Abstract
A resurrection has started in haploid and double haploid research in the twenty-first century. The haploid and double haploid could be achieved through in vivo and in vitro anther and microspore culture techniques. Fixing the homozygosity is the most striking benefit of androgenesis. Various factors like genotypic dependency, growth condition, developmental stage of the microspore, pre-treatment, culture media, regeneration media, growth hormones, and various chemicals have a direct effect. Wheat, rice, Brassica, and tobacco are the notable crops where anther and microspore culture has been utilized. These haploidy and double haploidy through anther culture served many purposes of basic and applied research. Especially, double haploid cultivars have been cultivating around the globe. In addition, for chromosome mapping, QTL mapping, marker-assisted selection, marker-assisted backcrossing, mutation breeding, genome-wide association study, genomic engineering, and genome editing, androgenesis based haploid and double haploid plants have been exploited due to the effectiveness. Recently, researchers are trying to explain albinism that happens during anther culture from an epigenetic perspective. Further prospects of haploid and doubled haploid research through anther culture have been described in this review.
-
Key words: Haploid, Anther culture, Microspore culture, Rice, Wheat, Brassica, Tobacco, Albinism
INTRODUCTION
Haploid plants are those which contain gametophytic chromosome number and it indicates plants contain a single set of chromosome (
Germanà 2011). When the haploid plants undergo spontaneous or induced chromosome dou-bling procedures like- colchicine treatment, nitric oxide, or oryzalin treatment they become double haploid or fertile homozygous diploid (Mishra
et al. 2016). Though the haploid plants are sexually sterile, the double haploids are fertile and pure homozygous lines (
Devaux and Pickering 2005). The haploid and double haploid plants could be obtained through in vivo and in vitro techniques.
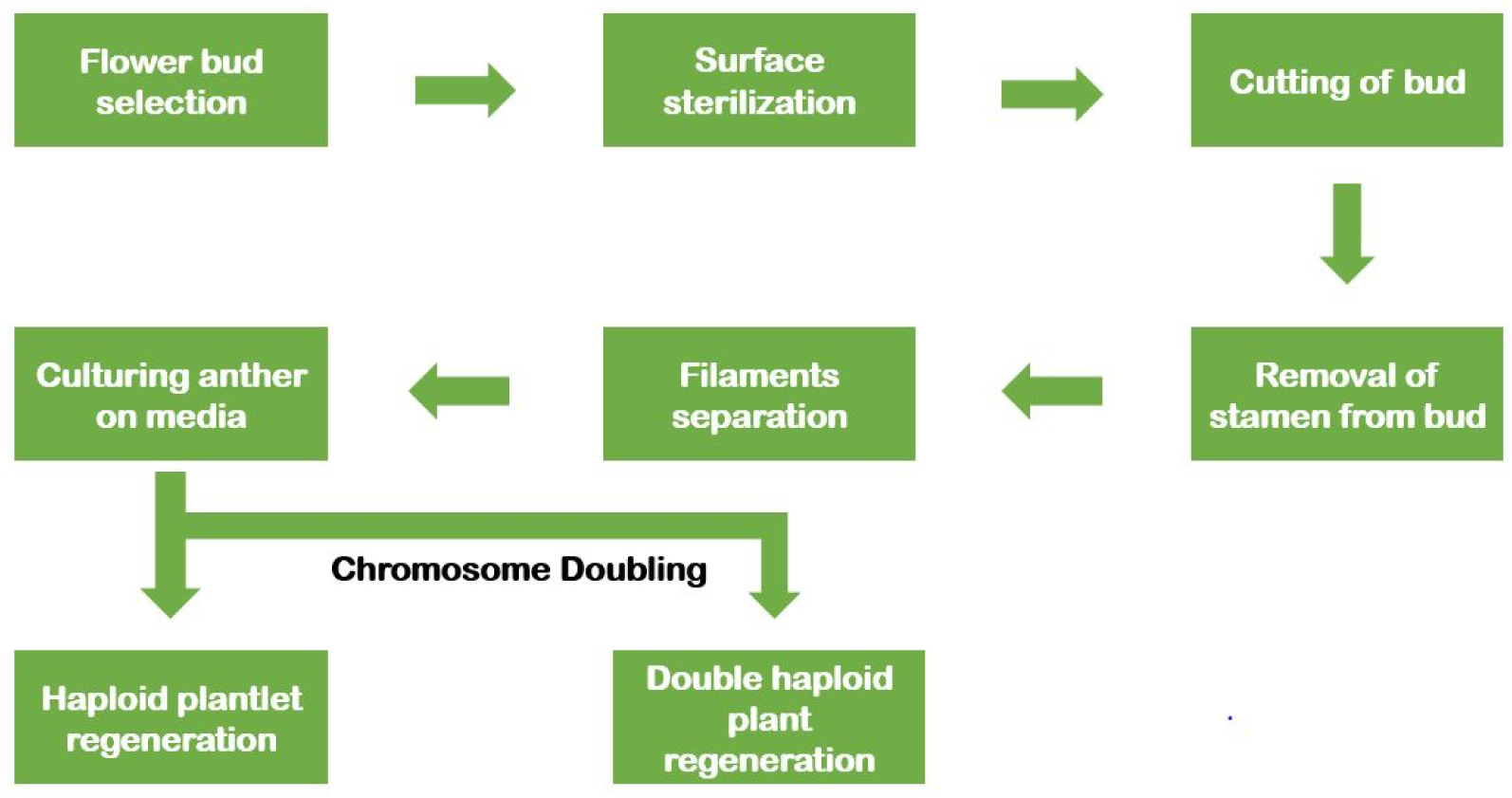
Androgenesis is the process of haploid production through the culturing of anther or microspores in vitro conditions (
Fig. 1). Anther culture is a widely utilized technique for developing haploid and double haploid plants because of its simplicity and effectiveness. It covers a vast range of genotypes with higher plant establishment (
Sopory and Munshi 1996). Also, haploid plant regeneration is possible through isolated microspore culture. Due to its delicacy, this technique demands sound technique and improved equipment compared to anther culture (
Germanà 2011).
In vitro anther culture is an empirical study where crops like wheat (
Triticum aestivum L.), barley (
Hordeum vulgare L.), tobacco (
Nicotiana spp.), rapeseed (
Brassica napus L.) showed high regeneration capability. This specialty made these crops model species to study the underlying mechanism (
Forster et al. 2007). The gametic embryogenesis to develop haploid plants had been studied by plant scientists for above around 5 decades and the notable review articles are
Magoon and Khanna (1963), Kasha (1974),
Zhang et al. (1990), Jain
et al. (1996–1997),
Smykal (2000), Maluszynski
et al. (2003a, b),
Andersen (2005), Palmer
et al. (2005),
Xu et al. (2007), Seguı`- Simarro and Nuez (2008a),
Touraev et al. (2009),
Dunwell (2010), Seguı`-Simarro (2010), Germanà (1997, 2006, 2007, 2009, 2011). Then,
Maluszynski et al. (2003a) stated fourty four protocols of developing double haploid from around thirty-three plant species.
This current review will try to address the history and crop-wise (rice, wheat, Brassica, and tobacco) success and utilization status of androgenesis along with albinism which is termed as the major drawback.
Importance of haploid and double haploid
The plant researchers have predicted the potentiality of haploid and double haploid (DH) in plant science research which was established in further years (
Sunderland 1978;
Forster and Thomas 2005). The pure line is mandatory for developing a hybrid variety and developing a pure line requires several generations by conventional approach (Keles
et al. 2015). Haploid and DH just cut off the duration and the homozygous pure line developed within a single generation and it is a major breakthrough in cultivar development (
Datta 2005;
Dunwell 2010). Afterward, in vegetable breeding, various parental lines show “inbreeding depression” and they only outyield in F1 condition after crossing with another parent (
Maluszynski et al. 2001;
Hochholdinger and Hoecker 2007). The hybrid perfor-mance could be fixed in a single line by using DH techno-logy (
Germanà 2011). Crops like rye (
Secale cereale) (
Immonen and Anttila 1996) and forage grasses (
Nitzche 1970) are recalcitrant to develop fertile homozygous by selfing. DH development of those crops is the best app-roach to confront the recalcitrance (
Germanà 2011).
Marker-assisted breeding along with the DH technique has been successfully utilized in the various breeding programs to cut short the breeding cycle (
Wessels and Botes 2014;
Tuvesson et al. 2007;
Mago et al. 2011). Then, the marker-assisted backcross breeding DH approach was integrated to reduce the duration and the target trait was introgressed in an elite line within the quickest time (
Toojinda et al. 1998).
The haploid and DH are also important tools in mutation breeding. The homozygous mutant line was developed by inducing mutation in the single gametic cells which gone through embryogenesis and double haploid mutant plant population achieved (
Szarejko and Forster 2006). Alterna-tively, the mutation was used in the gamete of the M1 plant to get the DH plant. In barley, rice, and wheat it has been utilized successfully (
Forster et al. 2007). Noteworthy, the unicellular microspore is recommended for mutation induction (
Germanà 2011). Then, the trait developed by mutation is recessive, and sometimes it is found in M2 or M3 generations. In that case, double haploidy is a good option; it just fixes the desired recessive trait (
Germanà 2011; Szarejko and Forster 2007). TILLING (Targeting Induced Local Lesions In Genomes;
McCallum et al. 2000; Perry
et al. 2003) is a reverse genetic approach which applied to relate the gene with the phenotypes by using chemical mutation along with the high throughput screen-ing method like SNPs (single nucleotide polymorphisms). Due to the inherent variation, a false-positive result could be achieved from the starting material (
Germanà 2011). To resolve this problem, in vitro methods like anther or microspore culture is a good alternative (
Tadele et al. 2010).
Double haploid and haploid have strengthened its position in applied research and basic research of breeding and genomics. The DH lines have homozygosity and uniformity; moreover, they can be replicated multiple times in various locations. This facility makes double haploid ideal for genetic mapping (
Tinker et al. 1996;
Khush and Virmani 1996). Likewise, in genetic and physical mapping the double haploid has considerable precision to locate the targeted gene (
Kunzel et al. 2000;
Wang et al. 2001). The chromosome map of various crops including barley, rice, rapeseed, and wheat had constructed using the haploid and double haploid plant (
Forster and Thomas 2005). In the ‘gametoclonal variation’ variation study the haploids have great potentiality as it points out the morphological, biochemical, and chromosomal variation within the plants derived through gametic cell culture (
Evans et al. 1984;
Morrison and Evans 1987). Also, the variations due to segregation, independent assortment, chromosome doubling procedure, and diploid level are explained from the ‘gametoclonal variation’ (
Morrison and Evans 1987;
Huang 1996).
Potrykus and his co-authors predicted the potentiality of the anther in genetic transformation as anther cultured isolated pollen has remarkable regeneration frequency (
Potrykus et al. 1985;
Potrykus 1988) which has proved further. In genetic transformation, the single-celled micro-spore, haploid embryo, or calli act as the recipient of the gene of interest. Thus, the obtained DH plant will contain the transgene in the homozygous state. Microinjection, electroporation, particle bombardment, and
Agrobacterium tumefaciens-mediated transformation were the gene deli-very technique was found effective in microspore based gene delivery (
Touraev et al. 2001;
Chen et al. 2006;
Chauhan and Khurana 2011; Li
et al. 2012; Ohnoutková and Vlčko 2020).
Genome editing is the state-of-the-art technology of the twenty-first century where transgene is not required to achieve genetic variation. Zinc Finger Nucleases (ZFNs), Transcription Activator-Like Effector Nucleases (TALENs), and Clustered Regularly Interspaced Short Palindromic Repeats and CRISPR-Associated Protein 9 (CRISPR/ Cas9) are the popular genome editing techniques. Among those, CRISPR/Cas9 is the most updated and efficient tool to edit any organism. Generally, immature embryos are used as the ex-plant of transformation. By contrast, studies show that anther or microspore has feasibility over the conventional method because the availability of gene-tically and physiologically identical microspore or anther is relatively easy. Then, after diploidization (spontaneous/ induced) the plants are in a homozygous state whether it might take several generations to achieve the homozy-gosity. Afterward, due to the haploidy status, the gene to be edited will be half in number which increases the efficacy (
Bhowmik et al. 2018;
Han et al. 2021). In essence, microspore or anther culture-based haploid transformation will save time, space, and money with great effectivity (
Bhowmik et al. 2018;
Han et al. 2021;
Bilichak et al. 2020;
Haque et al. 2018;
Brandt et al. 2020;
Borisjuk et al. 2019; Jansing 2019).
The haploidy status of the plant was first observed in sporophytic weed species
Datura stramonium (Jimson weed) in 1921 with proper cytological evidence (
Blakeslee et al. 1922). In another report,
Harland, 1920 claimed about the haploid presence in a sea island cotton commonly named as “Man cotton” (
Harland 1920,
1936,
1955). Then, the occurrence of the first androgenic haploid was stated by
Kostoff (1929) found in tobacco. That contemporary times, the concept of pure lines and gene theory proposed by Wilhelm Johannsen in 1903 and 1909 respectively, also the phenomenon of induced mutation by H.J.
Muller’s (1927) published. These concepts helped to fathom the poten-tiality of haploids and double haploids in genetics and plant breeding, as a result, haploid and double haploid study gained the center of interest in further decades (
Maheshwari 1996). From 1920 to 1950, plant scientists intensively tried to develop haploid plants but at that time anther culture was not a familiar tool to researchers (
Maheshwari 1996). Finally, in 1964, Guha and Maheshwari generated haploid embryos through in vitro anther culture from the immature anther of
Datura innoxia. This novel discovery coincided with the prediction of
Katayama and Nei (1964) where they stated the potentiality of pollens in generating haploid plants. This discovery helps to initiate extensive research in Poaceae, Brassicaceae, and Solanaceae family (Germena 2011). Subsequently, a similar result was reproduced for tobacco in France and Japan (
Bourgin and Nitsch 1967;
Nitsch and Nitsch 1969;
Nakata and Tanaka 1968). Then,
Niizeki and Oono (1968) successfully generated an andro-genic haploid rice plant which is a major cereal crop globally. These breakthrough findings raised a pinnacle of interest in haploids so that they could achieve homozygous pure-line within a single generation (Nitsch and Noreel 1973;
Nitsch 1974a, b). As interest rises, the first inter-national symposium was held at the University of Guelph, Canada with the title of ‘Haploids in Higher Plants’ in 1974 (Kasha 1974).
The first double haploid crop variety was “Maris Haplona” of rapeseed mustard (
Brassica napus) released at the begin-ning of the 1970s (
Thompson 1972). Afterward, a Barley (
Hordeum vulgare) cultivar named Mingo released in 1980 which was also a DH crop variety (
Ho and Jones 1980). Noteworthy, the early twenty-first century has experienced a resurrection in haploid research especially in higher plants (
Forster et al. 2007). Many initiatives had been taken to add value to this workforce. European Union ran a project COST 851 program (
https://www.cost.eu/actions/851/#tabs|Name:overview) where a research network de-veloped to work on the topic “Gametic cells and molecular breeding for crop improvement” for six years (Germena 2011). Significantly around 300 DH crop varieties have developed using various double haploid inducing techni-ques including in vitro anther culture (
Germanà 2011), and around 200 species regenerated from androgenesis of Gramineae, Solanaceae, and Cruciferae family to date (
Germanà 2011;
Dunwell 1986; Hu and Yang 1986).
Sarao and Gosal (2018) and
Mishra and Rao (2016) discussed thoroughly in their reviews regarding rice androgenesis. In rice anther culture, the genotype is the most deciding factor (
Mishra and Rao 2016). Among species, indica has a very limited response to anther culture (
Chen et al. 1991). By contrast, japonica species have higher regeneration efficiency (
He et al. 2006). Then, early or mid- to late-uninucleate are the best stages for haploid regeneration (
Datta and Wenzel 1998). Pretreatment of rice pollen is essential where various stresses like temperature shock, osmotic shock, sugar starvation were implied to a various extent (
Mishra and Rao 2016;
Datta 2001). Even though, gamma irradiation had utilized to treat anther which proved as a useful technique (
Zapata and Aldemita 1989;
Chen et al. 2001). Afterward, colchicine was used other than a mutagenic agent to promote gametic embryo-genesis in rice (Alemano and Guiderdoni 1994).
As culture media, N6 media with a trace amount of NH
4+ is widely used in rice anther culture. Especially for indica species,
Raina and Zapata (1997) recommended MO19 medium. The culture media needs carbohydrate source. Sucrose is commonly utilized for these purposes but maltose has supremacy in a few contexts compared to sucrose, such as albino plant frequency reduced by maltose utilization (
Reinert and Bajaj 1977;
Sen et al. 2011;
Park et al. 2013). Besides these - sorbitol, amino acid, and AgNO
3 were found beneficial for plant regeneration from andro-genesis in rice (
Yoshida et al. 1994;
Ogawa et al. 1995;
Faruq et al. 2014). Growth regulators are also used in the in vitro culture of anther and it has a positive impact on in vitro culture (
Mandal and Gupta 1995). The most com-monly utilized growth regulator for callus induction from rice anther is 2,4-dophenoxy acetic acid (2,4-D) and naphthalene acetic acid (NAA) where both of them are auxin (
Trejo-Tapia et al. 2002). But for direct androgenesis Indole acetic acid (IAA) and NAA might be used (
Ball et al. 1993). In special cases, growth regulators like kinetin and benzyl amino purine are recommended where 2,4-D or NAA cannot serve properly (
Mandal and Gupta 1995). Normally agar is used as a solidifying agent to solidify the media. Another gelling agent named ‘Ficoll’ showed a good response in anther culture. The purity status of ‘Ficoll’ is much better compared to agar (
Kao 1981). Ficoll is the synthetic polymer of sucrose and it has non-ionic status. It enhances the ratio of green plants to albino plant achievement (
Lashermes 1992).
Rice anther should be collected from the middle to lower part of the donor plant (
Jacquard et al. 2006). For pollen development, optimum nitrogen content should be main-tained. Optimum nitrogen content ensures quality micro-spores with high embryogenic potential (
Lapitan and Violeta 1999). Temperature variation in the booting stage adversely affects the microspore development process (
Lapitan and Violeta 1999;
Mishra and Rao 2016). The role of anther orientation had been found critical in rice anther culture (
Yang and Zhou 1979). Even though, anther from rationed rice had utilized in anther culture to assess the efficacy (
Fig. 2,
Guzmán and Arias 2000).
A great extent of work has been done in rice haploid and double haploid research. China is holding a strong position having more than 100 DH lines or varieties developed through anther culture. Likewise, other few countries like India, Japan, South Korea, Hungary have devolved DH lines using the anther culture (
Siddique 2015).
As mentioned before, anther and microspore cultured haploid and DH has used to enhance the basic research, quantitative, qualitative, resistance status, and so on in rice. The first salt-tolerant indica rice variety named PSBRc50 ‘Bicol’ was developed by anther culture and released for cultivating in the salinity-prone area (
Senadhira et al. 2002). Then, in 2003 salinity tolerant DH rice variety released utilizing six F1 hybrids where indica and japonica varieties were exploited (
Lee et al. 2003). A DH line named AC1 found from crosses of multiple salt-tolerant lines showed significant salinity tolerance. AC1 was found suitable for cultivating in saline-affected regions of Bangladesh (
Thomson et al. 2010). Afterward, drought is another menace for rice production. A rice variety named “Janka” developed through anther culture having drought tolerance in India (
Pauk et al. 2009). Likewise, cold tolerance at an early stage found in an Indian variety known as “Abel” has developed through anther culture (
Pauk et al. 2009). Also, anther culture has successfully derived indica rice variety tolerant to aluminum toxicity (
Dewi et al. 2009). Likewise, the DH rice line has developed having multiple stress tolerance; tolerant to blast, aluminum toxicity, and shade (
Purwoko et al. 2010).
Hybrid rice could outyield commonly cultivated rice varieties (
https://www.irri.org/hybrid-rice). But the yield potential of the hybrids depends on the purity of the parent line. But with the elapse of time, the parental line deter-iorates due to various factors and ultimately results in lower quality and yield (
Mishra and Rao 2016). A restorer line of a hybrid named Minghui 63 purified through anther culture and showed significant results in yield and resistance (
Bai et al. 1991;
Zhu et al. 1998). Anther culture utilized to develop restorer (B) lines for cytoplasmic male sterile (A) line (
Wang et al. 1994). Then,
Palanisamy et al. (2019) developed an improved R line with superior grain quality.
Then, a linkage map has developed in rice using the DH line. These maps were utilized to identify the molecular markers linked with the important genes which had significant resistance against rice blast, bacterial blight, and sheath blight disease (
Wang et al. 2001). Moreover, the DH line assisted to develop a chromosome map of rice (
Forster and Thomas 2005). Significant quantitative cha-racters of rice are controlled by quantitative trait loci (QTL) (
Datta 2005). The individual effect of QTL is very trivial and it is affected greatly by environmental factors. To overcome the issue, multiple replicated trials are needed to achieve the actual phenotype. As DH has true-breeding nature and convenience of producing large numbers, the DH plant is widely used for the identification and mapping of QTLs in rice (
Mishra and Rao 2016). Sheath blight resistance-related QTL has been identified in japonica rice by using the DH population (
Xu et al. 2011). Afterward, six brown planthoppers resistance-related QTL successfully mapped using the DH mapping population (
Soundararajan et al. 2004). Likewise, 5 panicle layer-uniformity related, 2 yields, and yield component related QTLs were identified through exploiting the DH population (
Ma et al. 2009). Additionally, rice root traits QTLs, plant growth, yield, and yield components QTLs were also mapped by utilizing the rice DH population (
Li et al. 2003;
Hittalmani et al. 2002).
Amylose content, protein content, and lipid content are the major rice quality indicators. Four amylose content related, 2 protein content-related, and 2 lipid content related QTLs had mapped using the DH rice population (
Lee et al. 2014). Alongside, Anther culture has a great potentiality in speed breeding and grain quality enrichment (
Xa and Lang 2011). A rice variety named Koshihikari gave superior grain and eating quality developed through DH technology (
Xa and Lang 2011). Likewise, Milyang 90, Guan 18 are another culture-derived genotype that showed superior grain quality. Notably, Guan 18 is a hybrid indica rice variety (
Chung 1987;
Zhu and Pan 1990).
In mutation breeding, Gamma irradiated rice anther lines gave an early and higher number of flower-producing plants (
Myint et al. 2005). Then, mutant double haploid plant resistance to Striga (a parasitic weed of rice) was achieved through anther culture (
Table 1) (
Shariatpanahi et al. 2018).
Iron and Zinc deficiency is a common problem around the world that leads to various avoidable diseases. Bio- fortification of the major cereals like rice is one of the probable solutions. Plant breeders are trying to develop new indica rice varieties enriched with zinc and iron by utilizing japonica donors. International Rice Research Institute had started a project to attain the goal and DH production through anther culture had considered as one of the probable approaches. IRRI had evaluated above 1500 DH lines to find the potential lines enriched with micro-nutrients like iron and zinc till 2009 (
Grewal 2009;
Mishra and Rao 2016).
Then, the DH population have utilized in marker-assisted selection (MAS) breeding considering the phenotypic precision (William
et al. 2002). A maintainer line has developed through the coupled utilization of anther culture and MAS. Notably, the cytoplasmic sterile maintainer had disease resistance, aroma, and red hull. Then, using this same approach dwarfing gene and glast resistance gene had introgressed in Basmati rice cultivar named ‘Ranbir Basmati’ (
Samal et al. 2019). Afterward, anther culture- based
Agrobacterium-mediated genetic transformation had conducted in rice (
Chen et al. 2006; Li
et al. 2007, 2010).
In 1973, the first anther culture-derived bread wheat (
T. aestivum) plant regenerated by three research groups (
Ouyang et al. 1973;
Chu et al. 1973; Picard and De Buyser 1973). Likewise rice, spontaneous doubling prevails in bread wheat ranging from 25 to 70% (
Maluszynski et al. 2003a, b).
The first DH wheat variety was developed by China in 1985 named ‘Jinghua No-1’ (
Hu et al. 1985). Within a short time, a DH variety named Florin developed in France (
De Buyser et al. 1987). In anther culture-driven DH wheat cultivar development, China showed good progress. Accord-ing to Hu (
Hu 1997), china developed 21 wheat cultivars until 1991 utilizing anther culture technique. Likewise, other countries developed DH wheat varieties and among those “Gk Delibab” (
Pauk et al. 1995), ‘SV Agaton’ (
Tuvesson et al. 2003), ‘McKenzi’ (
Graf et al. 2003), or ‘AC Andrew’ (
Sadasivaiah et al. 2004), ‘Huapei 8’ (Ming- hui
et al. 2011), ‘Kharoba’ (
Elhaddoury et al. 2012) and ‘GK Déva’ (
Pauk et al. 2020) are good examples. In addition, Hume, Gregory, Gladius, Axe, Crusader, Spitfire, Cobra, Gauntlet, Merlin, Fang, and Espada are few DH wheat varieties (
Broughton et al. 2014).
In Canada, 9 classes of Western Canadian Wheat are prevailing considering the milling quality (
https://www.grainscanada.gc.ca/en/grain-quality/grain-grading/wheat-classes.html). Among those classes, Canada Western Red Spring (CWRS) is one. According to
Dunwell (2010), 3 out of 5 wheat cultivars of CWRS were DH variety and a cultivar named “Lillian” (double haploid) covered 15% area of CWRS. Then “AC Andrew” named wheat cultivar covered the 99% cultivating area of Canada Western Soft White Spring (CWSWS) class which was also DH variety (
Dunwell 2010).
Factors affecting the wheat anther culture had thoroughly stated in Lantos and Pauk (2020). In brief, wheat has a strong genotypic dependency for in vitro androgenesis. Re-searchers had identified numerous wheat varieties amenable to anther culture (viz ‘Chris’, ‘Pavon’, ‘Svilena’, ‘Bob’) which become the desired test genotype (
Lazar et al. 1984;
Lantos et al. 2013;
Castillo et al. 2015;
Nielsen et al. 2015;
Seifert et al. 2016). In addition, few recalcitrant varieties have also been identified (
Torp et al. 2001;
Lantos et al. 2013;
Castillo et al. 2015). According to Lantos and Pauk (2020), both the amenable and recalcitrant genotypes will assist in developing a new protocol to mitigating the undesirable effect of genotype dependency and genotype × treatment interactions. Generally, two types of bread wheat are cultivated globally depending on the environment; winter wheat and spring wheat. Spring wheat shows more responsiveness compared to winter wheat during anther culture (
Weigt et al. 2020).
The growing condition of the donor plant is another deciding factor. Donor plants can be grown in controlled condition like a greenhouse year-round (
Ghaemi et al. 1995;
Torp et al. 2001;
Pauk et al. 2003;
Tuvesson et al. 2000,
2003;
Soriano et al. 2007, 2008;
Broughton 2008,
2011;
Redha and Suleman 2011;
Brew-Appiah et al. 2013;
Sanchez-Diaz et al. 2013). Alternatively, donor plants can be raised in field condition. Considering efficacy, a field growing plant is ideal for large scale production where controlled condition grown plants are helping researchers to optimize the protocol and excel applied research year-round (
Pauk et al. 2003;
Chauhan and Khurana 2011;
Lantos et al. 2013;
Zhao et al. 2015,
2017;
Weigt et al. 2016,
2019;
Lazaridou et al. 2017; Lantos and Pauk 2020). Mid- to late uninucleate stages of the microscope are the suggested stage for culturing anther (
Soriano et al. 2007, 2008;
Broughton 2008,
2011;
Chauhan and Khurana 2011;
Redha and Suleman 2011;
Rubtsova et al. 2013; Sanchez- Diaz
et al. 2013;
Zhao et al. 2015;
Castillo et al. 2015;
Echávarri and Cistué 2016;
Weigt et al. 2016,
2019;
Lazaridou et al. 2017;
Broughton et al. 2020;
Orlowska et al. 2020). Alternatively, early- and mid-uninucleate micro-spores were also utilized for wheat anther culture (
Datta and Wenzel 1987;
Datta and Wenzel 1987,
1995;
Tuvesson et al. 2000,
2003;
Datta 2005;
Lantos et al. 2013;
Lantos and Pauk 2016). Various stress factors like cold, heat, starva-tion, colchicine, osmotic shock, 2-HNA, DMSO, etc. had found beneficial (
Liu et al. 2001;
Barnabás 2003;
Shariatpanahi et al. 2006;
Echávarri and Cistué 2016). Short time cold treatment (3-8 days, 4-6℃) (
Ghaemi et al. 1995;
Broughton 2008,
2011;
Rubtsova et al. 2013;
Zhao et al. 2015,
2017;
Weigt et al. 2016,
2019;
Lazaridou et al. 2017; Sen 2017), long-duration cold treatment (2-5℃, 10 days-4 weeks) (
Pauk et al. 2003;
Lantos et al. 2013;
Lantos and Pauk 2016; Coelho
et al. 2018; Wang
et al. 2019), starvation (solely or combined with chemical treatment) (
Soriano et al. 2007, 2008;
Sanchez-Diaz et al. 2013;
Castillo et al. 2015;
Echávarri and Cistué 2016), heat treatment (3 days, 32℃) has implied successfully (
Ouyang et al. 1983;
Pauk et al. 2003;
Shariatpanahi et al. 2006;
Lantos et al. 2013;
Lantos and Pauk 2016). Previous studies indicated that colchicine has the capability to prompt microspore division and also can excel in the gametic embryogenesis in wheat (Barnabas
et al.1991). A similar effect has been found in the case of
Brassica (Mollers
et al. 1994), rice (Alemano and Guiderdoni 1994).
Various culture media has been used in wheat anther culture such as AM, C17, P2, P4, LIM, W14, MS3M (Lantos and Pauk 2020). Among those W14, and MS3M media have been using frequently (
Ouyang et al. 1989;
Lantos et al. 2013;
Rubtsova et al. 2013;
Lantos and Pauk 2016;
Lazaridou et al. 2017;
Zhao et al. 2017; Soriano 2007;
Sanchez-Diaz et al. 2013;
Castillo et al. 2015;
Echávarri and Cistué 2016). Lantos and Pauk (2020) have claimed that the modified W14 medium has found effective in wheat anther culture which has strengthened by previous findings (
Lantos et al. 2013, 2018, 2019;
Lantos and Pauk 2016;
Kanbar et al. 2020;
Pauk et al. 2020). Then, as a carbon source maltose has used frequently. The ficoll has been utilized as an osmotic agent (
Hunter 1987;
Datta and Wenzel 1987). Various growth regulators (2,4-D, benzyl adenine, centrophenoxine, dicamba, indole-3-acetic acid, kinetin, etc.) also exploited successfully to induce wheat plantlet (
Fig. 2; Lantos and Pauk 2020;
Castillo et al. 2015; Weight
et al. 2016, 2019;
Zhao et al. 2015,
2017;
Orlowska et al. 2020;
Rubtsova et al. 2013). In addition, a supplement like exogenous aliphatic polyamines (putrescine, cada-verine, spermidine, and spermine) was found beneficial including several wheat cultivars (
Mishra and Rao 2016;
Rajyalakshmi et al. 1995).
Utilizing the DH technique, QTL identification and genetic mapping of various traits of wheat have been done. Photoperiod, plant height, flowering time, yield compo-nents, and grain yield-related QTL has mapped using DH line (
Sourdille et al. 2000;
Heidari et al. 2012;
Cuthbert et al. 2008;
Kuchel et al. 2007a, b). Alongside, Fusarium Head Blight (FHB), Septoria tritici blotch resistance QTL had identified and mapped through DH (
Suzuki et al. 2012;
Kelm et al. 2012). Then, 6 lodgings resistance-related QTL has unveiled by utilizing DH mapping population of wheat developed by anther culture (
Hai et al. 2005). Then, QTL related to nitrogen uptake capacity also unfolded by DH mapping population of wheat developed by anther culture (
An et al. 2006). Then, 6 salt tolerance QTL had found where the mapping population was a cross combination of Excalibur and Kukri wheat variety (
Table 2) (
Asif et al. 2018).
Additionally, the DH wheat plant has the potentiality to develop hybrid wheat varieties (
Longin et al. 2014). A restorer line of hybrid wheat was bred using anther culture (
Shi-kuan and Ya-ying 1985). Similarly,
Shimada et al. (1994) developed a male sterility-maintainer line of wheat hybrid. In the case of genetic transformation, few reports had found where anther culture-derived haploid wheat embryo or callus had been used as an explant (
Haliloglu et al. 2004; Chauhan and Khurana. 2011;
Rustgi et al. 2020). Likewise, target mutagenesis was successfully achieved by genome editing utilizing the wheat microspore. The sgRNA of CRISPR/Cas9 had been delivered to wheat microspore and calli by electroporation and
Agrobacterium-mediated transformation (Bhowmik 2018;
Liu et al. 2020). Even though, for the first time in genome editing, the direct delivery of the Zinc finger nuclease (ZFN) protein in the intact wheat microspore had been claimed by
Bilichak et al. (2020). They used purified ZFN along with the cell‐penetrating peptides (CPP) to deliver to the microspore where the microspore wall remains intact.
Various factors have a direct effect on haploid and double haploid production of
Brassica and the principal factors had stated briefly by
Watts et al. (2020). Especially for white cabbage, the responsible factor had been opti-mized for the ease of microspore embryogenesis and double haploid production (
Bhatia et al. 2021). Firstly,
Brassica has a genotypic dependency. Notably, the geno-type dependency could be transferred from a highly respon-sive to a low responsive genotype. Especially, the respon-sible loci for embryogenesis have been identified in
B. napus and
B. campestris (
Cloutier et al. 1995;
Ajisaka et al. 1999). Then, the growth environment of the donor plant has a great role in anther or microspores derive embryogenesis (
Watts et al. 2020). In most cases, plants are grown at low temperatures responded significantly in
Brassica micro-scope embryogenesis (
Watts et al. 2020).
As the rate of spontaneous double haploid induction is very low in
Brassica, various chromosome doubling agents, for instance, colchicine, oryzalin, amiprophosmethyl, tri-fluralin, and propanamide are utilized to achieve a double haploid plant. Amongst, colchicine is widely utilized and factors like concentration, duration of treatment, and plant developmental stage are considered (
Watts et al. 2020). Notably, around 50-500 mg/L colchicine is used for 15-24 hours duration to yield double haploid (
Watts et al. 2020;
Mollers 1994;
Zhou et al. 2002).
The double haploid
Brassica lines have also been utilized to assess breeding potential, develop mapping population, QTL analysis, linkage mapping, and whole-genome se-quencing (
Table 2; Sing
et al. 2021;
Wang et al. 2011;
Yang et al. 2016;
Pink et al. 2008). Especially in mutation breeding, microspore has been used as the inception medi-um of mutation. After mutation induction, the changed segment of DNA, even if a recessive trait will be altered into a homozygous state due to the double haplodization (
Watts et al. 2020). In
Brassica, several reports have found where microspore-based mutations were conducted. A tho-rough discussion regarding microspore embryogenesis had discussed by
Ferrie and Möllers (2011). Notably, Ferrie
et al. (2008) treated the microspore of
B. rapa,
B. napus, and
B. juncea with a mutagenic agent and developed
Brassica lines with altered fatty acid content. Likewise, high oleic acid-containing
Brassica was also achieved (
Turner and Facciotti 1990;
Wong and Swanson 1991). Then, a similar approach was used to locate lower glucosinolate containing
B. napus germplasm (
Burbulis et al. 2001). Afterward, cold tolerant, salt-tolerant, and sclerotia-resistant mutant lines have developed through this In vitro mutagenesis approach (
Rahman et al. 1995;
Liu et al. 2005;
McClinchey and Kott 2008).
Tobaccos responsiveness to anther culture is compara-tively very high and demands less complexity (
Sunderland 2012). Several factors like donor plant growth condition, genotype, age, pre-treatment, photoperiod difference, anther developmental stage, and culture condition have a direct influence in increasing embryogenic response in tobacco (
Dunwell 1976;
Belogradova et al. 2009). Then, the pre-treatments such as chilling of buds, anaerobic anther treat-ment, and pre-treatment of the anther in water-saturated condition had found effective (
Nitsch and Norreel 1973; Duncan and Heberle 1976;
Sunderland 1978;
Imamura and Harada 1981;
Dunwell 1981). As culture media, N6 (Chu, 1978), MS (Murashige and Skoog 1962), H (
Nitsch 1972) media were utilized and
Sunderland (2012) recommended the N6 media. Especially, the presence of iron in culture media is vital because lacking iron inhibits embryogenesis in the globular stage (
Nitsch 1972). Using activated charcoal in the medium found beneficial in previous studies. The charcoal absorbs the inhibitory and toxic substances from medium or agar or from the wall of the senescing cell (
Anagnostakis 1974; Kohlenbach and Wernicke. 1978;
Horner et al. 1977).
Generally from one anther of tobacco 1-135 plantlet could be achieved (
Sunderland and Wicks 1971;
Nitsch 1972;
Horner et al. 1977). The growth stage of pollen is important. Empirically, it has been observed that bi-cellular pollen just after completing its first mitotic division gave consistent results (
Sunderland and Wicks 1969;
Sunderland 2012). Another, corolla length of tobacco could give an inkling about the anther responsiveness and the favorable range between 15 and 25 mm (Suntherland 2012).
Around 10-12% of plants derived from tobacco micro-spore culture are spontaneous DH. Then, colchicine could be to the diplodizing tobacco plant (Touraev and Heberle- Bors 2003). The ploidy level can be identified using flow cytometry or chromosome counting by cytological proce-dure (
Touraev and Heberle-Bors 2003). Overall, the detailed procedure of tobacco anther and microspore culture has been stated thoroughly in
Touraev and Heberle-Bors (2003) and Suntherland (2012). Especially,
Sunderland (2012) stated the protocols of developing plantlets from tobacco anther culture. The first protocol Involves using a solid agar medium and the second one utilizes liquid media where float culture is practiced and pretreatment is man-datory (
Sunderland 2012).
In genetic transformation, maize cytokinin-specific b- glucosidase gene was transferred to tobacco to observe the activity of b-glucosidase. Results showed that the overac-tivity of b-glucosidase dwindled the anther culture-derived pollen embryo regeneration (
Dubová 1996). Similarly, using CRISPR/Cas9 based genome editing, a genetically fixed double haploid line had achieved. The microspore of tobacco and the leaf explant was the recipient of the sgRNA specifically constructed for the green fluorescent protein (GFP) gene (
Table 4) (
Schedel et al. 2017).
Though there is no deep and clear indication about albinism occurrence, many researchers tried to unfold the behind mystery (
Makowska and Zimny 2015). Scientists attempted to explain it by cytological study, plastid genome study, and nuclear genome study (
Makowska and Oleszczuk 2014). Especially from the cytological aspect, the struc-tural change of chloroplast at various stages of anther or microspore culture which resist the chloroplast genesis by comparing the in vitro derived green plant and albino plant (Caredda
et al. 1999, 2000, 2004). Likewise, it has attempted to find out the difference in the plastid genome and nuclear genome by comparing the green and the albino plant (
Hofinger et al. 2000;
Muñoz-Amatriaín et al. 2008,
2009;
Makowska and Zimny 2015). A thorough discussion re-garding this research area had done by
Makowska and Oleszczuk (2014). Apart from these assumptions, Duarte- Aké
et al. (2016) tried to describe albinism from the epigenetics view. In brief, they found three types of plantlets (green, variegated, and albino) by culturing the somaclones of
Agave angustifolia Haw and observed different types of DNA methylation patterns. Finally, they concluded that in vitro culture induces “Epigenetic Stress Memory” which might favor the development of the albino and variegated shoot by a chromatic shift.
Genotypic variation has a direct role in albino plant occurrence. For example, in winter barley the frequency of albino plant occurrence is lower than the spring wheat. This phenomenon could be explained by the evolutionary adap-tation as the winter one gone through low-temperature stress (
Caredda et al. 2000;
Castillo et al. 2000;
Makowska and Zimny 2015).
To overcome albinism many attempts had taken and those empirical studies suggested the calibration of the parameters in in vitro anther culture. Among those, donor plant culturing environment, the growth stage of micro-spore, pretreatment, culture method, and media composi-tion is notable (
Makowska and Zimny 2015). Similarly,
Sriskandarajah et al. (2015) stated that optimization of the culture method will help to reduce this phenomenon, as it has a genotypic dependency.
For instance, cytokinins like thidiazuron and dicamba are used in induction media, and meta-topoline is utilized in regeneration media in barley microspore culture. The result showed that albino plant achievement dwindled signifi-cantly (
Esteves et al. 2014). Afterward, Calic
et al. (2012), showed that abscisic acid has a significant role in achieving green plant over the albino plant in horse chestnut (
Aesculus hyppocastanum). Then, Mannitol along with colchicine or DMSO gave a higher number of green plantlets and DH plants compared to the starvation solely induced by mannitol (
Soriano et al. 2007;
Echávarri and Cistué 2016).
As mentioned, there is no clear and in-depth under-standing of the albino plant formation in androgenesis.
Makowska and Oleszczuk (2014) stated the plastid behav-ior during androgenesis. They inferred that due to the failure of the reprogramming from gametophyte to sporo-phyte, chloroplast might not develop. Likewise, they inferred that early-stage microspore might reduce the albino plant but no findings ossified this assumption. Then, still, it is unknown when the plastid genome was modified that leads to albino plants (
Makowska and Oleszczuk 2014). A few numbers of QTL had identified in the nuclear genome which has a direct effect on the green and albino plant ratio. Still now this number is very trivial (
Yamagishi et al. 1998;
Makowska and Oleszczuk 2014).
So, it is evident that unfolding the mechanism of albino plant formation is the important part. This could forward the way of androgenesis research. Notably, researchers are trying to explain it from the epigenetic aspect. In the future, the extensive epigenetic study will give pace to fathom albinism.
CONCLUSION
Androgenesis is successfully exploited for more than 5 decades and many haploid and doubled haploid plants had regenerated for basic and applied research. Most impor-tantly, various crop cultivars (barley, wheat, rice,
Brassica, etc) have been cultivating around the world which has de-veloped through androgenesis. By 2050, the world popula-tion will be around 9.7 billion (
https://www.worldometersinfo/world-population/). Additionally, the intensity of bio-tic and abiotic stresses will threaten the global food produc-tion. To tackle these issues, a more updated bio-fortified, climate-resilient, disease, and pest-resistant cultivar is needed. Double haploid developed through androgenesis will be one of the important tools to achieve it. The haploid and double haploid will be con-tinued to contribute to gene mapping, QTL mapping, Genome-Wide Association Study (GWAS), mutation breed-ing, Marker Assisted Selection (MAS), Marker-Assisted Backcross (MAB) breeding. Similarly, in genetic transfor-mation and genome editing, it will be exploited, especially in the lines where the zygotic embryo is recalcitrant to receive the trans-gene or sgRNA. Finally, to dissect the phenomenon of albinism, the scien-tist will try to utilize epigenetics extensively along with the existing approaches. Finally, cutting-edge techniques like the reverse genetic approach, genome editing especially CRISPR/Cas9 will be a game-changer.
Fig. 1Schematic diagram showing the anther culture technique.
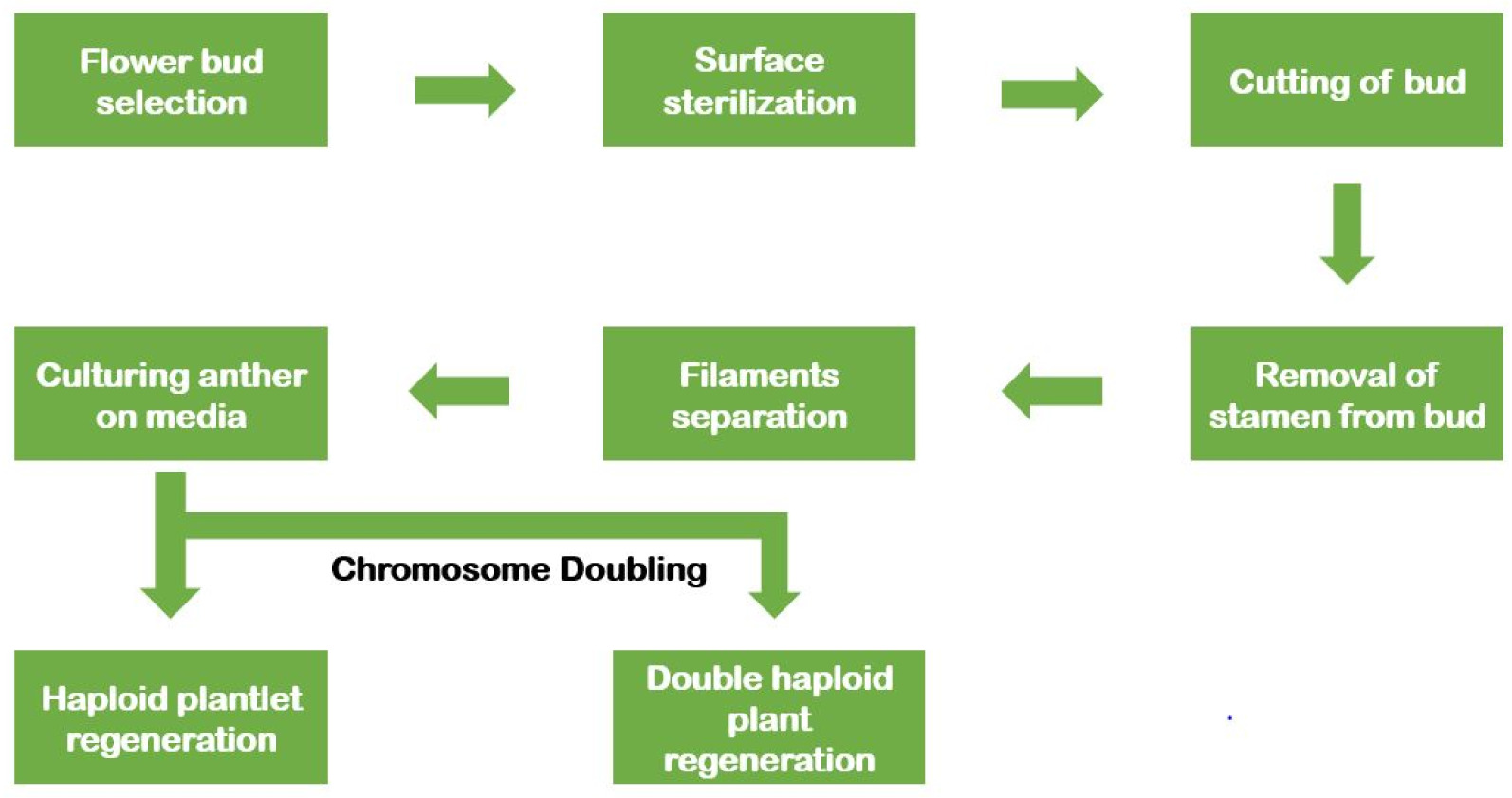
Fig. 2Embryogenic calli (a), Embryo Like Structure (ELS), (b) and plant regenerating (c) from ELS of rice anther culture (
Guzmán and Arias 2000); ELS (d, e), regenerating plantlets (f), regenerated plantlet (g), haploid (n) and double haploid (2n) wheat plant (h) by anther culture (
Rubtsova et al. 2013); green and albino plantlet achieved through barley anther culture (i) (
Sriskandarajah et al. 2015); Segregating green and albino plantlet found by tobacco anther culture (j) (
Dunwell 2010).

Table 1List of rice varieties developed through anther culture having stress tolerance and other agronomic special features (Adopted from Sarao and Gosal et al. 2018).
Table 1
|
Variety name |
Characteristics |
Country |
Reference |
|
Huayu I, Huayu II, Xin Xiu, Late Keng 959, Tunghua 1, Tunghua 2, Tunghua 3, Zhonghua 8, Zhonghua 9, Huahanzao, Huajian 7902, Tanghuo 2, Shanhua 7706, Huahanzao 77001, Nanhua 5, Noll, Hua 03 |
Rice blast and bacterial blight resistance along with quality grain |
China |
Zang 1980; Hu and Zeng 1984; Chen 1986; Loo and Xu 1986; Yang and Fu 1989
|
|
Guan 18 |
Early maturity; good quality and disease resistance |
China |
, Zhu and Pan 1990
|
|
Huayu 15 |
Resistant to lodging and diseases; good quality |
China |
, Shouyi and Shouyin 1991
|
|
Milyang 90 |
Brown planthopper and stripe rust-resistant with better grain quality |
China |
, China Chung 1987
|
|
Hwacheongbyeo, Joryeongbyeo, Hwajinbyeo |
Resistant to brown planthopper, rice stripe tenui virus, blast, and bacterial blight |
China |
, Lee et al. 1989
|
|
Risabell |
High milling and cooking quality; resistant to blast |
India |
, Pauk et al. 2009
|
|
CR Dhan 10 (CRAC2221-43), Satyakrishna |
Resistant to neck blast, sheath-rot, and yellow stem borer |
India |
CRRI Annual Report 2007–2008; Rao and Gosal 2018 |
|
CR Dhan 801 (CRAC2224-1041, IET18720), Phalguni |
Resistant to leaf blast, gall midge; moderately resistant to sheath rot, rice stripe tenuivirus, yellow stem borer, brown spot, and sheath blight |
China |
CRRI Annual Report 2009–2010;
Rao and Gosal 2018 |
Table 2Double haploid technology in wheat breeding.
Table 2
|
Application |
Trait |
Reference |
|
Development of cultivars |
Improving flour quality, higher molecular weight glutenin, rust resistance |
Ushiyama T 2008; Touraev et al. 2009; Bakhshi et al. 2012
|
|
Mapping quantitative trait loci |
Foliar disease of wheat, important agronomic traits, Fusarium head blight, number of tillers, number of spikes per plant and spike length, leaf rust resistance, photoperiod response, heading time, crossability among different wheat genome |
, Chu et al. 2008a, b; Yang et al. 2005; Li et al. 2010; Suenaga et al. 2005; Huang et al. 2003; Sourdille et al. 2000; Tixier et al. 1998
|
|
Production of transgenic plants |
Salt tolerance, drought tolerance |
, Chauhan and Khurana 2010, 2011; Khurana et al. 2011 |
Table 3List of Brassica species with mapping traits using double haploid technology.
Table 4Haploid and double haploid technology in tobacco breeding.
Table 4
|
Technology |
Application |
Reference |
|
Haploid |
Genome mapping, identification of quantitative trait loci (QTL), genetic transformation, marker assisted selection in breeding for new cultivars |
, Wernsman 1993; Tai 2005; Milla et al. 2005
|
|
Haploid & double haploid |
Environment-friendly F1 hybrid system |
, Ribarits et al. 2007
|
References
- Abdollahi MR, Moieni A, Mousavi AA, Salmanian AH. 2011. High frequency production of rapeseed transgenic plants via combination of microprojectile bombardment and secondary embryogenesis of microspore-derived embryos. Mol Biol Rep.. 38(2): 711-719.
- Abdollahi MR, Moieni A, Salmanian AH, Mousavi A. 2009. Secondary embryogenesis and transient expression of the b-glucuronidase gene in hypocotyls of rapeseed micros-pore derived embryos. Biol Plant.. 53: 573-577.
- Agache S, Bechelier B, De Buyser J, Henry Y, Snape J. 1989. Genetic analyses of anther culture response in wheat using aneuploid, chromosome substitution and translocation lines. Theor Appl Genet.. 77: 7-11.
- Ai SL, Chen ZJ. 1981. Genetic studies on tobacco breeding with haploid developed by anther culture. I ch'uan hsueh pao.= Acta genetica Sinica..
- Ajisaka H, Kuginuki Y, Shiratori M, Ishiguro K, Enomoto S, Hirai M. 1999. Mapping of loci affecting the culturing efficiency of microspore culture of Brassica rapa L. syn. Campestris L. using DNA polymorphism. Breed Sci. 49: 187-192.
- Alemanno L, Guiderdoni E. 1994. Increased doubled haploid plant regeneration from rice (Oryza sativa L.) anthers cultured on colchicine-supplemented media. Plant Cell Rep.. 13(8): 432-436.
- Al-Yasiri S, Rogers OM. 1971. Attempting chemical induction of haploidy using toluidine blue. J Am Soc Hortic Sci.. 96(1): 126-127.
- An D, Su J, Liu Q, Zhu Y. Mapping QTLs for nitrogen uptake in relation to the early growth of wheat (Triticum aestivum L.). Plant Soil. 284(1):pp. 73-84.
- Anagnostakis SL. 1974. Haploid plants from anthers of tobacco—enhancement with charcoal. Planta 281-283..
- Andersen SB. 2005. Haploids in the improvement of woody species. In: Palmer CE, Keller WA, Kasha K (eds) Hap-loids in crop improvement II, vol 56. Springer. Heidelberg 243-257:
- Asif MA, Schilling RK, Tilbrook J, Brien C, Dowling K, Rabie H, et al. 2018. Mapping of novel salt tolerance QTL in an Excalibur× Kukri doubled haploid wheat population. Theor Appl Genet.. 131(10): 2179-2196.
- Asker S. 1983. A monopolid of Potentilla argentea. Hereditas. 99(2): 303-304.
- Babbar SB, Agarwal PK, Sahay S, Bhojwani SS. 2004. Isolated microspore culture of Brassica: an experimental tool for developmental studies and crop improvement. Indian J Plant Biotechnol.. 3: 185-202.
- Bai HS, Wang BH, Ge MF. 1991. Study on purification of Minghui 63. Crop Res. 5(1): 10-11 (in Chinese).
- Baillie AMR, Epp DJ, Hucheson D, Keller WA. 1992. In vitro culture of isolated microspores and regeneration of plants in Brassica campestris. Plant Cell Rep.. 11: 234-237.
- Bakhshi T, Bozorgipour R, Mostafavi K, Kashani HH. 2012. Wheat yellow rust resistance improvement in wheat and maize cross progenies using double haploid method. Scient Res Essa.. 7: 2708-2712.
- Ball ST, Zhou HP, Konzak CF. 1993. Influence of 2,4-D, IAA, and duration of callus induction in anther cultures of spring wheat. Plant Sci.. 90(2): 195-200.
- Pfahler PL. Barnabás BKovács G1991. Direct effect of colchicine on the microspore embryogenesis to produce dihaploid plants in wheat (Triti-cum aestivum L.). Theor Appl Genet.. 81: 675-678.
- Barnabás B.2003. Protocol for producing doubled haploid plants from anther culture of wheat (Triticum aestivum L. ). In Doubled haploid production in crop plants. Springer. Dordrecht 65-70.:
- Barro F, Martin A. 1999. Response of different genotypes of Brassica carinata to microspore culture. Plant Breed.. 118: 79-81.
- Belogradova K, Lewicka I, Heberle-Bors E, Touraev A. 2009. An overview on tobacco doubled haploids. Advances in haploid production in higher plants 75-85..
- Bettey M, Finch-Savage WE, King GJ, Lynn JR. 2000. Quan-titative genetic analysis of seed vigour and pre-emer-gence seedling growth traits in Brassica oleracea. New Phytol.. 148(2): 277-286.
- Bhatia R, Dey SS, Parkash C, Sharma K, Sood S, Kumar R. 2018. Modification of important factors for efficient microspore embryogenesis and doubled haploid pro-duction in field grown white cabbage (Brassica oleracea var. capitata L. ) genotypes in India. SciHortic. 233: 178-187.
- Bhatia R, Sharma K, Parkash C, Pramanik A, Singh D, Singh S, et al. 2021. Microspore derived population developed from an inter-specific hybrid (Brassica oleracea × B. carinata) through a modified protocol provides insight into B genome derived black rot resistance and inter- genomic interaction. Plant Cell Tissue Organ Cult.. 145(2): 417-434.
- Bhowmik P, Ellison E, Polley B, Bollina V, Kulkarni M, Ghanbarnia K, et al. 2018. Targeted mutagenesis in wheat microspores using CRISPR/Cas9. Sci Rep.. 8(1): 1-10.
- Bilichak A, Sastry‐Dent L, Sriram S, Simpson M, Samuel P, Webb S, et al. 2020. Genome editing in wheat micro-spores and haploid embryos mediated by delivery of ZFN proteins and cell‐penetrating peptide complexes. Plant Biotechnol J.. 18(5): 1307-1316.
- Blakeslee AF, Belling J, Farnham ME, Bergner AD. 1922. A haploid mutant in the Jimson weed, Datura stramonium. Science. 55: 646-647.
- Borisjuk N, Kishchenko O, Eliby S, Schramm C, Anderson P, Jatayev S, et al. 2019. Genetic modification for wheat improvement: from transgenesis to genome editing. Biomed Res Int..
- Bosemark NO. 1971. Haploids and homozygous diploids, triploids and tetraploids in sugar beet. Hereditas. 69(2): 193-203.
- Bourgin JP, Nitsch JP. 1967. Obtention de Nicotiana haploids à partir détamines cultivées in vitro. Ann Physiol.. 9: 377-382.
- Brandt KM, Gunn H, Moretti N, Zemetra RS. 2020. A Stream-lined Protocol for Wheat (Triticum aestivum) Protoplast Isolation and Transformation With CRISPR-Cas Ribonu-cleoprotein Complexes. Front Plant Sci.. 11: 769
- Brew-Appiah RAT, Ankrah N, Liu W, Konzak CF, Von Wettstein D, Rustgi S. 2013. Generation of doubled hap-loid transgenic wheat lines by microspore transformation. PLoS One. 8: e80155
- Broughton S, Castello M, Liu L, Killen J, Hepworth A, O'learly R. 2020. The effect of Caffeine and Triflurain on chromosome doubling in wheat anther culture. Plants. 9: 105
- Broughton S. 2008. Ovary co-culture improves embryo and green plantlets production in anther culture of Australian spring wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult.. 95: 185-195.
- Broughton S. 2011. The application of n-butanol improves embryo and green plant production in anther culture of Australian wheat (Triticum aestivum L.) genotypes. Crop Pasture Sci.. 62: 813-822.
- Broughton S, Sidhu PK, Davies PA. 2014. In vitro culture for doubled haploids: tools for molecular breeding. Methods Mol Biol.. 1145: 167-189.
- Burbulis N, Malinauskaite R, Kott L. 2001. Oil quality im--provement through in vitro mutagenesis and haploid selection in Lithuanian winter Brassica napus germplasm. Proc Latv Acad Sci.. 55: 197-200.
- Ćalić D, Bohanec B, Devrnja N, Milojević J, Tubić L, Kostić I, et al. 2013. Impact of abscisic acid in overcoming the problem of albinism in horse chestnut androgenic embryos. Trees. 27(3): 755-762.
- Caredda S. Clément C.1999. Androgenesis and albinism in Poaceae: influence of genotype and carbohydrates. In Anther and Pollen. Springer. Berlin, Heidelberg 211-228.:
- Caredda S, Devaux P, Sangwan RS, Proult I. Clément C2004. Plastid ultrastructure and DNA related to albinism in androgenetic embryos of various barley (Hordeum vulgare L.) cultivars. Plant Cell Tissue Organ Cult.. 76(1): 35-43.
- Caredda S, Doncoeur C, Devaux P, Sangwan RS. Clément C2000. Plastid differentiation during androgenesis in albino and non-albino producing cultivars of barley (Hordeum vulgare L.). Sex Plant Reprod.. 13(2): 95-104.
- Carl CM Jr, Yawney HW. 1972. Multiple seedlings in Acer saccharum. J Torrey Bot 142-144..
- Castillo AM, Sanchez-Diaz RA, Valles MP. 2015. Effect of ovary induction on bread wheat anther culture: ovary genotype and developmental stage, and candidate gene association. Front Plant Sci.. 6: 402
- Castillo AM, Valles MP. Cistué L2000. Comparison of anther and isolated microspore cultures in barley. Effects of culture density and regeneration medium. Euphytica. 113(1): 1-8.
- Cegielska-Taras T, Pniewski T, Szala L. 2008. Transforma-tion of microspore-derived embryos of winter oilseed rape (Brassica napus L.) by using Agrobacterium tume-faciens. J Appl Genet.. 49: 343-347.
- Central Rice Research Institute (CRRI) Annual Report2007.
- Central Rice Research Institute (CRRI) Annual Report2009.
- Chase SS. 1969. Monoploids and monoploid-derivatives of maize (Zea mays L.). Bot Rev.. 35(2): 117-168.
- Chauhan H, Khurana P. 2010. Development of drought tolerant transgenic doubled haploid in wheat through Agrobacterium-mediated transformation. Plant Biotech J. 9. 408-417.:
- Chauhan H, Khurana P. 2011. Use of doubled haploid technology for development of stable drought tolerant bread wheat (Triticum aestivum L.) transgenics. Plant Biotechnol J.. 9(3): 408-417.
- Chen CC, Tsay HS, Huang CR. 1991. Factors affecting andro-genesis in rice (Oryza sativa L.). Bajajv YPS (ed) Rice. Bio-technology in Agriculture and Forestry. Springer, Berlin, Heidelberg, New York. 14: 192-215.
- Chen JJ, Tsay HS. 1986. The callus forming ability of rice anther derived from tillers of different orders and from spikelets of different positions and branches in the panicle. J Agric Res China.. 33: 354-362.
- Chen QF, Wang CL, Lu YM, Shen M, Afza R, Duren MV. et al2001. Anther culture in connection with induced muta-tions for rice improvement. Euphytica. 120(3): 401-408.
- Chen C, Xiao H, Zhang W, Wang A, Xia Z, Li X, et al. 2006. Adapting rice anther culture to gene transformation and RNA interference. Science in China Series C: Life Sci. 49(5): 414-428.
- Cho MC, Chang KY. 1984. Genetic studies on the dihaploids made from anther culture of the F1 of aromatic tobacco (N. tobacum L.). Korean J Breed Sci..
- Chu CC, Wang CC, Sun CS, Chien NF, Yin KC, Hsu C. 1973. Investigations on the induction and morphogenesis of wheat (Triticum aestivum) pollen plants. Acta Bot Sin.. 15: 1-11.
- Chu CG, Friesen TL, Xu SS, Faris JD. 2008a. Identification of novel tan spot resistance loci beyond the known host- selective toxin insensitivity genes in wheat. Theor Appl Genet. 117. 873-881.:
- Chu CG, Xu SS, Friesen TL, Faris JD. 2008b. Whole genome mapping in a wheat doubled haploid population using SSRs and TRAPs and the identification of QTL for agronomic traits. Mol Breed. 22. 251-256.:
- Chung GS. 1987. Application of anther culture technique for rice (Oryza sativa L.) improvement. In: Korea-China Plant Tissue Culture Symposium. Beijing. China: Academia Sinica Publishers 36-56.
- Chuong PV, Beversdorf WD. 1985. High frequency embryo-genesis through isolated microspore culture in Brassica napus L. and B. carinata Braun. Plant Sci. 39: 219-226. 219-226.
- Chuong PV, Deslauriers C, Kott LS, Beversdorf WD. 1988. Effects of donor genotype and bud sampling on micro-spore culture of Brassica napus. Can J Bot.. 6: 1653-1657.
- Cloutier S, Cappadocia M, Landry BS. 1995. Study of micro-spore-culture responsiveness in oilseed rape (Brassica napus L.) by comparative mapping of a F2 population and two microspore derived populations. Theor Appl Genet.. 91: 841-847.
- Craig IL. 1974. Haploid plants (2n = 21) from in vitro anther culture of Triticum aestivum. Can J Genet Cytol.. 16: 697-700.
- Cuthbert JL, Somers DJ, Brown PD, Crow GH. Brûlé-Babel AL2008. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor Appl Genet.. 117(4): 595-608.
- Da Silva Dias JC. 1999. Effect of activated charcoal on Brassica oleracea microspore culture embryogenesis. Euphytica. 108: 65-69.
- Sharp WR. DA.Yamada Y (eds) Handbook of plant cell culture: III Crop Species.Macmillan, New York 65-90. Macmillan. New York 65-90.
- Daoru T, Xinghua J. 1991. Mutant in tobacco anther culture induced by sup 60 Co. gamma-rays. Henong Xuebao (Acta Agriculturae Nucleatae Sinica); (China). 5(4)..
- Datta SK, Wenzel G. 1998. Single microspore derived emb-ryogenesis and plant formation in barley (Hordeum vulgare). Single microspore derived emb-ryogenesis and plant formation in barley (Hordeum vulgare). Archiv für zuechtungsforschung 18: 125-131. 125-131.
- Datta SK. 2001. Androgenesis in cereals. In: Bhojwani SS, Soh WY Current Trends in the Embryology of Angio-sperms. Netherlands: Springer 471-488.
- Datta SK. 2005. Androgenic haploids: factors controlling development and its application in crop improvement. Curr Sci.. 89: 1870-1878.
- Datta SK, Wenzel G. 1987. Isolated microspore derived plant formation via embryogenesis in Triticum aestivum L. Plant Sci.. 48: 49-54.
- De Buyser J, Henry Y, Lonnet P, Herzog R, Hespel A. 1987. Florin - a doubled haploid wheat variety developed by the anther culture method. Plant Breed.. 98: 53-56.
- Devaux P. 2003. The Hordeum bulbosum (L. ) method. In Doubled haploid production in crop plants. Springer. Dordrecht 15-19.:
- Devaux P, Pickering R. 2005. Haploids in the improvement of Poaceae. In Haploids in crop improvement II. Springer. Berlin, Heidelberg 215-242.:
- Dewi IS, Purwoko BS, Aswidinnoor H, Somantri IH, Chozin MA. 2009. Plant regeneration from anther cultures of several genotypes of indica rice tolerant to aluminum toxicity. Indones J Agric Sci.. 2(1): 1-5.
- Dhaka N, Rout K, Yadava SK, Sodhi YS, Gupta V, Pental D, et al. 2017. Genetic dissection of seed weight by QTL analysis and detection of allelic variation in Indian and east European gene pool lines of Brassica juncea. Theor Appl Genet.. 130(2): 293-307.
- Dormann M, Wang HM, Oelck M. 2001. Transformed embryo-genic microspores for the generation of fertile homozy-gous plants. USA Patent US 6. 316,694 B1..
- Duarte-Aké F, Castillo-Castro E, Pool FB, Espadas F, Santamaría JM, Robert ML, et al. 2016. Physiological differences and changes in global DNA methylation levels in Agave angustifolia Haw. albino variant somaclones during the micropropagation process. Plant Cell Rep.. 35(12): 2489-2502.
- Dubová J.1996. The haploid embryoids development in anther culture of transformed tobacco (Nicotiana tabacum L. ). Acta Soc Bot Pol. 65(1-2): 31-35..
- Pochard E. Dumas de Vaulx R.1974. Effect of nitrous oxide on the induction of haploidy in the garden pepper (Capsicum annum L. ). In Annales de l'Amelioration des Plantes..
- Duncan EJ, Heberle-Bors E. 1976. Effect of temperature shock on nuclear phenomena in microspores of Nicotiana tabacum and consequently on plantlet production. Protoplasma. 90: 173-177.
- Dunford R, Walden RM. 1991. Plastid genome structure and plastid-related transcript levels in albino barley plants derived from anther culture. Curr Genet.. 20(4): 339-347.
- Dunwell JM. 1986. Pollen, ovule and embryo culture, as tools in plant breeding. In: Withers LA, Alderson PG (eds) Plant tissue culture and its agricultural applications. Butterworths. London 375-404:
- Dunwell JM. 1976. A comparative study of environmental and developmental factors which influence embryo induction and growth in cultured anthers of Nicotiana tabacum. Environ Exp Bot. 16(2-3): 109-118..
- Dunwell JM. 1981. Stimulation of pollen embryo induction in tobacco by pretreatment of excised anthers in a water- saturated atmosphere. Plant Sci Lett.. 21(1): 9-13.
- Dunwell JM. 2010. Haploids in flowering plants: origins and exploitation. Plant Biotechnol J.. 8(4): 377-424.
- Echávarri B, Cistué L2016. Enhancement in androgenesis efficiency in barley (Hordeum vulgare L.) and bread wheat (Triticum aestivum L. ) by the addition of dimethyl sulfoxide to the mannitol pretreatment medium. Plant Cell Tissue Organ Cult.. 125: 11-22.
- Echávarri B, Cistué L2016. Enhancement in androgenesis efficiency in barley (Hordeum vulgare L.) and bread wheat (Triticum aestivum L. ) by the addition of dimethyl sulfoxide to the mannitol pretreatment medium. Plant Cell Tissue Organ Cult.. 125(1): 11-22.
- Ekiz H, Konzak CF. 1991a. Nuclear and cytoplasmic control of anther culture response in wheat: I. analyses of all-oplasmic lines. Crop Sci.. 31: 1421-1427.
- Ekiz H, Konzak CF. 1991b. Nuclear and cytoplasmic control of anther culture response in wheat: III. Common wheat crosses. Crop Sci.. 31: 1432-1436.
- Elhaddoury J, Lhaloui S, Udupa SM, Moatissim B, Taiq R, Rabeh M, et al. 2012. Registration of 'Kharoba': A bred wheat cultivar developed through doubled haploid breed-ing. J. Plant Regist.. 6: 169-173.
- Esteves P, Clermont I, Marchand S, Belzile F. 2014. Im-proving the efficiency of isolated microspore culture in six-row spring barley: II-exploring novel growth regulators to maximize embryogenesis and reduce albinism. Plant Cell Rep.. 33(6): 871-879.
- Evans DA, Sharp WR, Medina-Filho HP. 1984. Somaclonal and gametoclonal variation. Am J Bot.. 71: 759-774.
- Faruq G, Shamsuddin F, Jenifer A, Jusoh SH, Nezhadahmadi A, Khalid N. 2014. Optimization of media and cold pre-treatment for anther culture using japonica/indica and indica/indica hybrids and their callus induction com-parisons in different rice crosses. Indian J Sci Technol.. 7(11): 1861-1870.
- Ferrie AMR. Möllers C2011. Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tissue Organ Cult.. 104: 375-386.
- Ferrie AMR, Taylor DC, Mackenzie SL, Keller WA. 1999. Microspore embryogenesis of high sn-2 erucic acid Bras-sica oleracea germplasm. Plant Cell Tissue Organ Cult.. 57: 79-84.
- Ferrie AMR, Taylor DC, MacKenzie SL, Rakow G, Raney JP, Keller WA. 2008. Microspore mutagenesis of Brassica species for fatty acid modifications: a preliminary evalua-tion. Plant Breed.. 127: 501-506.
- Ferrie AM. Möllers C2011. Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tissue Organ Cult.. 104(3): 375-386.
- Forster BP, Thomas WTB. 2005. Doubled haploids in genetics and plant breeding. Plant Breed Rev.. 25: 57-88.
- Forster BP, Herberle-Bors E, Kasha KJ, Touraev A. 2007. The resurgence of haploids in higher plants. Trends Plant Sci.. 12(8): 368-375.
- Fukuoka H, Ogawa T, Matsuoka M, Ohkawa Y, Yano H. 1998. Direct gene delivery into isolated microspores of rapeseed (Brassica napus L.) and the production of fertile transgenic plants. Plant Cell Rep.. 17: 323-328.
- Furusho M, Suenaga K, Nakajima K. 1991. Production of haploid barley from barley × maize and barley × Italian ryegrass crosses. Jpn J Breed.. 41: 175-179.
- Garcia Cruz H, Garcia V, Pino LA, Espino E. 1999. 'Burley Habana-13'('BH-13') first commercial tobacco variety obtained in Cuba by anther culture resistant to blue mold (Peronospora tabacina), black shank (Phytophthora parasitica variety nicotianae) and Tobacco Mosaic Virus (TMV). Cubatabaco. Ciencia y Tecnica (Cuba)..
- GENCER AS. 2002. Developing Tobacco Lines Resistant to Powdery Mildew (Erysiphe cichoracearum L.) by Anther Culture Technique for the Aegean Region. Turk J Agric For.. 26(2): 63-69.
- Germanà MA2011. Anther culture for haploid and doubled haploid production. Plant Cell Tissue Organ Cult.. 104(3): 283-300.
- Chiancone B. Germanà MA2001. Gynogenetic haploids of Citrus after in vitro pollination with triploid pollen grains. Plant Cell Tissue Organ Cult.. 66(1): 59-66.
- Germanà MA.1997. Haploidy in Citrus. In: Jain SM, Sopory SK, Veilleux RE (eds) In vitro haploid production in higher plants, vol 5. Kluwer. Dordrecht 195-217:
- Germanà MA2006. Doubled haploid production in fruit crops. Plant Cell Tissue Organ Cult.. 86: 131-146.
- Germanà MA.2007. Haploidy. In: Khan I (ed) Citrus. Gene-tics, breeding and biotechnology. CABI. Wallingford 167-196:
- Germanà MA.2009. Haploid and doubled haploids in fruit trees. In: Touraev A, Forster B, Jain M (eds) Advances in haploid production in higher plants. Springer. Heidelberg 241-263:
- Ghaemi M, Sarrafi A, Morris R. 1995. Reciprocal substitu-tions analyses of embryo induction and plant regenera-tion from anther culture in wheat (Triticum aestivum L.). Genome. 38: 158-165.
- Gil-Humanes J, Barro F. 2009. Production of doubled hap-loids in Brassica. In: Touraev et al (eds) Advances in haploid production in higher pants. Springer. New York 65-73:
- Graf RJ, Hucl P, Orshinsky BR, Kartha KK. 2003. "McKenzi" hard red spring wheat. Can J Plant Sci.. 83: 565-569.
- Grewal D. 2009. Doubled haploids: from laboratory to field. Rice Today. 8: 46.
- Guerche P, Charbonnier M, Jouanin L, Tourneur C, Paszkowski J, Pelletier G. 1987. Direct gene transfer by electropora-tion in Brassica napus. Plant Sci.. 52: 111-116.
- Guha S, Maheshwari SC. 1964. In vitro production of embryos from anthers of Datura. Nature. 204: 497-498.
- Guo YD, Pulli S. 1996. High-frequency embryogenesis in Brassica campestris microspore culture. Plant Cell Tissue Organ Cult.. 46: 219-225.
- Arias FJZ. Guzmán M2000. Increasing anther culture ef-ficiency in rice (Oryza sativa L.) using anthers from ratooned plants. Plant Sci.. 151(2): 107-114.
- Hai L, Guo H, Xiao S, Jiang G, Zhang X, Yan C, et al. 2005. Quantitative trait loci (QTL) of stem strength and related traits in a doubled-haploid population of wheat (Triticum aestivum L.). Euphytica. 141(1): 1-9.
- Haliloglu K, Baenziger PS, Mitra A. 2004. Genetic transfor-mation of wheat (Triticum aestivum L.) anther culture- derived embryos by electroporation. Biotechnol Biotechnol Equip.. 18(2): 62-68.
- Han Y, Broughton S, Liu L, Zhang XQ, Zeng J, He X, et al. 2021. Highly efficient and genotype-independent barley gene editing based on anther culture. Plant Commun.. 2(2): 100082
- Haque E, Taniguchi H, Hassan M, Bhowmik P, Karim MR, Śmiech M, et al. 2018. Application of CRISPR/Cas9 genome editing technology for the improvement of crops cultivated in tropical climates: recent progress, prospects, and challenges. Front Plant Sci.. 9: 617
- Harland SC. 1920. A note on a peculiar type of "rogue" in Sea Island cotton. Agr. News, Barbados. 19: 29.
- Harland SC. 1936. Haploids in polyembryonic seeds of Sea Island cotton. J Hered.. 27: 229-231.
- Harland SC. 1955. Plant Breeding: Present Position and Future Perspective. Third Bateson Lecture. Cambridge: University Press 15..
- He T, Yang Y, Tu SB, Yu MQ, Li XF. 2006. Selection of interspecific hybrids for anther culture of indica rice. Plant Cell Tissue Organ Cult.. 86: 271-277.
- Heidari B, Saeidi G, Tabatabaei BES, Suenaga K. 2012. QTLs involved in plant height, peduncle length and heading date of wheat (Triticum aestivum L.). J Agric Sci Technol.. 14(5): 1093-1104.
- Hittalmani S, Shashidhar HE, Bagali PG, Huang H, Sidhu JS, Singh VP, et al. 2002. Molecular mapping of quantitative trait loci for plant growth, yield and yield related traits across three diverse locations in a doubled haploid rice population. Euphytica. 125(2): 207-214.
- Ho KM, Jones GE. 1980. Mingo barley. Can J Plant Sci.. 60: 279-280.
- Hochholdinger F, Hoecker N. 2007. Towards the molecular basis of heterosis. Trends Plant Sci.. 12: 427-432.
- Hofinger BJ, Ankele E, Heberle-Bors E, Pfosser MF. Gülly Ch.2000. The involvement of the plastid genome in albino plant regeneration from microspores in wheat. In: Bohanec B (ed) Biotechnological approaches for utili-zation of gametic cells-COST 824. OP-EUR. Luxembourg 215-228.
- Horner M, McComb JA, McComb AJ, Street HE. 1977. Ethylene production and plantlet formation by Nicotiana anthers cultured in the presence and absence of charcoal. J Exp Bot.. 28: 1365-1372.
- Hu H. Yang HY (eds).1986. Haploids in higher plants in vitro. China Academic Publishers/Springer. Beijing/Berlin..
- Hu H, Zeng JZ. 1984. Development of new varieties via anther culture. In: Ammirato PV. Evans.
- Hu DF, Yuan ZD, Tang Y, Liu JP. 1985. Jinghua No 1., a winter wheat variety derived from pollen sporophyte. Sci Sin.. 28: 733-745.
- Hu H. 1997. In vitro induced haploids in wheat. In In vitro haploid production in higher plants Springer. Dordrecht 73-97.:
- Huang B. 1996. Gametoclonal variation in crop improvement. In: Jain SM, Sopory SK, Veilleux RE (eds) In vitro haploid production in higher plants, vol 2. Kluwer. Dordrecht 73-91:
- Huang L, Brooks SA, Li WL, Fellers JP, Trick HN, Gill BS. 2003. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics 164. 655-664.:
- Hunter CP. 1987. Plant regeneration method. Europen patent, 1987. Application No. 872007737.
- Imamura J, Harada H. 1981. Stimulation of tobacco pollen embryogenesis by anaerobic treatments. Z Pflanzenphysiol.. 103(3): 259-263.
- Immonen S, Anttila H. 1996. Success in rye anther culture. Votr Pflanzenzuchtg. 35: 237-244.
- Jacquard C, Asakaviciute R, Hamalian AM, Sangwan RS, Devaux P, Clement C. 2006. Barley anther culture: Effects of annual cycle and spike position on microspore em-bryogenesis and albinism. Plant Cell Rep.. 25(5): 375-381.
- Jagannath A, Bandyopadhyay P, Arumugam N, Gupta V, Burma PK, Pental D. 2001. The use of a spacer DNA fragment insulates the tissue specific expression of a cytotoxic gene (barnase) and allows high frequency generation of transgenic male sterile lines in Brassica juncea L. Mol Breed.. 8: 11-23.
- Jain SM, Sopory SK, Veilleux RE, 1996-1997. editors. In vitro haploid production in higher plants. Kluwer. Dordrecht: pp. 1-5.
- Jansing J, Sack M, Augustine SM, Fischer R, Bortesi L. 2019. CRISPR/Cas9‐mediated knockout of six glycosyltrans-ferase genes in Nicotiana benthamiana for the production of recombinant proteins lacking b‐1, 2‐xylose and core a‐1, 3‐fucose. Plant Biotechnol. J. 17(2): 350-361.
- Jardinaud MF, Alibert G. Souvré A.1993. Transient GUS gene expression in Brassica napus electroporated micro-spores. Plant Sci. 93(1-2): 177-184..
- Jensen CJ. 1983. Producing haploid plants by chromosome elimination. Cell and tissue culture techniques for cereal crop improvement. Science Press. Beijing 55-79..
- Johannsen W. 1903. Uber Erblichkeit in Populationen und in reinen Linien. Gustav Fischer. Jena..
- Johannsen W. 1909. Elemente der exacten Erblichkeitslehre. Gustav Fischer. Jena.:
- Jones-Villeneuve E, Huang B, Prudhomme I, Bird S, Kemble R, Hattori J, et al. 1995. Assessment of microinjection for introducing DNA into uninuclear microspores of rapeseed. Plant Cell Tissue Organ Cult.. 40: 97-100.
- Kameya T, Hinata K. 1970. Induction of haploid plants from pollen grains of Brassica. Jpn J Breed.. 20: 82-87.
- Kanbar OZ, Lantos C, Kiss E, Pauk J. 2020. Androgenic responses of winter wheat combinations in in vitro anther culture. Genetika-Belgrade. 52: 337-350.
- Kao KN. 1981. Plant formation from barley anther cultures with ficoll media. J Plant Physiol.. 103(5): 437-443.
- Kasha KJ (ed).1974. Haploids in higher plants: advances and potential. The Office of Continuing Education. University of Guelph Press, Guelph..
- Kasha KJ, Kao KN. 1970. High frequency haploid production in barley (Hordeum vulgare L.). Nature (London). 225: 874-875.
- Kasha KJ, Simion E, Oro R, Shim YS. 2003b. Barley isolated microspore culture protocol. In: Maluszymski M, Kasha KJ, Forster BP, Szarejko I (eds) Doubled haploid production in crop plants: a manual. Kluwer Academic. Dordrecht 43-48:
- Kasha KJ, Kao KN. 1970. High frequency haploid production in barley (Hordeum vulgare L.). Nature. 225(5235): 874-876.
- Kasha KJ (Ed).Haploids in Higher Plants: Advances and Potential. Proceedings of the First International Symposium, June 10-14. 1974, University of Guelph, Guelph.
- Katayama Y, Nei M. 1964. Studies on the haploidy in higher plant, Rep. Lab. Plant Breed Fac Agr Miyazaki Univ. 2: 1-78.
- Pınar H, Ata A, Yıldız S. Keleş DTaşkın HBüyükalaca S2015. Effect of pepper types on obtaining spontaneous doubled haploid plants via anther culture. HortScience. 50(11): 1671-1676.
- Keller WA, Rajhathy T, Lacapra J. 1975. In vitro production of plants from pollen in Brassica campestris. Can J Genet Cytol.. 17: 655-665.
- Kelm C, Ghaffary SMT, Bruelheide H, Roder MS, Miersch S, Weber WE, et al. 2012. The genetic architecture of seedling resistance to Septoria tritici blotch in the winter wheat doubled-haploid population Solitar × Mazurka. Mol Breed.. 29(3): 813-830.
- Khurana P. Tuteja N, Gill SS, Tubercio AF, Tuteja R, 2011. Functional genomics of abiotic stress tolerance in wheat. editors. Improving crop resistance to abiotic stress. Wiley-Blackwell, Wiley-VCH Verlag GmbH & Co. Germany: pp. 1-43.
- Khush GS, Virmani SS. Jain SM, Sopory SK, Veilleux RE. 1996. Haploids in plant breeding. In vitro haploid production in higher plants, vol 1. Kluwer. Dordrecht: pp. 11-33.
- Kohlenbach W, Wernicke W, Lang H. 1978. Androgenetic development of isolated microspores of Hyoscyamus and Nicotiana [drug plants, tobacco]. In Proceedings of an International Symposium on plant cell culture. Tuebingen. Germany, FR.
- Kostoff D. 1929. An androgenic Nicotiana haploid. Zeit. Zellforschg. 9: 640-642.
- Kott LS, Beversdorf WD. 1990. Enhanced plant regeneration from microspore-derived embryos of Brassica napus by chilling, partial desiccation and age selection. Plant Cell Tissue Organ Cult.. 23: 187-192.
- Kott LS, Polsoni L, Berversdorf WD. 1988. Cytological aspects of isolated microspore culture of Brassica napus. Can J Bot.. 66: 1658-1664.
- Kuchel H, Williams K, Langridge P, Eagles HA, Jefferies SP. 2007. Genetic dissection of grain yield in bread wheat. II. QTL-by-environment interaction. Theor Appl Genet.. 115(7): 1015-1027.
- Kumari M, Clarke HJ, Small I, Siddique KH. 2009. Albinism in plants: a major bottleneck in wide hybridization, androgenesis and doubled haploid culture. Critical Reviews in Plant Sci.. 28(6): 393-409.
- Kunzel G, Korzuna L, Meistera A. 2000. Cytologically inte-grated physical restriction fragment length polymorphism maps for the barley genome based on translocation breakpoints. Genetics. 154: 397-412.
- Lacadena JR. 1974. Spontaneous and induced parthenogenesis and androgenesis. Haploids in higher plants: advances and potential. The University of Guelph. Guelph: pp. 13-32.
- Lantos C, Pauk J. 2016. Anther culture as an effective tool in winter wheat (Triticum aestivum L.) breeding. Russ J Genet.. 52: 794-801.
- Lantos C, Pauk J. Bóna LNagy ÉBékés F2018. Induction of in vitro androgenesis in anther and isolated microspore culture of different spelt wheat (Triticum spelta L.) geno-types. Plant Cell Tissue Organ Cult.. 133: 385-393.
- Lantos C, Purgel S, Ács K, Langó B, Bóna L, Boda K, et al. 2019. Utilization of in vitro anther culture in spelt wheat breeding. Plants. 8: 436
- Lantos C, Weyen J, Orsini JM, Gnad H, Schlieter B, Lein V, et al. 2013. Efficient application of in vitro anther culture for different European winter wheat (Triticum aestivum L.) breeding programs. Plant Breed.. 132: 149-154.
- Lantos , Csaba , János P, et al. 2020. “Factors influencing the efficiency of wheat anther culture.”. Acta Biol Crac Ser Bot.. 2020: 7-16.
- Lapitan PC, Violeta NV. 1999. Anther culture response of some genotypes with varying amylase content to nitrogen fixation. Phill J Crop Sci.. 24(3): 79-84.
- Lashermes P. 1992. Improved anther culture method for ob-taining direct regeneration in wheat (Triticum aestivum L.). J Genet Breed.. 46: 99-102.
- Lazar MD, Baenziger PS, Schaeffer GW. 1984. Combining abilities and heritability of callus formation and plantlet regeneration in wheat (Triticum aestivum L.) anther cultures. Theor Appl Genet.. 68: 131-134.
- Lazaridou TB, Pankou CI, Xynias IN, Roupakias DG. 2017. Effects of 1BL.1RS wheat-rye translocation on the andro-genic response in spring bread wheat. Cytol Genet.. 51: 485-490.
- Lee GH, Yun BW, Kim KM. 2014. Analysis of QTLs associated with the rice quality related gene by double haploid populations. Int J Genom.. 2014: 1-6.
- Lee SY, Lee JH, Kwon TO. 2003. Selection of salt-tolerant doubled haploids in rice anther culture. Plant Cell Tissue Organ Cult.. 74(2): 143-149.
- Lee YT, Lim MS, Kim HS, et al. 1989. An anther derived new high quality variety with disease and insect resistance 'Hwacheong byeo'. Res Rep Rural Dev Admin Rice.. 31: 27-34.
- Li Z, Peng T, Xie Q, Han S, Tian J. 2010. Mapping of QTL for tiller number at different stages of growth in wheat using double haploid and immortalized F2 populations. J Gene. 89: 409-415.
- Li ZK, Yu SB, Lafitte HR, Huang N, Courtois B, Hittalmani S, et al. 2003. QTL × environment interactions in rice: I. Heading date and plant height. Theor Appl Genet.. 108(1): 141-153.
- Li ZHU, Fu YP, Liu WZ, Hu GC, Si HM, Tang KX, et al. 2007. Rapid generation of selectable marker-free trans-genic rice with three target genes by co-transformation and anther culture. Rice Sci.. 14(4): 239-246.
- Lichter R. 1982. Induction of haploid plants from isolated pollen of Brassica napus. Z Pflanzenphysiol.. 105: 427-434.
- Liu S, Wang H, Zhang J, Fitt BDL, Xu Z, Evans N, et al. 2005. In vitro mutation and selection of doubled-haploid Brassica napus lines with improved resistance to Sclerotinia sclerotiorum. Plant Cell Rep.. 24: 133-144.
- Liu W, Zheng MY, Konzak CF. 2001. Improving green plant production via isolated microspore culture in bread wheat (Triticum aestivum L.). Plant Cell Rep.. 20: 821-824.
- Liu H, Wang K, Jia Z, Gong Q, Lin Z, Du L, et al. 2020. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized Agrobacterium-mediated CRISPR system. J Exp Bot.. 71(4): 1337-1349.
- Liu NT, Jane WN, Tsay HS, Wu H, Chang WC, Lin CS. 2007. Chloroplast genome aberration in micropropagation- derived albino Bambusa edulis mutants, ab1 and ab2. Plant Cell Tissue Organ Cult.. 88(2): 147-156.
- Longin CFH, Mi X, Melchinger AE, Reif JC. Würschum T2014. Optimum allocation of test resources and comparison of breeding strategies for hybrid wheat. Theor Appl Genet.. 127: 2117-2126.
- Loo SW, Xu ZH. 1986. Rice: anther culture for rice improvement in China. In: Bajaj YPS (ed) Biotechnology in agriculture and forestry: II. Crops I. Springer. Verlag, Berlin 139-156:
- Ma LY, Bao JS, Guo LB, Zeng DL, Li XM, Ji ZJ, et al. 2009. Quantitative trait loci for panicle layer uniformity iden-tified in doubled haploid lines of rice in two environ-ments. J Integr Plant Biol.. 51(9): 818-824.
- McCallum CM, Comai L, Greene EA, Henikoff S. 2000. Targeting induced locallesions in genomes (TILLING) for plant functional genomics. Plant physiol.. 123(2): 439-442.
- Mago R, Tabe L, McIntosh RA, Pretorius Z, Kota R, Paux E, et al. 2011. A multiple resistance locus on chromosome arm 3BS in wheat confers resistance to stem rust (Sr2), leaf rust (Lr27) and powdery mildew. Theor Appl Genet.. 123(4): 615-623.
- Magoon ML, Khanna KR. 1963. Haploids. Caryologia. 16(1): 191-235.
- Maheshwari SC. 1996. The discovery of anther culture tech-nique for the production of haploids. In In Vitro Haploid Production in Higher Plants. Springer. Dordrecht 1-10.:
- Makowska K, Oleszczuk S. 2014. Albinism in barley andro-genesis. Plant Cell Rep.. 33(3): 385-392.
- Makowska K, Zimny J. 2015. Albinism-common phenomenon during cereal androgenesis. BioTechnologia 96(1)..
- Maluszynski M, Kasha KJ, Forster BP. Szarejko I (eds).2003a. Doubled haploid production in crop plants: a manual. Kluwer. Dordrecht.:
- Maluszynski M, Kasha KJ, Szarejko I. 2003b. Published doublehaploid protocols in plant species. In: Maluszynski M, Kasha KJ, Forster BP, Szarejko I (eds) Haploid production in crop plants: a manual. Kluwer. Dordrecht 309-335:
- Maluszynski M, Szarejko I, Barriga P, Balcerzyk A. 2001. Heterosis in crop mutant crosses and production of high yielding lines using doubled haploid systems. Euphytica. 120: 387-398.
- Mandal N, Gupta S. 1995. Effect of genotype and culture medium on androgenic callus formation and green plant regeneration in indica rice. Indian J Exp Biol.. 33: 761-765.
- McClinchey SL, Kott LS. 2008. Production of mutants with high cold tolerance in spring canola. Euphytica. 162: 51-67.
- Milla SR, Levin JS, Lewis RS, Rufty RC. 2005. RAPD and SCAR markers linked to an introgressed Gene Condi-tioning Resistance to Peronospora tabacina D.B Adam in tobacco. Crop Sci.. 45: 2346-2354.
- Ming-hui K, Yan H, Bing-yan H, Yong-ying Z, Shi-jie W, Li-juan M, et al. 2011. Breeding of newly licensed wheat variety Huapei 8 and improved breeding strategy by anther culture. Afr J Biotechnol.. 10(85): 19701-19706.
- Mishra R, Rao GJN. 2016. In-vitro androgenesis in rice: advantages, constraints and future prospects. Rice Sci.. 23(2): 57-68.
- Mollers CC. 1994. Efficient production of doubled haploid Brassica napus plants by colchicine treatment of micro-spores. Euphytica. 75: 95-104.
- Morrison RA, Evans DA. 1987. Gametoclonal variation. Plant Breed Rev.. 5: 359-391.
- Muller HJ. 1927. Artificial transmutation of the genes. Science. 66: 84-87.
- Castillo AM, Chen XW. Muñoz-Amatriaín MCistué LVallés MP2008. Identification and validation of QTLs for green plant percentage in barley (Hordeum vulgare L.) anther culture. Mol Breed.. 22(1): 119-129.
- Svensson JT, Castillo AM, Close TJ. Muñoz-Amatriaín MVallés MP2009. Microspore embryogenesis: assignment of genes to embryo formation and green vs. albino plant production. Funct Integr Genomic.. 9(3): 311-323.
- Murovec J, Bohanec B. 2011. Haploids and doubled haploids in plant breeding. In: Ibrohim Y. Dr. Ibrokhim Abdu-rakhmonov (Ed.) Plant Breeding, 2011.-Р 87-106.
- Myint SK, Aung KO, Soe K. 2005. Development of a short duration upland rice mutant line through anther culture of gamma irradiated plants. Mutat Breed Newslett Rev.. 1: 13-14.
- Nakata K, Tanaka M. 1968. Differentiation of embryoids from developing germ cells in anther culture of tobacco. Jpn J Genet.. 43: 65-71.
- Nehlin L, Bergman P, Glimelius K. Möllers C2000. Transient b-gus and gfp gene expression and viability analysis of microprojectile bombarded microspores of Brassica napus L. J Plant Physiol.. 156(2): 75-183.
- Nezhevenko LI, Shumnyi VK. 1970. Twinning method in raising haploid plants. Genetika..
- Nichols WA, Rufty RC. 1992. Anther culture as a probable source of resistance to tobacco black shank caused by Phytophthora parasitica var 'nicotianae'. Theor Appl Genet. 84(3-4): 473-479..
- Nielsen NH, Andersen SU, Stougaard J, Jensen A, Backes G, Jahoor A. 2015. Chromosomal regions associated with the in vitro culture response of wheat (Triticum aestivum L.) microspores. Plant Breed.. 134: 255-263.
- Niizeki H, Oono K. 1968. Induction of haploid rice plant from anther culture. Proc Jpn Acad.. 44: 554-557.
- Nitsch JP. 1972. Haploid plants from pollen. Z Pflanzenzüchtg. 67: 3-18.
- Nitsch C, Norreel B. 1973. Effect d'un choc thermique sur la pouvoir embryogene du pollen de Datura innoxia cultive dans I'anthere ou isole de l'anthere. CR Acad Sci Paris. 276D: 303-306..
- Nitsch C, Norreel B. 1973. Factors favoring the formation of androgenetic embryos in anther culture. In Genes. Enzymes, and Populations. Springer, Boston, MA 129- 144.:
- Nitsch C. 1974a. La culture de pollen isole sur milieu synthetique. CR Acad Sci Paris. 278D: 1031-1034..
- Nitsch C. 1974. Pollen culture-a new technique for mass production of haploid and homozygous plants. Haploids in higher plants: advances and potential 123-135..
- Nitsch JP, Nitsch C. 1969. Haploid plants from pollen grains. Science. 163: 85-87.
- Nitzche W. 1970. Herstellung haploider Pflanzen aus Festuca- Lolium Bastarden. Sci Nat.. 57: 199-200.
- Ogawa T, Fukuwa H, Ohakawa Y. 1995. Plant regeneration through direct culture of isolated pollen grains in rice. Breed Sci.. 45(3): 301-307.
- Ohkawa Y, Bevis E, Keller WA. 1988. Validity study of microspore culture method in Brassica crops. Cruciferae Newslett.. 13: 75.
- Ohnoutková, L, Vlčko T2020. Homozygous Transgenic Barley (Hordeum vulgare L.) Plants by Anther Culture. Plants. 9(7): 918
- Oiyama I, Kobayashi S. 1993. Haploids obtained from diploid × triploid crosses of citrus. Jpn Soc Hort Sci.. 62(1): 89-93.
- Orlov PA, Becker D, Shewe G, Lorz H. 1999. Cytoplasmic effects on pollen embryogenesis induction in wheat microspore culture. Cereal Res Commun.. 27: 357-363.
- Orlowska R, Pachota KA, Machczinska J, Niedziela A, Makowska K, Zimny J, et al. 2020. Improvement of anther cultures conditions using the Taguchi method in three cereal crops. Electron J Biotechnol.. 43: 8-15.
- Ouyang JW, Hu H, Chuang CC, Tseng CC. 1973. Induction of pollen plants from anthers of Triticum aestivum L. cultured in vitro. Sci Sin.. 16: 79-95.
- Ouyang JW, Jia SE, Zhang C, Chen X, Fen G. 1989. A new synthetic medium (W14) for wheat anther culture. Annual Report. 91-92, Institute of Genetics, Academia Sinica Beijing..
- Ouyang JW, Zhou SM, Jia SE. 1983. The response of anther culture to culture temperature in Triticum aestivum. Theor Appl Genet.. 66: 101-109.
- Padmaja LK, Agarwal P, Gupta V, Mukhopadhyay A, Sodhi YS, Pental D, et al. 2014. Natural mutations in two homoeologous TT8 genes control yellow seed coat trait in allotetraploid Brassica juncea (AABB). Theor Appl Genet.. 127: 339-347.
- Palanisamy D, Marappan S, Ponnuswamy RD, Mahalingam PS, Bohar R, Vaidyanathan S. 2019. Accelerating hybrid rice breeding through the adoption of doubled haploid technology for R-line development. Biologia. 74(10): 1259-1269.
- Palmer CD, Keller WA. 2005. Overview of haploidy. In Haploids in crop improvement II. Springer. Berlin, Heidelberg 3-9..
- Panjabi P, Yadava SK, Sharma P, Kaur A, Kumar A, Arumugam N, et al. 2010. Molecular mapping reveals two independent loci conferring resistance to Albugo candida in the east European germplasms of oilseed mustard Brassica juncea. Theor Appl Genet.. 121: 137-145.
- Panjabi-Massand P, Yadava SK, Sharma P, Kaur A, Kumar A, Arumugam N, et al. 2010. Molecular mapping reveals two independent loci conferring resistance to Albugo candida in the east European germplasm of oilseed mustard Brassica juncea. Theor Appl Genet.. 121(1): 137-145.
- Park SG, Ubaidillah M, Kim KM. 2013. Effect of maltose concentration on plant regeneration of anther culture with different genotypes in rice (Oryza sativa L.). Am J Plant Sci.. 4(11): 2265-2270.
- Pauk J, Janeso M, Simon-Kiss I. 2009. Rice doubled haploids and breeding. In: Touraev A, Forster BP, Jain SM (eds) Advances in haploid production in higher plants. Springer. Netherlands 189-197:
- Pauk J, Beke B, Matuz J. Kertész ZBóna LCsősz M1995. New winter wheat variety- "GK Délibáb" developed via combining conventional breeding and in vitro andro-genesis. Cereal Res Commun.. 23: 251-256.
- Pauk J, Lantos C, Cseuz L, Papp M, Óvári J, Beke B, et al. 2020. 'GK Déva' dihaploid módszer segítségével előállított új őszi búzafajta ('GK Déva', new released winter wheat variety using dihaploid method). XXVI. Növénynemesítési Tudományos Napok. Szeged, Hungary: pp. 04-05.
- Pauk J, Manninen O, Mattila I, Salo Y, Pulli S. 1991. Androgenesis in hexaploid spring wheat F2 populations and their parents using a multiple-step regeneration system. Plant Breed.. 107: 18-27.
- Pauk J, Puolimatka M. Mihály R.2003. Protocol for wheat (Triticum aestivum L. ) anther culture. In Doubled haploid production in crop plants. Springer. Dordrecht 59-64.:
- Paz MMM, Carpena AL, Javier EL. 1994. Chromosome behavior of tobacco (Nicotiana tabacum L.) diploids recon-stituted from anther culture-derived haploids. Philipp J Crop Sci.. 19(1): 45-52.
- Paz MMM, Carpena AL, Javier EL. 1994. Diploidization of Anther Culture-Derived Tobacco (Nicotiana tabacum L.) Haploids. Philipp J Crop Sci.. 19(1): 39-45.
- Pechan PM. 1989. Successful cocultivation of Brassica napus microspores and proembryos with Agrobacterium. Plant Cell Rep.. 8: 387-390.
- Pechan PM, Keller WA. 1988. Identification of potentially embryogenic microspores in Brassica napus. Physiol Plant.. 74: 377-384.
- Picard E, de Huyser J. 1973. Obtention de plantlets haploldes de Triticum aestivum L. a. partir de cultures d'antheres in vitro. CR Acad Sci Paris. 277: 1463-1466.
- Pickering RA. 1980. Attempts to overcome partial incompati-bility between Hordeum vulgare L. and H. bulbosum L. L. Euphytica. 29: 369-377.
- Pink D, Bailey L, McClement S, Hand P, Mathas E, Buchanan- Wollaston V, et al. 2008. Double haploids, markers and QTL analysis in vegetable brassicas. Euphytica. 164: 509-514.
- Potrykus I. 1988. Direct gene transfer to plants. In: Appli-cation of Plant Cell Tissue Culture (Ciba Foundation Symposium 137). John Wiley Sons, New York 144-162:
- Potrykus IJ, Paszkowski M, Saul S, Kruger-Lebus T, Muller R, Schacher I, et al. Freeling M., 1985. editor. Direct gene transfer to protoplasts: an efficient and generally applicable method for stable alterations of plant genomes. In: M. Freeling (Ed.), Plant Genetics, A.R. Liss. New York 181-199.
- Potrykus I, Shillito RD, Saul MW, Paszkowski J. 1985. Direct gene transfer State of the Art and Future Potential. Plant Mol Biol Rep.. 3(3): 117-128.
- Purwoko BS, Dewi IS, Khumaida K. 2010. Rice anther cul-ture to obtain doubled-haploids with multiple tolerances. Asia Pacific J Mol Biol Biotechnol.. 18(1): 55-57.
- Qin G, Gu H, Ma L, Peng Y, Deng XW, Chen Z, et al. 2007. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis. Cell Res.. 17: 471-482.
- Rahman MH, Krishnaraj S, Thorpe TA. 1995. Selection for salt tolerance in vitro using microspore-derived embryos of Brassica napus cv Topas, and the characterization of putative tolerant plants. In Vitro Cell Dev Biol Plant.. 31: 116-121.
- Raina SK, Zapata FJ. 1997. Enhanced anther culture effi-ciency of indica rice (Oryza sativa L.) through modifi-cation of the culture media. Plant Breed.. 116(4): 305-315.
- Rajyalakshmi K, Chowdhry CN, Maheshwari N, Maheshwari SC. 1995. Anther culture response in some Indian wheat cultivars and the role of polyamines in induction of haploids. Phytomorphology: Int J Plant Morphol. 45: 139-145.
- Ramchiary N, Bisht NC, Gupta V, Mukhopadhyay A, Arumugam N, Sodhi YS, et al. 2007b. QTL analysis reveals context-dependent loci for seed glucosinolate trait in the oilseed Brassica juncea: importance of recurrent selection backcross scheme for the identification of 'true' QTL. Theor Appl Genet.. 116: 77-85.
- Ramchiary N, Padmaja KL, Sharma S, Gupta V, Sodhi YS, Mukhopadhyay A, et al. 2007a. Mapping of yield influencing QTL in Brassica juncea: implications for breeding of a major oilseed crop of dryland areas. Theor Appl Genet.. 115: 807-817.
- Redha A, Suleman P. 2011. Effects of exogenous application of polyamines on wheat anther culture. Plant Cell Tissue Organ Cult.. 105: 345-353.
- Reinert J, Bajaj YPS. 1977. Anther culture: Haploid produc-tion and its significance. In: Reinert J, Bajaj YPS. Applied and Fundamental Aspects of Plant Cell, Tissue and Organ Culture. Berlin Heidelberg: Springer 251-267.
- Research Group 303Institute of GeneticsAcademia Sinica1972. A preliminary report on 96 H. Hu the induction of pollen plants from rice and wheat. Genet Commun Beijing.. 1: 1-4.
- Ribarits A, Mamun AN, Li S, Resch T, Fiers M, Heberle-Bors E, et al. 2007. Combination of reversible male sterility and doubled haploid production by targeted inactivation of cytoplasmic glutamine synthetase in developing anthers and pollen. Plant Biotechnol J.. 5: 483-494.
- Rout K, Yadav BG, Yadava SK, Mukhopadhyay A, Gupta V, Pental D, et al. 2018. QTL landscape for oil content in Brassica juncea: analysis in multiple bi-parental popu-lations in high and "0" erucic background. Front Plant Sci.. 9: 1448
- Rubtsova M, Gnad H, Melzer M, Weyen J, Gils M. 2013. The auxins centrophenoxine and 2, 4-D differ in their effects on non-directly induced chromosome doubling in anther culture of wheat (T. aestivum L. ). Plant Biotechnol Rep.. 7(3): 247-255.
- Rudolf K, Bohanec B, Hansen N. 1999. Microspore culture of white cabbage, Brassica oleracea var. capitata L. genetic improvement of non-responsive cultivars and effect of genome doubling agents. Plant Breed.. 118: 237-241.
- Rustgi S, Kashyap S, Ankrah N, von Wettstein D. 2020. Use of microspore-derived calli as explants for biolistic transformation of common wheat. In Biolistic DNA delivery in plants. Humana. New York, NY 263-279.:
- Sadasivaiah RS, Perkovic SM, Pearson C, Postman B, Beres BL. 2004. Registration of "AC Andrew" wheat. Crop Sci.. 44: 696-697.
- Samal P, Pote TD, Krishnan SG, Singh AK, Salgotra RK, Rathour R. 2019. Integrating marker-assisted selection and doubled haploidy for rapid introgression of semi- dwarfing and blast resistance genes into a basmati rice variety 'ranbir basmati'. Euphytica. 215(9): 1-13.
- Sanchez-Diaz RA, Castillo AM, Valles MP. 2013. Micro-spore embryogenesis in wheat: new marker genes for early, middle and late stages of embryo development. Plant Rep.. 26: 287-296.
- Sarao NK, Gosal SS. 2018. In vitro androgenesis for accele-rated breeding in rice. In Biotechnologies of Crop Improvement. Volume 1. Springer, Cham 407-435.:
- Schedel S, Pencs S, Hensel G, Rutten T, Kumlehn J. Müller A2017. RNA-guided Cas9-induced mutagenesis in tobacco followed by efficient genetic fixation in doubled haploid plants. Front Plant Sci.. 7: 1995
- Seguı'-Simarro JM, Nuez F. 2008a. How microspores trans-form into haploid embryos: changes associated with em-bryogenesis induction and microsporederived embryoge-nesis. Physiol Plant.. 134: 1-12.
- Seguı'-Simarro JM. 2010. Androgenesis revisited. Bot Rev.. 76: 377-404.
- Seifert F, Kumlehn J, Gnad H, Scholten S. Bössow S2016. Analyses of wheat microspore embryogenesis induction by transcriptome and small RNA sequencing using the highly responsive cultivar "Svilena". BMC Plant Biol.. 16: 97
- Sen C, Singh RP, Singh MK, Singh HB. 2011. Effect of cold pretreatment on anther culture of boro rice hybrids. Int. J. Plant Rep Biol.. 3: 69-73.
- Senadhira D, Zapata-Arias FJ, Gregorio GB, Alejar MS, De La Cruz HC, Padolina TF, et al. 2002. Development of the first salt tolerant rice cultivar through indica/indica anther culture. Field Crops Res. 76(2/3): 103-110..
- Shariatpanahi ME, Bal U, Heberle-Bors E, Touraev A. 2006. Stresses applied for the re-programming of plant mmicro-spores towards in vitro embryogenesis. Physiol Plant.. 127: 519-534.
- Tajedini S, Oroojloo M, Ghanim AMA, Fakheri B, Mahdinejad N. Shariatpanahi.ME.2018. Haploidy in Rice (Oryza Sativa L. ) Mutation Breeding for Striga Resistance (No. IAEA-CN--263)..
- Shi-kuan H, Ya-ying L. 1985. Breeding the Restoring Lines of the Hybrid T-type Wheat with Anther Culture. Chinese Bulletin of Botany. 3(06): 49.
- Shimada T, Toriyama K, Tsunewaki K, Nonaka S, Koba T, Otani M, et al. 1994. Breeding of candidate lines for male sterility-maintainer by anther culture for hybrid wheat production using an Sv type cytoplasm and a 1BL-1RS chromosome. Jpn J Breed.. 44(1): 23-28.
- Shouyi L, Shouyin H. 1991. Huayu 15 A high yielding rice variety bred by anther culture. In Bajaj YPS, Biotechnol Agric For.. 14: 230-247.
- Siddique R. 2015. Impact of different media and genotypes in improving anther culture response in rice (Oryza sativa) in Bangladesh. Eur Sci J.. 11(6): 164-169.
- Siebel J, Pauls KP. 1989. A comparison of anther and microspore culture as a breeding tool in Brassica napus. Theor Appl Genet.. 78: 473-479.
- Singh S, Bhatia R, Kumar R, Das A, Ghemeray H, Behera TK, et al. 2021. Characterization and genetic analysis of OguCMS and doubled haploid based large genetic arsenal of Indian cauliflowers (Brassica oleracea var. botrytis L. ) for morphological, reproductive and seed yield traits revealed their breeding potential. Genet Resour Crop Evol.. 68(4): 1603-1623.
- Smykal P. 2000. Pollen embryogenesis—the stress mediated switch from gametophytic to sporophytic development. Current status and future prospects. Biol Plant.. 43: 481-489.
- Sopory SK, Munshi M. 1996. In vitro haploid production in higher plants, 145-176.
- Soriano M, Castillo AM. Cistué L2008. Enhanced induction of microspore embryogenesis after n-butanol treatment in wheat (Triticum aestivum L.) anther culture. Plant Cell Rep.. 27: 805-811.
- Soriano M, Valles MP, Castillo AM. Cistué L2007. Effects of colchicine on anther and microspore culture of bread wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult.. 91(3): 225-234.
- Soundararajan RP, Kadirvel P, Gunathilagaraj K, Maheswaran M. 2004. Mapping of quantitative trait loci associated with resistance to brown planthopper in rice by means of a doubled haploid population. Crop Sci.. 44: 2214-2220.
- Sourdille P, Snape JW, Cadalen T, Charmet G, Nakata N, Bernard S, et al. 2000. Detection of QTLs for heading time and photoperiod response in wheat using a doubled- haploid population. Genome. 43(3): 487-494.
- Sriskandarajah S, Sameri M, Westerbergh A. Lerceteau‐Köhler E2015. Increased recovery of green doubled haploid plants from barley anther culture. Crop Sci.. 55(6): 2806-2812.
- Suenaga K, Khairallah M, William HW, Hoisington DA. 2005. A new intervarietal linkage map and its application for quantitative trait locus analysis of "gigas" features in bread wheat. Genome 48. 65-75.:
- Sunderland N. 1978. Strategies in the improvement of yields in anther culture. In Proceedings of symposium on plant tissue culture. Science Press 65-86..
- Sunderland N. 2012. Anther culture of Nicotiana tabacum. Cell Culture and Somatic Cell Genetics of Plants Vol.1. 283.:
- Sunderland N, Wicks FM. 1969. Cultivation of haploid plants from tobacco pollen. Nature. 2.:
- Sunderland N, Wicks FM. 1971. Embryoid formation in pollen grains of Nicotiana tabacum. J Exp Bot.. 22(1): 213-226.
- Suzuki T, Sato M, Takeuchi T. 2012. Evaluation of the effects of five QTL regions on Fusarium head blight resistance and agronomic traits in spring wheat (Triticum aestivum L.). Breed Sci.. 62(1): 11-17.
- Pauk J. Szakács ÉKovács GBarnabás B1988. Substitution analyses of callus induction and plant regeneration from anther culture in wheat (Triticum aestivum L.). Plant Cell Rep.. 7: 127-129.
- Szarejko I, Forster BP. 2006. Doubled haploidy and induced mutation. Euphytica. 158: 359-370.
- Tadele Z, Chikelu MBA, Till BJ. 2010. TILLING for mutations in model plants and crops. In Molecular Techniques in crop improvement. Springer. Dordrecht 307-332.:
- Tai GCC. 2005. Haploids in the improvement of Solanaceous species. In: Palmer CE, Keller WA, Kasha KJ (eds) Haploids in Crop Improvement II. Springer. Berlin/Heidelberg, pp 173-190:
- Takahata Y, Keller WA. 1991. High frequency embryogenesis and plant regeneration in isolated microspore culture of Brassica oleracea L. Plant Sci.. 74: 235-242.
- Thomas E, Wenzel G. 1975. Embryogenesis from microspores of Brassica napus. Z Pflanzenzuecht.. 74: 77-81.
- Thompson KF. 1972. Oil-seed rape. In: Reports of the Plant Breeding Institute. Cambridge University Press. Cambridge 94-96.
- Thomson MJ, de Ocampo M, Egdane J, Rahman MA, Sajise AG, Adorada DL, et al. 2010. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice. 3(2): 148-160.
- Tinker NA, Mather DE, Rossnagel BG, Kasha KJ, Kleinhofs A, Hayes PM, et al. 1996. Regions of the genome that affect agronomic performance in two‐row barley. Crop Sci.. 36(4): 1053-1062.
- Tixier MH, Sourdille P, Charmet G, Gay G, Jaby C, Cadalen T, et al. 1998. Detection of QTLs for crossability in wheat using a doubled-haploid population. Theor Appl Genet. 97. 1076-1082.:
- Tong D, Jia X. 1991. Mutant in tobacco anther culture induced by 60 Co g-rays. Acta Agriculturae Nucleatae Sinica. 5(4): 193-198.
- Toojinda T, Baird E, Booth A, Broers L, Hayes P, Powell W, et al. 1998. Introgression of quantitative trait loci (QTLs) determining stripe rust resistance in barley: an example of marker-assisted development. Theor Appl Genet.. 96: 123-131.
- Torp AM, Hansen AL, Andersen SB. 2001. Chromosomal regions associated with green plant regeneration in wheat (Triticum aestivum L.) anther culture. Euphytica. 119: 377-387.
- Torp AM, Andersen SB. 2009. Albinism in microspore culture. In Advances in haploid production in higher plants. Springer. Dordrecht 155-160.:
- Touraev A, Forster BP. Jain SM (eds).2009. Advances in haploid production in higher plants. Springer. Berlin.:
- Touraev A, Heberle-Bors E. 2003. Anther and microspore culture in tobacco. In Doubled haploid production in crop plants (223-228). Springer, Dordrecht. 24(5225): 1227-1229.
- Touraev A, Indrianto A, Wratschko I, Vicente O, Heberle- Bors E. 1996b. Efficient microspore embryogenesis in wheat (Triticum aestivum L.) induced by starvation at high temperatures. Sex Plant Reprod.. 9: 209-215.
- Touraev A, Pfosser M, Heberle-Bors E. 2001. The microspore: a haploid multipurpose cell. Adv Bot Res.. 35: 53-109.
- Touraev A, Vicente O, Heberle-Bors E. 1997. Initiation of microspore embryogenesis by stress. Trends Plant Sci.. 2: 297-302.
- Touraev A, Pfosser M, Heberle-Bors E. 2001. The microspore: a haploid multipurpose cell.
- Trejo-Tapia G, Amaya UM, Morales GS, Sanchez ADJ, Bonfil BM, Rodriguez-Monroy M, et al. 2002. The effects of cold-pretreatment, auxins and carbon source on anther culture of rice. Plant Cell Tiss Organ Cult.. 71: 41-46.
- Turcotte EL, Feaster CV. 1963. Haploids: High-frequency production from single-embryo seeds in a line of Pima cotton. Science. 140(3574): 1407-1408.
- Turcotte EL, Feaster CV. 1969. Semigametic Production of Haploids in Pima Cotton 1. Crop Sci.. 9(5): 653-655.
- Turner J, Facciotti D. 1990. High oleic acid Brassica napus from mutagenized microspores. In: McFerson JR, Kresovich S, Dwyer SG (eds) Proc 6th Crucifer Genet Workshop. Geneva 24.
- Tuvesson IKD, Peterson S, Anderson SB. 1989. Nuclear genes affecting albinism in wheat (Triticum aestivum L.) anther culture. Theor Appl Genet.. 78: 879-961.
- Tuvesson S, Ljungberg A, Johasson N, Karlsson KE, Suijs LW, Posset JP. 2000. Large-scale production of wheat and triticale double haploid through the use of a single- anther culture method. Plant Breed.. 119: 455-459.
- Tuvesson S, Von Post R, Ljungberg A. 2003. Wheat anther culture. In: Maluszynski M. Kasha KJ, Forster BP, Szarejko I (eds), Doubled Haploid Production in Crop Plants, a manual, Kluwer Academic Publishers, Dordrecht, The Netherlands 71-76:
- Tuvesson S, Dayteg C, Hagberg P, Manninen O, Tanhuanpää P, Tenhola-Roininen T, et al. 2007. Molecular markers and doubled haploids in European plant breeding programmes. Euphytica. 158(3): 305-312.
- Ushiyama T. Studies on haploid breeding in wheat. United Graduate School of Agricultural Science 2008. Tokyo University of Agriculture and Technology 1-60.
- Uzunova M, Ecke W, Weissleder K. Röbbelen G1995. Mapping the genome of rapeseed (Brassica napus L.): construction of an RFLP linkage map and localization of QTLs for seed glucosinolates content. Theor Appl Genet.. 90: 194-204.
- Wang JY, Zhang ZX, Xiang YW, Zhang AZ, Zhou XM. 1994. Effects of integrated breeding techniques on the selection of restorer lines in hybrid rice. Southwest China J Agric Sci.. 7(2): 17-21.
- Wang X, Wang H, Wang J, Sun R, Wu J, Liu S. 2011. Brassica rapa genome sequencing project consortium. The genome of the mesopolyploid crop species Brassica rapa. Nat Genet.. 43: 1035-1039.
- Wang Z, Taramino G, Yong D, Liu G, Tingey SV, Miao GL, et al. 2001. Rice ESTs with disease-resistance gene- or defense-response gene-like sequences mapped to regions containing major resistance genes or QTLs. Mol Genet Genom.. 265(2): 302-310.
- Watts A, Sankaranarayanan S, Raipuria RK, Watts A. 2020. Production and Application of Doubled Haploid in Brassica Improvement. In Brassica Improvement. Springer. Cham: pp. 67-84.
- Weigt D, Kiel A, Nawracala J, Pluta M, Lacka A. 2016. Solid-stemmed spring wheat cultivars give better andro-genic response than hollow-stemmed cultivars in anther culture. In Vitro Cell Dev Biol Plant.. 52: 619-625.
- Weigt D, Kiel A, Siatkowski I, Zyprych-Walczak J, Tomkowiak A, Kwiatek M. 2020. Comparison of androgenic response of spring and winter wheat. Plants. 9: 49
- Weigt D, Niemann J, Siatkowski I, Zyprych-Walczak J, Przemyslaw O, Kurasiak-Popowska D. 2019. Effect of zearalenone and hormone regulators on microspore embryogenesis in anther culture of wheat. Plants. 8: 487
- Wernsman EA. 1993. Varied roles for the haploid sporophyte in plant improvement. In: Stalker HT, Murphy JP (eds) Plant Breeding in the 1990s. CAB International. Wallingford, pp 461-484.
- Wessels E, Botes WC. 2014. Accelerating resistance breeding in wheat by integrating marker-assisted selection and doubled haploid technology. South African J Plant Soil.. 31(1): 35-43.
- Williams K, Taylor S, Bogacki P, Pallotta M, Bariana H, Wallwork H. 2002. Mapping of the root lesion nematode (Pratylenchus neglectus) resistance gene Rlnn1 in wheat. Theor Appl Genet.. 104(5): 874-879.
- Wong RSC, Swanson E. 1991. Genetic modification of canola oil: high oleic acid canola. In: Haberstroh C, Morris CE (eds) Fat and cholesterol reduced food. Gulf. Houston 154-164.
- Xa TT, Lang NT. 2011. Rice breeding for high grain quality through anther culture. Omonrice. 18: 68-72.
- Xu L, Najeeb U, Tang GX, Gu HH, Zhang GQ, He Y, et al. 2007. Haploid and doubled haploid technology. Adv Bot Res.. 45: 181-216.
- Xu Q, Yuan XP, Yu HY, Wang YP, Tang SX, Wei XH. 2011. Mapping quantitative trait loci for sheath blight resistance in rice using double haploid population. Plant Breed.. 130(3): 404-406.
- Yadava SK, Arumugam N, Mukhopadhyay A, Sodhi YS, Gupta V, Pental D, et al. 2012. QTL mapping of yield- associated traits in Brassica juncea: meta-analysis and epistatic interactions using two different crosses between east European and Indian gene pool lines. Theor Appl Genet.. 125(7): 1553-1564.
- Yamagishi M, Otani M, Higashi M, Fukuta Y, Fukui K, Shimada T. 1998. Chromosomal regions controlling anther culturability in rice (Oryza sativa L.). Euphytica. 103(2): 227-234.
- Yang J, Liu D, Wang X, Ji C, Cheng F, Liu B, et al. 2016. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat Genet.. 48(10): 1225-1232.
- Yang HY, Zhou C. 1979. Experimental research on the two pathways of pollen development in Oryza sativa L. Act Bot Sin.. 21: 345-351.
- Yang XR, Fu HH. 1989. Hua-03: a high protein indica rice. Intl Rice Res Newslett.. 14: 14-15.
- Yang ZP, Gilbert J, Fedak G, Somers DJ. 2005. Genetic characterization of QTL associated with resistance to Fusarium head blight in a doubled-haploid spring wheat population. Genome 48. 187-196.:
- Yoshida KT, Fujii S, Sakata M, Takeda G. 1994. Control of organogenesis and embryogenesis in rice calli. Breed. Sci.. 44(4): 355-360.
- Zapata FJ, Aldemita RR. 1989. Induction of salt tolerance in highyielding rice varieties through mutagenesis and anther culture. In: Maluszynski M. Current Options for Cereal Improvement: Doubled Haploid. Mutants and Heterosis 193-202:
- Zhang YX, Lespinasse Y, Chevreau E. 1990. Induction of haploidy in fruit trees. Acta Hortic.. 280: 293-304.
- Zhang JF, Stewart JM. 2004. Semigamy gene is associated with chlorophyll reduction in cotton. Crop Sci.. 44(6): 2054-2062.
- Zhao L, Liu L, Wang J, Guo H, Gu J, zhao S, et al. 2015. Development of a new wheat germplasm with high anther culture ability by using of gamma-ray irradiation and anther culture. J Sci Food Agr.. 95: 120-125.
- Zhao P, Wang K, Zhang W, Liu HY, du LP, Hu HR, et al. 2017. Comprehensive analyses of differently expressed genes and proteins in albino and green plantlets from a wheat anther culture. Biol Plant.. 61: 255-265.
- Zhenji ASC. 1981. Genetic Studies on Tobacco Breeding with Haploid Developed by Anther Culture. Act Genet Sin. 04..
- Zhongxin W, Tongqing Z, Genhuai Y, Fangyang G. 1994. Selection of Tobacco Lines Resistant to Alternaria alterata by Anther Culture. J Agric Biotech..
- Zhou WWJ, Tang GGX, Hagberg PP. 2002. Efficient produc-tion of doubled haploid plants by immediate colchicine treatment of isolated microspores in winter Brassica napus. Plant Growth Regul.. 37: 185-192.
- Zhu DY, Pan XG. 1990. Rice (Oryza sativa L.): Guan 18: An improved variety through anther culture. In: Bajaj Y P S. Biotechnology in Agriculture and Forestry 12: Rice. Berlin Heidelberg, Springer. 12: Rice. Berlin.
- Zhu DY, Sun ZX, Pan XG, Ding XH, Shen XH, Wan Y, et al. 1998. Use of anther culture in hybrid rice breeding. Adv Hybrid Rice Technol.. 21: 265-281.