Abstract
Increasing the productivity of Colocasia esculenta var. Antiquorum is very necessary to enhance the mass of tubers. This can be achieved through polyploid mutation method using colchicine to improve tuber size by increasing the number of chromosomes. Therefore, this study aimed to determine the effect of colchicine on morphology and polyploid changes of Colocasia esculenta var. Antiquorum at various concentrations and duration of immersion In vitro. The analysis was carried out as a completely randomized factorial design, with the first factor being colchicine concentration, consisting of 0.0%, 0.05%, 0.075%, and 0.1%. The second factor was the immersion time which consisted of 8 and 16 hours. The results showed that the best combination of concentration and immersion time on somatic morphology was 0.075% colchicine with 8 hours and 16 hours of immersion. Meanwhile, the best combination of chromosomal changes was 0.05% colchicine with 16 hours of immersion. These results indicated that the treatment of 0.05% colchicine concentration with a soaking time of 16 hours can be recommended for induction of polyploid mutations in Colocasia esculenta.
-
Key words: Colocasia esculenta, Polyploid mutation, Colchicine, Diploid, Tetraploid, In vitro
INTRODUCTION
Colocasia esculenta var. Antiquorum is a type of taro with a small tuber size (small corm taro), also known as Japanese taro (
Maretta et al. 2016). The plant exhibits a high level of resistance to weather changes, which allows it to adapt to different seasons. Based on the data from the Food Security Service of South Sulawesi Province, the productivity of
Colocasia esculenta var. Antiquorum in 2020 was 7.214 tons. However, the exports of
Colocasia esculenta L. tubers only amount to 300 tons/ha due to the low productivity of the plant (
Nagano et al. 2016).
Increasing the productivity of
Colocasia esculenta var. Antiquorum can be done through plant breeding by induc-tion of polyploid mutations. Polyploid induction aims to increase crop productivity because polyploid (tetraploid) plants have a more significant figure, fruit size, tuber, and flower than diploid plants (
Suryo 2007) and are also resis-tant to specific environmental stresses (
Deng et al. 2012).
Polyploid can be carried out by giving chemical mutagens such as colchicine to plant meristem tissue. Colchicine changes the number of plant chromosomes and causes gene mutations on the scale of seeds and vegeta-tively propagated plants. It also induces polyploid by inhibiting the formation of spindle fibers during cell division. However, the number of chromosomes increases, but cell division does not occur, thereby producing poly-ploid cells with a doubled number of chromosomes (
Manzoor et al. 2018,
2019; Ade
et al. 2010).
Colchicine is an important mutagen that prevents micro-tubule formation and doubles the number of chromosomes. It is commonly used to develop polyploid plants and func-tions as a mitotic toxin, which produces multiple plant muta-genic effects (El-Nashar
et al. 2015). Colchicine not only helps in doubling chromosomes but also causes plant muta-tions, resulting in the formation of Colchis-mutants (Ari
et al. 2015). The concentration of colchicine for seed treatment usually ranges from 0.1% to 0.8%, but high doses cause malformations and reduce the production of tetra-ploid plants. Therefore, it is recommended to use colchi-cine with the lowest possible concentration (
Pirkoohi et al. 2011).
The success of polyploid induction depends on the explant, plant species, colchicine concentration, and dura-tion of exposure. However, induction must be carried out carefully by plant breeders as excessively high concentra-tions cause problems in the form of abnormalities in developing seedlings (
Manzoor et al. 2018,
2019). Several studies on polyploid induction using colchicine have been successful with different concentrations, such as Papaver bracteatum L. (
Esfahani et al. 2020), Allium Sativum L. (
Ayu et al. 2019), Colchicum chalcedonicum Azn. (
Karlik et al. 2020), Neolamarckia cadamba (
Eng et al. 2021), Dionaea muscipula Ellis (
Jala 2014), Nigella sativa (
Gupta et al. 2021), and Rhododendron fortunei L. (
Mo et al. 2020). This shows that polyploid induction can be used to create or select better offspring for further use.
Based on the description above, it is necessary to in-vestigate induction of polyploid mutations in Colocasia esculenta var. Antiquorum. Therefore, this study aims to determine the effect of colchicine on morphology and chromosome polyploid of Colocasia esculenta var. Anti-quorum plantlets on the interaction of concentration and immersion time In vitro. The novelty of this study is that it is the first to explore induction of polyploid mutations in Colocasia esculenta var. Antiquorum.
MATERIALS AND METHODS
Experimental design
This study was performed at the Tissue Culture Labo-ratory, Department of Agricultural Cultivation, Faculty of Agriculture, Hasanuddin University, Makassar, South Sulawesi, 90245, Indonesia. The analysis employed a 2-factor factorial (F2F) experiment with a Completely Randomized Design (CRD) as the environmental design, with three replications in each of the eight treatments. The combination of treatment between colchicine concentra-tion c0 = 0%, c1 = 0.05%, c2 = 0.075%, c3 = 0.1%) and immersion time (t1 = 8 hours; t2 = 16 hours). Each replication consisted of three shoots, resulting in a total of 82 individual shoots, as the experimental unit observed.
Experimental conditional
The plantlets used as material for polyploid induction were eight weeks old Colocasia esculenta var. Antiquorum shoots, which were selected as explants. The in vitro shoots were removed and the petioles were cut to a size of 1-3 cm. The immersion was carried out in laminar and the equip-ment used was sterile. The shoots were immersed in dif-ferent concentrations of colchicine solution, namely 0% (sterile distilled water) (c0), 0.05% (c1), 0.075% (c2), and 0.1% (c3). Each shoot was immersed in a culture bottle containing 15 mL of colchicine solution and shaken at 100 rpm on a shaker for 8 hours (c1) and 16 hours (c2). After immersion, the explants were washed with sterile distilled water 3-4 times and planted into shoot propagation medium, namely MS media with 2 mg L‒1 BAP and 1 mg L‒1IBA. The medium contained sugar (30 g L‒1), with the pH being adjusted to 5.8, and compacted with 7 g L‒1 agar. The shoots were kept in an incubation room at a temperature of 25-26℃ with 16 hours/day irradiation and an irradiation inten-sity of ± 1,000-2,000 lux (Modification of Dwiyani 2015). Subsequently, the shoots were incu-bated for 16 Weeks After Planting (WAP) and observations were made every week from outside the culture bottle for 16 WAP.
Observation of parameters
Several parameters observed from shoot explants were the percentage of living shoots (%), the number of living leaves, the number of roots and the number of compound shoots.
Percentage of living explants =
Chromosome analysis with flow cytometry
Polyploid level analysis was performed using flow cytometry. For each treatment, samples from the best plantlet vigor were collected from the leaves of the treated plants, measuring ± 25 mm
2. The samples were ground using a razor blade in a petri dish with 0.2 mL of extraction buffer (solution A from Partec Kit) and incubated for 10 minutes at room temperature. Subsequently, the superna-tants from the samples were filtered using nylon kesh measuring 30 m and placed in a cuvette, which was added to 1 mL of DAPI solution. The samples were incubated again for 1 minute and inserted into the Flow cytometry tool. The tool read the light intensity of the measured sample through the graph displayed to show the number of ploidy changes that occurred (
Escobedo-Gracia-Medrano et al. 2018).
The data analysis was carried out using the F test to determine the interaction between colchicine concentra-tion and immersion time. When the variance obtained had a significant effect, further analysis was conducted using the Duncan’s Multiple Range Test (DMRT) at a 5% significance level to determine the effect of differences between treatments. Data processing was carried out using the Statistical Tool for Agricultural Research (STAR) program and Microsoft Office Excel 2019. The data were analyzed by correlation analysis to determine the relation-ship between the parameters (
Anshori et al. 2022).
RESULTS
Percentage of living shoots of Colocasia esculenta var Antiuorum in vitro
The results of the living shoots in
Table 1 and
Fig. 1 showed that the control shoots had a very high percentage of survival compared to the combination of concentration and immersion time. In this study, there were differences in the number of shoots planted due to contamination of the media before treatment. Therefore, out of the 18 propa-gated shoots, there was a variation in the number of shoots that grew before colchicine immersion treatment. How-ever, this can be corrected through the percentage of living shoots, which were analyzed via graphs.
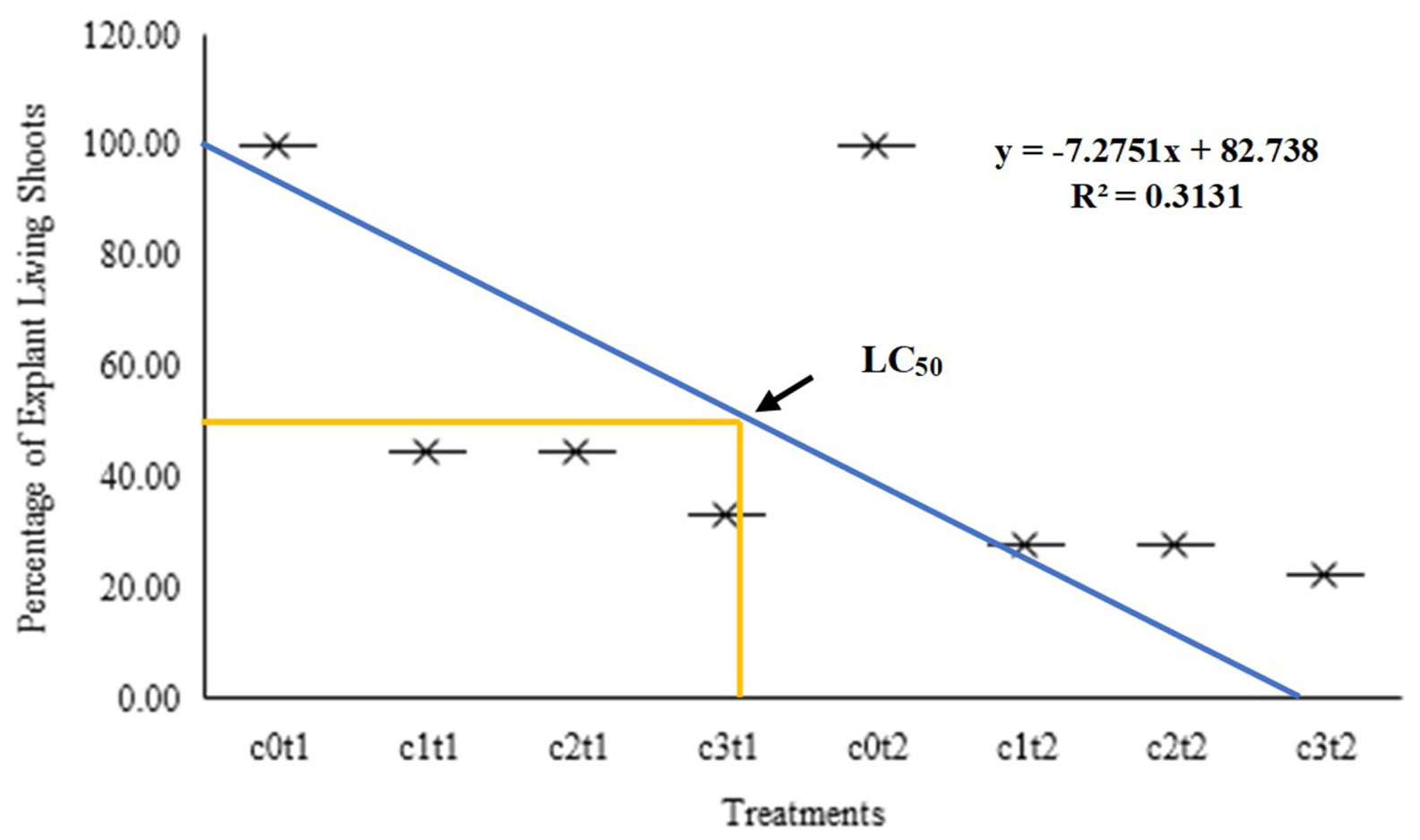
Based on the graph of living shoot percentage, this study showed a negative graph with an increase in colchicine concentration and time of immersion. The graph formed a linear regression with the formula Y = 7.2751x + 82.378, and the coefficient of determination (R2) was 0.3131, as presented in
Fig. 2. Based on this formula, the lethal 50 was in 0.1% colchicine concentration with 8 hours of immersion. The relationship between the percentage of surviving shoot explants was inversely proportional to the concentration and duration of colchicine immersion. As the concentration and duration of immersion of colchicine increased, the number of explants that survived decreased. However, the survival plantlet in the chemical mutants had unique trait changes, making it necessary to achieve a balance between the plantlet regeneration and optimum diversity in chemi-cal mutation induction. Therefore, the lethal 50 served as a practical point to meet both of these objectives.
The analysis of the mean square of variance in polyploid mutation induction was presented in
Table 2. The results showed that the treatment duration of colchicine immer-sion in
Colocasia esculenta var. Antiquorum and concen-tration levels significantly affected all observation parame-ters. The interaction effect between concentration level (c) and immersion time (t) also significantly affected the number of leaves, roots, and shoots, as well as the coef-ficient of diversity in the medium category.
The results of the DMRT test (a = 0.05) in
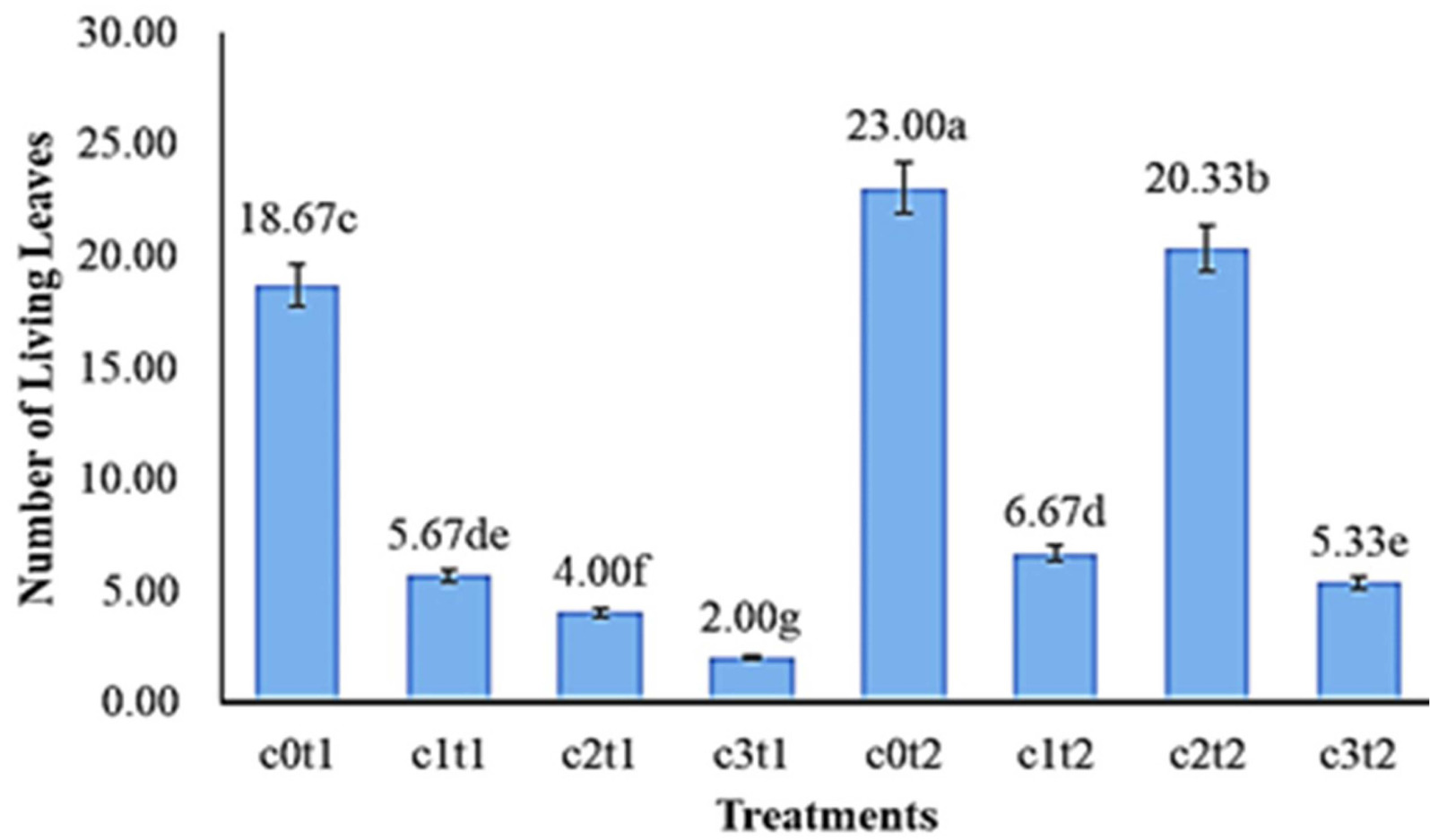
Fig. 3 showed the number of living leaves of
Colocasia esculenta var. Antiquorum in different treatments. The best treatment was c0t2 with 23.00 leaves, compared to c2t2, c1t2, and c3t2 with 20.33, 6.67, and 5.33 leaves, respectively. The lowest mean number of living leaves of
Colocasia esculenta var Antiquorum in the treatment c3t1 (2.00 leaves) was significantly different from c2t1 (4.00 leaves), c1t1 (5.67 leaves), c0t1 (18.67 leaves).
The results of the DMRT test (a = 0.05) in
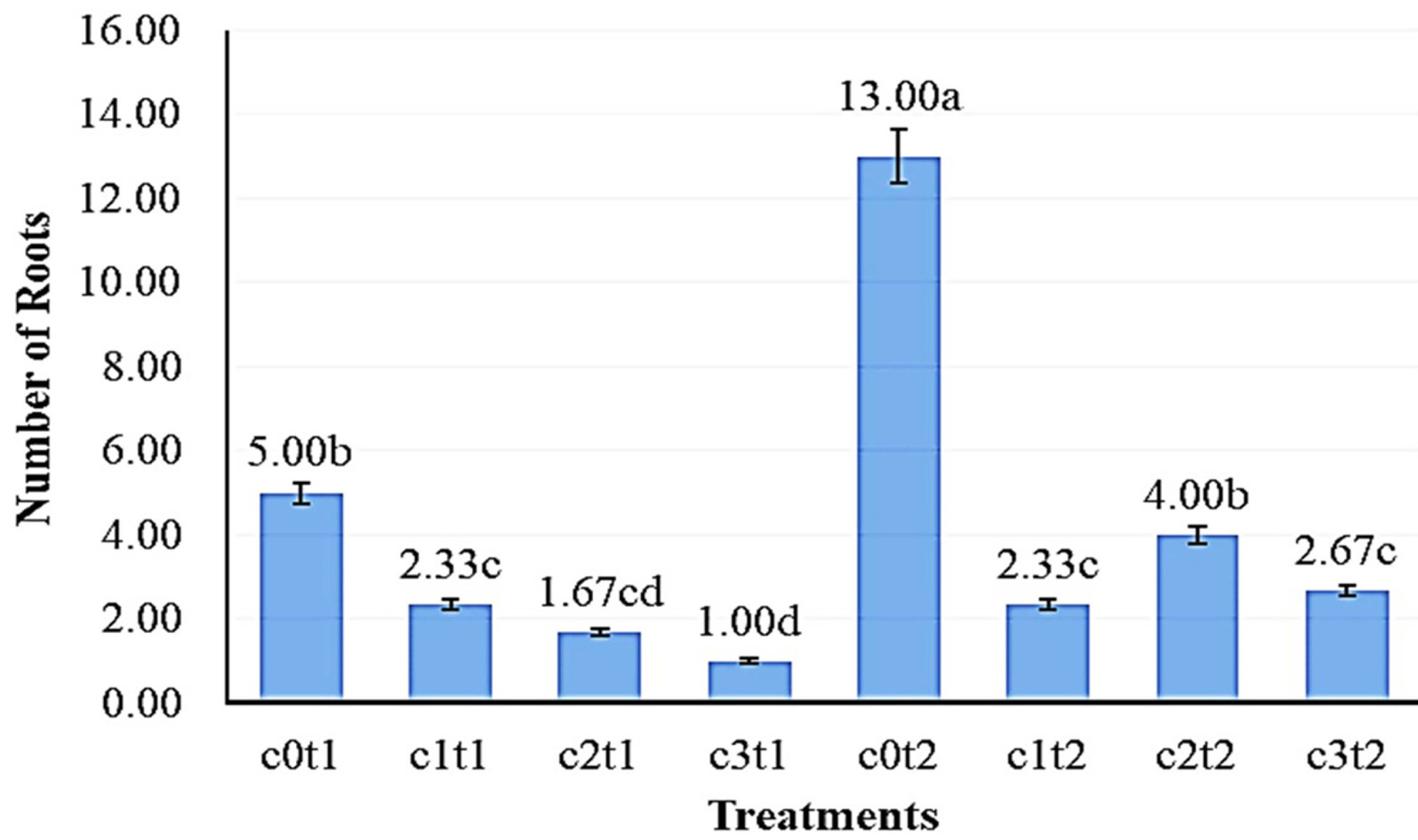
Fig. 4 showed the number of roots of the best
Colocasia esculenta var Antiquorum in the treatment c0t2 with 13.00 roots were significantly different from c2t2, c3t2, and c1t2 with 4.00, 2.67, and 2.33 leaves, respectively. The lowest mean number of roots of
Colocasia esculenta var Antiquorum in the treatment c3t1 with 1.00 roots was significantly different from the 5.00 roots of c0t1 but not from c1t1 and c2t1 at 2.33 and 1.67 roots, respectively.
The results of the DMRT test (a = 0.05) in
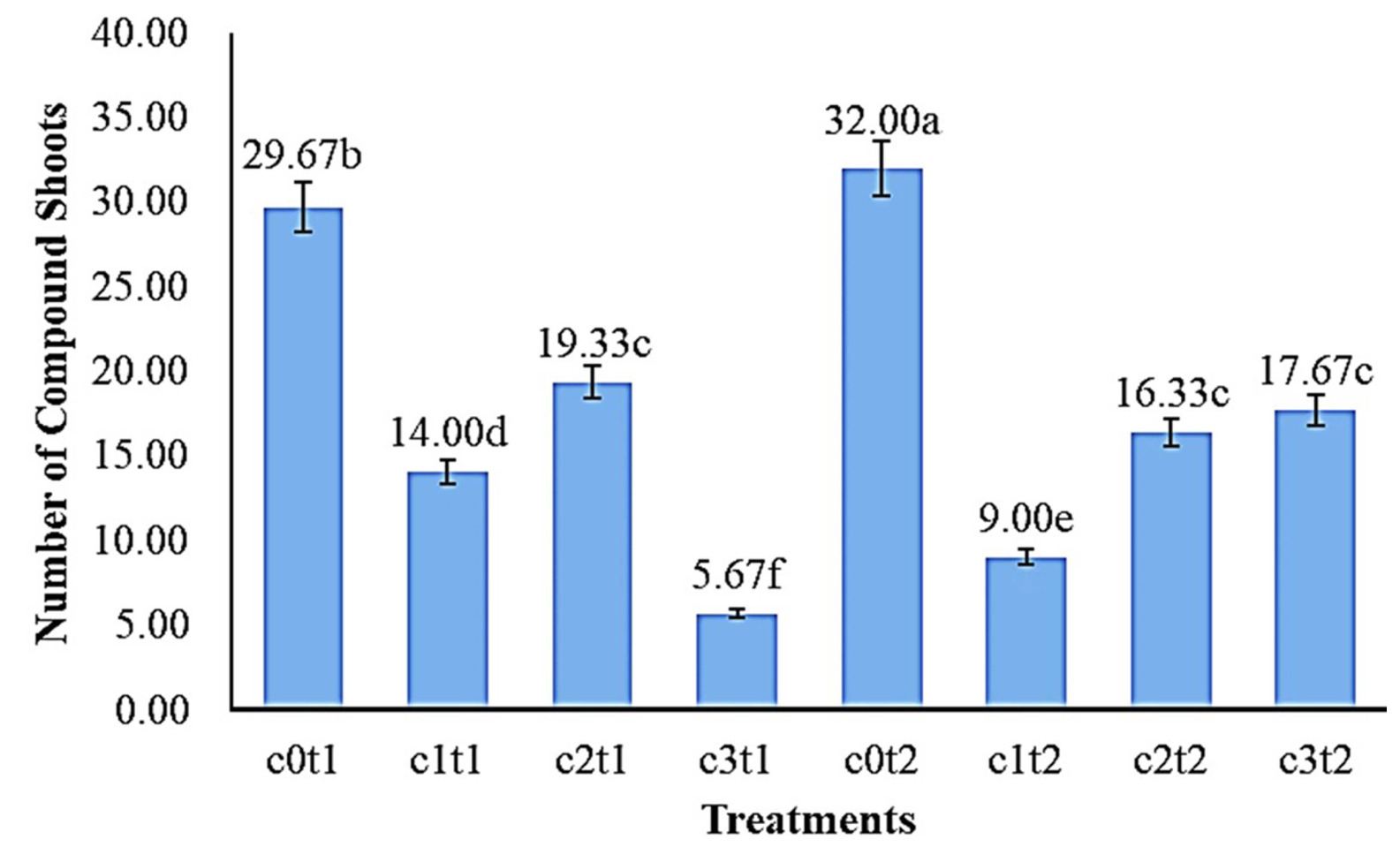
Fig. 5 showed the number of the best
Colocasia esculenta var. Antiquo-rum compound shoots in the treatment c0t2 with 32.00 shoots were significantly different from c3t2 (17.67 shoots), c2t2 (16.33 shoots), c1t2 (9.00 shoots), but c3t2 (17.67 shoots) was not different from c2t2 (16.33 shoots). The lowest average number of shoots of
Colocasia esculenta var. Antiquorum in the treatment c3t1 with 5.67 shoots was significantly different from c1t1, c2t1, and c0t1 with 14.00, 19.33, and 29.67 shoots, respectively.
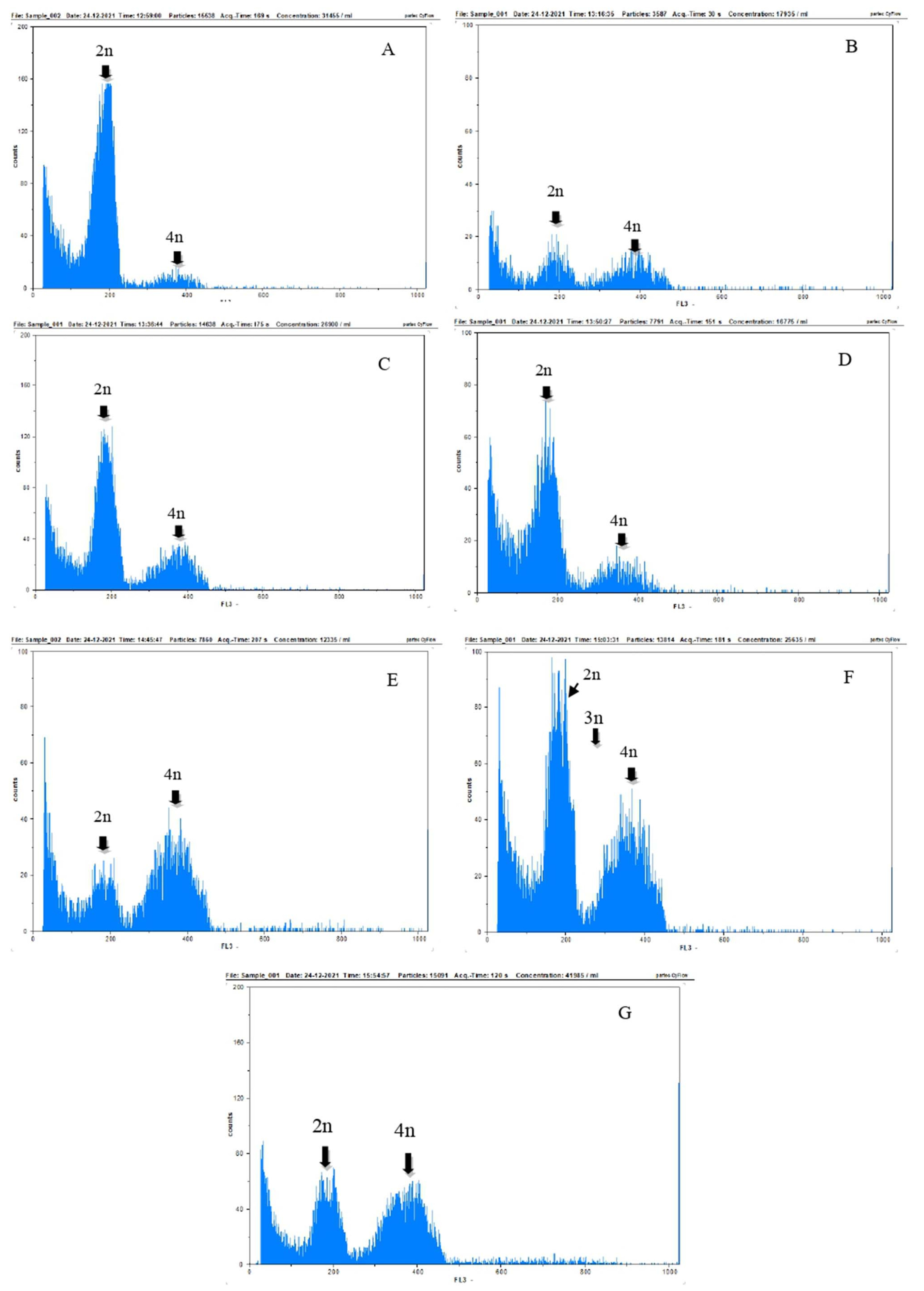
Polyploid analysis using flow cytometry on
Colocasia esculenta var. Antiquorum in
Table 3 showed that the difference in concentration and duration of colchicine immersion affected plant chromosomes. The treatments, namely c0t1, c1t1, c2t1, c3t1, c0t2, c1t2, and c3t1b showed an increase in the chromosomes from 2n-4n = 24 (diploid) to 48 (tetraploid). In the c2t2 treatment (0.075% colchicine, immersion 16 hours) the detected chromosomes ranged from 2n-3n-4n = 24 (diploid), 36 (triploid) to 48 (tetrap-loid).
Based on the percentage of gated chromosomes in the treatment c0t1 and c0t2, the dominant chromosome was 2n (diploid) with a percentage of 67.70%, which was signi-ficantly different from the 4n chromosome (tetraploid) at 4.17%. Therefore, it can be concluded that the chromo-somes in the treatments were diploid. In the c1t1 treatment (0.05% colchicine, immersion 8 hours), the dominant chromosome was 4n (tetraploid) with a percentage of 36.97% but not significantly different from 2n (diploid) at 28.35%. This showed that the chromosome was putative tetraploid because it allowed ploidy selection to occur. In c2t1 and c3t1, the dominant chromosomes were 2n (dip-loid) with a percentage of 52.94% and 55.26%, different from 4n (tetraploid) at 24.99% and 16.25%, which was mixoploid. In c1t2 and c3t2, the dominant chromosome was 4n (tetraploid) with a percentage of 54.10% and 44.36%, significantly different from 2n (diploid) at 20.27% and 28.24%, which did not allow ploidy selection to occur, namely tetraploid. In c2t2 treatment (0.075% colchicine, immersion 16 hours), the dominant chromosome was 4n (tetraploid) at 35.50% but not significantly different from chromosome 2n (diploid) at 29.94% and 3n (triploid) at 15.39%, making it putative-tetraploid because it allowed ploidy selection.
The results of the flow cytometry analysis in
Fig. 6 showed the chromosome peaks in
Colocasia esculenta var. Antiquorum plants in all treatments, which were interp-reted as standards for diploid (2n), tetraploid (4n), and triploid cells (3n) were also seen. Although all seven treat-ments exhibited diploid (2n), triploid (3n), and tetraploid (4n) cells, the peak height of each cell was different among treatments.
The results of the correlation analysis in
Table 4 showed the relationship between the number of leaf parameters, which can be evaluated
In vitro, in polyploid of
Colocasia esculenta var. Antiquorum. The number of leaves was positively correlated with the number of roots and com-pound shoots at 0.80 and 0.75, respectively.
DISCUSSION
Polyploid is a plant breeding technique carried out using mutagens such as colchicine to obtain plants with a chromosome number of more than 2n (diploid). Colchicine is one of the chemicals known to induce somaclonal variation in polyploidization in cultured plant cells. In plants, colchicine binds to tubulin, one of the main con-stituents of microtubules, which inhibits the development of spindle fibers and further mitotic division, leading to induction of polyploid (
Kharde et al. 2017). It has also been used for several years to induce polyploid, which increases the potential for metabolite production and facilitates the generation of valuable compounds in plants (
Bernard et al. 2012).
Flow cytometry analysis is a fast and efficient modern method for determining the ploidy level in almost all plant tissue regeneration and is particularly suitable for studies involving large sample sizes (
De et al. 2010;
Eeckhaut et al. 2005). For example,
Jones et al. (2008) and
De et al. (2004) used flow cytometry to determine the ploidy level and genome size of 200 diverse species and Rhododendron cultivars, creating a valuable database for breeders. This method has also been used to determine the ploidy level of induced polyploid adventitious shoots from three Rhodo-dendron hybrids.
The results of flow cytometry analysis in
Table 3 and
Fig. 6 showed that the number of chromosomes in the plant cell of
Colocasia esculenta var. Antiquorum doubled to tetraploid (4n). This indicated that the plant can be induced using polyploid by treating with colchicine concentration and immersion time, with the multiplication of chromo-somes varied in each treatment. Visual observations of plantlets in
Fig. 1 confirmed the doubling of chromosomes in
Colocasia esculenta var. Antiquorum plant. The increase in leaf size and petiole was observed in the c1t2 treatment (0.05% colchicine, immersion 16 hours), with an improve-ment in the size of shoots formed. This result was supported by
Mahyuni et al. (2015), who stated that diploid plants had a smaller morphology size than polyploid plants.
This study showed that the doubling of the chromosomes occurred randomly, leading to a non-uniform effect on an individual cell. The best chromosomal doubling occurred at low concentrations, namely 0.05% colchicine with 16 hours immersion time. This led to a significantly higher percentage of 54.10% compared to 44.36% and 35.50% at 0.1% and 0.075% colchicine concentration, respectively. Similarly, at 8 hours of immersion, more tetraploid chro-mosomes were formed at a low concentration of 0.05% by 36.97%, indicating different mutations in the same plant.
Heo et al. (2016) also concluded that the most prolonged immersion with low colchicine concentrations induced the highest percentage of polyploid production efficiency among other treatments on L. leichtlinii plants. In orchid plants, higher concentrations of colchicine produced shor-ter pods, with some dry and dead pods due to colchicine toxicity at high concentrations (1000 and 2000 mg/L) (
Sudirman et al. 2021).
Table 3 and
Fig. 6 showed that the doubling of the chromosomes formed were diploid, mixoploid, and tetra-ploid. Mixoploid plants had diploid and tetraploid nuclei, assessed based on the relative number of nuclei (
Cimen 2020;
Koutoulis et al. 2005). In polyploid induction, a high percentage of mixoploid yield was a procedure drawback because the unstable polyploid state often returned wholly or partially to diploid state after successive cell division cycles (
Esfahani et al. 2020). Furthermore, developing two different cells in a mixoploid bud caused competition during its growth. According to
Kainth and Grosser (2010), diploid cells had a faster division rate than autotetraploid cells, allowing the resulting mixoploid shoots to become diploid again.
Tetraploid are polyploid containing four sets of chromo-somes that can produce more secondary metabolites and biomass compared to diploid and are also drought-tolerant and disease-resistant (
Sattler et al. 2016;
Xu et al. 2014). Tetraploid can be artificially induced using mitotic spindle inhibitors such as colchicine (
Wang et al. 2016), with their plants having giant leaf and shoot sizes than diploid (
Kurtz et al. 2020).
Wulandari et al. (2015) conclude that 0.1% colchicine treatment for 1-hour is the most efficient in producing tetraploid Pumelo Nambangan citrus plants.
Asif (2001) and Tesfaye (2005) reported that chromo-somes were doubled in Musa balbisiana, various Musa acuminata subspecies, and Ensete ventricosum species using seeds, respectively. In polyploid induction, colchi-cine was found to be very efficient (Ganga 2002) de-pending on the variable amount of media, antimitotic agent, type of explant, exposure time, and concentration. Flow cytometry was the pre-eminent method used to induce evaluation polyploidization, while other methods such as chromosome number and morphology observa-tions had also been applied.
Urwin (2014) reported that a concentration of 0.1% and 6 hours of immersion can duplicate chromosomes in Lavandins plants (
Lavandula ×
intermedia cvs. Grosso and Seal). According to
Sadhukhan et al. (2014), colchicine at a concentration of 0.0005% with 6 hours of immersion duplicated chromosomes in African marigold (
Tagetes erecta) plants. The method was carried out by placing the entire plant with roots in colchicine solution and applying it to the apical shoots on cotton soaked in colchicine.
In this study, different concentrations of colchicine treat-ment and immersion time were found to affect morphology characteristics of regenerated plants under
In vitro condi-tions. According to
Jala (2014), the effect of colchicine concentration and duration of time for immersion in D. muscipula can be observed in changes in morphology and growth rate, survival rate, and variations that occurred at high concentrations of colchicine and immersion time.
The results of the DMRT analysis in
Figs. 3-
5 showed differences in the number of living leaves, roots, and compound shoots. The graph indicated that the highest data were obtained in the c0t2 treatment (0.0% colchicine, immersion 16 hours) and the lowest was in the c3t1 treatment (0.1% colchicine, immersion 8 hours). This indicated that adding a small concentration of colchicine and increasing immersion time significantly affected mor-phology of
Colocasia esculenta var. Antiquorum, caused more exposed cells to be damaged or fail during growth. This was supported by
Allum et al. (2007), who stated that at the concentration, the chromosomes will be doubled in the cell.
The best morphology in this study was obtained at a concentration of 0.0% due to the stress on the plant in colchicine treatment. However, in subsequent subcultures, other differences were observed between the concentration of 0.0% and colchicine at 0.05%, 0.075%, and 0.1% because the plants were no longer under stress due to colchicine administration. The nature of colchicine mutagen influenced morphology differences in each treatment due to its high toxicity to plants. Therefore, low doses with a long exposure period were considered reliable to reduce its toxic effect and increase the rate of polyploid production (
Sajjad et al. 2013). The optimal amount of colchicine used for polyploid output varied widely, with concentrations ranging from 0.01% (
Thao et al. 2003) to 1.0% (
Demtsu et al. 2013).
In the correlation analysis in
Table 4, the closeness between the variables was measured without considering the influence or the magnitude of one variable on others. In this study, the main character was the number of leaves because it can be used as a character to identify tetraploid plants. The results of the correlation analysis showed that four characters were positively correlated, including a very significant positive correlation on the number of roots (0.80) and shoots (0.75), speed of shoots (0.69), and speed of rooting (0.66). This correlation proved the extent to which a relationship exists between one or more variables (
Astuti 2017). According to (Rohaeni 2012), correlation with strong character conditions can be used as an indirect selection tool for the main character.
Based on the results, colchicine with the best effect on the formation of polyploid of Safira taro was the interaction between 0.05% colchicine concentration and 16 hours immersion time (c1t2), which resulted in tetraploid (4n) of 54.10%. This was followed by the combination of 0.1% colchicine concentration with 16 hours of immersion (c3t2), yielding tetraploid (4n) of 44.36%. Furthermore, colchicine with the best effect on morphology parameters was obtained from the interaction between colchicine concentration of 0.075% with 16 hours of immersion (c2t2), which produced 20.33 leaves and 4.00 roots, while a concentration of 0.075% with 8 hours of immersion (c2t1) yielded 19.33 shoots.
Fig. 1Colocasia esculenta var. antiquorum plants with several concentrations of colchicine and In vitro immersion time after 16 weeks. (A) c0t1 (0% colchicine, 8 hours immersion); (B) c1t1 (colchicine 0.05%, immersion 8 hours); (C) c2t1 (colchicine 0.075%, immersion 8 hours); (D) c3t1 (colchicine 0.1%, immersion 8 hours); (E) c0t2 (0% colchicine, immersion 16 hours); (F) c1t2 (colchicine 0.05%, immersion 16 hours); (G) c2t2 (colchicine 0.075%, immersion 16 hours); (H) c3t2 (colchicine 0.1%, immersion 16 hours).

Fig. 2Graph letal concentrasion of living shooting per-centage of Colocasia esculenta var. antiquorum from first week to sixteenth week after In vitro induc-tion. c0t1 (0% colchicine, immersion 8 hours); c1t1 (colchicine 0.05%, immersion 8 hours); c2t1 (colchi-cine 0.075%, immersion 8 hours); c3t1 (colchi-cine 0.1%, immersion 8 hours); c0t2 (0% colchi-cine, im-mersion 16 hours); c1t2 (colchicine 0.05%, immer-sion 16 hours); c2t2 (colchicine 0.075%, immersion 16 hours); c3t2 (colchicine 0.1%, im-mersion 16 hours).
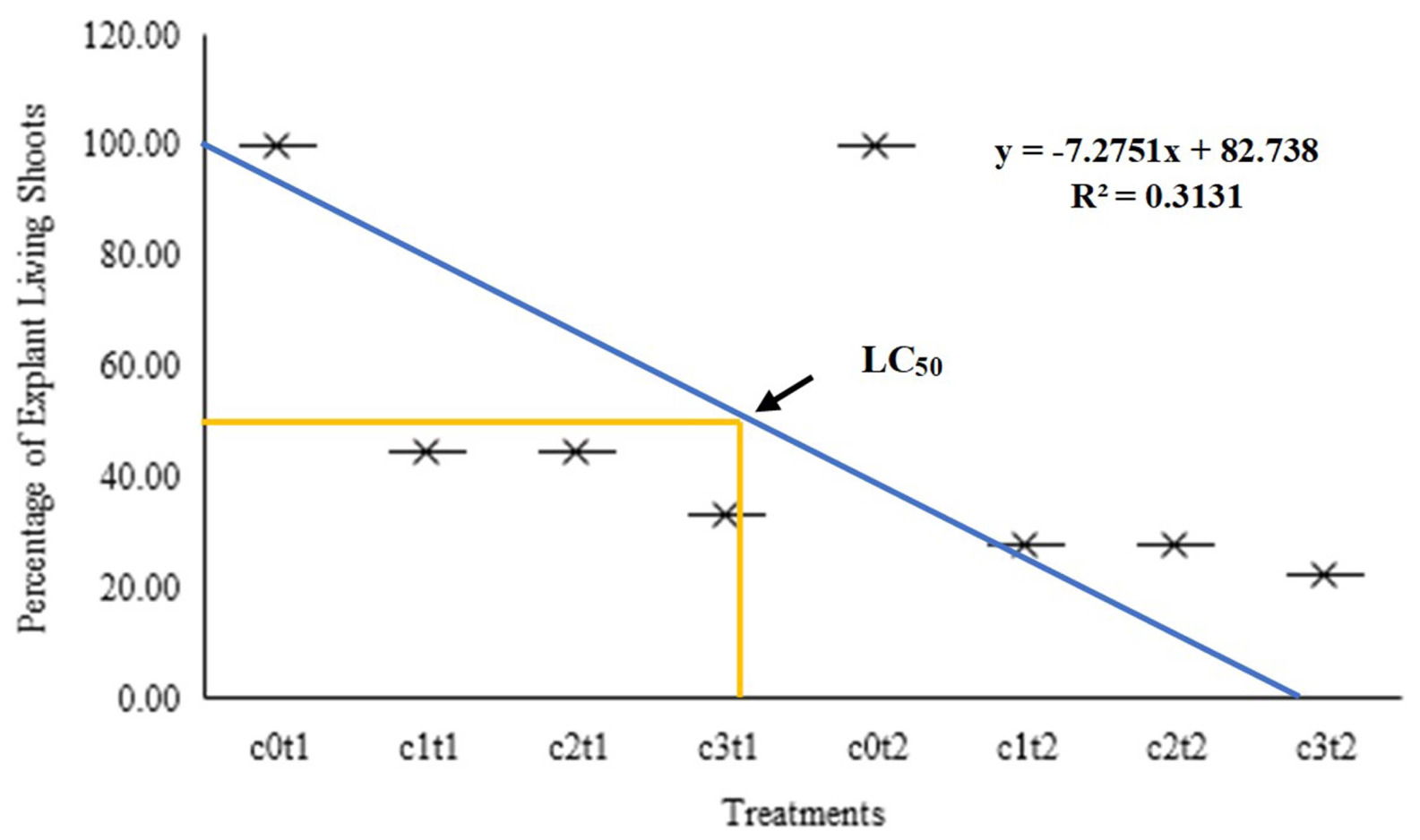
Fig. 3The effect of colchicine concentration and immer-sion time on the number of living leaves of Colo-casia esculenta var antiquorum plants In vitro. Numbers followed by the same letter (a-g) mean that they are not significantly different in the DMRT test with a level of = 0.05.
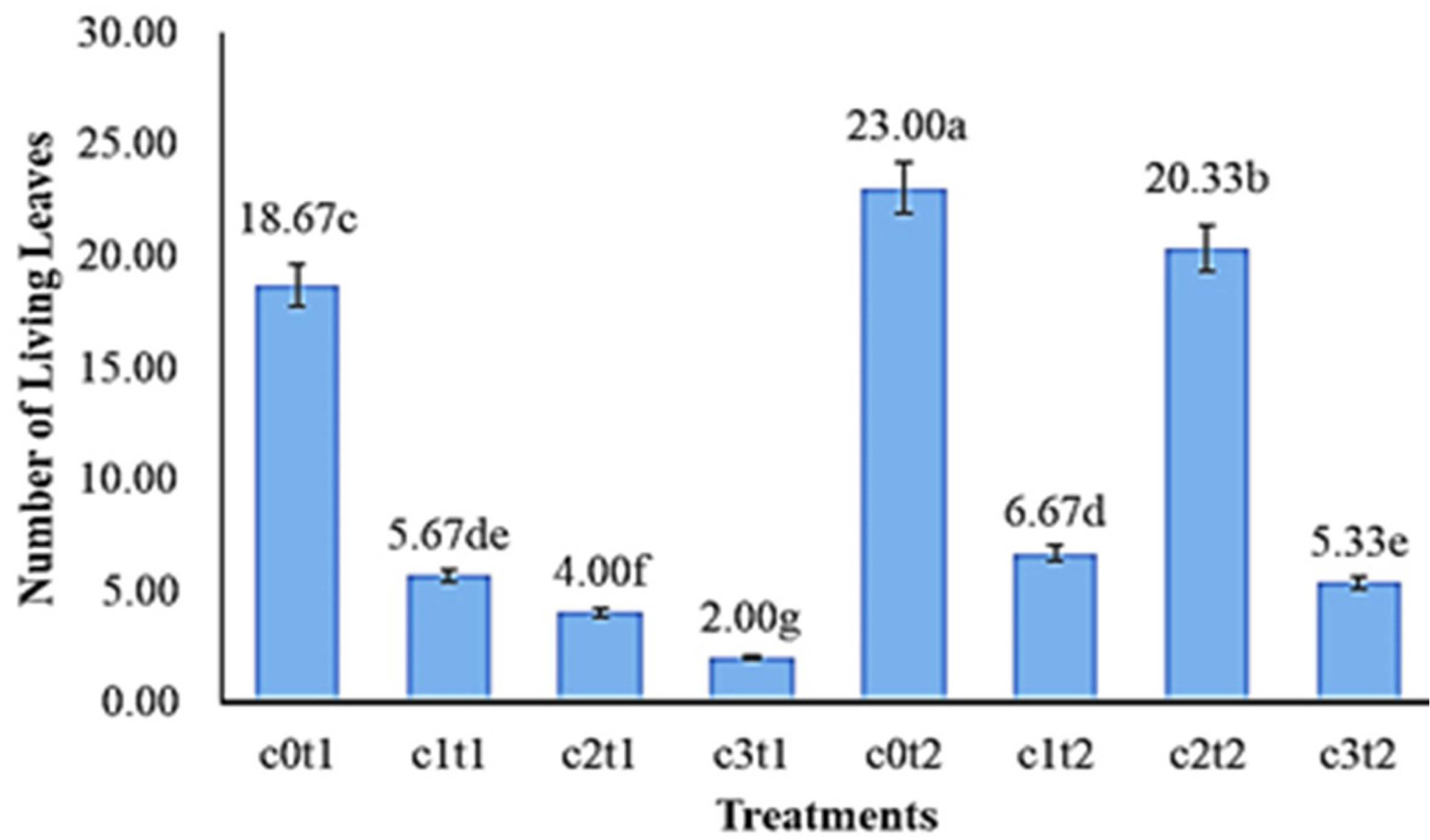
Fig. 4Effect of colchicine concentration and immersion time on root number of Colocasia esculenta var. antiquorum In vitro. Numbers followed by the same letter (a-d) mean that they are not signi-ficantly different in the DMRT test with a level of = 0.05.
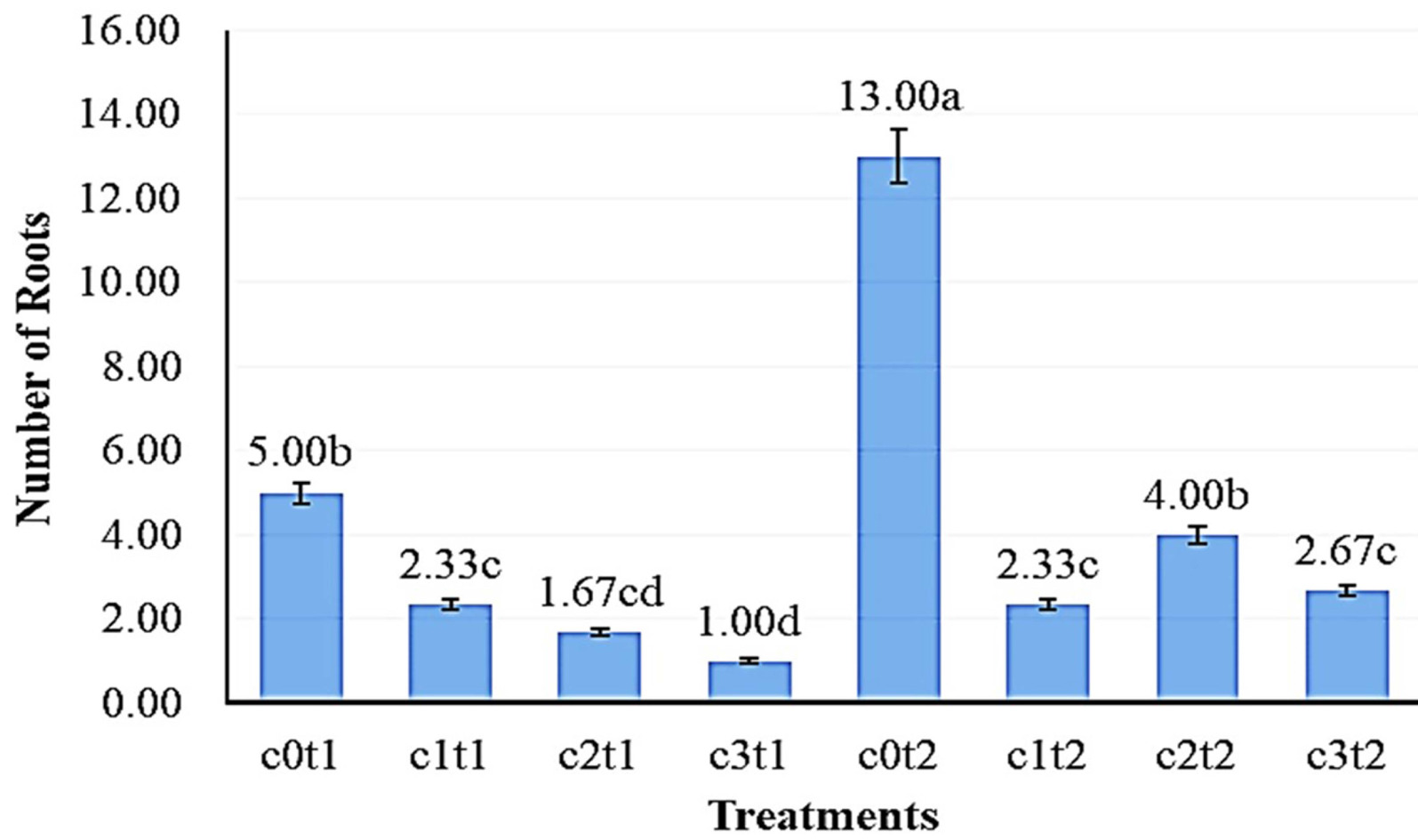
Fig. 5The effect of colchicine concentration and immer-sion time on the number of compound shoots of Colocasia esculenta var. antiquorum plants In vitro. Numbers followed by the same letter (a-f) mean that they are not significantly different in the DMRT test with a level of = 0.05.
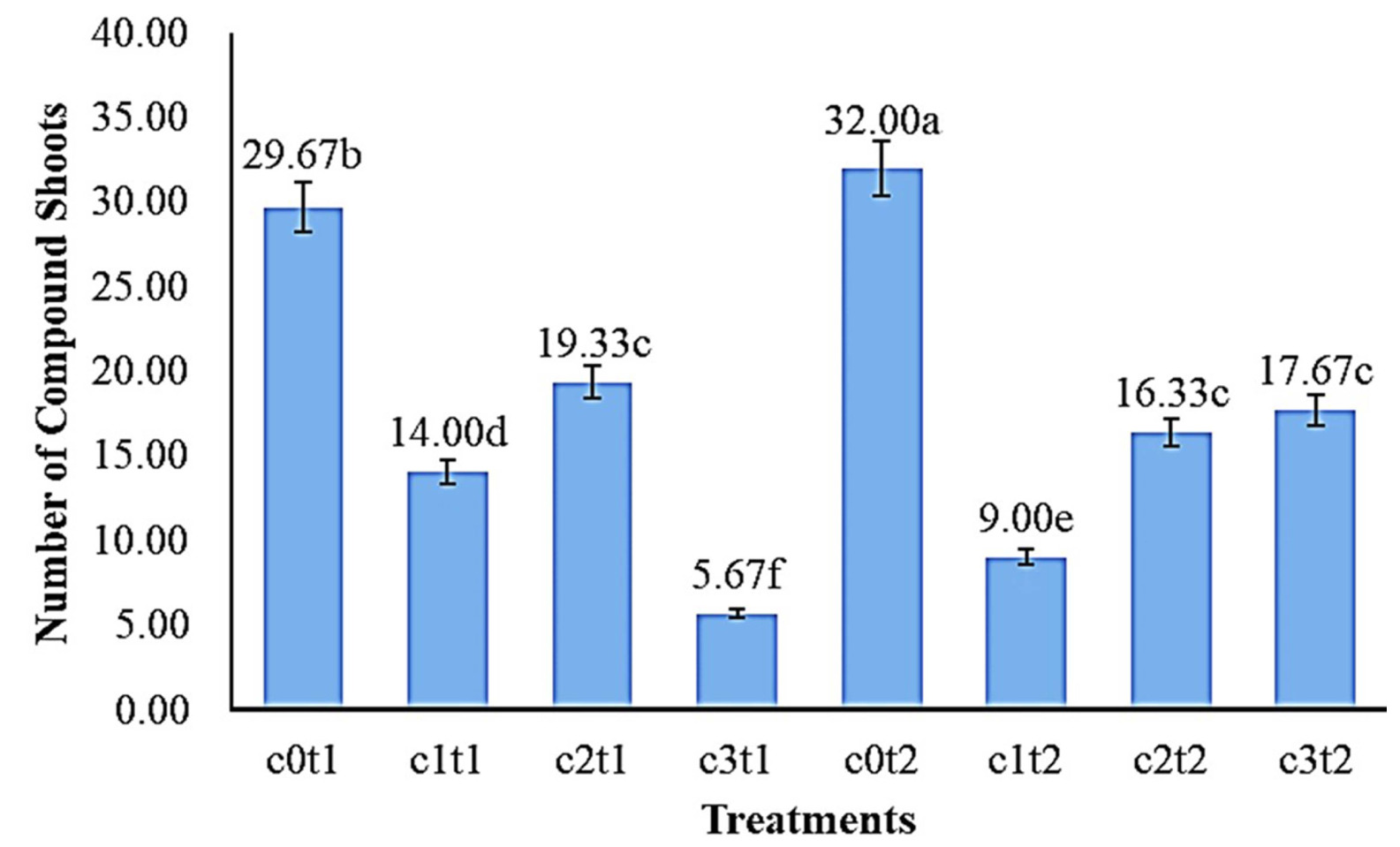
Fig. 6Histogram of ploidy degree results with flow cytometry of Colocasia esculenta var. antiquorum plant resulting in In vitro colchicine induction. (A) c0t1 (0% colchicine, immersion 8 hours) and c0t2 (0% colchicine, immersion 16 hours); (B) c1t1 (colchicine 0.05%, immersion 8 hours); (C) c2t1 (colchicine 0.075%, immersion 8 hours); (D) c3t1 (colchicine 0.1%, immersion 8 hours); (E) c1t2 (colchicine 0.05%, immersion 16 hours); (F) c2t2 (colchicine 0.075%, immersion 16 hours); (G) c3t2 (colchicine 0.1%, immersion 16 hours).
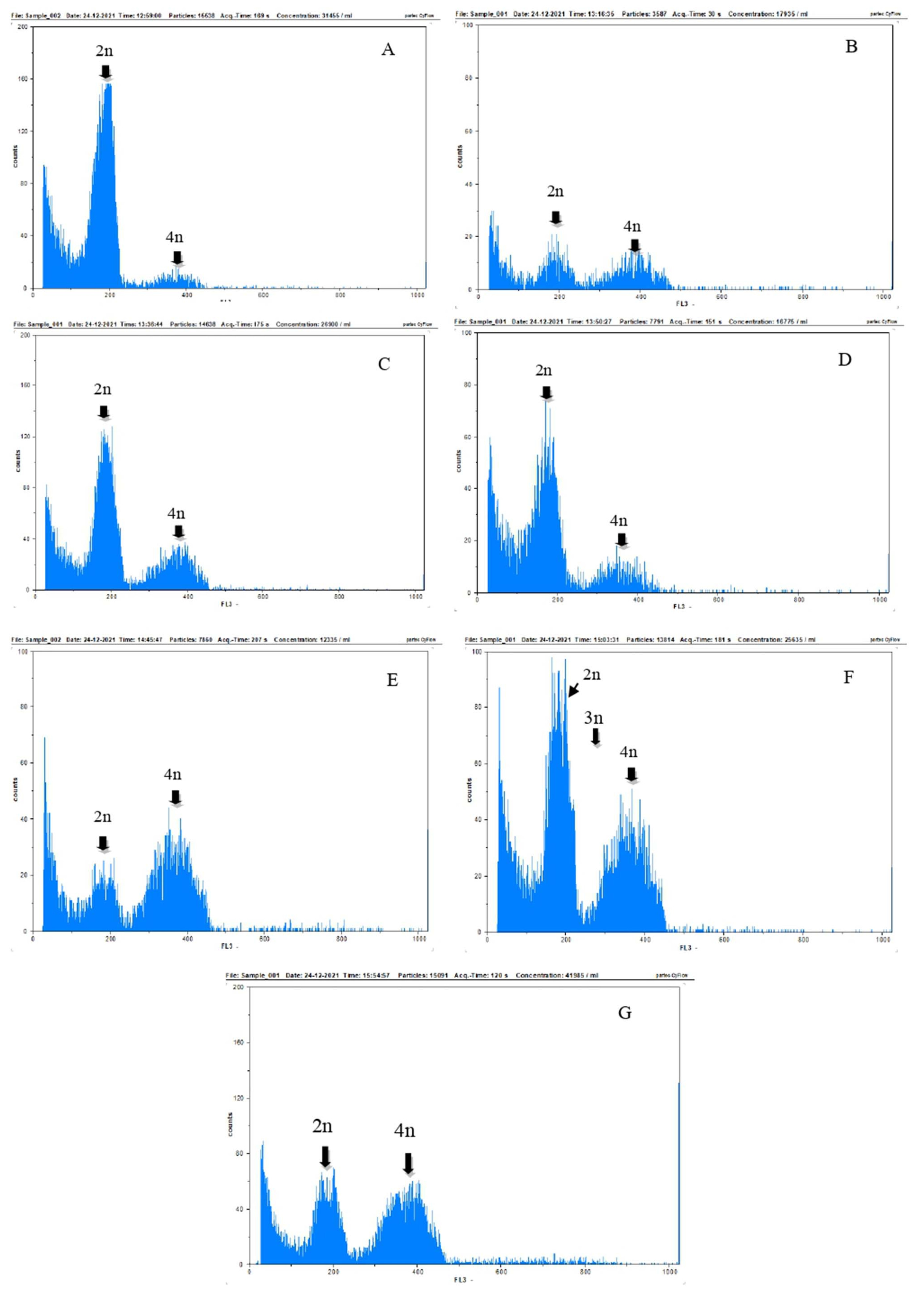
Table 1Percentage of living shoots on each treatment of colchicine concentration and time of immersion.
Table 1
|
Treatments |
Shoot planted |
Living shoots |
Dead shoots |
Living percentage (%) |
|
c0t1 |
13 |
13 |
0 |
100.00 |
|
c1t1 |
9 |
4 |
5 |
44.44 |
|
c2t1 |
9 |
4 |
5 |
44.44 |
|
c3t1 |
12 |
4 |
8 |
33.33 |
|
c0t2 |
13 |
13 |
0 |
100.00 |
|
c1t2 |
18 |
5 |
13 |
27.78 |
|
c2t2 |
18 |
5 |
13 |
27.78 |
|
c3t2 |
18 |
4 |
12 |
22.22 |
|
Total |
110 |
52 |
56 |
47.27 |
Table 2Mean square analysis of variance on several observation parameters in In vitro polyploid induction experiments of Colocasia esculenta var. Antiquorum plants.
Table 2
|
Source |
DF |
Mean square in annalysis of variance (ANNOVA) |
|
Number of living leaves |
Number of roots |
Number of compound shoots |
|
Time immersion (T) |
1 |
234.375**
|
54.000**
|
15.042ns
|
|
Colchicine concentration (C) |
3 |
349.708**
|
67.667**
|
494.153**
|
|
Interaction of T × C |
3 |
70.708**
|
18.111**
|
86.708**
|
|
Error |
16 |
4.333 |
0.542 |
12.458 |
|
CV (%) |
16.82% |
19.44% |
18.40% |
Table 3Results of polyploidy analysis with flow cytometry in Colocasia esculenta var. Antiquorum plants In vitro induction of colchicine.
Table 3
|
Treatment |
Chromosomes detected |
% gated |
Dominant chromosomes |
Explanation (*) |
|
Control (c0t1, c0t2) |
2n-4n |
67.70 + 4.17 |
2n |
Diploid |
|
c1t1 |
2n-4n |
28.35 + 36.97 |
4n |
Putatif-Tetraploid |
|
c2t1 |
2n-4n |
52.94 + 24.99 |
2n |
Mixoploid |
|
c3t1 |
2n-4n |
55.26 + 16.25 |
2n |
Mixoploid |
|
c1t2 |
2n-4n |
20.27 + 54.10 |
4n |
Tetraploid |
|
c2t2 |
2n-3n-4n |
29.94 + 15.39 + 35.50 |
4n |
Putatif-Tetraploid |
|
c3t2 |
2n-4n |
28.24 + 44.36 |
4n |
Tetraploid |
Table 4Results of correlation analysis on all observation parameters of Colocasia esculenta var. Antiquorum plants at various concentrations and duration of In vitro immersion of colchicine.
Table 4
|
Number of compound shoots |
Number of roots |
Number of living leaves |
|
Number of compound shoots |
1.00 |
0.79**
|
0.75*
|
|
Number of roots |
|
1.00 |
0.80**
|
|
Number of leaves |
|
|
1.00 |
References
- Ade R, Rai MK. 2010. Review: Colchicine, current advances and future prospects. Nusantara Biosci.. 2: 90-96. https.
- Allum JF, Bringloe DH, Robers AV. 2007. Chromosome doubling in a rosa rugosa thub. hybrid by exposure of In vitro nodes to oryzalin: The effects of node lenght, oryzalin concentration and exposure time. Plant Cell Rep.. 26: 1977-1984.
- Anshori MF, Purwoko BS, Dewi IS, Suwarno WB, Ardie SW. 2022. Salinity tolerance selection of double-haploid rice lines based on selection index and factor analysis. AIMS Agric. Food. 7(3): 520-535.
- Ari E, Djapo H, Mutlu N, Gurbuz E, Karaguzel O. 2015. Creation of variation through gamma irradiation and poly-ploidization in Vitex agnus-castus L. Sci. Hortic.. 195: 74-81. http.
- Asif MJ, Othman RY. 2001. In vitro zygotic embryo culture of wild Musa Acuminata Spp. malaccensis and factors affecting germination and seedling growth. Plant Cell Tissue Organ Cult.. 67: 267
- Astuti CC. 2017. Correlation analysis to determine the close-ness of the relationship between student activity and final learning outcomes. Journal of Information and Computer Technology Education. 1: 1-7. https.
- Ayu GM, Nurwahyuni I. Elimasni2019. Effect of con-centration and duration of colchicine treatment to garlic (Allium Sativum L.) Cv. Doulu. Int. J. Sci. Technol. Res.. 8: 172-175.
- Bernard F, Moghbel N, Hassannejad S. 2012. Treatment of licorice seeds with colchicine: changes in seedling DNA levels and anthocyanin and glycyrrhizic acid contents of derived callus cultures. Nat. Prod. Commun.. 7: 1457-1460. https.
- Cimen B. 2020. Induction of polyploidy in c35 citrange through In vitro colchicine treatments of seed-derived explants. Int. J. Fruit Sci.. 20: 1929-1941.
- De KK, Saha A, Tamang R, Sharma B. 2010. Investigation on relative genome sizes and ploidy levels of darjeeling- himalayan rhododendron species using flow cytometer. Indian J. Biotechnol.. 9: 64-68.
- De SS, Leus L, Eeckhaut T, Van BE, Debergh P, De LM. 2004. Somatic polyploid petals: Regeneration offers new roads for breeding belgian pot azaleas. Plant Cell Tissue Organ Cult.. 76: 183-188.
- Demtsu B, Taychasinpitak T, Wongchaochant S, Manochai B. 2013. Induced mutation by colchicine treatment of somatic embryos in 'namwa' banana (Musa sp. ABB). International Transaction Journal of Engineering, Mana-gement, and Applied Sciences and Technologies. 4: 311-320.
- Deng B, Du W, Liu C, Sun W, Tian S, Dong H. 2012. Antioxidant response to drought, cold and nutrient stress in two ploidy levels of tobacco plants: Low Resource Requirement Confers Polytolerance in polyploids?. . Plant Growth Regul. 66. 37-47. https://doi.org/10.1007/s10725-011-9626-6 66(1) 37-47:
- Eeckhaut TL, Leus HJ. Van2005. Exploitation of flow cytometry for plant breeding. Acta Physiol. Plant.. 27: 743-750.
- El-Nashar YI, Ammar MH. 2015. Mutagenic influences of colchicine on phenological and molecular diversity of Calendula officinalis L. Genet. Mol. Biol.. 15: 1-15. https.
- Eng WH, Ho WS, Ling KH. 2021. In vitro Induction and Identification of Polyploid Neolamarckia Cadamba Plants by Colchicine Treatment. Peer J. 1-26. https://doi.org/10.7717/peerj.12399.
- Escobedo-Gracia-Medrano RM, Burgos-Tan MJ, Ku-Cauich JR, Quiroz-Moreno A. 2018. Using flow cytometry analysis in plant tissue culture derived plants, p 1815: 317-332.
- Esfahani ST, Karimzadeh G, Naghavi MR. 2020. In vitro polyploidy induction in persian poppy (Papaver brac-teatum Lindl.). Int. J. Cytol. Cytosystematics Cytogenet.. 73: 133-144. https.
- Esfahani ST, Karimzadeh G, Naghavi MR. 2020. In vitro polyploidy induction in persian poppy (Papaver bractea-tum Lindl.). Caryologia. 73: 133-144. https.
- Ganga M, Chezhiyan N. 2002. Influence of the antimitotic agents colchicine and oryzalin on In vitro regeneration and chromosome doubling of diploid bananas (Musa spp.). J. Hortic. Sci. Biotechnol.. 77: 572-575. https.
- Gupta G, Memon AG, Pandey B, Khan MS, Shariq M, Srivastava JK. 2021. Colchicine induced mutation in ni-gella sativa plant for the assessment of morpho-physiolo-gical and biochemical parameter vis-a-vis In vitro anti- inflammatory activity. Open Biotechnol. J.. 15: 173-182. https.
- Heo JY, Jeong SH, Choi HR, Park SM. 2016. Polyploidy production in lilium leichtlinii var. maximowiczii using colchicine. Anim. Plant Sci.. 26: 1111-1116.
- Jala A. 2014. Colchicine and duration time on survival rate and micropropagation of Dionaea muscipula Ellis. Afr. J. Plant Sci.. 8: 291-297. https.
- Jones JR, Ranney GT, Eaker TA. 2008. A novel method for inducing polyploidy in rhododendron seedlings. J. Am. Rhododendron Soc.. 62: 130-135.
- Kainth D, Grosser JW. 2010. Induction of autotetraploid in pumelo (Citrus grandis L. Osbeck) through colchicine treatment of meristematically active seeds In vitro. Annu. Meet. Fla. State Hort. Soc.. 123: 44-48.
- Karlik E, Deger M, Uzen E, Gozukirmizi N. 2020. Pioneering In vitro studies for callus formation of Colchicum chalcedonicum Azn. Trak. Univ. J. Nat. Sci.. 21: 131-137.
- Kharde AV, Chavan NS, Chandre MA, Autade RH, Khetmalas MB. 2017. In vitro enhancement of bacoside in brahmi (Bacopa monnieri) using colchicine. Plant Physiol. Biochem. Physiol.. 5: 1-6. https.
- Koutoulis A, Roy AT, Price A, Sheriff L, Legget G. 2005. DNA ploidy level of colchicines-treated hops (Humulus lupulus L.). Sci. Hortic.. 105: 263-268.
- Kurtz LE, Brand MH, Lubell-Brand JD. 2020. Production of tetraploid and triploid hemp. Hort. Sci.. 55: 1703-1707. https.
- Mahyuni R, Girsang ESB, Hanafiah DS. 2015. The effect of colchicine on morphology and number of chromosome of binahong plant (Anredera cordifolia Tenn. Steenis.). Jurnal Agroekoteknologi. 4(1): 1815-1821.
- Manzoor A, Ahmad T, Bashir MA, Baig MMQ, Quresh AA, Shah MKN, Hafiz IA. 2018. Induction and identification of colchicine induced polyploidy in gladiolus grandiflorus white prosperity. J. Fol. Hor.. 30: 307-319. https.
- Manzoor A, Ahmad T, Bashir MA, Hafiz IA, Silvestri C. 2019. Studies on colchicine induced chromosome doubl-ing for enhancement of quality traits in ornamental plants. Plants (Basel).. 8(7): 194. doi.
- Maretta D, Dwi PH, Henti R, Amelia T. 2016. Shoot multiplication and induction of micro satoimo tubers (Colocasia esculenta (L.) Schott) at several concentrations of sucrose and benzylaminopurine. Jurnal Bioteknologi dan Biosains Indonesia. 3: 81-88.
- Mo L, Chen J, Lou X, Xu Q, Dong R, Tong Z, Huang H, Lin E. 2020. Colchicine-induced polyploidy in Rhododendron fortunei Lindl. Plants. 9(4): 424. https.
- Nagano M, Nur ES. Sunaryo2016. Study of the application of UB compost on the growth and yield of taro (Colocasia esculenta (L.) Schott var. Antiquorum) grown on dry land during the dry season. Jurnal Produksi Tanaman. 4(7): 570-577.
- Pirkoohi MH, Keyvanloo M, Hassanpur M. 2011. Colchicine induced polyploidy in mint by seed treatment. Int. J. Agric. Crop Sci.. 3: 102-104.
- Rohaeni WR, Permadi DANK. 2012. Path analysis of several yield component characteristics of lowland rice yields in the agrisimba application. Journal on Agri-culture Science. 2: 185-190.
- Sadhukhan R, Ganguly A, Singh PK, Sarkar HK. 2014. Study of induced polyploidy in african marigold (Tagetes ecrecta L.). Environ. Ecol.. 32: 1219-1222.
- Sajjad Y, Jaskani MJ, Mehmood A, Ahmad I, Abbas H. 2013. Effect of colchicine on In vitro polyploidy induction in african marigold (Tagetes erecta). Pak. J. Bot.. 45: 1255-1258.
- Sattler MC, Carvalho CR, Clarindo WR. 2016. The polyploidy and its key role in plant breeding. Planta.. 243: 281-296. https.
- Amier N, Amal MAI. Sudirman2021. Review: Induction of diversity based on biotechnology in orchids (Orchids Sp.). J. Agrivigor. 12: 75.
- Suryo.2007. Sitogenetika. Gadjah Mada University Press. Yogyakarta..
- Tesfaye, M2005. Agronomic and yield performance of induced-autotetraploid ensete (Ensete ventricosum) clones. Plant Genet. Resour. Newsl.. 143: 56-58.
- Thao NTP, Ureshino K, Miyajima I, Ozaki Y, Okubo H. 2003. Induction of tetraploids in ornamental alocasia through colchicine and oryzalin treatments. Plant Cell, Tissue and Organ Cult.. 72: 19-25. https.
- Urwin NA. 2014. Generation and characterization of colchicine- induced polyploid lavandula x intermedia. Euphytica. 197: 331
- Wang X, Cheng ZM, Zhi S, Xu F. 2016. Breeding triploid plants: A Review. Czech J. Genet. Plant Breed.. 52: 41-54. https.
- Wulandari DR, Ermayanti TM, Purwito A, Susanto S, Husni A. 2015. In vitro induction of tetraploid pummelo 'Nam-bangan' (Citrus maxima (Burm.) Merr. ) by colchicine treatment using germinated seed, shoot tip and coty-ledonary node as eplants. Annales Bogorienses. 19: 5-7.
- Xu CG, Tang TX, Chen R, Liang CH, Liu XY, Wu CL, Yang YS, Yang DP, Wu H. 2014. A Comparative study of bioactive secondary metabolite production in diploid and tetraploid Echinacea purpurea (L.) moench. Plant Cell Tissue Organ Cult.. 116: 323-332.