59
- Guo R, Yu F, Gao Z, An H, Cao X, Guo X. 2011. GhWRKY3, a novel cotton (Gossypium hirsutum L.) WRKY gene, is involved in diverse stress responses. Mol. Biol. Rep. 38(1): 49-58.
- Hamani AKM, Chen J, Soothar MK, Wang G, Shen X, Gao Y, et al. 2021. Application of Exogenous Protectants Mitigates Salt-Induced Na(+) Toxicity and Sustains Cotton (Gossypium hirsutum L.) Seedling Growth: Comparison of Glycine Betaine and Salicylic Acid. Plants (Basel, Switzerland). 10(2): 380
- Hamani AKM, Wang G, Soothar MK, Shen X, Gao Y, Qiu R, et al. 2020. Responses of leaf gas exchange attributes, photosynthetic pigments and antioxidant enzymes in NaCl-stressed cotton (Gossypium hirsutum L.) seedlings to exogenous glycine betaine and salicylic acid. BMC Plant Biol. 20(1): 434
- Hayat Q, Hayat S, Irfan M, Ahmad A. 2010. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 68(1): 14-25.
- He P, Zhao P, Wang L, Zhang Y, Wang X, Xiao H, et al. 2017. The PIN gene family in cotton (Gossypium hirsutum): genome-wide identification and gene expression analyses during root development and abiotic stress responses. BMC Genomics. 18(1): 507
- Jia MZ, Li ZF, Han S, Wang S, Jiang J. 2022. Effect of 1-aminocyclopropane-1-carboxylic acid accumulation on Verticillium dahliae infection of upland cotton. BMC Plant Biol. 22(1): 386
- Kim TJ, Lim GH. 2023. Salicylic Acid and Mobile Regulators of Systemic Immunity in Plants: Transport and Metabolism. Plants (Basel, Switzerland). 12(5): 1013
- Koche D, Gandhi R, Rathod S, Shirsat R. 2021. An update on role of salicylic Acid (SA) in abiotic stress tolerance in crop plants: a review. AG. BI. R. 37(6): 219-26.
- Kumar M, Shukla AK, Singh H, Tuli R. 2009. Development of insect resistant transgenic cotton lines expressing cry1EC gene from an insect bite and wound inducible promoter. J. Biotechnol. 140(3-4): 143-8.
- Kumar V, Parkhi V, Joshi SG, Christensen S, Jayaprakasha GK, Patil BS, et al. 2012. A novel, conditional, lesion mimic phenotype in cotton cotyledons due to the expression of an endochitinase gene from Trichoderma virens. Plant Sci. 183: 86-95.
- Li B, Zhang L, Xi J, Hou L, Fu X, Pei Y, et al. 2022. An Unexpected Regulatory Sequence from Rho-Related GTPase6 Confers Fiber-Specific Expression in Upland Cotton. Int. J. Mol. Sci. 23(3): 1087
- Liu J, Wang Y, Zhao G, Zhao J, Du H, He X, et al. 2017. A novel Gossypium barbadense ERF transcription factor, GbERFb, regulation host response and resistance to Verticillium dahliae in tobacco. P. M. B. P. 23(1): 125-134.
- Liu Z, Fu M, Li H, Chen Y, Wang L, Liu R. 2019. Systematic analysis of NAC transcription factors in Gossypium barbadense uncovers their roles in response to Verticillium wilt. PeerJ. 7: e7995
- Marmey P, Jalloul A, Alhamdia M, Assigbetse K, Cacas JL, Voloudakis AE, et al. 2007. The 9-lipoxygenase GhLOX1 gene is associated with the hypersensitive reaction of cotton Gossypium hirsutum to Xanthomonas campestris pv malvacearum. P. P. B. 45(8): 596-606.
- Mbiza NIT, Hu Z, Zhang H, Zhang Y, Luo X, Wang Y, et al. 2022. GhCalS5 is involved in cotton response to aphid attack through mediating callose formation. Front. Plant Sci. 13: 892630
- Mo H, Wang X, Zhang Y, Yang J, Ma Z. 2015. Cotton ACAULIS5 is involved in stem elongation and the plant defense response to Verticillium dahliae through thermospermine alteration. Plant Cell Rep. 34(11): 1975-85.
- Pei Y, Li X, Zhu Y, Ge X, Sun Y, Liu N, et al. 2019. GhABP19, a Novel Germin-Like Protein From Gossypium hirsutum, Plays an Important Role in the Regulation of Resistance to Verticillium and Fusarium Wilt Pathogens. Front. Plant Sci. 10: 583
- Pei Y, Zhu Y, Jia Y, Ge X, Li X, Li F, et al. 2020. Molecular evidence for the involvement of cotton GhGLP2, in enhanced resistance to Verticillium and Fusarium Wilts and oxidative stress. Sci. Rep. 10(1): 12510
- Qanmber G, Ali F, Lu L, Mo H, Ma S, Wang Z, et al. 2019. Identification of Histone H3 (HH3) Genes in Gossypium hirsutum Revealed Diverse Expression During Ovule Development and nStress Resposes. Genes. 10(5): 355
- Qin T, Liu S, Zhang Z, Sun L, He X, Lindsey K, et al. 2019. GhCyP3 improves the resistance of cotton to Verticillium dahliae by inhibiting the E3 ubiquitin ligase activity of GhPUB17. Plant Mol. Biol. 99(4-5): 379-393.
- Rivas-San Vicente M, Plasencia J. 2011. Salicylic acid beyond defence: its role in plant growth and development. J. Exp. Bot. 62(10): 3321-3338.
- Shah ST, Pang C, Fan S, Song M, Arain S, Yu S. 2013. Isolation and expression profiling of GhNAC transcription factor genes in cotton (Gossypium hirsutum L.) during leaf senescence and in response to stresses. Gene. 531(2): 220-34.
- Shi G, Wang S, Wang P, Zhan J, Tang Y, Zhao G, et al. 2022. Cotton miR393-TIR1 Module Regulates Plant Defense Against Verticillium dahliae via Auxin Perception and Signaling. Front. Plant Sci. 13: 888703
- Stotz HU, Koch T, Biedermann A, Weniger K, Boland W, Mitchell-Olds T. 2002. Evidence for regulation of resistance in Arabidopsis to Egyptian cotton worm by salicylic and jasmonic acid signaling pathways. Planta. 214(4): 648-52.
- Su Y, Wang G, Huang Z, Hu L, Fu T, Wang X. 2022. Silencing GhIAA43, a member of cotton AUX/IAA genes, enhances wilt resistance via activation of salicylic acid-mediated defenses. Plant Sci. 314: 111126
- Tang Y, Zhang Z, Lei Y, Hu G, Liu J, Hao M, et al. 2019. Cotton WATs Modulate SA Biosynthesis and Local Lignin Deposition Participating in Plant Resistance Against Verticillium dahliae. Front. Plant Sci. 10: 526
- Wang P, Sun Y, Pei Y, Li X, Zhang X, Li F, et al. 2018. GhSNAP33, a t-SNARE Protein From Gossypium hirsutum, Mediates Resistance to Verticillium dahliae Infection and Tolerance to Drought Stress. Front. Plant Sci. 9: 896
- Wang Z, Dong S, Teng K, Chang Z, Zhang X. 2022. Exogenous Salicylic Acid Optimizes Photosynthesis, Antioxidant Metabolism, and Gene Expression in Perennial Ryegrass Subjected to Salt Stress. Agron. 12(8): 1920
- Xu L, Zhang W, He X, Liu M, Zhang K, Shaban M, et al. 2014. Functional characterization of cotton genes responsive to Verticillium dahliae through bioinformatics and reverse genetics strategies. J. Exp. Bot. 65(22): 6679-92.
- Yan Z, Xingfen W, Wei R, Jun Y, Zhiying M. 2016. Island Cotton Enhanced Disease Susceptibility 1 Gene Encoding a Lipase-Like Protein Plays a Crucial Role in Response to Verticillium dahliae by Regulating the SA Level and H(2)O(2) Accumulation. Front. Plant Sci. 7: 1830
- Yu D, Qanmber G, Lu L, Wang L, Li J, Yang Z, et al. 2018. Genome-wide analysis of cotton GH3 subfamily II reveals functional divergence in fiber development, hormone response and plant architecture. BMC Plant Biol. 18(1): 350
- Yu F, Huaxia Y, Lu W, Wu C, Cao X, Guo X. 2012. GhWRKY15, a member of the WRKY transcription factor family identified from cotton (Gossypium hirsutum L.), is involved in disease resistance and plant development. BMC Plant Biol. 12: 144
- Yu HL, Zhang YJ, Pan WL, Guo YY, Gao XW. 2007. Identification of volatiles from field cotton plant under different induction treatments. J. Appl. Ecol. 18(4): 859-64.
- Zhang P, Zhu X, Huang F, Liu Y, Zhang J, Lu Y, et al. 2011. Suppression of jasmonic acid-dependent defense in cotton plant by the mealybug Phenacoccus solenopsis. PLoS One. 6(7): e22378
- Zhang Y, Gao Y, Liang Y, Dong Y, Yang X, Yuan J, et al. 2017. The Verticillium dahliae SnodProt1-Like Protein VdCP1 Contributes to Virulence and Triggers the Plant Immune System. Front. Plant Sci. 8: 1880
- Zhao Y, Jing H, Zhao P, Chen W, Li X, Sang X, et al. 2021a. GhTBL34 Is Associated with Verticillium Wilt Resistance in Cotton. Int. J. Mol. Sci. 22(17): 9115
- Zhao YP, Shen JL, Li WJ, Wu N, Chen C, Hou YX. 2021b. Evolutionary and Characteristic Analysis of RING-DUF1117 E3 Ubiquitin Ligase Genes in Gossypium Discerning the Role of GhRDUF4D in Verticillium dahliae Resistance. Biomol. 11(8): 1145
- Zhen XH, Li YZ. 2004. Ultrastructural changes and location of beta-1, 3-glucanase in resistant and susceptible cotton callus cells in response to treatment with toxin of Verticillium dahliae and salicylic acid. J. Plant Physiol. 161(12): 1367-77.