Abstract
Pathogens are the major causes of wheat crop yield losses, including the fungus Fusarium graminearum, an agent of Fusarium Head Blight (FHB). A better understanding of the relationship between plant morphological and biochemical traits and resistance to FHB can be effective in implementing a successful breeding program. This study investigated the relationship between FHB resistance as well as the morphological and biochemical traits in 20 durum wheat lines. Both morphological and biochemical traits were investigated using statistical tools. Therefore, analyses of variance, mean, as well as the correlation between the traits were con-sidered. In addition, for the morphological traits, cluster analyses were performed to identify similar genotypes in control and infected conditions. Furthermore, machine learning (ML) classification techniques, including Support Vector Machine (SVM), were proposed to detect the infected plants using morphological traits. The results show a great promise for the application of data-driven ML-based methods in plant breeding and disease detection.
-
Key words: Fusarium head blight, Durum wheat, Plant pathology, Machine learning, Deep learning, Support vector machine
INTRODUCTION
With the growing global population, which would reach 9 billion by 2050, food security will be the major challenge for governments (
Azimi 2018). Wheat is the second most important crop globally with a total estimated world pro-duction of approximately 663 million tons during 2011-2012 (
http://faostat.fao.org). Currently, about 95% of the wheat grown worldwide is hexaploidy bread wheat, with most of the remaining 5% being tetraploid durum wheat. The durum wheat is the most cultivated species of wheat with significant economic and nutritional impacts due to its unique characteristics. Its protein and gluten contents are crucial for diverse food products, such as pasta that is the most common durum wheat final product consumed in Europe and North America (
Pinheiro et al. 2013).
Fusarium Head Blight (FHB) caused by
Fusarium graminearum is a fungal disease that mostly affects wheat, corn, and barley (
Figs. 1,
2). It was first discovered in England in 1884, and because of the chalky, lifeless appearance of the infected kernels, it was called wheat scab. Due to the production of mycotoxins, FHB causes significant yield loss (i.e., 10%-70% during epidemic years (
Matthies and Buchenauer 2000)). The amount of such contaminants in wheat (and other cereals) is of great concern because their presence in foods is often associated with chronic or acute mycotoxicoses in livestock, which could threaten human health (
Visconti and Pascale 2010).
Additionally, the disease is favored by extended periods of moderate to high temperature (15-30℃), high moisture (humidity > 90%), and frequent rainfall (
Wiśniewska et al. 2014;
Lenc 2015). For example, two-third of provinces in China, with approximately 7 million hectares, are suscepti-ble to FHB infection (
Wang et al. 1982;
Ze et al. 2017). Seventeen species of
Fusarium, with different climatic re-quirements and genetic and environmental adaptations, cause FHB (Stępień and Chełkowski 2010). The source of primary inoculations are the fungal colonies as well as overwintering of crops such as wheat, maize, and rice residues (
Sutton 1982;
Adams 1921;
Parry et al. 1995), which make the spores to be deposited on or inside wheat florets where they germinate and initiate infection (
Bai and Shaner 1994;
Bushnell et al. 2003). Soon after infection, dark-brown spots appear on the infected florets, and later, the entire florets become blighted. Typically, the first noticeable symptom is bleaching of some or all parts of the spikelets. Studies have shown that the use of fungicides and agronomic operations can partially reduce the loss, how-ever, the use of resistant cultivars is the most efficient and cost-effective strategy to overcome FHB (
Parry et al. 1995;
McMullen et al. 1997). The future breeding efforts would consider developing new FHB-resistant cultivars as the most important goal.
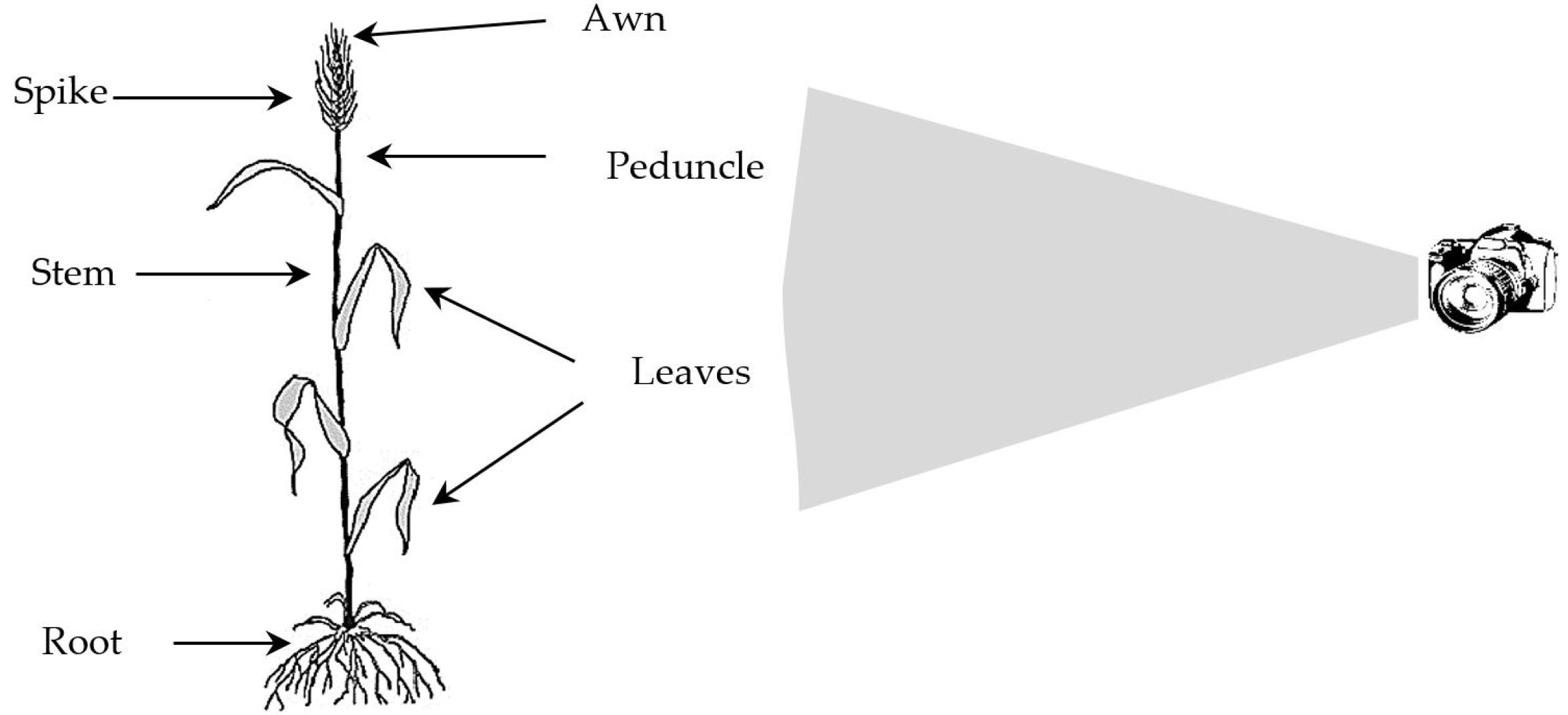
In this study, in order to investigate the resistant and sensitive genotypes of durum wheat lines to FHB and to study the genetic diversity of FHB disease, an experimental greenhouse study was conducted based on a completely randomized design. The disease indices, as well as the mor-phological-biochemical traits, were measured. The data were used during the clustering and classification proce-dure through statistical and machine learning (ML) met-hods. The aim of the ML-based classification methods in this study is to promote automated crop monitoring using high-tech machines. However, the morphological traits were measured by the human in this study, using a digital camera, the proposed technique can be improved and automated using the framework shown in
Fig. 3.
MATERIALS AND METHODS
Greenhouse operation
This study was carried out in the greenhouse with a tem-perature of 20 ± 3℃ during the day (16 hours), and 16 ± 3℃ during the night (8 hours). For this experimental study, 20 Durum wheat lines from the plant gene bank were selected (
Table 1). The seeds were planted at the same temperature, light, moisture, soil, and fertilizer conditions.
In this study, The Wegener’s method (
Wegener 1992) was used to prepare the fungal inoculum for spraying (
Fig. 4a). Using this method, 5 grams of straw powder was mixed with 125 mL of distilled water in an Erlenmeyer flask (250 mL), and the mixture was placed in an autoclave at 120℃ and pressure of one atmosphere for 30 minutes and three times in 48 hours. Then, a small piece of Fusarium mycelium disc with 3-5 millimeters diameter along with the culture medium was poured into the Erlenmeyer flask and placed in an incubator shaker at 25-30℃ (
Fig. 4b). The spores of the Fusarium fungus were produced and stored at 4℃ after 96 hours. The concentration of suspension was adjusted to 5.104 spores per milliliter. When the plants reached 50% of flowering, the prepared inoculum was sprayed on spikes using a manual agitator (
Fig. 4c). After inocula-tion, to ensure the contamination, spikes were hand-misted with a hand-fogging nozzle for 30 minutes. After inocula-tion, spikes were covered with plastic bags to simulate humidity for 24 hours (
Fig. 4d). This procedure was repeated twice after three days interval. After two weeks, the morphological traits were recorded by observing the symp-toms of the disease. For the biochemical traits, the sam-pling was done according to Bernardo
et al. (
Bernardo et al. 2012).
Symptoms of the disease begin with brown spots along with the spikes and ultimately lead to bleaching of the entire spikes (
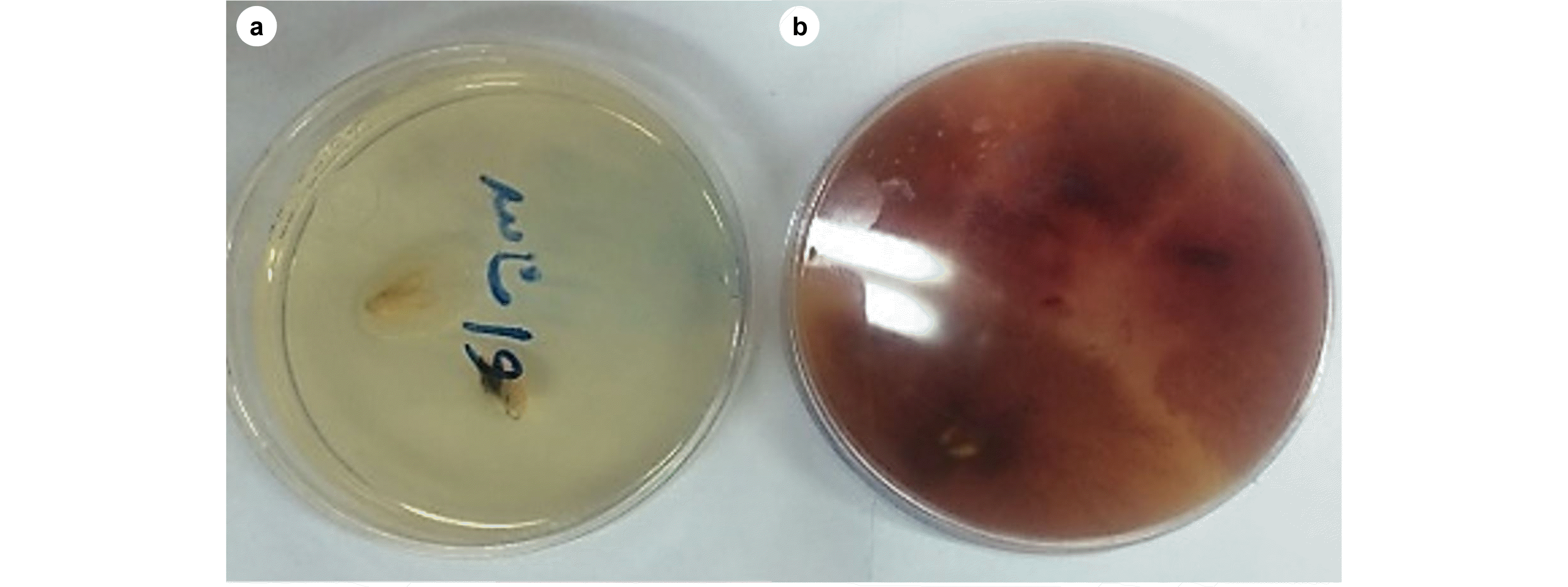
Siranidou et al. 2002). Different parts of spikes of the control and infected plants were removed using a blade and disinfected using sodium hypochlorite 0.5% for 30-60 seconds, and washed with distilled water. Then, they were transferred to the PDA after drying. The plates were placed in the incubator at 25℃ (12 hours in dark and 12 hours in light conditions). The growth of the Fusarium fungus was observable around the cultivars after 48 hours (
Fig. 5).
The tissue samples were used to measure soluble carbo-hydrates (
Irigoyen et al. 1992). The proline content in leaf samples was estimated according to Bates
et al. (
Bates et al. 1973). The tissue sample was ground and homogenized in 5 mL of 3% sulfosalicylic acid. 2 mL of the extract was mixed with 2 mL of glacial acetic acid and 2 mL of nin-hydrin reagent. The mixture was boiled in a water bath at 100℃ for 30 minutes and cooled down slowly. When the mixture was cool, the color developed was extracted with 6 mL of toluene and read at 520 nm. Using the standard graph, the amount of proline was estimated.
To prepare the extract of antioxidant enzymes from the plant tissue, the tissues were ground after freezing in liquid nitrogen. Then, 5-10 mL of buffer was added to homo-genize. Finally, the extract was placed in a centrifuge at 15,000 rpm for 10 minutes at 4℃. Based on Kar and Mishra (
Kar and Mishra 1976), the catalase and peroxidase enzyme activities were measured.
Bradford’s method (
Bradford 1976) was used to mea-sure the protein concentration of the green leaves. There-fore, a 0.1 g of leaf sample with 1.0 mL phosphate buffer were mixed, ground, and placed in a microtube with 12,000 rpm at 4℃ for 15 minutes. 100 µL of the obtained solution was added to the test tube along with 5 mL of the Bradford indicator. After 5 minutes, using the spectrophotometer, the index was read at 595 nm wavelength in the room temperature.
The support vector machine (SVM) is a supervised classifier that optimally separates each labeled class using a hyperplane (
Hsu and Lin 2002;
Pan et al. 2017;
Alisaac et al. 2018;
Golhani et al. 2018;
Pan et al. 2018;
Qiu et al. 2018). Some advantages of SVM over other statistical methods include (1) regularization to avoid overfitting, (2) using several kernels, and (3) convex optimization to avoid local minima that are common in AI-based methods (
Azimi and Pekcan 2019;
Azimi et al. 2020). More detailed infor-mation regarding ML-based classifiers can be obtained from Nisbet
et al. (
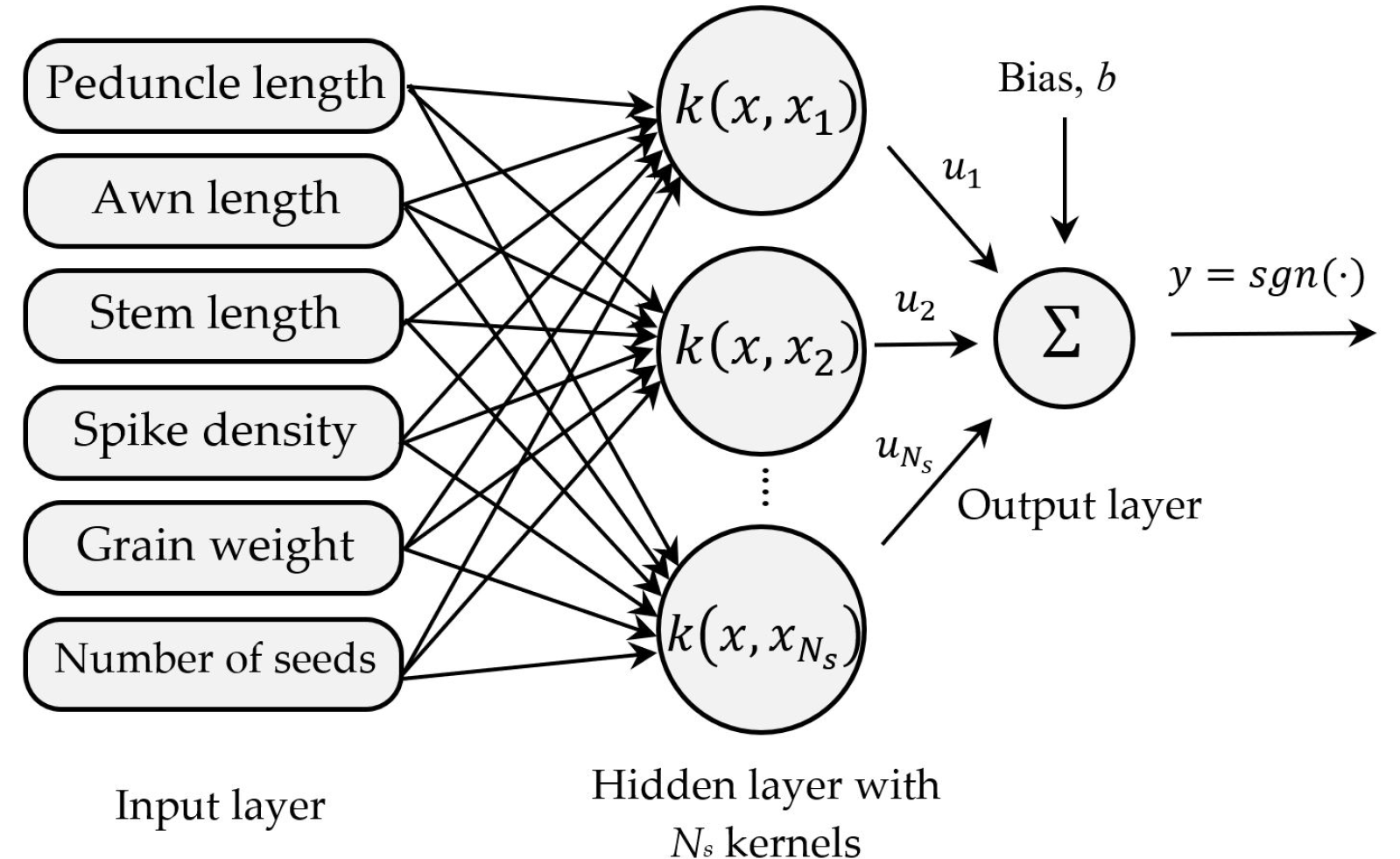
Nisbet et al. 2018). The SVM can be considered as an optimization problem, which finds the optimal hyperplane with the maximum margin. This pro-cess can be summarized as finding a decision function,
f (
x,
y), that maps the inputs
x to labels
y (
Fig. 6). Therefore, for a new label
xn, the discriminant function
f (
xn) can classify it as
yn.
for which, the constraints are
where (xi, yi) are the input pairs of the total m training samples, and a is a vector of the importance factors of the training samples. C and k are the regularization parameter and the kernel function, respectively. The discriminant function, f (xn), to classify new data, xn, is defined as:
In this study, the SVM classifier is compared with three other popular classifiers; k-nearest neighbors (KNN), decision tree (DT), and Naive Bayes (NB). Algorithms of these classification methods are embedded in MATLAB that was used in this study. For the SVM model, a linear kernel function was used; and for the KNN model, the number of neighbors is selected as 5.
To avoid overfitting and to generalize the models, the k-fold cross-validation technique was used, which has ad-vantages over the other regularization methods. Using the k-fold cross-validation, (1) the entire dataset is split into k folds; (2) for each kth fold, the model is trained based on the k-1 folds (train set), and the trained model is tested for the kth fold (test set); and (3) the average k recorded accuracy scores is considered as the performance metric. For each run, 80% of the whole data samples were used to train each classifier, and the remaining 20% were used to test the accuracy.
RESULTS
Analysis of variance and mean values of the morphological traits
The results of the analysis of variance for all traits are given in
Table 2. The maximum and minimum variation are for the stem length and the number of seeds per spike, respectively. The results of the mean values are given in
Table 3, which are based on Duncan’s method for the significance level of
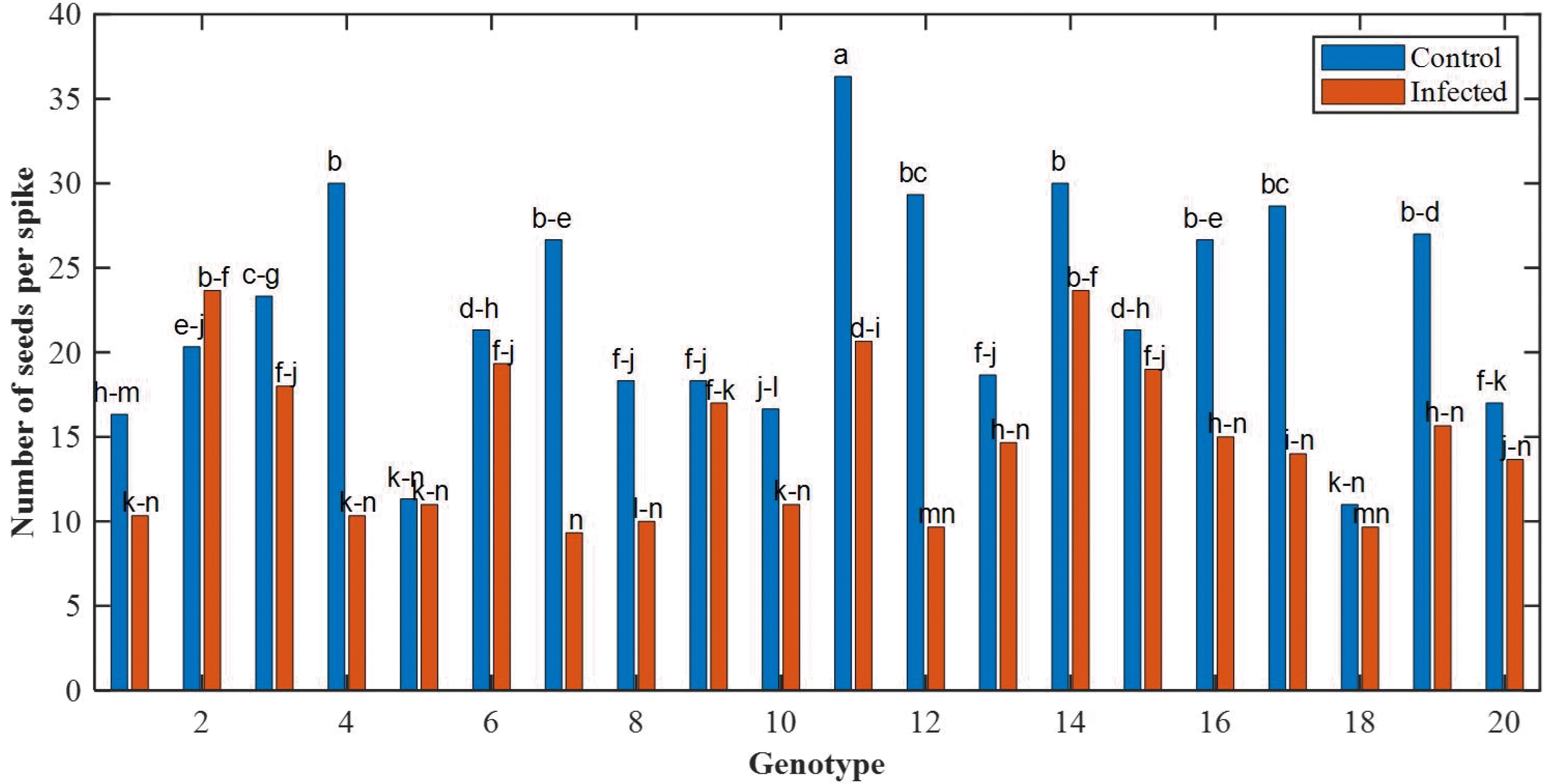
P < 0.05. The results show that under the stress, the peduncle length of the cultivars is reduced. The mean values of the number of seeds per spike for different genotypes are shown in
Fig. 7.
Results of the analysis of variance and mean values of biochemical traits (protein, carbohydrate, proline, peroxidase, and catalase) are given in
Tables 4,
5, respectively. Accord-ing to the results, the effect of stress and interaction effects of stress × genotype is not significant. The highest and lowest coefficient of variation is related to proline and catalase, respectively. The highest protein content is ob-tained for genotype 2 and the lowest amount is observed for genotype 12. The highest amount of proline is observed for genotype 2 and the lowest amount for genotype 7.
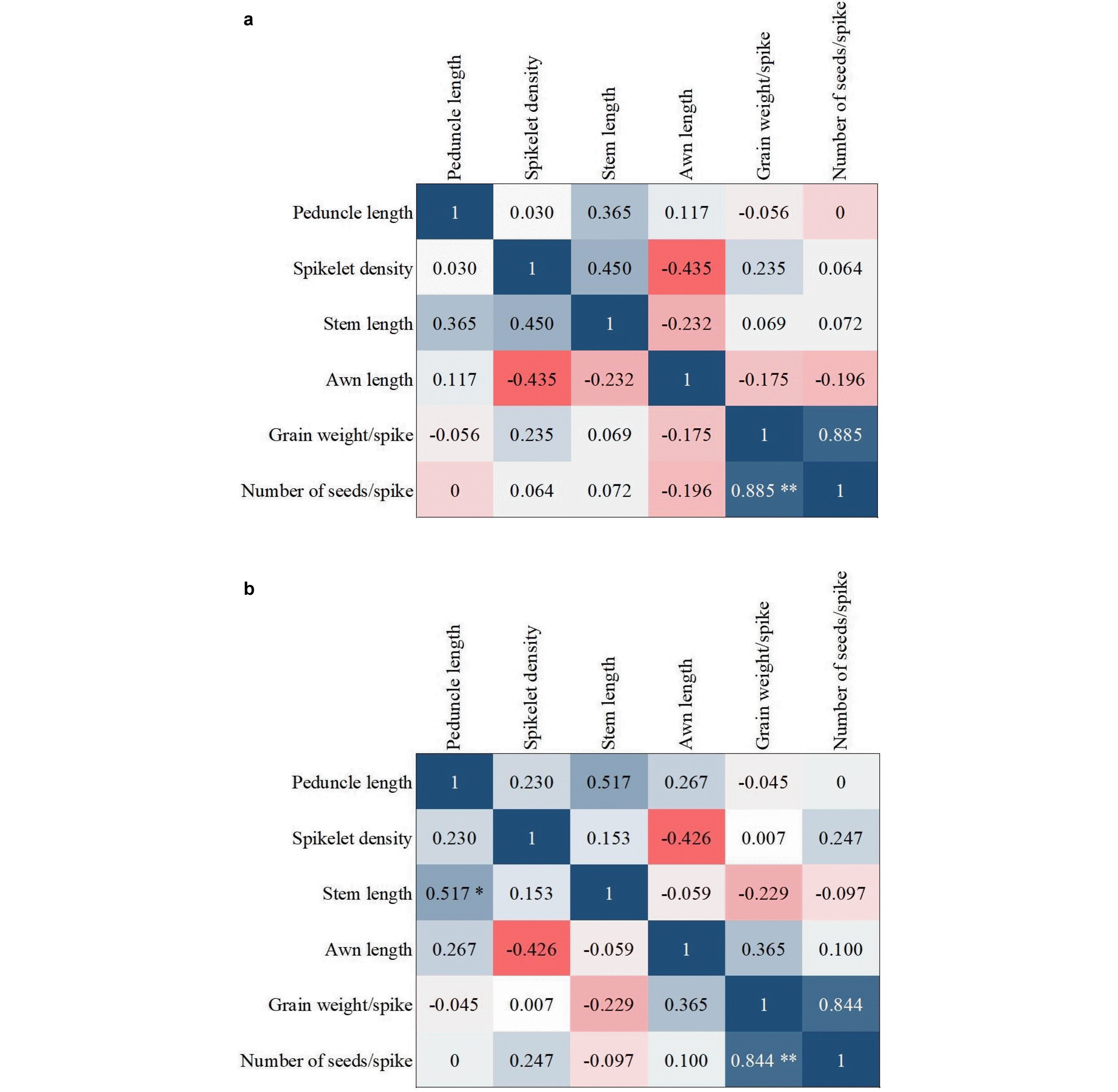
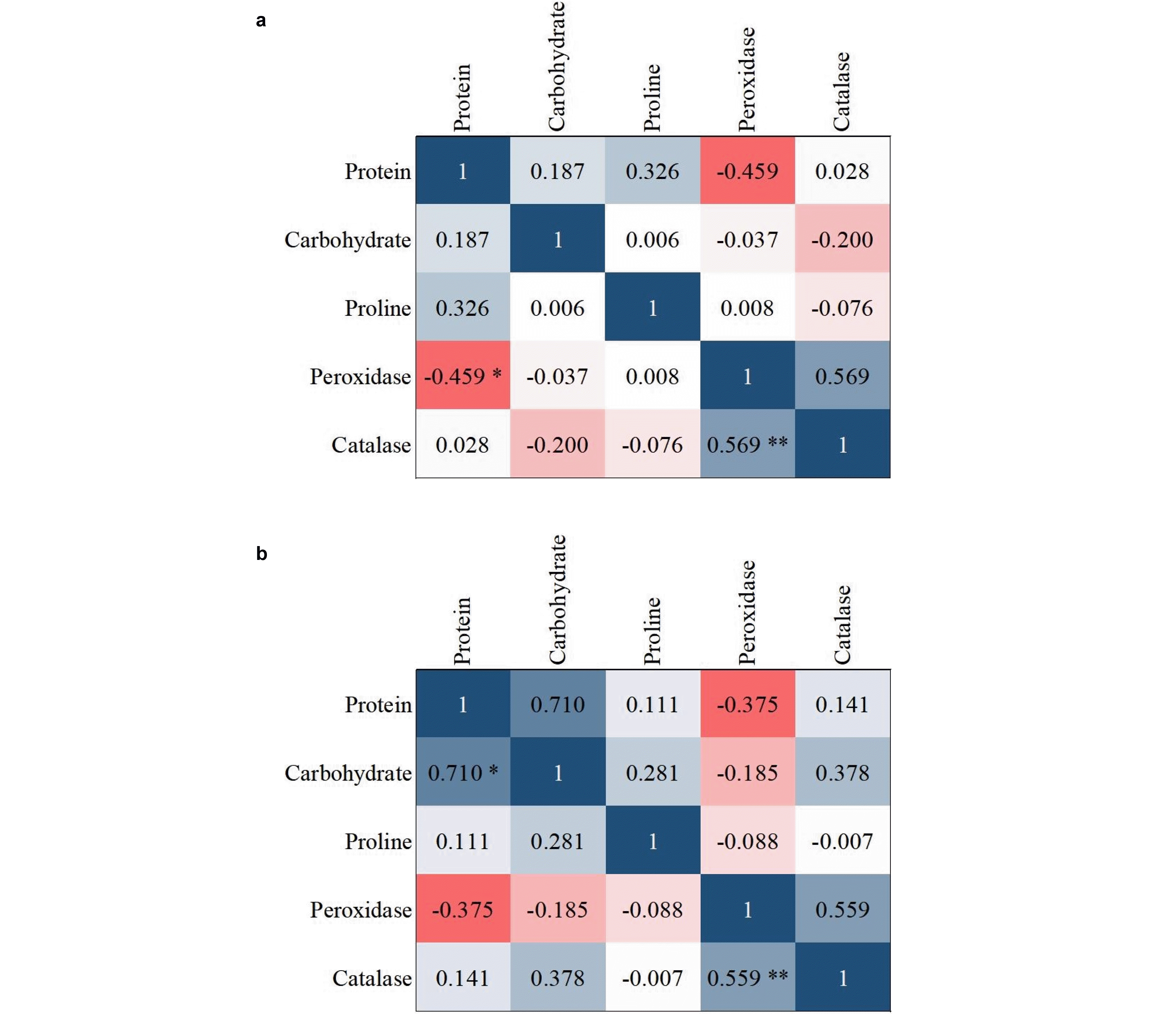
Insufficient information about the relationship between different traits leads to poor results and decisions. Hence, in designing and implementing breeding programs, it is ne-cessary to pay attention to the correlation between different traits. There are significantly positive and negative cor-relations between morphological traits and grain yield in most of the plants and crops such as wheat. Correlation coefficients between the morphological traits as well as the biochemical traits are given in
Figs. 8,
9, respectively. A positive coefficient is shown in blue color and a negative coefficient is shown in red color.
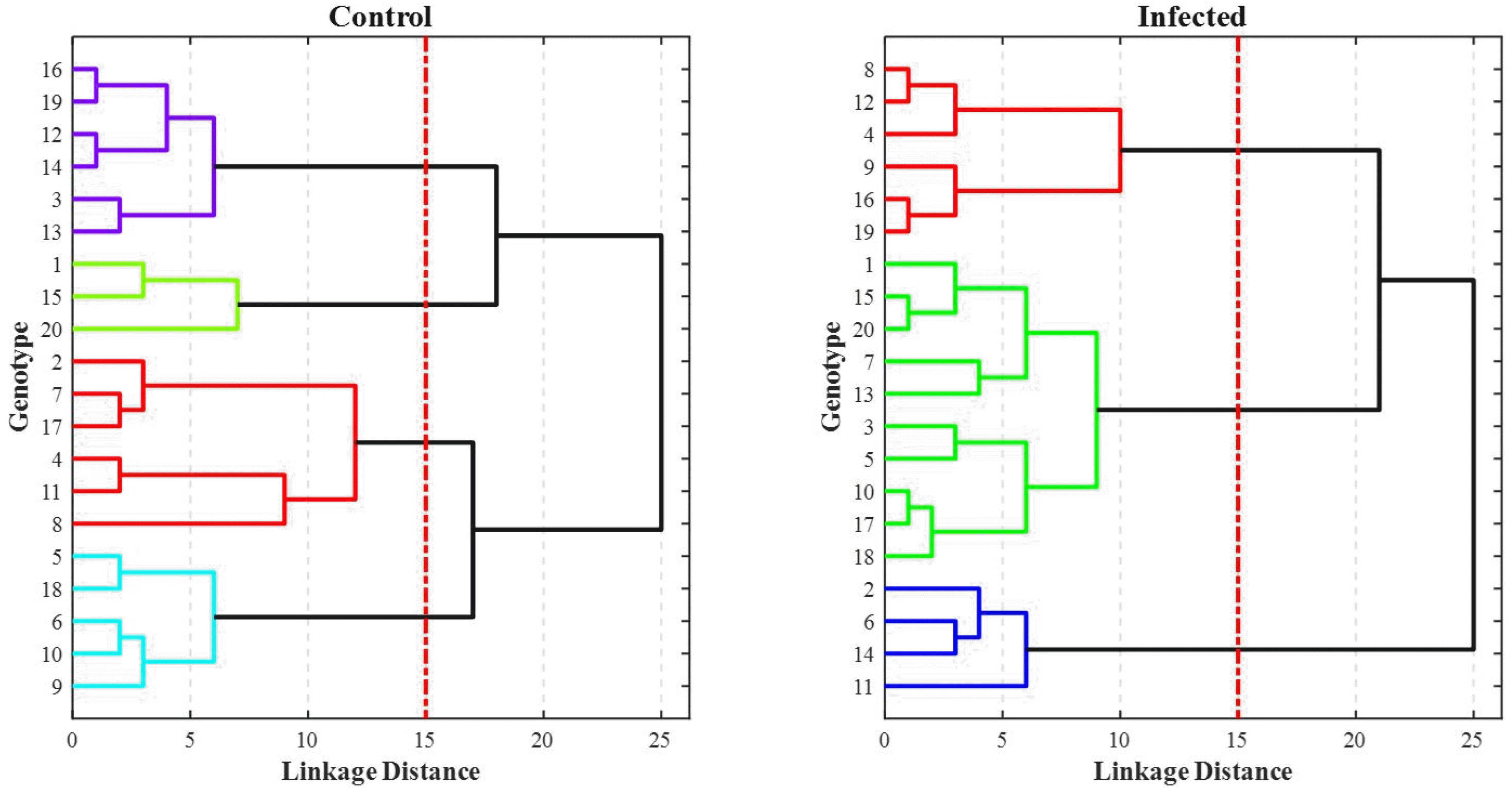
Cluster analysis is one of the most commonly used multivariate statistical methods in the study of genetic diversity and grouping of individuals and populations. Varieties in any plant have a high diversity in terms of quantitative and qualitative traits, and judgments based on one or more morphological traits that often have mono-genic heredity do not seem to be sufficient. The cut-off location was selected in each of the dendrograms, where the groups were well separated; i.e., below the distance of 15. Considering the proper structure of the grouping of the genotypes and the separation distance of the clusters from each other, the cut-off location was selected with four clusters for control and three clusters for infected condi-tions (
Fig. 10). In order to determine the characteristics of the studied traits, the mean values were calculated (
Tables 6,
7).
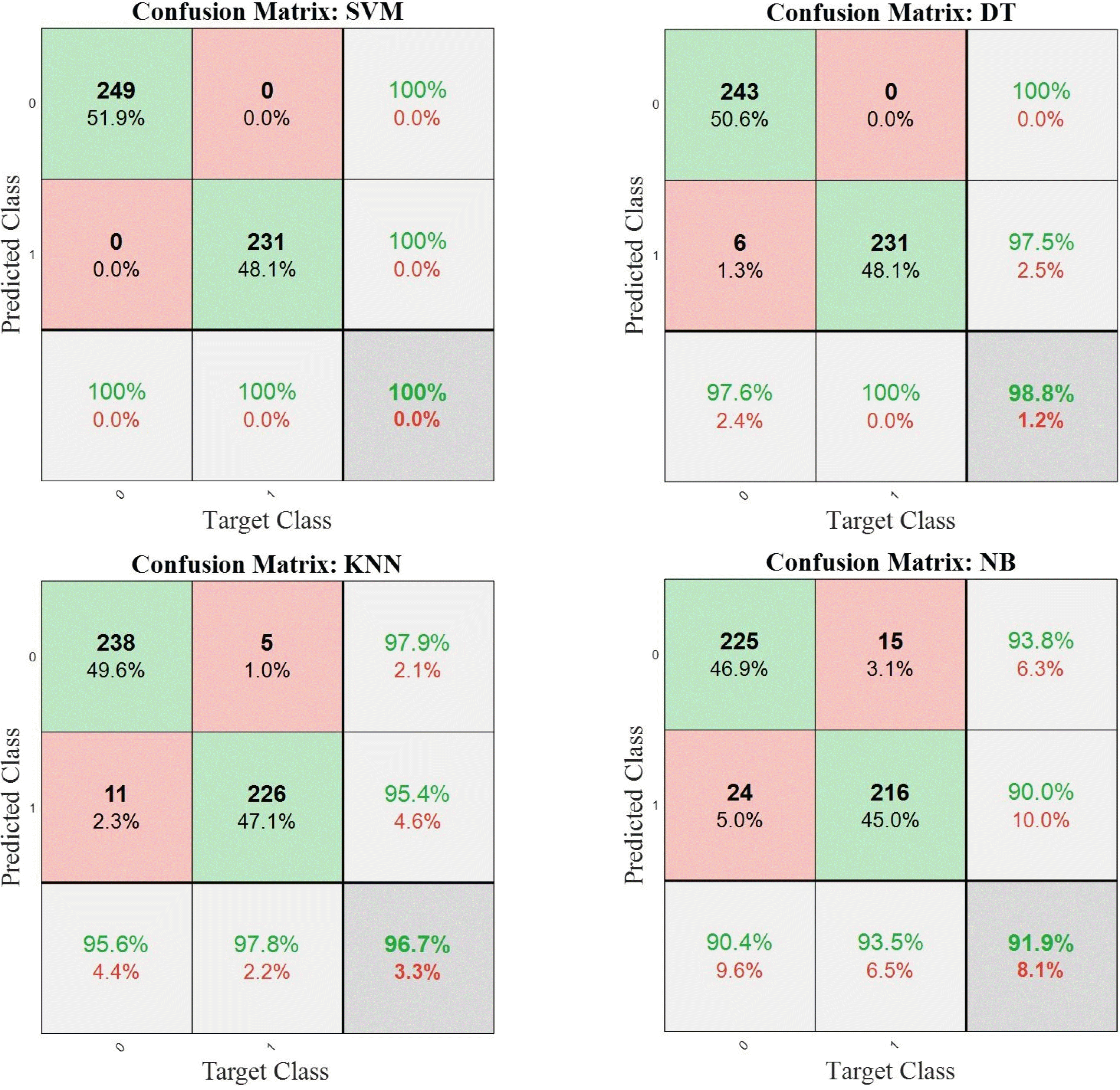
An SVM model was developed and trained using MATLAB. In order to show the effectiveness of the SVM classifier over other ML methods, three of the most popular classifiers were trained using the same datasets;
k-nearest neighbors (KNN), decision tree (DT), and Naive Bayes (NB) (
Golhani et al. 2018;
Li et al. 2018). To study the performance of the classifiers, the confusion matrices for all the models are given in
Fig. 11. Since the
k-fold cross-validation was applied to each procedure, the sum of the integers in each confusion matrix indicates the total size of samples (i.e., the sum of 5 confusion matrices from each run). The overall accuracy scores are given in dark-shaded down-right cells of each matrix. The diagonal integers (in green) show the correctly classified samples, or True Posi-tive (TP) and True Negative (TN), while the off-diagonal integers (in red) indicate the misclassified samples, or False Positive (FP) and False Negative (FN). The percentage of each integer with respect to the whole data size is given, as well. From
Fig. 11, it can be seen that the SVM and DT models can classify the data with accuracy scores of 100% and 98.8%, respectively. On the other hand, the KNN model misclassifies 3% of the same test data compared to 8% misclassification using the NB model. Therefore, for this specific classification task for FHB detection, the SVM model achieves a high accuracy score.
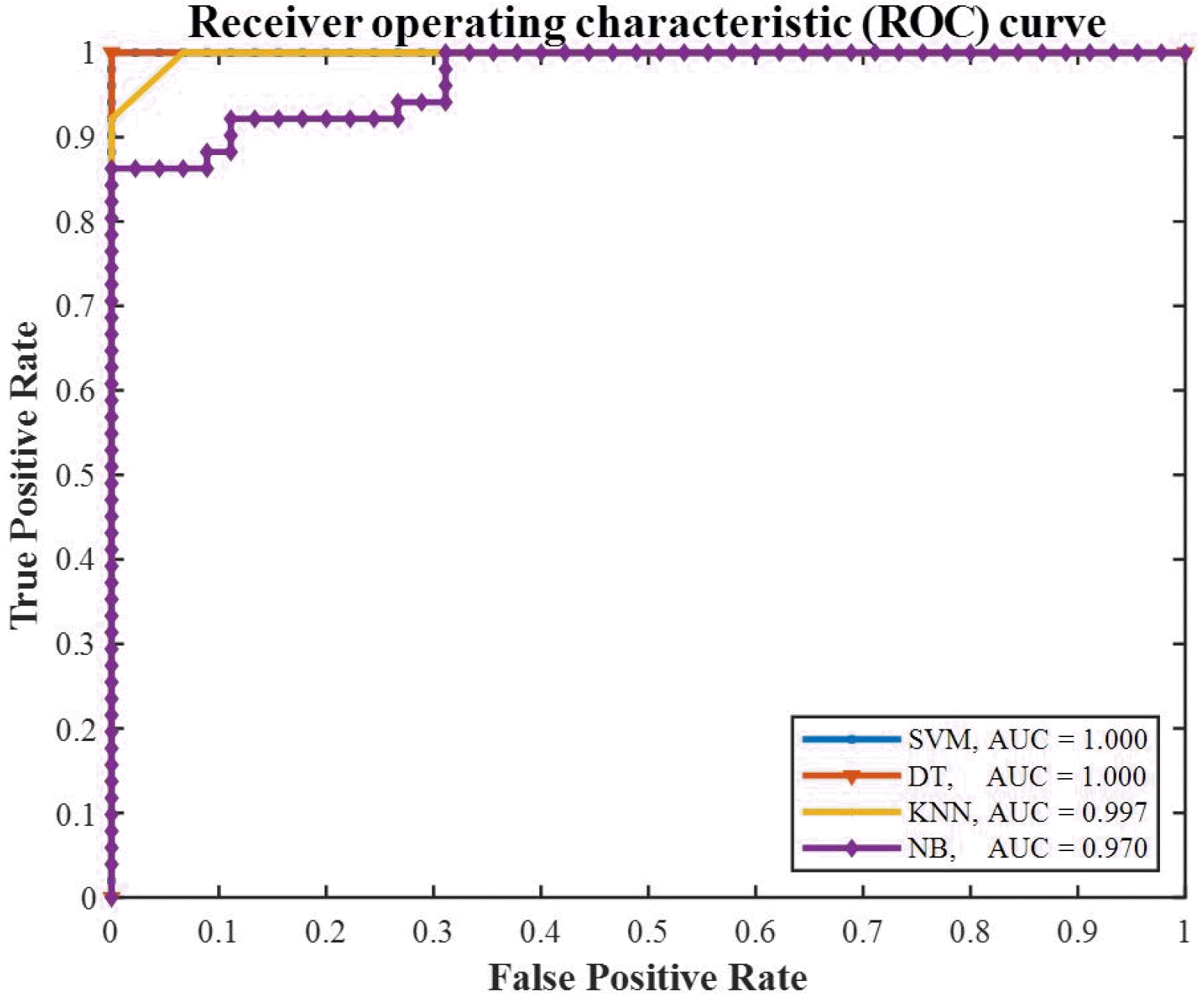
Furthermore, the receiver operating characteristic (ROC) curves are plotted in
Fig. 12 to compare the overall performance of the models. For a model with higher accuracy, the ROC curve is closer to the upper-left corner, and the area under the curve (AUC) is nearly equal to one. Considering the advances in software platforms, an accuracy of AUC ≥ 90% is expected from the selected classification models. It is obvious that SVM and DT models lead to a perfect classification, while NB and KNN have the lower AUC indices.
DISCUSSION
The results of the analysis of variance show that the studied lines have a significant difference for all traits at the 0.01 probability level (
Table 2). The interaction effect of genotype × stress in all the studied traits was significant at the 0.01 probability level. In other words, different geno-types show different effects at each level of stress. The mean value analysis results show that Genotypes with longer peduncle length, such as Genotype 2, have more resistance to the disease; this trait is one of the morpho-logical traits associated with FHB resistance (
Mesterhazy 1995). Peduncle length is one of the important agricultural traits that is effective not only in spike growth and plant cross-pollination but also has a significant effect on photo-synthesis and grain growth on spike (
Rao et al. 2007). Con-sidering that dense spikelets can trap more moisture be-tween the spikelets, it can be expected that the density of the spikelets is associated with the disease indicators (
Buerstmayr et al. 2000;
Rudd et al. 2001).
Taking into account that the high spike density can be considered as a positive and effective trait in yield; thus, resistant high-density spike genotypes, such as line 9 (3.64), could be desirable genotypes in breeding programs. A comparison of the mean values of stem height shows that the highest stem length was observed in normal conditions in the genotype 10 (59.1a). Holzapfel
et al. (
Holzapfel et al. 2008) showed that short spikes are the main reason for the sensitivity of genotypes to FHB. Therefore, genotypes with both shorter and better resistance to FHB spikes could be used in breeding programs. It was also observed that FHB leads to complexity and reduces the length of awn so that the lowest awn length was found in Genotype 16. Further-more, the comparison of grain weight and the number of seed per spike indicates that FHB caused a decrease in weight and seed number so that the genotype 7 had the lowest grain weight (
Fig. 7). The effect of FHB on the re-duction of wheat yield may be due to early prevalence and premature spikes maturation, which leads to wrinkling, re-duced grain quality, and reduced grain yield (
Stack 2000).
Analysis of variance shows that all biochemical traits were influenced by genotype effects (i.e., significance level at
P < 0.01). The results of the mean comparison using Duncan’s multiple range test (MRT) at a 0.05 sig-nificance level show that the occurrence of FHB caused a decrease in total protein. Increasing the rate of protein degradation may be due to the effects of stress (
Bolen and Baskakov 2001). Proline plays a major role in protecting cellular structures, which results in the stability of enzymes (
Kishor et al. 2005). Therefore, proline may increase due to its protective effect on proteins and enzymes. The highest and lowest amount of proline is observed for genotypes 2 and 7, respectively. The increase in carbohydrate acts as a metabolic message under stress conditions, which increases the expression of defense genes and reduces photosynthesis. In addition, carbohydrates play other ecological roles in plant protection against scarring as well as detoxification of external compounds. Carbohydrate content is high in geno-type 5, and the lowest in genotype 12. Further, the results of the mean value comparison show that the highest activity of the peroxidase enzyme is in the genotypes 18. Increasing activity of this enzyme is one of the defensive responses against oxidative stress, and peroxidase iso-enzymes play key roles in stress tolerance (
Ahmad and Haddad 2011). The lowest activity of the catalase enzyme is for genotype 19. Magbanua
et al. (
Magbanua et al. 2007) measured the effect of catalase enzyme activity of corn under the in-fluence of
Aspergillus flavus disease and showed that in-creased activity of this enzyme was affected by the disease. As a result, the genotypes with higher levels of carbohy-drates, proline, and enzymes can be used in breeding pro-grams.
A correlation between two traits may indicate that the genes controlling the two traits are located on the same chromosome. Regarding the qualitative traits, the correla-tion between traits depends exclusively on the location of the controlling gene and their relationship to chromo-somes, which can be described as the linkage of genes or non-allelic gene interaction. The existence of a correlation between the pairs of attributes is very important in breeding programs based on the traits. Between morphological traits in control and infected conditions, there was a positive and significant correlation between the number of seeds and the grain weight. In addition, a positive and significant correla-tion was observed between the peduncle and stem length (
Fig. 8). For biochemical traits in the control condition, there was a positive and significant correlation between catalase and protein and peroxidase. In the presence of disease, there was a correlation between proteins with car-bohydrates and peroxidase with catalase (
Fig. 9).
The results of the cluster analysis revealed a large vari-ation among the groups. In the control condition, lines 16, 19, 12, 14, 3, and 13 were placed in group one. This group had the lowest mean awn and peduncle length. Lines 1, 15, and 20 were in the second group, which had the highest mean for the awn length and the lowest mean spikelet density. Lines 2, 7, 17, 4, 11, and 8 were placed in the third group, which had the highest mean grain weight, number of seeds per spike, and spikelet density, which indicates good agricultural performance. Lines of the fourth group 5, 18, 6, 10, and 9 had the lowest grain weight and maximum stem length and showed lower yield. In inoculated (infected) conditions, the lines were clustered into three groups. In the first group, the lines 8, 12, 4, 9, 16, and 19 showed the lowest mean in most of the studied traits and this group showed lower yield, while the second group included lines 1, 15, 20, 7, 13, 3, 5, 10, 17, and 18 with a better per-formance compared to the other two groups. The third group consists of lines 2, 6, 14, and 11 with the highest mean values for traits such as peduncle length, the number of seeds per spike, stem length, and grain weight. This group had better performance. Genotypes 2, 11, 7, and 17 showed the highest yield at the control and inoculated conditions. So that they have been able to resist this level of stress and continue to perform better than other varieties, however, genotype 8 had the highest yield at the control level did not show resistance to infection and showed a decrease in yield. Genotype 12 has had a poor performance in both the control infected conditions.
The results of the machine learning models show that data-driven approaches would speed up the data processing and classification to automate FHB detection in durum wheat. Considering the advances in computer engineering as well as the availability of advanced sensors and cameras for data collection, real-time disease detection of plants would be possible through portable smartphones and lap-tops. The support vector machine and decision tree algorithms have the highest accuracies, which shows the potentials for implementing such algorithms in plant breeding and pro-tection programs.
In summary and conclusions, This study investigates the relationship between FHB disease resistance and mor-phological and biochemical traits in 20 durum wheat lines. The results of the analyses of variance show a significant genetic variation. Biochemical traits such as the activity of catalase, peroxidase, protein, and proline were affected by disease stress, however, the interaction of stress × genotype was not significant. There is a positive and significant correlation between the number of seeds and grain weight in control and disease conditions as well as a positive and significant correlation between peduncle length and stem length. Among the biochemical traits in the control con-dition, there is a positive and significant correlation be-tween the catalase, proteins, and peroxidases, as well as a positive and significant correlation between protein and carbohydrate. The cluster analysis of morphological traits shows that from the considered lines, the genotypes 2, 11, 7, 17 showed the highest yields at the control and inocu-lated levels. Therefore, they have been able to withstand the level of stress and continue to perform better than other varieties. While the genotype 8 had the highest yield at the control level, it did not show a decrease in yield and re-sistance to disease. The genotype 12 has had poor perform-ance at both control and infected stress levels.
This study aims to highlight the efficiency of machine learning (ML) methods in FHB detection using morpho-logical traits. On the other hand, it tries to lead the future studies to consider a broad range of ML-based applications in agricultural engineering, particularly, in automated crop monitoring and disease detection using camera-equipped systems. Therefore, in addition to the comparison of the statistical results, and inspired by the similar studies in engineering, data-driven machine learning (ML) models were trained and tested using the support vector machine (SVM), k-nearest neighbors (KNN), decision tree (DT), and Naive Bayes (NB) algorithms. The results showed that SVM has the highest classification accuracy, while NB does not perform well with 90.6% of accuracy. However, high accuracy of classification was obtained in this study using SVM, further investigations are essential to consider environmental and uncontrolled real-world conditions. Furthermore, more data is needed to improve the regulari-zation of the studied ML models.
Both morphological and biochemical traits were mea-sures carefully by a human in this study, using a digital camera, the future studies would fill this gap by developing the automated robotic system that can constantly monitor the crop using embedded vision-based deep learning (DL), and collect the morphological traits including peduncle length, awn length, stem length, spike density, grain weight, number of seeds. In addition, vision-based DL models would eliminate the necessity of traits such as grain weight by taking advantage of pixel-level investigation of images from the wheat plant.
-
FUNDING
This research received no external funding.
-
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Fig. 1FHB symptom development on durum wheat spikes (a-d).

Fig. 2FHB effects on durum wheat grains (small, shriveled pale, and white).

Fig. 3Recording the morphology of wheat plants using the robotic digital camera.
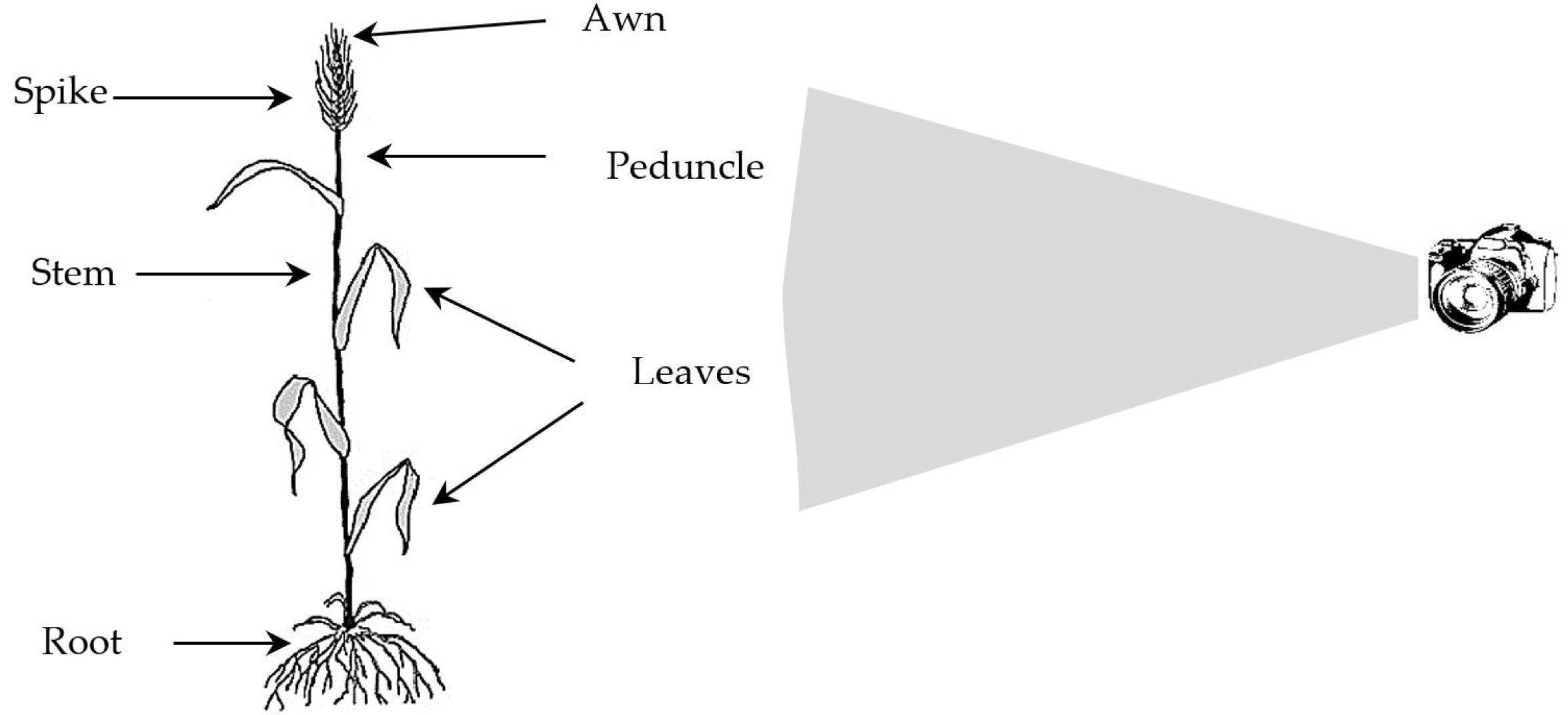
Fig. 4(a) Fusarium graminearum colony growth; (b) Macroconidial suspension preparation; (c) flowering stage of spikes; (d) covered spiked with plastic bags.

Fig. 5Growth of the Fusarium fungus. (a) Control; (b) Infected.
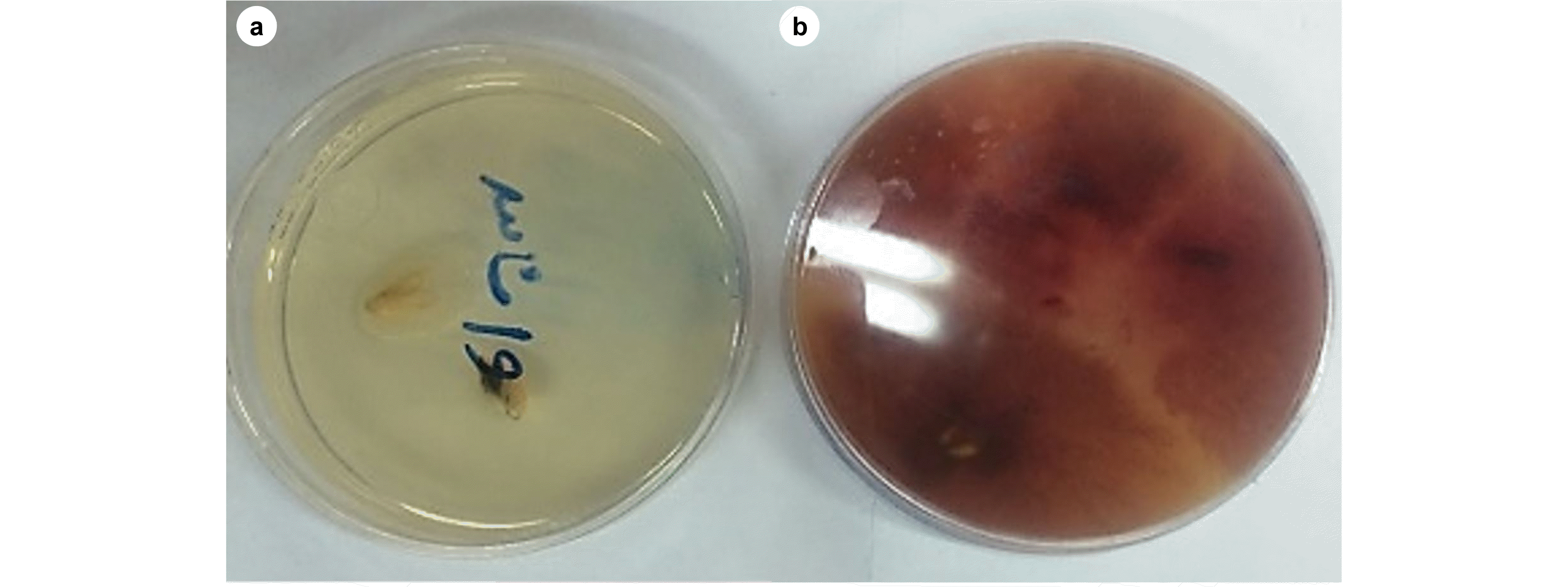
Fig. 6The architecture of the SVM classifier using the morphological traits.
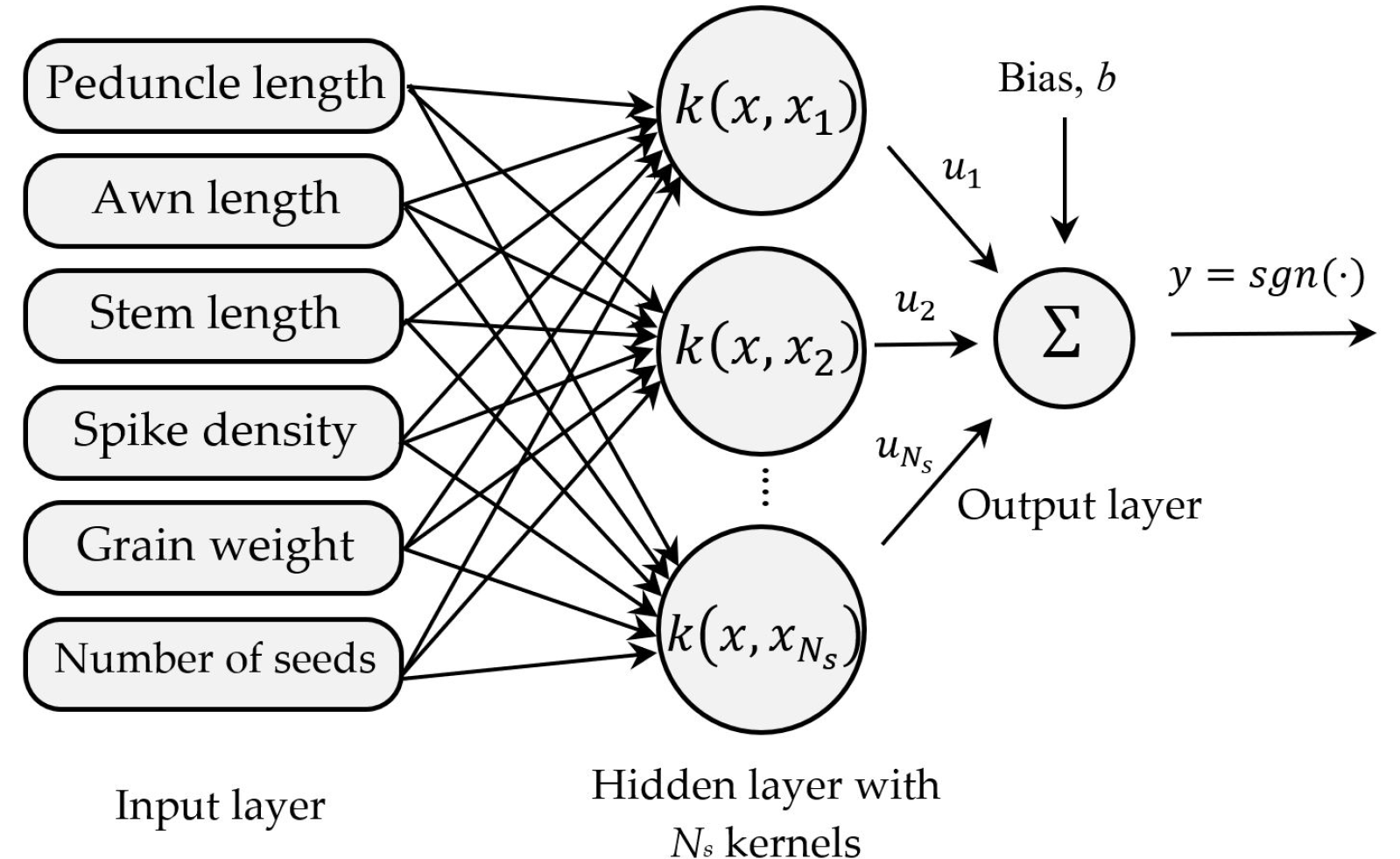
Fig. 7Mean values of the number of seeds per spike in different genotypes.
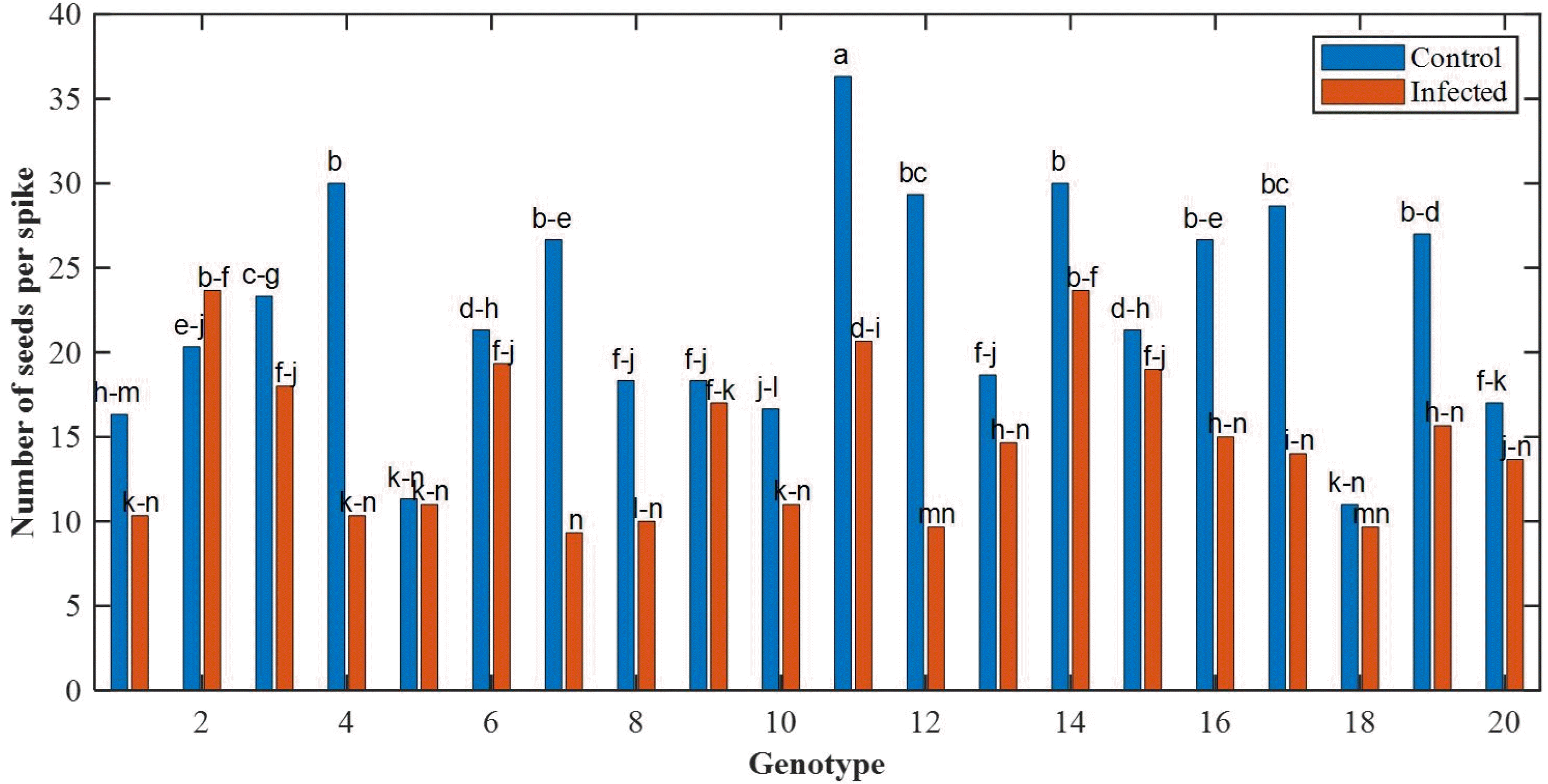
Fig. 8Correlation coefficients between the morphological traits (* and ** indicate the significance level at 0.05 and 0.01 probability levels, respectively). (a) Control; (b) Infected.
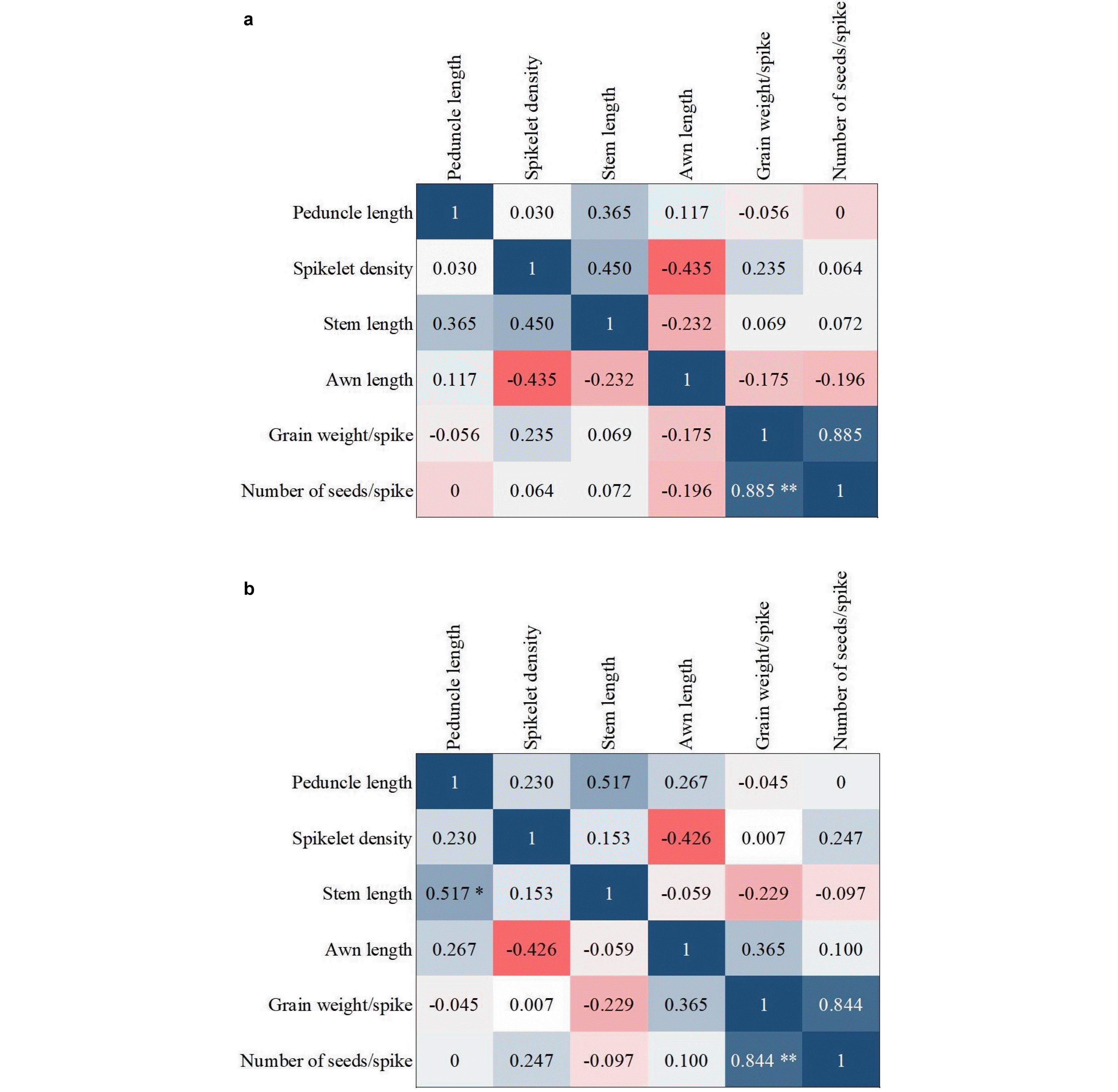
Fig. 9Correlation coefficients between the biochemical traits (* and ** indicate the significance level at the 0.05 and 0.01 probability levels, respectively). (a) Control; (b) Infected.
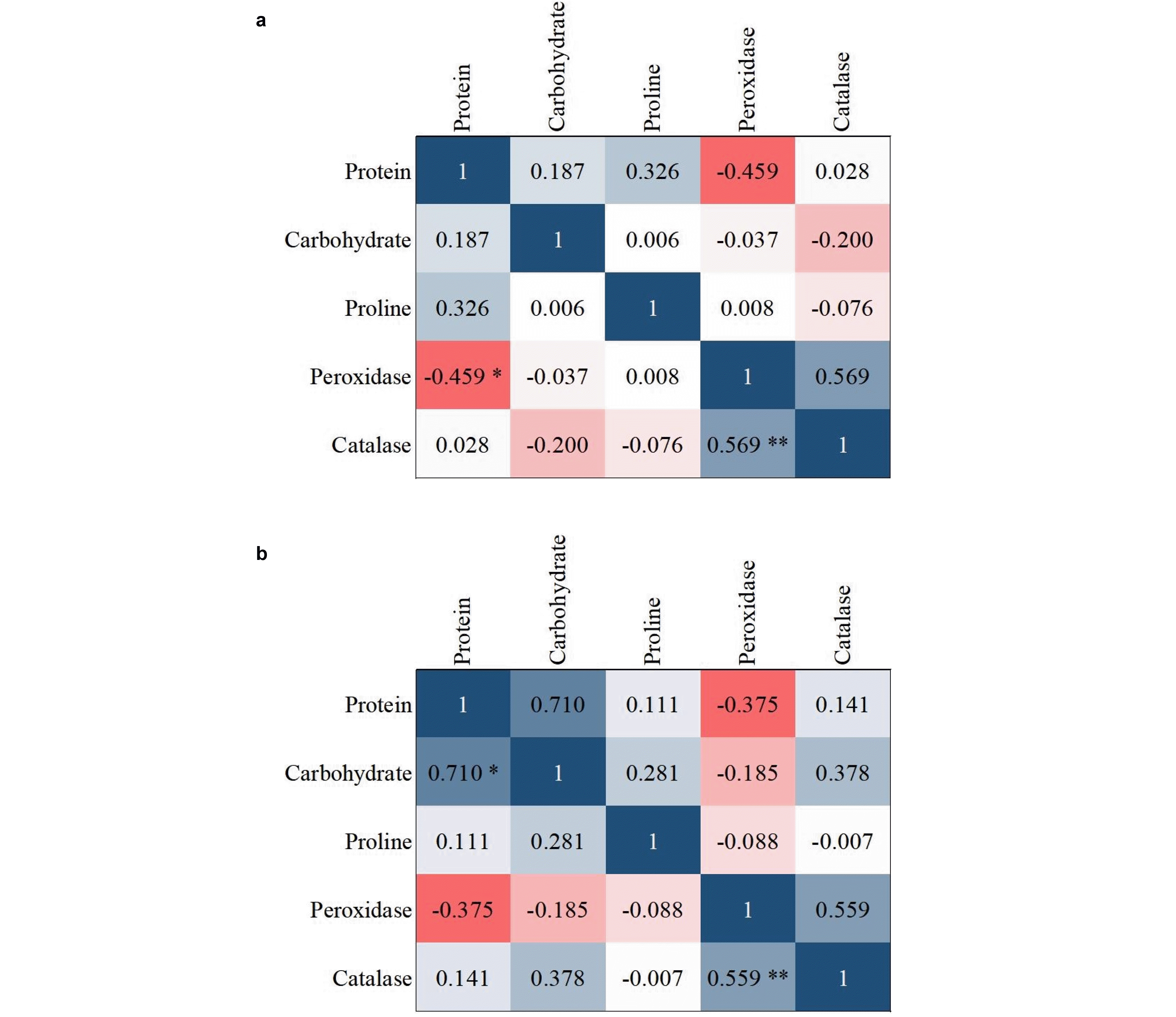
Fig. 10Hierarchical cluster analysis based on the morphological traits and Ward’s method.
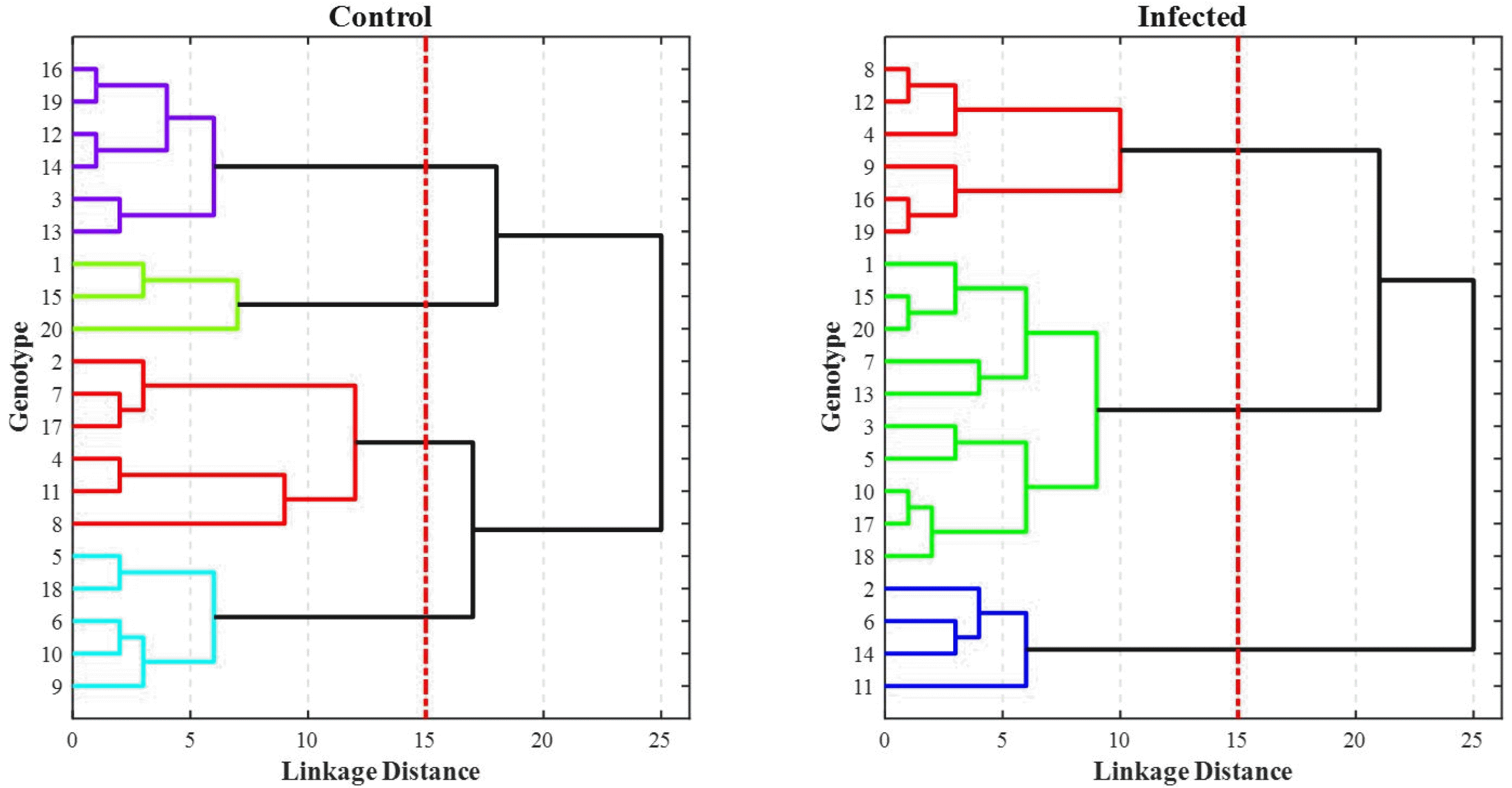
Fig. 11Total confusion matrices of the SVM, DT, KNN, and NB classifiers using test datasets (‘0’ and ‘1’ indicate control and infected samples, respectively).
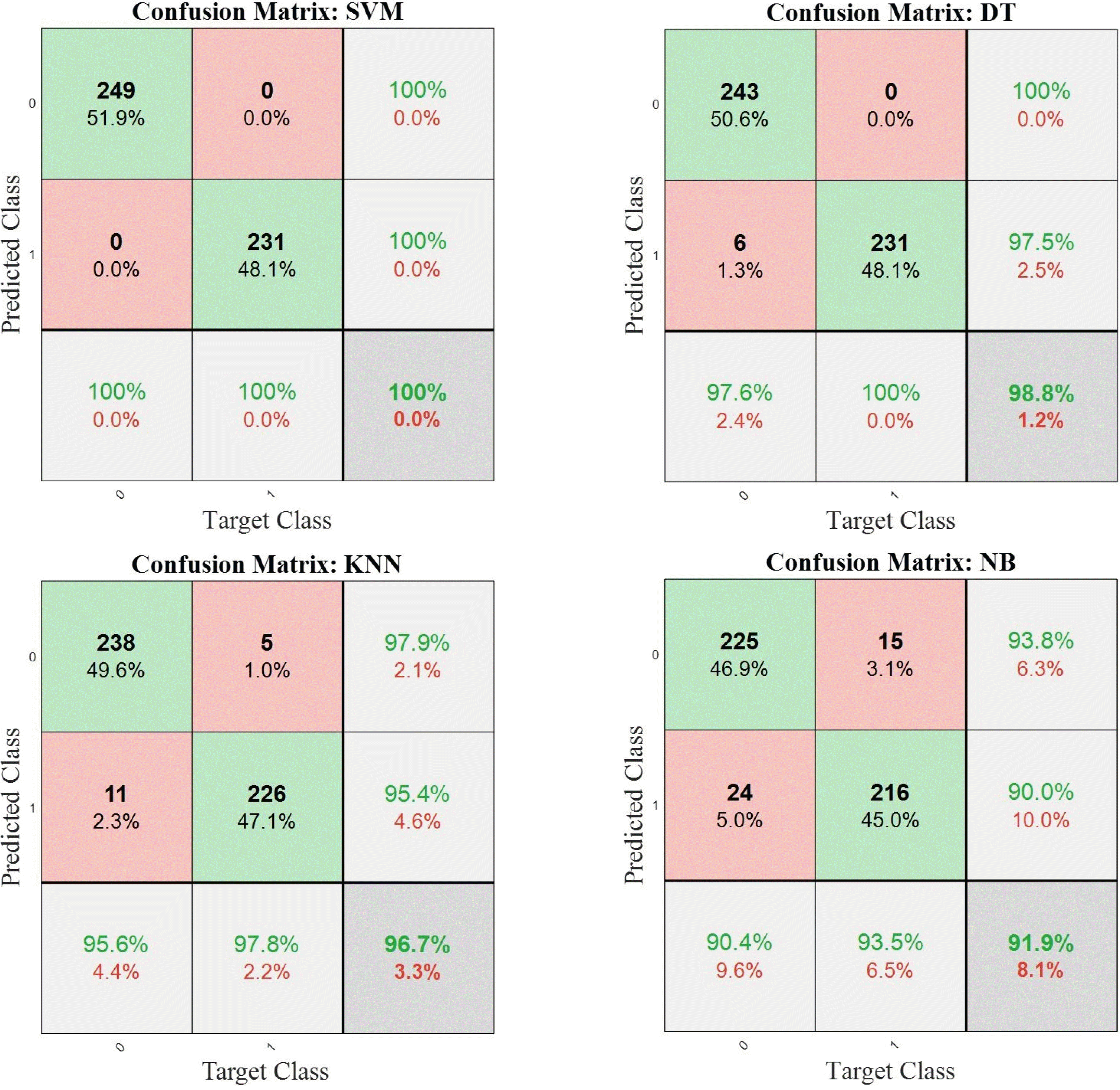
Fig. 12The ROC curves of the selected classifiers; SVM, DT, KNN, and NB.
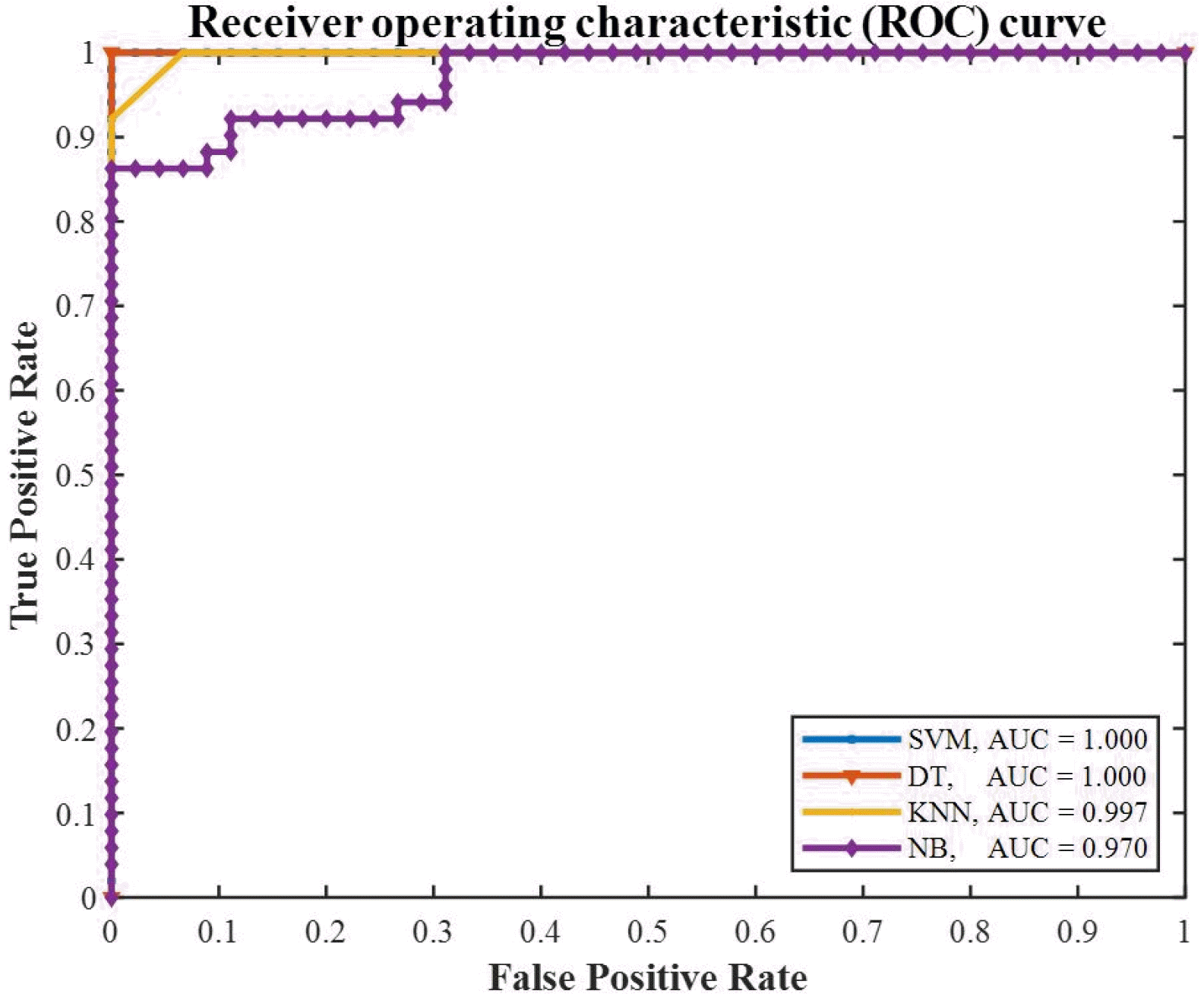
Table 1
Table 1
|
Line number |
Family tree |
|
1 |
SRN_1/6/FGO/DOM//NACH/5/ALTAR84/4/GARZA/AFN//CRA/3/GGOVZ394/7/GEDIZ/FGO//GTA/3/CN D/8/GREEN_38/9/2*STOT//ALTAR 84/ALDCDSSOOBOO227T-0TOPY-0B-6Y-0M-0Y-1B |
|
2 |
ALTAR 84/BINTEPE 85/3/ALTAR 84/STINT// SILVER_45/4/LHNKE/RASCON//CONA-DCDSS99B 01265T0TOPY-0M-0Y-12Y-0M-0Y-1M-0Y |
|
3 |
AINZEN-1/SORD_3CDSS99B00317S-0M-0Y-104Y-0M0Y |
|
4 |
STAT//ALTAR 84/ALD*2/3/AUK/GUIL// GREENCDSS00Y00786-0TOPB-9Y-0BLR-5Y-0B-0Y-1M-0Y |
|
5 |
LLARETA
INIA/YEBAS_8/3MINIMUS_6/PLATA_16//MMERCDSS00Y01047T-0TOPB-5Y-OBLR-1Y-0B-0Y-1B-0Y 1B-0Y |
|
6 |
GEDIZ/FGO//GTA/3/SRN_1/4TOTUS/5/ENTE/MEXI_2/HUI/3/YAV_/GEDIZ/6/SOMBRA_20/7/STAT//ALTAR84/ALDCDSS97Y00835 S 0TOPM-4Y-0M-0Y-0B-0B-3Y-0BLR-1Y-0B27 RASCON_21/3/MQUE/ALO// FOJACDSS94Y00099 -7M-0Y-0B-1Y-0B-0BLR-5Y-0B |
|
7 |
GAUNT_10/SNITANCDSS97Y0038S-4Y-0M-0Y-0B-0B-3Y-0BLR-1Y-0B |
|
8 |
ALTAR 84/STINT// SILVER_45/3/CBC 503 CHLE/4/AUK/GUIL//GREENCDSS99B0 1115T-0TOPY-0M-0Y-Y-0M-0Y-1B-0Y |
|
9 |
SRN_1/6/FGO/DOM//NACH/5/ALTAR84/4/GARZA/AFN//CRA/3/GGOVZ394/7/GEDIZ/FGO//GTA/3/CNDO/8/GREEN_38/9/2*STOT//ALTAR-84/ALDSS00B00227T-0TOPY-0B |
|
10 |
STOT//ALTAR 84/ALD/3/GREEN_18/FOCHA_1//AIR0N_1CDSS 99B00467S0-0M-0Y-75Y-0M-0Y-2M-0Y |
|
11 |
RASCON_21/3/MQUE/ALO//FOJA/4/GREEN_38/BUSHEN_4/5/CADOO/BOOMER_33CDSS99B01055T-0TOPY0M-0Y-10Y-0M-0Y-1M-0Y |
|
12 |
SOMO/CROC_4//LOTUS_I/3KITTI/4/STOT//ALTAR 84/ALDCDSS99Y0063S-0M-0Y-34Y-0M-0Y-0B |
|
13 |
CMH82A.1062/3/GGOVZ394//SBA81/PLC/4/AAZ_1/CREX/5/HUI//CIT71/CH/6/STOT/ALTAR84/ALD CDSS99Y00643S-0M-0Y-16Y-0M-0Y-0B |
|
14 |
Dehdasht |
|
15 |
ACUATICO_1/RASCON_33//ACUATICO_1/3/AJAIA_12/F3LOCAL.(SEL.ETHIO.135.85)//PLATA_13CDSS96Y00 |
|
16 |
SAIMAREH |
|
17 |
STOT//ALTAR 84/ALD/3/THB/CEP7780//2*MUSK_4CDSS99Y00366 S-3Y-0M-0Y-0BLR-1Y-0B-1M-0Y |
|
18 |
ALTAR 84/STINT//SILVER_ 45/3/STOT// ALTAR 84/ALDCDSS99Y 0037S-7Y-0M-0Y-0BLR-0B-1B-0Y |
|
19 |
RASCON_21/3/MQUE/ALO//FOJACDSS94Y00099S-7M-0Y-0B-1Y-0B-0BLR-5Y-0B |
|
20 |
PLATA_8/4/GARZA/AFN//CRA/3/GTA/5/RASCON/6/CADO/BOOMER_33/7/STOT//ALTAR84/ALDCDSS99B0 0843S-0TOPY-0M-0Y-5Y-0M-0Y-1B-0Y |
Table 2Variance analysis for morphological traits of durum wheat lines (units: cm, gram).
Table 2
|
Variation source |
Degree of freedom |
Peduncle length |
Spikelet density |
Stem
length |
Awn length |
Grain weight per spike |
Number of seeds per spike |
|
Genotype |
19 |
10** |
0.358** |
40.79** |
7.59** |
0.071** |
132.51** |
|
Stress |
1 |
36.41** |
0.276** |
64.68** |
15.48** |
8.17** |
1755.7** |
|
Genotype × Stress |
19 |
2.41** |
0.172** |
37.33** |
1.37** |
0.052** |
69.95** |
|
Error |
78 |
1.017 |
0.043 |
10.14 |
0.249 |
0.12 |
11.73 |
|
Variation factor (%) |
|
10.67 |
10.52 |
18.96 |
13.4 |
9.5 |
8.21 |
Table 3Mean values of the morphological traits, using Duncan’s method at the significance level of P < 0.05 (units: cm, gram).
Table 3
|
Genotype |
Peduncle length |
|
Spikelet density |
|
Stem length |
|
Awn length |
|
Grain weight
per spike |
|
Number of seeds per spike |
|
|
|
|
|
|
|
0 |
1 |
0 |
1 |
0 |
1 |
0 |
1 |
0 |
1 |
0 |
1 |
|
1 |
28a-h |
27.4d-k |
|
2.23ij |
2.35h-j |
|
47.16h-j |
55.6e-d |
|
11.33cd |
11.1cd |
|
0.70e-h |
0.23n-p |
|
16.33h-m |
10.33k-n |
|
2 |
29.9a |
27.8b-i |
|
2.47e-j |
2.61d-j |
|
52b-i |
54.2a-g |
|
10.50d-g |
8.9i-o |
|
0.84c-f |
0.54h-l |
|
20.33e-j |
23.66b-f |
|
3 |
24.5n |
25.3l-n |
|
2.44e-j |
2.44e-j |
|
48.1g-j |
50.03c-j |
|
8.70j-o |
8.1o-q |
|
0.93a-d |
0.41k-p |
|
23.33c-g |
18f-j |
|
4 |
26.8f-m |
25.5i-n |
|
3.05bc |
3.05bc |
|
56.4a-c |
52.6b-j |
|
9.80f-i |
7.5pq |
|
1.7a |
0.23n-p |
|
30b |
10.33k-n |
|
5 |
29.5a-c |
28a-h |
|
2.51e-j |
2.32h-j |
|
56a-d |
54.8a-f |
|
8.3n-p |
8.1o-q |
|
0.65f-i |
0.25n-p |
|
11.33k-n |
11k-n |
|
6 |
29.1a-d |
28.6a-f |
|
2.43e-j |
2.78b-j |
|
53.1a-h |
50.4c-j |
|
9.3i-o |
10.7c-e |
|
0.63f-j |
0.44j-n |
|
21.33d-h |
19.33f-j |
|
7 |
29.8ab |
26.3h-n |
|
2.63d-i |
2.28h-j |
|
59a |
47.33h-j |
|
11.66bc |
11.1cd |
|
1.06ab |
0.21p |
|
26.66b-e |
9.33n |
|
8 |
27.6c-j |
27e-m |
|
3.64a |
2.80b-f |
|
52.66b-i |
50.7c-j |
|
8.1o-q |
8.4l-p |
|
0.90a-e |
0.23n-p |
|
18.33f-j |
10l-n |
|
9 |
28.1a-h |
27e-m |
|
2.79a-g |
2.45e-j |
|
52.8a-i |
46.5ij |
|
10.2e-h |
9.4h-k |
|
0.704e-h |
0.39k-p |
|
18.32f-j |
17f-k |
|
10 |
28.5a-g |
28.9a-e |
|
2.57d-j |
2.83b-e |
|
59.1a |
51.6c-i |
|
10e-h |
8.9k-o |
|
0.71e-h |
0.25n-p |
|
16.66j-l |
11k-n |
|
11 |
26.8f-m |
28.8a-f |
|
2.67c-h |
3.15b |
|
56.3a-c |
53.9a-g |
|
9.2i-n |
10e-h |
|
1.10a |
0.47i-m |
|
36.33a |
20.66d-i |
|
12 |
27.1d-l |
25mn |
|
2.30h-j |
2.56d-j |
|
52.03b-i |
52.46b-i |
|
9.6g-j |
8.4k-p |
|
0.94a-d |
0.22op |
|
29.33bc |
9.66mn |
|
13 |
26.5j-m |
25.5k-n |
|
2.31h-j |
2.25h-j |
|
48g-j |
49.1g-j |
|
9.3i-o |
8.9i-o |
|
0.075d-g |
0.33l-p |
|
18.66f-j |
14.66h-n |
|
14 |
28.1a-h |
25mn |
|
2.66c-h |
2.96b-d |
|
50.3c-j |
49.7d-j |
|
8.8i-o |
8.7j-o |
|
1.02a-c |
0.54h-l |
|
30b |
23.66b-f |
|
15 |
29.1a-d |
28.6a-f |
|
2.29h-j |
2.55d-j |
|
44.66j |
52.96a-i |
|
12.2ab |
11.3cd |
|
0.906a-e |
0.43j-o |
|
21.33d-h |
19f-j |
|
16 |
27.3d-k |
25.8j-n |
|
2.40f-j |
2.77a-g |
|
50.8c-j |
44.5j |
|
9.3h-l |
8.8i-o |
|
1a-c |
0.34k-p |
|
26.66b-e |
15h-n |
|
17 |
29.9a |
28a-h |
|
2.49e-j |
2.94b-d |
|
55.3a-e |
52.9a-i |
|
9.3h-l |
8.3m-p |
|
1.1a |
0.32m-p |
|
28.66bc |
14i-n |
|
18 |
27.8c-i |
26.8f-n |
|
2.59d-j |
2.47e-j |
|
58.1ab |
53a-h |
|
10.6d-f |
10.1e-h |
|
0.55j-k |
0.22op |
|
11k-n |
9.66mn |
|
19 |
27.6c-j |
25.5k-n |
|
2.37g-j |
2.78b-j |
|
52b-i |
48.6f-j |
|
9.6g-j |
7.4q |
|
.86b-e |
0.36k-p |
|
27b-d |
15.66h-n |
|
20 |
25.6k-n |
24.5n |
|
2.21j |
2.62d-j |
|
47.3h-j |
51.03c-i |
|
12.5a |
10.2e-h |
|
0.72e-h |
0.31m-p |
|
17f-k |
13.66j-n |
Table 4Variance analysis for biochemical traits of durum wheat lines.
Table 4
|
Variation source |
Degree of freedom |
Protein |
Carbohydrate |
Proline |
Peroxidase |
Catalase |
|
Genotype |
19 |
3.04** |
2.63** |
39.93** |
4.81** |
11.07** |
|
Stress |
1 |
5.05 ns |
6.43 ns |
12.6 ns |
6.53 ns |
8.53 ns |
|
Genotype × Stress |
19 |
3.29 ns |
9.55 ns |
18.8 ns |
2.13 ns |
8.03 ns |
|
Error |
78 |
0.08 |
0.091 |
11.83 |
0.51 |
0.58 |
|
Coefficient of Variation |
|
13.8% |
12.2% |
15.2% |
8.92% |
7.12% |
Table 5Mean values of the biochemical traits using the Duncan method at the 0.05 significance level.
Table 5
|
Genotype |
Protein |
Carbohydrate |
Proline |
Peroxidase |
Catalase |
|
1 |
0.227a-d
|
62.1b-f
|
4.79a-d
|
4.53bc
|
0.48a-c
|
|
2 |
0.26a
|
78.3a-d
|
6.12a
|
3.89c
|
0.16a-c
|
|
3 |
0.25ab
|
85.3a
|
5.04a-d
|
4.89a-c
|
0.64ab
|
|
4 |
0.191cd
|
72.2a-e
|
3.79a-e
|
4.01c
|
0.42a-c
|
|
5 |
0.228a-d
|
88.9a
|
3.07b-e
|
7.30a
|
0.66ab
|
|
6 |
0.217a-d
|
82.8ab
|
5.03a-d
|
6.42a-c
|
0.69a
|
|
7 |
0.186d
|
53.4ef
|
1.94e
|
4.82a-c
|
0.19a-c
|
|
8 |
0.212a-d
|
54.3ef
|
3.63a-e
|
6.20a-c
|
0.28a-c
|
|
9 |
0.24a-c
|
71.5a-e
|
2.35de
|
4.22c
|
0.39a-c
|
|
10 |
0.223a-d
|
85.2a
|
5.29a-c
|
5.26a-c
|
0.40a-c
|
|
11 |
0.213a-d
|
73.3a-e
|
2.47de
|
6.41a-c
|
0.71a
|
|
12 |
0.173d
|
44.3f
|
2.71ce
|
6.99ab
|
0.35a-c
|
|
13 |
0.185d
|
56.4ef
|
5.31a-c
|
5.56a-c
|
0.12bc
|
|
14 |
0.219a-d
|
60.5d-f
|
3.58a-e
|
6.50a-c
|
0.69a
|
|
15 |
0.217a-d
|
71a-e
|
5.62ab
|
5.48a-c
|
0.52ab
|
|
16 |
0.194b-d
|
82.5a-c
|
3.04b-e
|
4.96a-c
|
0.13bc
|
|
17 |
0.215a-d
|
82.3a-c
|
2.55de
|
4.67a-c
|
0.24a-c
|
|
18 |
0.179d
|
58.6d-f
|
4.75ad
|
7.31a
|
0.56ab
|
|
19 |
0.208a-d
|
61.5c-f
|
4.47a-e
|
4.17c
|
0.80c
|
|
20 |
0.225a-d
|
70.6a-e
|
2.36de
|
5.27a-c
|
0.62ab
|
Table 6The mean and standard deviation for morphological traits of durum wheat (infected).
Table 6
|
Spike |
Lines |
|
Peduncle length |
Spikelet density |
Stem length |
Awn length |
Grain weight per spike |
Number of seeds per spike |
|
1 |
8, 12, 4, 9, 16, 19 |
µ |
26.05 |
2.73 |
49.23 |
8.33 |
0.29 |
12.94 |
|
σ |
0.73 |
0.19 |
3.00 |
0.69 |
0.07 |
3.01 |
|
2 |
1, 15, 20, 7, 13, 3, 5, 10, 17, 18 |
µ |
26.95 |
2.50 |
51.85 |
9.64 |
0.30 |
13.06 |
|
σ |
7.66 |
0.70 |
14.24 |
2.83 |
0.10 |
4.23 |
|
3 |
2, 6, 14, 11 |
µ |
27.57 |
2.87 |
52.09 |
9.59 |
0.50 |
21.83 |
|
|
σ |
8.08 |
0.89 |
15.25 |
2.81 |
0.17 |
7.24 |
Table 7The mean and standard deviation for morphological traits of durum wheat (control).
Table 7
|
Spike |
Lines |
|
Peduncle length |
Spikelet density |
Stem length [cm] |
Awn length
[cm] |
Grain weight per spike [gr] |
Number of seeds per spike |
|
1 |
16, 19, 12, 14, 3, 13 |
µ |
26.88 |
2.42 |
50.22 |
9.26 |
0.92 |
25.83 |
|
σ |
1.18 |
0.12 |
1.63 |
0.37 |
0.09 |
3.86 |
|
2 |
1, 15, 20 |
µ |
27.60 |
2.25 |
46.38 |
12.06 |
0.78 |
18.22 |
|
|
σ |
1.47 |
0.03 |
1.21 |
0.50 |
0.09 |
2.22 |
|
3 |
2, 7, 17, 4, 11, 8 |
µ |
28.49 |
2.83 |
55.32 |
9.81 |
1.02 |
26.72 |
|
σ |
1.42 |
0.41 |
2.40 |
1.10 |
0.10 |
6.03 |
|
4 |
5, 18, 6, 10, 9 |
µ |
28.63 |
2.58 |
55.87 |
9.73 |
0.65 |
15.73 |
|
σ |
0.62 |
0.12 |
2.55 |
0.82 |
0.06 |
4.02 |
References
- Adams J. 1921. Observations on wheat scab in Pennsylvania and its pathological histology. Phytopathology. 11: 115- U119.
- Ahmad ST, Haddad R. 2011. Study of silicon effects on anti-oxidant enzyme activities and osmotic adjustment of wheat under drought stress. Czech J. Genet. Plant Breed.. 47: 17-27.
- Alisaac E, Behmann J, Kuska MT, Dehne HW, Mahlein AK. 2018. Hyperspectral quantification of wheat resistance to Fusarium head blight: Comparison of two Fusarium species. Eur. J. Plant Pathol.. 152: 869-884.
- Azimi M, Eslamlou AD, Pekcan G. 2020. Data-driven struc-tural health monitoring and damage detection through deep learning: State-of-the-art review. Sensors. 20: 1-34.
- Azimi M, Pekcan G. 2019. Structural health monitoring using extremely compressed data through deep learning. Comput aided Civ. Inf.. 35: 597-614.
- Azimi N. 2018. Assessment of genetic variability of durum wheat resistance to Fusarium head blight disease using molecular markers. M. Sc.. University of Mohaghegh Ardabili. Ardabil, Iran.
- Bai G, Shaner G. 1994. Scab of wheat: Prospects for control. Plant Dis.. 78: 760-766.
- Bates LS, Waldren RP, Teare ID. 1973. Rapid determination of free proline for water-stress studies. Plant Soil. 39: 205-207.
- Bernardo AN, Ma H, Zhang D, Bai G. 2012. Single nu-cleotide polymorphism in wheat chromosome region harboring Fhb1 for Fusarium head blight resistance. Mol. Breed.. 29: 477-488.
- Bolen DW, Baskakov IV. 2001. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol.. 310: 955-963.
- Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.. 72: 248-254.
- Buerstmayr H, Steiner B, Lemmens M, Ruckenbauer P. 2000. Resistance to Fusarium head blight in winter wheat: Heritability and trait associations. Crop Sci.. 40: 1012-1018.
- Bushnell WR, Hazen BE, Pritsch C, Leonard K. 2003. His-tology and physiology of Fusarium head blight. Fusarium head blight of wheat and barley. APS Press, U.S.A..
- Golhani K, Balasundram SK, Vadamalai G, Pradhan B. 2018. A review of neural networks in plant disease detection using hyperspectral data. Inf. Process. Agric.. 5: 354-371.
- Holzapfel J, Voss HH, Miedaner T, Korzun V, Häberle J, Schweizer G, et al. 2008. Inheritance of resistance to Fusarium head blight in three European winter wheat populations. Theor. Appl. Genet.. 117: 1119-1128.
- Hsu CW, Lin CJ. 2002. A comparison of methods for multi-class support vector machines. IEEE Trans. Neural Netw.. 13: 415-425.
- Irigoyen JJ, Einerich DW, Sánchez-Díaz M. 1992. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant.. 84: 55-60.
- Kar M, Mishra D. 1976. Catalase, peroxidase, and polyphe-noloxidase activities during rice leaf senescence. Plant Physiol.. 57: 315-319.
- Kishor PBK, Sangam S, Amrutha RN, Laxmi PS, Naidu KR, Rao KRSS, et al. 2005. Regulation of proline biosyn-thesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress toler-ance. Curr. Sci.. 88: 424-438.
- Lenc L. 2015. Fusarium Head Blight (FHB) and Fusarium populations in grain of winter wheat grown in different cultivation systems. J. Plant Prot. Res.. 55: 94-109.
- Li J, Xi B, Li Y, Du Q, Wang K. 2018. Hyperspectral classi-fication based on texture feature enhancement and deep belief networks. Remote Sens.. 10: 1-20.
- Magbanua ZV, De Moraes CM, Brooks TD, Williams WP, Luthe DS. 2007. Is catalase activity one of the factors as-sociated with maize resistance to Aspergillus flavus? Mol. Plant Microbe Interact.. 20: 697-706.
- Matthies A, Buchenauer H. 2000. Effect of tebuconazole (Folicur) and prochloraz (Sportac) treatments on Fusarium head scab development, yield and deoxynivalenol (DON) content in grains of wheat following artificial inoculation with Fusarium Culmorum. J. Plant Dis. Protect.. 107: 33- 52.
- McMullen M, Jones R, Gallenberg D. 1997. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis.. 81: 1340-1348.
- Mesterhazy A. 1995. Types and components of resistance to Fusarium head blight of wheat. Plant Breed.. 114: 377- 386.
- Mohammadnia S, Asghari A, Motayagheni Y. 2015. Evalua-tion of end season drought tolerance in durum wheat advanced lines and its relation with RAPD marker. Inter-national Journal of Biology, Pharmacy and Allied Sciences. 4: 55-79.
- Nisbet R, Miner G, Yale K. Nisbet R., Miner G., Yale K., 2018. Chapter 8 - Advanced algorithms for data mining. editors. Handbook of statistical analysis and data mining applications (Second Edition). Academic Press. Boston, U.S.A: pp. 149-167.
- Pan H, Azimi M, Gui G, Yan F, Lin Z. Conte J., Astroza R., Benzoni G., Feltrin G., Loh K., Moaveni B., 2017. Vibration-based support vector machine for structural health monitoring. editors. Experimental vibration ana-lysis for civil structures. EVACES 2017. Lecture notes in civil engineering, Vol 5. Springer. Cham, Switzerland: pp. 167-178.
- Pan H, Azimi M, Yan F, Lin Z. 2018. Time-frequency-based data-driven structural diagnosis and damage detection for cable-stayed bridges. J. Bridge Eng.. 23: 1-22.
- Parry DW, Jenkinson P, McLeod L. 1995. Fusarium ear blight (Scab) in small grain cereals-A review. Plant Pathol.. 44: 207-238.
- Pinheiro N, Costa R, Almeida AS, Coutinho J. 2013. Durum wheat breeding in Mediterranean environments-influence of climatic variables on quality traits. Emir. J. Food Agric.. 25: 962-973.
- Qiu R, Yang C, Moghimi A, Anderson J, Steffenson B, Marchetto P. 2018. Detection of Fusarium head blight in small grains using hyperspectral imaging. National Fusarium Head Blight Forum: Saint Louis. MO, U.S.A: pp. 32-36.
- Rao HS, Basha OP, Singh NK, Sato K, Dhaliwal HS. 2007. Frequency distributions and composite interval mapping for QTL analysis in 'Steptoe' × 'Morex' barley mapping population. Barley Genet. Newsl.. 37: 5-20.
- Rudd JC, Horsley RD, McKendry AL, Elias EM. 2001. Host plant resistance genes for Fusarium head blight: Sources, mechanisms, and utility in conventional breeding systems. Crop Sci.. 41: 620-627.
- Siranidou E, Kang Z, Buchenauer H. 2002. Studies on symp-tom development, phenolic compounds and morphological defense responses in wheat cultivars differing in resist-ance to Fusarium head blight. J. Phytopathol.. 150: 200- 208.
- Stack R. 2000. Return of an old problem: Fusarium head blight of small grains. American Pathology Society. APSnet Feature.
- Stępień Ł, Chełkowski J. 2010. Fusarium head blight of wheat: Pathogenic species and their mycotoxins. World Mycotoxin J.. 3: 107-119.
- Sutton J. 1982. Epidemiology of wheat head blight and maize ear rot caused by Fusarium graminearum. Can. J. Plant Pathol.. 4: 195-209.
- Visconti A, Pascale M. 2010. An overview on Fusarium mycotoxins in the durum wheat pasta production chain. Cereal Chem.. 87: 21-27.
- Wang YZ, Yong XN, Xiao QP. 1982. The improvement of identification technique of scab resistance of wheat and the development of resistant sources. Sci. Agric. Sin.. 5: 67-77.
- Wegener M. 1992. Optimierung von saatgutpillierungen mit mikrobiellen antagonisten zur biologischen bekampfung von Fusarium culmorum in weizen. Diplomarbeit. Uni-versitat Gottingen Gottingen, Germany.
- Wiśniewska H, Stępień Ł, Waśkiewicz A, Beszterda M, Góral T, Belter J. 2014. Toxigenic Fusarium species infecting wheat heads in Poland. Cent. Eur. J. Biol.. 9: 163-172.
- Ze L, Li Z, Xianfang H, Jianlai W. 2017. Screening fungi-cides for controlling Fusarium head blight of winter wheat. Agricultural Science & Technology. 18: 2495-2502.