Abstract
In this study, we detected the morphological variation of 83 Perilla accessions from Korea and China by observing the measurements of seven quantitative and eight qualitative characters. Most accessions of cultivated var. frutescens from Southern China showed higher plant height than accessions from Korea and Northern China. The accessions of cultivated var. frutescens from Southern China have a relatively late flowering time, while there were three types of accessions for cultivated var. frutescens from Korea and Northern China: early-maturity, middle-maturity, and late-maturity. These results indicate that latitude with geographical distribution may be an essential factor affecting the flowering time in Perilla species in East Asia. In principal components analyses (PCAs), four quantitative traits and five qualitative characters contributed in the positive direction on the first axis. The accessions of cultivated var. frutescens were clearly separated from accessions of cultivated and weedy types of var. crispa in the PCAs, while the accessions of cultivated and weedy types of var. crispa revealed no separation. In addition, most accessions of weedy type of var. frutescens were not clearly separated with the accessions of cultivated and weedy types of var. crispa. In this study, we have provided the information regarding the morphological variation of accessions of two cultivated types of Perilla crop and their weedy types from Korea and China. The findings provide further insight into the morphological variations and differentiation of Perilla accessions with different geographical distributions in Korea and China.
-
Key words: Perilla frutescens, Cultivated and weedy types, Geographical distribution, Morphological traits, Principal components analysis
INTRODUCTION
Perilla frutescens Britt. is an annual crop from the self-fertilizing species Labiatae. This
Perilla crop includes two varieties based on their morphology and uses: One is
P. frutescens var.
frutescens, an oil crop or vegetable crop (Ren in Chinese and Deulkkae in Korean). The other is
P. frutescens var.
crispa, a Chinese medicine or vegetable crop (Zisu in Chinese and Cha-jo-ki in Korean) (
Makino 1961;
Nitta and Ohnishi 1999;
Lee and Ohnishi 2001,
2003;
Ma et al. 2019). These two
Perilla varieties are widely cultivated in the Himalayan hill areas and Asian regions, such as Southeast and East Asia (i.e., Nepal, India, Vietnam, Laos, China, Korea, and Japan) (
Makino 1961;
Lee and Ohnishi 2001,
2003;
Nitta et al. 2003;
Anjula and Bhatt 2008). In East Asia,
Perilla crop are cultivated on a large scale and used widely; therefore, East Asia is considered to be the birth place of
Perilla species (
Makino 1961;
Li 1969;
Nitta 2001;
Nitta et al. 2003). China has been assumed to be the primary center of biodiversity for the
Perilla crop (
Li 1969; Zeven and de Wet 1982;
Lee and Ohnishi 2003).
Lee and Ohnishi (2001, 2003) suggested that China was the original place for the cultivation of var.
frutescens and Korea was a secondary center of diffusion due to var.
frutescens’s large-scale cultivation, many uses, high morphological variation, and genetic diversity.
Today,
Perilla frutescens, in particular var.
frutescens, has become an essential economic crop in Korea. The var.
frutescens is extensively cultivated and used not only as an oil crop but also as a leafy vegetable. The leaves are rich in vitamin B and C, and they are a favored salad vegetable eaten with meat and used for pickles. Like sesame seeds, var.
frutescens seeds can be used for oil or seasoning, and even as flavoring agents in traditional Korean food. On the other hand, var.
crispa is not cultivated as much as var.
frutescens due to the decreased use of Chinese medicine. Var.
crispa is occasionally found as a relict form in Korea (
Lee and Ohnishi 2001;
Lee et al. 2002;
Sa et al. 2013), while
Perilla crop can be found in multiple provinces throughout China (i.e., Heilongjiang, Liaoning, Shanxi, Ningxia, Gansu, Anhui, Hubei, Sichuan, Taiwan, Hunan, Jiangxi, Jiangsu, Anhui, Henan, and Hulunbuir city of Inner Mongolia) (
Liu et al. 1996;
Liu and Zhang 1998;
Zhang et al. 2009;
Hu et al. 2010). The main cultivated areas for
P. frutescens var.
crispa are concentrated in Guizhou and Sichuan provinces (
Zhang et al. 2009). In the northern areas of China, the largest cultivated areas of
P. frutescens var.
frutescens are Changchun, Jilin, and Songyuan of Jilin province. In the Yanbian area of Northeast China, inhabited by Chinese-Koreans particularly, var.
frutescens is cultivated on a larger scale (
Ma and Lee 2017b).
These two cultivated types of
Perilla crop show differential morphological characteristics. Var.
frutescens is taller, has larger seeds (above 2 mm), green leaves and stem, non-wrinkled leaves, and a fragrance specific to the var.
frutescens, while var.
crispa is smaller in plant height and seed size (below 2 mm), has red or green coloration in the leaves and stem, wrinkly or non-wrinkly leaves, and a fragrance specific to the var.
crispa (
Lee and Ohnishi 2001;
Nitta 2001;
Nitta et al. 2003). Until now, many taxonomic studies have been carried out to distinguish the two cultivated types of the
Perilla species. Their chromosome numbers are the same 2n = 40 (
Yamane 1950;
Honda et al. 1994). A cross experiment between two varieties by
Nagai (1935) and
Honda et al. (1990, 1994) showed that they are cross-fertile with each other by means of artificial pollination. Further studies by
Koezuka et al. (1985, 1986),
Honda et al. (1990,
Lee and Ohnishi (2001) failed to distinguish the two varieties because of the existence of intermediate types. Essential oils, anthocyanines, leaf color, and seed hardness cannot be used as key characters when attempting to separate the two as unique types. It is still difficult to distinguish the two varieties. In East Asia, although a wild ancestor of the two cultivated types of
Perilla crop has not been certified, weedy plants of the
Perilla crop were reported by
Nitta and Ohnishi (1999),
Lee and Ohnishi (2001,
Lee et al. (2002),
Nitta et al. (2003), and
Sa et al. (2013). These weedy plants naturally grow in habitats such as roadsides, wastelands, open fields, and farms. In previous studies by
Nitta and Ohnishi (1999) and
Lee and Ohnishi (2003), RAPD and AFLP analyses showed that these weedy plants can be also grouped into two types; one belonging to the group of var.
frutescens and another belonging to the group of var.
crispa, respectively. However, concrete evidence for the origin of the weedy plants was not presented in their analyses. Therefore, there is a need to clarify the origin of the weedy plant for the
Perilla crop.
The investigation of variation in genetic resources of landrace crops in relation to their geographical distribution is a fundamental approach to understanding crop origin and genetic differentiation. Comparison of morphological characters expands our empirical understanding of differentiation and it is a popular step in taxonomic studies of plant species that begins the search for a common wild ancestor. It is necessary to investigate the genetic differentiation of the two cultivated types of Perilla crop (i.e., var. frutescens and var. crispa) in East Asia (Korea and China) in order to identify their common origin. Therefore, the objective of this study was to identify the morphological differences among cultivated and weedy types of Perilla accessions from Korea and China based on 15 morphological characters.
MATERIALS AND METHODS
Plant materials
The materials for this study consist of 83 accessions (47 cultivated var.
frutescens, 20 weedy var.
frutescens, 4 cultivated var.
crispa, and 12 weedy var.
crispa) collected in different areas of Korea and China. The number of accessions and the name of the collection places for
Perilla accessions collected in Korea and China are shown in
Table 1.
To assess the morphological variation among the accessions of cultivated and weedy types of Perilla crop from Korea and China, ten individuals of each accession were grown in a field at the College of Agriculture and Life Sciences, Kangwon National University, Chuncheon, Gangwon-do, in Korea. Approximately twenty seeds from each accession were sown in a nursery bed in early May and kept in a glass house for a month. Ten seedlings of each accession were then transplanted into a field in early June. We examined seven quantitative and eight qualitative characters that were selected on the basis of a previous report by
Lee and Ohnishi (2001) and evaluated at the appropriate growth stages as described in detail in
Table 2. Measurements of the seven quantitative characters and observation of eight qualitative characters were made on 10 individuals for each accession.
The measurements of each trait were compared among the accessions using one-way analysis of variance (ANOVA) followed by the Student-Newman-Keuls test for multiple comparison of means among the accessions of cultivated var. frutescens, weedy var. frutescens, cultivated var. crispa and weedy var. crispa collected from different areas of Korea and China. Principal component analyses (PCAs) were conducted to detect differences among Perilla accessions. A geographical comparison of the accessions of cultivated var. frutescens collected from Korea and China was carried out using univariate and multivariate analyses. The Microsoft Excel Statistical Analysis System Program was used for univariate and multivariate analyses.
RESULTS
Morphological variation among cultivated types of Perilla crop and their weedy types from Korea and China
Descriptive statistics for the seven quantitative traits (QN1-QN7) and morphological descriptions for the eight qualitative traits (QL1-QL7) following the accession of the cultivated and weedy types of var.
frutescens and var.
crispa are shown in
Table 3. Based on the survey of quantitative traits, the average plant height (QN1) was 108.6 cm (45.0-163.0 cm), 111.9 cm (90.4-126.4 cm), 99.1 cm (93.6-106.4 cm), and 106.2 cm (86.4-128.4 cm) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average days from the seedling stage to the flowering stage (QN2) was 112.8 days (78.0-146.0 days), 120.6 days (108.0-139.0 days), 116.0 days (110.0-122.0 days), and 115.6 days (98.0-130.0 days) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average effective number of branches (QN3) was 18.1 (3.5-24.0), 19.2 (12.8-26.4), 24.2 (21.2-28.0), and 19.9 (15.2-25.6) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average number of branches (QN4) was 25.6 (7.0-32.0), 28.4 (21.2-34.8), 30.9 (26.8-35.2), and 27.4 (21.0-32.0) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average number of internodes (QN5) was 13.8 (4.5-17.4), 15.1 (11.6-18.4), 16.3 (14.4-18.6), and 15.3 (12.6-17.8) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average length of the largest inflorescence (QN6) was 9.9 cm (4.7-23.2 cm), 9.4 cm (4.0-15.6 cm), 10.6 cm (8.6-11.8 cm), and 11.9 cm (4.8-16.8 cm) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively. The average floret number of the largest inflorescence (QN7) was 34.5 (20.8-60.0), 32.5 (20.0-47.2), 33.6 (23.2-37.6), and 36.3 (22.4-48.0) for cultivated var.
frutescens, weedy var.
frutescens, cultivated var.
crispa, and weedy var.
crispa, respectively.
Mean score analysis revealed that the accessions of cultivated var. frutescens and cultivated var. crispa were different at a statistically significant level (P < 0.05) when comparing the effective number of branches (QN3), the number of branches (QN4) and the number of internodes (QN5). Statistically significant differences existed for the effective number of branches (QN3) when comparing the accessions of cultivated var. crispa and cultivated and weedy types of var. frutescens, and when comparing cultivated var. crispa with weedy var. crispa. However, no differences were found between the cultivated and weedy types of var. frutescens or var. crispa when comparing plant height (QN1), flowering time (QN2), length of the largest inflorescence (QN6) and the number of florets of inflorescence (QN7). In addition, no differences existed between cultivated var. frutescens and cultivated var. crispa, or cultivated var. frutescens and weedy types of var. crispa when comparing the effective number of branches (QN3), the number of branches (QN4) and the number of internodes (QN5). The cultivated and weedy types of var. crispa revealed no significant difference in any trait observed in this study.
On the basis of the detection of qualitative traits, for seed size (QL1) and hardness (QL2), 46 accessions of cultivated var. frutescens and 2 accessions of weedy var. frutescens showed a large seed size (QL1), while one accession of cultivated var. frutescens, 18 accessions of weedy var. frutescens, and 16 accessions of cultivated and weedy types of var. crispa showed small sizes (QL1). Seed hardness (QL2) in the accessions of cultivated var. frutescnes was found to be both soft (44 accessions) and hard (3 accessions), while the accessions of weedy var. frutescens produced both soft (2 accessions) and hard (18 accessions) seeds, and all the accessions of cultivated and weedy types of var. crispa produced only hard seeds. Concerning seed color (QL3), the accessions of cultivated var. frutescens included white (6 accessions), gray (29 accessions), and brown (12 accessions) seeds. The accessions of weedy var. frutescens displayed gray (5 accessions), brown (12 accessions), and dark brown (3 accessions) colored seeds. The accessions of cultivated var. crispa were gray (1 accession) and brown (3 accessions), while the accessions of weedy var. crispa displayed gray (1 accession), brown (6 accessions), and dark brown (5 accessions) colored seeds. Flower colors were found on 44 cultivated accessions (QL4): 13 weedy accessions of var. frutescens, one cultivated accession of var. crispa and one weedy accession of var. crispa that were white; one cultivated accession and three weedy accessions of var. frutescens that were white/ purple; and two cultivated accessions and four weedy accessions of var. frutescens, three cultivated accessions and 11 weedy accessions of var. crispa that were purple. The stem color (QL5) on the accessions of cultivated var. frutescens included green (44 accessions) and green/purple (3 accessions) colors. The stem on the accessions of weedy var. frutescens had green (15 accessions) and green/purple (5 accessions) colors, while the cultivated accessions of var. crispa had green (1 accession), green/purple (2 accessions), and purple (1 accessions) colors. The stem color on weedy accessions of var. crispa had green/purple (11 accessions) and purple (1 accession) colors. For the leaf surface color (QL6), 30 accessions of cultivated var. frutescens, 16 accessions of weedy var. frutescens, and five accessions of weedy var. crispa had a green leaf surface color. 17 accessions of cultivated var. frutescens, two accessions of weedy var. frutescens, three accessions of cultivated var. crispa and four accessions of weedy var. crispa exhibited a deep green color on the leaf surface. Two accessions of weedy var. frutescens and three accessions of weedy var. crispa exhibited a light purple leaf surface color. Only one accession of cultivated var. crispa had a purple color on the leaf surface. For the color on the reverse side of the leaves (QL7), the accessions of cultivated var. frutescens had green (33 accessions), deep green (9 accessions), light purple (4 accessions), and purple (1 accessions) colors. The accessions of weedy var. frutescens had green (11 accessions), deep green (2 accessions), and purple (7 accessions) colors, while cultivated accessions of var. crispa had deep green (1 accession), light purple (1 accession), and purple (2 accessions) colors, while the accessions of weedy var. crispa had green (1 accession), light purple (2 accessions), and purple (9 accessions) colors. For the pubescent degree (QL8), the accessions of cultivated var. frutescens had slight pubescence (8 accessions), average pubescence (32 accessions), and heavy pubescence (7 accessions). The accessions of weedy var. frutescens had slightly pubescence (6 accessions), average pubescence (12 accessions), and heavily pubescence (2 accessions), while all accessions of cultivated var. crispa were slightly pubescent, and the accessions of weedy var. crispa displayed slightly pubescence (7 accessions) and average pubescence (5 accessions).
Geographical differences in cultivated var. frutescens between Korea and China
Statistical analysis was performed on the seven quantitative characters on the accessions of cultivated var.
frutescens collected from Korea and China in order to understand the geographical difference between the accessions of cultivated var.
frutescens from these regions. The mean, standard deviation, and range values of the seven quantitative traits are summarized in
Table 4.
As shown in
Table 4, QN1 was 110.1 cm (89.0-124.0 cm) and 107.2 cm (45.0-163.0 cm) for accessions of cultivated var.
frutescens from Korea and China, respectively. QN2 was 114.9 cm (101.0-128.0 cm) and 110.8 cm (78.0-146.0 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. QN3 was 20.4 cm (16.0-24.0 cm) and 15.9 cm (3.5-23.2 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. QN4 was 28.0 cm (23.6-32.0 cm) and 23.3 cm (7.0-29.6 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. QN5 was 15.1 cm (12.8-17.4 cm) and 12.7 cm (4.5-15.8 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. QN6 was 10.0 cm (8.0-14.0 cm) and 9.7 cm (4.7-23.2 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. QN7 was 34.6 cm (28.0-40.0 cm) and 34.4 cm (20.8-60.0 cm) in accessions of cultivated var.
frutescens from Korea and China, respectively. Results from mean score comparison are as follows: significant differences (
P < 0.05) were found between accessions of cultivated var.
frutescens from Korea and China for effective number of branches (QN3), number of branches (QN4), and number of internodes (QN5). However, no significant difference between accessions of cultivated var.
frutescens from Korea and China were found in plant height (QN1), flowering time (QN2), length of the largest inflorescence (QN6), or the number of florets of inflorescence (QN7).
In this study, principal components analysis (PCA) was used to detect the morphological variation among the
Perilla accessions from Korea and China. The first (principal component 1) and second (principal component 2) components accounted for 35.4% and 23.4% of the total variance, respectively (
Table 5). Four quantitative traits (QN2, QN3, QN4, and QN5) and five qualitative characters (QL1, QL2, QL4, QL5, and QL7) greatly contributed in the positive direction on the first axis (
Table 5). The other quantitative and qualitative characters contributed in both the positive and negative direction on the first axis.
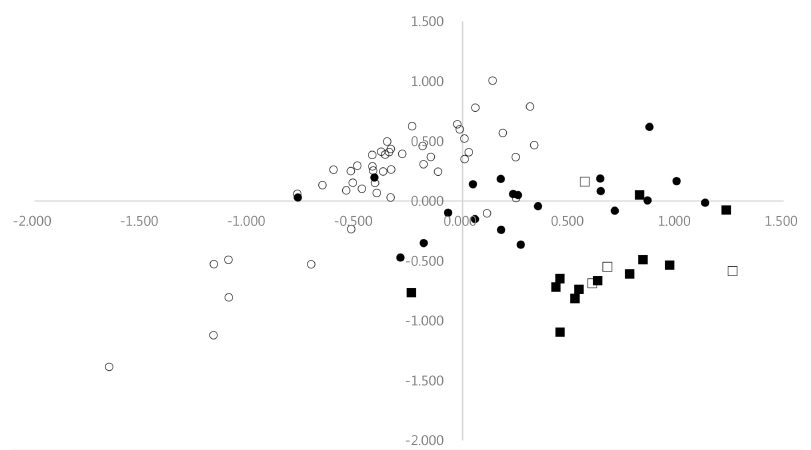
As shown in
Fig. 1, most accessions of cultivated var.
frutescens from Korea and China were clearly separated with the accessions of weedy type of var.
frutescens in the first axis, except for a few outliers. Most accessions of cultivated var.
frutescens from Korea and China were situated to the left on the first axis, and most accessions of weedy var.
frutescens from Korea and China were situated to the right on the first axis. In addition, most accessions of cultivated and weedy types of var.
crispa were situated to the right on the first axis, except for one. Most accessions from cultivated type of var.
frutescens clearly separated with the accessions of cultivated and weedy types of var.
crispa in the first axis. The accessions of cultivated and weedy types of var.
crispa were not clearly separated from one another by the first axis. Furthermore, the first axis did not separate most accessions of weedy type of var.
frutescens from the accessions of cultivated and weedy types of var.
crispa. However, the accessions of weedy type of var.
frutescens were comparatively more clearly separated from the accessions of cultivated and weedy types of var.
crispa by the second axis. Most accessions of weedy var.
frutescens were in the upper part of the plane, whereas most accessions of cultivated and weedy types of var.
crispa were in the lower part of the plane (
Fig. 1).
DISCUSSION
Morphological variation among cultivated and weedy types of Perilla crop from Korea and China
Morphological variation between cultivated and weedy or wild type in some crops within their geographic distribution areas has been of considerable interest in the study of the evolution and differentiation of crop. Domestication is an evolutionary process through which domesticated plants become morphologically and physiologically divergent from their wild ancestors (
Schwanitz 1966;
Gould and Johnstn 1972;
Wyatt and Antonovics 1981;
Harlan 1992). Therefore, this study was conducted to understand the morphological variation and differentiation process of two cultivated types of
Perilla crop and their weedy types from Korea and China.
In this study, seven quantitative characters and eight qualitative characters were examined for 83
Perilla accessions (47 cultivated var.
frutescens, 20 weedy var.
frutescens, 4 cultivated var.
crispa, and 12 weedy var.
crispa) from Korea and China. On the results for the analysis of seven quantitative characters, the four quantitative characters (QN1, QN2, QN6, QN7) did not show significant differences between accessions of cultivated and weedy types of
Perilla crop, while the accessions of cultivated var.
frutescens were significantly different from the accessions of weedy type of var.
frutescens and cultivated and weedy types of var.
crispa in two quantitative characters (QN4, QN5). The accessions of cultivated var.
crispa were significantly different from the accessions of cultivated and weedy types of var.
frutescens and weedy type of var.
crispa only in one quantitative character (QN3). In addition, the accessions of cultivated var.
frutescens did not show significant differences from the accessions of weedy var.
frutescens and var.
crispa. Furthermore, cultivated and weedy types of var.
crispa showed no significant difference in most of the investigated traits, except for QN3 (
Table 3). These results suggest that morphological variation is not significant in the accessions of cultivated and weedy types of
Perilla crop collected under similar climatic conditions in Korea and China. However, in the case of flowering time, the accessions collected in the southern and northern parts of China showed a significant variation. For example, accessions collected from Korea include middle-maturity (100 days < flowering time < 130 days) and northern part of China displayed early-maturity (flowering time < 100 days), middle-maturity (100 days < flowering time < 130 days), and late-maturity (flowering time > 130 days), while most accessions collected from southern China showed only late-maturity. These results indicate that latitude with geographical distribution might be an important factor affecting flowering time in
Perilla species, as reported in the previous study of
Lee and Ohnishi (2001) and
Ma et al. (2017b).
Based on the examination of qualitative traits in
Table 3, morphological differences between accessions of cultivated var.
frutescens and accessions of cultivated var.
crispa were revealed in seed size (QL1), seed hardness (QL2), color of stem (QL5), color of leaf (QL6, QL7), and pubescence degree (QL8). Except for several exceptional accessions, the differences between cultivated and weedy types of var.
frutescens were only shown in seed size (QL1) and seed hardness (QL2). Meanwhile, differences between accessions of cultivated and weedy types of var.
crispa showed no significant difference in most of the morphological traits we investigated. Many taxonomic studies have been carried out to distinguish the two cultivated types of
Perilla crop and their weedy types.
Lee and Ohnishi (2001) suggested that seed size was considered to be the most reliable trait in discriminating between cultivated var.
frutescens and cultivated var.
crispa or between cultivated and weedy types of var.
frutescens. Many researchers have failed to distinguish the two varieties of
Perilla crop because of the existence of intermediate types. However, seed size is identified to be a reliable character for the differentiation between accessions of cultivated var.
frutescens and cultivated var.
crispa, as previously reported of
Lee and Ohnishi (2001). In addition, discrimination between the cultivated and weedy types of var.
crispa is impossible by the morphological key characters (
Lee and Ohnishi 2001), by RAPD markers (
Nitta and Ohnishi 1999) and by AFLPs (
Lee and Ohnishi 2003). Therefore, cultivated var.
crispa might not be completely domesticated, and a wild-type has been used for cultivation in the case of Japan in East Asia.
With regard to the crop evolution process, seed dormancy is strongly selected against by farmers during domestication (
Hancock 1992). On the previous reports by
Lee and Ohnishi (2001),
Jung et al. (2009), and
Sa et al. (2012,
2018), cultivated and weedy types of var.
crispa have strong seed dormancy, but accessions of cultivated var.
frutescens type have weak or almost no dormancy. Thus, cultivated var.
frutescens may be regarded as a relatively highly domesticated type among the two cultivated types of
Perilla species (
Sa et al. 2012). In the PCA analysis (
Fig. 1), the accessions of weedy type of var.
frutescens and also cultivated and weedy types of var.
crispa were situated far away from the accessions of cultivated var.
frutescens, except for several exceptional accessions. However, no significant difference was found between cultivated and weedy types of var.
crispa in PCA analysis.
In our study, we have observed several Perilla accessions that showed intermediate types between cultivated var. frutescens and cultivated var. crispa. Thus, these accessions were very difficult to distinguish between var. frutescens and var. crispa by their morphological characteristics. For example, in the case of cultivated var. frutescens collected in China, some accessions produced plant fragrance such as cultivated var. frutescens, but the color of the leaves were purple (e.g., cultivated var. crispa). In addition, some accessions of weedy types of var. frutescens and var. crispa collected in China did not produce a fragrance, preventing us from making conclusions about fragrance in this study. However, most accessions of cultivated var. frutescens collected in Korea have shown characteristics of cultivated var. frutescens as described in the introduction. These results probably indicate that cultivated var. frutescens has been adapted under stronger selection pressure than var. crispa by Korean farmers. For instance, weedy var. frutescens cannot be eaten so they are removed when found by farmers in Korea. However, in China, cultivated var. frutescens is not used as much as in Korea so various plants of Perilla species still exist in their natural state.
Geographical variation of cultivated var. frutescens between Korea and China
Previous research by
Lee and Ohnishi (2001) found that the formation of morphological variation was closely related to the geographic origin of accessions of
Perilla species. For example, most accessions of cultivated var.
frutescens from China showed significant differences from accessions of cultivated var.
frutescens from Korea and Japan, such as in plant height, internode number, and flowering time (
Lee and Ohnishi 2001). And also,
Ma and Lee (2017a) reported that the accessions of cultivated var.
frutescens from high-latitude areas (Northeast China) and middle-latitude areas (Northwest China and North China) of China showed significant differences in quantitative traits, such as branch number, internode number, length of largest inflorescence, and flowering time etc. The accessions of cultivated var.
frutescens from Korea and China showed significant differences in several morphological traits, such as effective number of branches (QN3), branch number (QN4), and internode number (QN5) (
Table 4). However, unlike previous reports by
Lee and Ohnishi (2001) and
Ma and Lee (2017a,
b), there were no differences in plant height and flowering time in this study. Most accessions of cultivated var.
frutescens from Southern China showed higher plant height than those accessions from Korea and Northern China. These accessions of cultivated var.
frutescens from Southern China have a single late flowering time, while the accessions cultivated from var.
frutescens in Korea and Northern China have three types: early-maturity, middle-maturity, and late-maturity. These results indicate that latitude with geographical distribution may be an important factor affecting the flowering time in
Perilla crop, and a contributing factor to why
Perilla is considered a short-day crop. The geographic differentiation of cultivated var.
frutescens in East Asia may be the result of the diffusion of cultivation of
Perilla crop and local selection for flowering habit by farmers using traditional agricultural practices over long periods. The findings demonstrated in this study provide researchers furtherer understanding of the morphological variations and differentiation of the two cultivated types of
Perilla crop and their weedy types in relation to their geographical distributions in Korea and China.
ACKNOWLEDGEMENTS
This study was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (2016R1D1A1B01006461), and the Cooperative Research Program for Agriculture Science and Technology Development (Project nos. PJ01422 7032019 and PJ0142272019), Rural Development Ad ministration, Republic of Korea.
Fig. 1Projection of 83 accessions of the two cultivated types of Perilla crop and their weedy types collected from Korea and China in the first and second principal components (○: Cultivated type of var. frutescens; ●: Weedy type of var. frutescens; □: Cultivated type of var. crispa; ■: Weedy type of var. crispa).
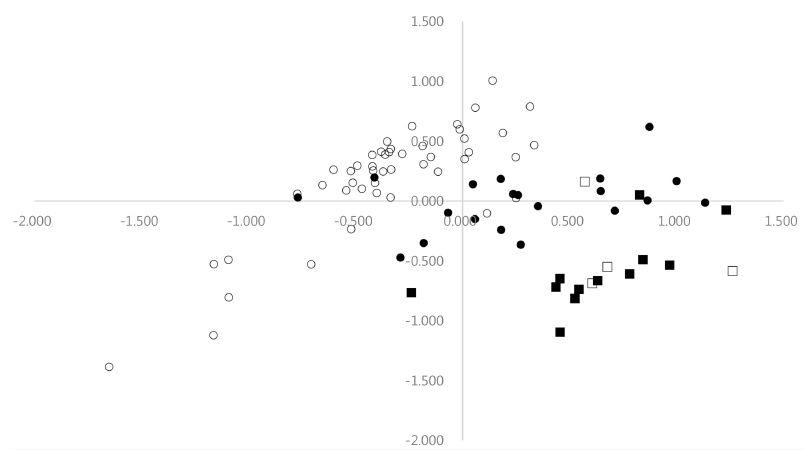
Table 1Accessions of two cultivated types of Perilla crop and their weedy types used for morphologic analysis.
Table 1
|
Code No. |
Accession no. |
Village, town or city |
Country |
Type |
|
1 |
PF08016 |
Donghae-si, Kangwon-do |
Korea |
Cultivated type of var. frutescens
|
|
2 |
PF08118 |
Gangneung-si, Kangwon-do |
Korea |
Cultivated type of var. frutescens
|
|
3 |
PF11098 |
Yanggu-gun, Kangwon-do |
Korea |
Cultivated type of var. frutescens
|
|
4 |
PF15009 |
Pyeongchang-gun, Kangwon-do |
Korea |
Cultivated type of var. frutescens
|
|
5 |
PF06264 |
Pocheon-si, Gyeonggi-do |
Korea |
Cultivated type of var. frutescens
|
|
6 |
PF06239 |
Gapyeong-gun, Gyeonggi-do |
Korea |
Cultivated type of var. frutescens
|
|
7 |
PF98052 |
Hamyang-gun, Gyeongsangnam-do |
Korea |
Cultivated type of var. frutescens
|
|
8 |
PF98062 |
Yangsan-si, Gyeongsangnam-do |
Korea |
Cultivated type of var. frutescens
|
|
9 |
PF13054 |
Yeongyang-gun, Gyeongsangbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
10 |
PF14022 |
Mungyeong-si, Gyeongsangbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
11 |
PF14026 |
Uiseong-gun, Gyeongsangbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
12 |
PF17045 |
Damyang-gun, Jeollanam-do |
Korea |
Cultivated type of var. frutescens
|
|
13 |
PF98070 |
Goheung-gun, Jeollanam-do |
Korea |
Cultivated type of var. frutescens
|
|
14 |
PF98073 |
Gangjin-gun, Jeollanam-do |
Korea |
Cultivated type of var. frutescens
|
|
15 |
PF13137 |
Iksan-si, Jeollabuk-do |
Korea |
Cultivated type of var. frutescens
|
|
16 |
PF13168 |
Jeongeup-si, Jeollabuk-do |
Korea |
Cultivated type of var. frutescens
|
|
17 |
PF13190 |
Jinan-gun, Jeollabuk-do |
Korea |
Cultivated type of var. frutescens
|
|
18 |
PF12071 |
Seogwipo-si, Jeju-do |
Korea |
Cultivated type of var. frutescens
|
|
19 |
PF12080 |
Jeju-si, Jeju-do |
Korea |
Cultivated type of var. frutescens
|
|
20 |
PF16030 |
Hongseong-gun, Chungcheongnam-do |
Korea |
Cultivated type of var. frutescens
|
|
21 |
PF09072 |
Jecheon-si, Chungcheongbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
22 |
PF12105 |
Chungju-si, Chungcheongbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
23 |
PF12123 |
Jeungpyeong-gun, Chungcheongbuk-do |
Korea |
Cultivated type of var. frutescens
|
|
24 |
PF14001 |
Wonju-si, Kangwon-do |
Korea |
Weedy type of var. frutescens
|
|
25 |
PF08083 |
Yangpyeong-gun, Gyeonggi-do |
Korea |
Weedy type of var. frutescens
|
|
26 |
PF09140 |
Yeoju-si, Gyeonggi-do |
Korea |
Weedy type of var. frutescens
|
|
27 |
PF08096 |
Hadong-gun, Gyeongsangnam-do |
Korea |
Weedy type of var. frutescens
|
|
28 |
PF13073 |
Cheongsong-gun, Gyeongsangbuk-do |
Korea |
Weedy type of var. frutescens
|
|
29 |
PF17065 |
Jangseong-gun, Jeollanam-do |
Korea |
Weedy type of var. frutescens
|
|
30 |
PF13189 |
Jinan-gun, Jeollabuk-do |
Korea |
Weedy type of var. frutescens
|
|
31 |
PF12072 |
Seogwipo-si, Jeju-do |
Korea |
Weedy type of var. frutescens
|
|
32 |
PF12081 |
Jeju-si, Jeju-do |
Korea |
Weedy type of var. frutescens
|
|
33 |
PF15103 |
Dangjin-si, Chungcheongnam-do |
Korea |
Weedy type of var. frutescens
|
|
34 |
PF15082 |
Cheonan-si, Chungcheongnam-do |
Korea |
Weedy type of var. frutescens
|
|
35 |
PF12149 |
Okcheon-gun, Chungcheongbuk-do |
Korea |
Weedy type of var. frutescens
|
|
36 |
PF15020 |
Yeongwol-gun, Kangwon-do |
Korea |
Weedy type of var. crispa
|
|
37 |
PF08082 |
Yangpyeong-gun, Gyeonggi-do |
Korea |
Weedy type of var. crispa
|
|
38 |
PF09139 |
Yeoju-si, Gyeonggi-do |
Korea |
Weedy type of var. crispa
|
|
39 |
PF13077 |
Pohang-si, Gyeongsangbuk-do |
Korea |
Weedy type of var. crispa
|
|
40 |
PF08097 |
Gwangyang-si, Jeollanam-do |
Korea |
Weedy type of var. crispa
|
|
41 |
PF13171 |
Jeongeup-si, Jeollabuk-do |
Korea |
Weedy type of var. crispa
|
|
42 |
PF12099 |
Jeju-si, Jeju-do |
Korea |
Weedy type of var. crispa
|
|
43 |
PF15106 |
Seosan-si, Chungcheongnam-do |
Korea |
Weedy type of var. crispa
|
|
44 |
PF09114 |
Goesan-gun, Chungcheongbuk-do |
Korea |
Weedy type of var. crispa
|
|
45 |
CH10 |
Anguo, Hebei |
China |
Cultivated type of var. frutescens
|
|
46 |
CH14 |
Siping, Jilin |
China |
Cultivated type of var. frutescens
|
|
47 |
CH24 |
Baishan, Jilin |
China |
Cultivated type of var. frutescens
|
|
48 |
CH31 |
Huai'an, Jiangsu |
China |
Cultivated type of var. frutescens
|
|
49 |
CH37 |
Tianshui, Gansu |
China |
Cultivated type of var. frutescens
|
|
50 |
CH40 |
Longnan, Gansu |
China |
Cultivated type of var. frutescens
|
|
51 |
CH41 |
Haozhou, Anhui |
China |
Cultivated type of var. frutescens
|
|
52 |
CH46 |
Heihe, Hei Longjiang |
China |
Cultivated type of var. frutescens
|
|
53 |
CH47 |
Baoding, Hebei |
China |
Cultivated type of var. frutescens
|
|
54 |
CH49 |
Pingliang, Gansu |
China |
Cultivated type of var. frutescens
|
|
55 |
CH5 |
Shenyang, Liaoning |
China |
Cultivated type of var. frutescens
|
|
56 |
CH51 |
Qingyang, Gansu |
China |
Cultivated type of var. frutescens
|
|
57 |
CH56 |
Helong, Jilin |
China |
Cultivated type of var. frutescens
|
|
58 |
CH67 |
Helong, Jilin |
China |
Cultivated type of var. frutescens
|
|
59 |
CH7 |
Liaoyang, Liaoning |
China |
Cultivated type of var. frutescens
|
|
60 |
CH71 |
Yanji, Jilin |
China |
Cultivated type of var. frutescens
|
|
61 |
CH81 |
Longjing, Jilin |
China |
Cultivated type of var. frutescens
|
|
62 |
CH82 |
Longjing, Jilin |
China |
Cultivated type of var. frutescens
|
|
63 |
CH9 |
Cangzhou, Hebei |
China |
Cultivated type of var. frutescens
|
|
64 |
CSY18 |
Dali,Yangbi, Huan'an cun |
China |
Cultivated type of var. frutescens
|
|
65 |
CSY22 |
Dali,Eryuan, Fengyuzhen |
China |
Cultivated type of var. frutescens
|
|
66 |
CSY31 |
Lijiang, Baishazhen |
China |
Cultivated type of var. frutescens
|
|
67 |
CSY37 |
Lijiang, Yongshengxian |
China |
Cultivated type of var. frutescens
|
|
68 |
CSY4 |
Chuxiong, Cangling |
China |
Cultivated type of var. frutescens
|
|
69 |
CSY1 |
An'ning-si, Qinglong |
China |
Weedy type of var. frutescens
|
|
70 |
CSY2 |
Chuxiong, Jiuzhuang |
China |
Weedy type of var. frutescens
|
|
71 |
CSY20 |
Dali,WanqiaoZhen, Zhongzhuang cun |
China |
Weedy type of var. frutescens
|
|
72 |
CSY24 |
Dali,Eryuan, Niujiexiang |
China |
Weedy type of var. frutescens
|
|
73 |
CSY32 |
Lijiang, Baishazhen |
China |
Weedy type of var. frutescens
|
|
74 |
CSY41 |
Lijiang, Yongshengxian, Qinazhen |
China |
Weedy type of var. frutescens
|
|
75 |
CSY45 |
Dali,Binchuanxian, Daluocheng |
China |
Weedy type of var. frutescens
|
|
76 |
CSY5 |
Dali,Yangbi, Pingpo zhen |
China |
Weedy type of var. frutescens
|
|
77 |
CH2 |
Harbin, Hei Longjiang |
China |
Cultivated type of var. crispa
|
|
78 |
CH52 |
Yantai, Shandong |
China |
Cultivated type of var. crispa
|
|
79 |
CH53 |
Zhaoyuan, Shandong |
China |
Cultivated type of var. crispa
|
|
80 |
CH54 |
Zhaoyuan, Shandong |
China |
Cultivated type of var. crispa
|
|
81 |
CSY23 |
Dali, Eryuan |
China |
Weedy type of var. crispa
|
|
82 |
CSY48 |
Jinhua, Zhejiang |
China |
Weedy type of var. crispa
|
|
83 |
CSY7 |
Dali, Yangbi, Pingpo zhen |
China |
Weedy type of var. crispa
|
Table 2Characters used in the morphological analysis of Perilla crop and their weedy types.
Table 2
|
Abbreviation |
Character |
When/how measured |
Category |
|
QN1 |
Plant height |
at flowering stage |
cm |
|
QN2 |
Days from seedling to flowering |
at flowering stage |
day |
|
QN3 |
Effective number of branches |
at flowering stage |
number |
|
QN4 |
Number of branches |
at flowering stage |
number |
|
QN5 |
Number of internodes |
at flowering stage |
number |
|
QN6 |
Length of the largest inflorescence at flowering stage |
at flowering stage |
cm |
|
QN7 |
Number of floret of the inflorescence at flowering stage |
at flowering stage |
number |
|
QL1 |
seed size |
after harvest |
(> 2 mm)-1, (2 < mm)-2 |
|
QL2 |
seed hardness |
after harvest |
soft-1, hard-2 |
|
QL3 |
seed color |
after harvest |
white-1, gray-2, brown-3, dark brown-4 |
|
QL4 |
flower color |
at flowering stage |
white-1, white/purple-2, purple-3 |
|
QL5 |
stem color |
at flowering stage |
green-1, green/purple-2, purple-3 |
|
QL6 |
Color of leaf surface |
at flowering stage |
green-1, deep green-2, light purple-3, purple-4 |
|
QL7 |
Color of reverse side of leaf |
at flowering stage |
green-1, deep green-2, light purple-3, purple-4 |
|
QL8 |
degree of pubescence |
at flowering stage |
slightly pubescence-1, average pubescence-2, heavily pubescence-3 |
Table 3Mean, standard deviation, range and accession number for 7 quantitative and 8 qualitative characters among 83 accessions of the two cultivated types of Perilla crop and their weedy types.
Table 3
|
Morphological character |
Cultivated var. frutescens (N=47) |
Weedy var. frutescens (N=20) |
Cultivated var. crispa (N=4) |
Weedy var. crispa (N=12) |
Statistical comparison*
|
|
QN1 |
108.6 ± 18.3 (45.0-163.0) |
111.9 ± 11.8 (90.4-126.4) |
99.1 ± 5.4 (93.6-106.4) |
106.2 ± 13.8 (86.4-128.4) |
ns |
|
QN2 |
112.8 ± 12.9 (78.0-146.0) |
120.6 ± 10.0 (108.0-139.0) |
116.0 ± 4.9 (110.0-122.0) |
115.6 ± 10.1 (98.0-130.0) |
ns |
|
QN3 |
18.1 ± 4.1 (3.5-24.0) |
19.2 ± 3.4 (12.8-26.4) |
24.2 ± 2.9 (21.2-28.0) |
19.9 ± 3.2 (15.2-25.6) |
A = B = D ≠ C |
|
QN4 |
25.6 ± 4.7 (7.0-32.0) |
28.4 ± 3.7 (21.2-34.8) |
30.9 ± 3.6 (26.8-35.2) |
27.4 ± 3.5 (21.0-32.0) |
A ≠ C, A = B = D, B = C = D |
|
QN5 |
13.8 ± 2.3 (4.5-17.4) |
15.1 ± 1.8 (11.6-18.4) |
16.3 ± 1.7 (14.4-18.6) |
15.3 ± 1.6 (12.6-17.8) |
A ≠ C, A = B = D, B = C = D |
|
QN6 |
9.9 ± 2.9 (4.7-23.2) |
9.4 ± 3.3 (4.0-15.6) |
10.6 ± 1.4 (8.6-11.8) |
11.9 ± 3.4 (4.8-16.8) |
ns |
|
QN7 |
34.5 ± 6.5 (20.8-60.0) |
32.5 ± 6.8 (20-47.2) |
33.6 ± 7.0 (23.2-37.6) |
36.3 ± 7.0 (22.4-48.0) |
ns |
|
QL1 |
large (46), small (1) |
large (2), small (18) |
small (4) |
small (12) |
|
QL2 |
soft (44), hard (3) |
soft (2), hard (18) |
hard (4) |
hard (12) |
|
QL3 |
white (6), gray (29), Brown (12) |
gray (5), brown (12), dark brown (3) |
gray (1), brown (3) |
gray (1), brown (6), dark brown (5) |
|
QL4 |
white (44), white/purple (1), purple (2) |
white (13), white/purple (3), purple (4) |
white (1), purple (3) |
white (1), purple (11) |
|
QL5 |
green (44), green/purple (3) |
green (15), green/purple (5) |
green (1), green/purple (2), Purple (1) |
green/purple (11), purple (1) |
|
QL6 |
green (30), deep green (17) |
green (16), deep green (2), light purple (2) |
deep green (3), purple (1) |
green (5), deep green (4), light purple (3) |
|
QL7 |
green (33), deep green (9), light purple (4), purple (1) |
green (11), deep green (2), purple (7) |
deep green (1), light purple (1), purple (2) |
green (1), light purple (2), purple (9) |
|
QL8 |
slightly pubescence (8), pubescence (32), heavily pubescence (7) |
slightly pubescence (6), pubescence (12), heavily pubescence (2) |
slightly pubescence (4) |
slightly pubescence (7), pubescence (5) |
Table 4Mean, standard deviation, range and accession number for 7 quantitative and 8 qualitative characters among 47 cultivated type of var. frutescens accessions from Korea and China.
Table 4
|
Morphological characters |
Cultivated var. frutescens in Korea (N = 23) |
Cultivated var. frutescens in China (N = 24) |
Statistical Comparison*
|
|
QN1 |
110.1 ± 10.0 (89.0-124.0) |
107.2 ± 23.9 (45.0-163.0) |
ns |
|
QN2 |
114.9 ± 6.2 (101.0-128.0) |
110.8 ± 17.0 (78.0-146.0) |
ns |
|
QN3 |
20.4 ± 2.4 (16.0-24.0) |
15.9 ± 4.2 (3.5-23.2) |
A ≠ B |
|
QN4 |
28.0 ± 2.5 (23.6-32.0) |
23.3 ± 5.1 (7.0-29.6) |
A ≠ B |
|
QN5 |
15.1 ± 1.3 (12.8-17.4) |
12.7 ± 2.6 (4.5-15.8) |
A ≠ B |
|
QN6 |
10.0 ± 1.5 (8.0-14.0) |
9.7 ± 3.9 (4.7-23.2) |
ns |
|
QN7 |
34.6 ± 3.1 (28.0-40.0) |
34.4 ± 8.7 (20.8-60.0) |
ns |
|
QL1 |
large (23) |
large (23), small (1) |
|
QL2 |
soft (21), hard (2) |
soft (23), hard (1) |
|
QL3 |
white (4), gray (12), brown (7) |
white (2), gray (17), brown (5) |
|
QL4 |
white (23) |
white (21), white/purple (1), purple (2) |
|
QL5 |
green (23) |
green (21), green/purple (3) |
|
QL6 |
green (18), deep green (5) |
green (12), deep green (12) |
|
QL7 |
green (20), deep green (2), light purple (1) |
green (13), deep green (7), light purple (3), purple (1) |
|
QL8 |
slightly pubescence (1), pubescence (18), heavily pubescence (4) |
slightly pubescence (7), pubescence (14), heavily pubescence (3) |
Table 5Cumulative variances of first and second principal components and the loadings of 15 qualitative and quantitative characters on each principal component.
Table 5
|
Morphological character |
Eigenvectors |
|
|
1 |
2 |
|
Number of floret of inflorescense (QN7) |
‒0.382 |
‒0.527 |
|
Length of largest inflorescense (QN6) |
‒0.241 |
‒0.597 |
|
Degree of pubescence (QL8) |
‒0.035 |
0.638 |
|
Seed color (QL3) |
0.394 |
‒0.467 |
|
Color of leaf surface (QL6) |
0.409 |
‒0.266 |
|
Plant height (QN1) |
0.425 |
0.586 |
|
Flowering time (QN2) |
0.634 |
0.49 |
|
Stem color (QL5) |
0.654 |
‒0.502 |
|
Flower color (QL4) |
0.687 |
‒0.499 |
|
Color of reverse side of leaf (QL7) |
0.717 |
‒0.416 |
|
Seed hardness (QL2) |
0.726 |
‒0.396 |
|
Number of branches (QN4) |
0.731 |
0.448 |
|
Seed size (QL1) |
0.736 |
‒0.438 |
|
Effective number of branches (QN3) |
0.75 |
0.438 |
|
Number of internodes (QN5) |
0.784 |
0.425 |
|
Cumulative variance (%) |
35.4 |
23.4 |
References
Reference
- Anjula P, Bhatt KC. 2008. Diversity distribution and collection of genetic resources of cultivated and weedy type in Perilla frutescens (L.) Britton var. frutescens and their uses in Indian Himalaya. Genet. Resour. Crop Evol.. 55: 883-892.
- Gould SJ, Johnston RF. 1972. Geographic variation. Ann. Rev. Ecol. Syst.. 3: 457-498.
- Hancock JF. 1992. Plant evolution and the origin of crop species. Prentice Hall, Englewood Cliffs. New Jersey, U.S.A..
- Harlan JR. Chapman GP., 1992. Origins and processes of domestication. Grass Evolution and Domestication, Cambridge University Press. Cambridge, U.K.: pp. 159-175.
- Honda G, Koezuka Y, Tabata M. 1990. Genetic studies of fruit color and hardness in Perilla frutescens
. Jpn. J. Breed.. 40: 469-474.
- Honda G, Yuba A, Kojima T, Tabata M. 1994. Chemotaxonomic and cytogenetic studies on Perilla frutescens var. citiodora ("Lemon Egoma"). Natural Medicines. 48: 185-190.
- Hu Y, Sun LW, Neo MC, Zhang YX, Wen CX, Xie XL, et al. 2010. Primary identifications and palynological observations of Perilla L. in China. J. Syst. Evol.. 48: 133-145.
- Jung JN, Yu CY, Kim JH, Lee JK. 2009. Variation of seed germination among cultivated and weedy types of Perilla crop in Korea and Japan. J. Crop Sci.. 54: 270-278.
- Koezuka Y, Honda G, Tabata M. 1986. Genetic control of the chemical composition of volatile oils in Perilla frutescens
. Phytochemistry. 26: 859-863.
- Koezuka Y, Honda G, Sakamoto S, Tabata M. 1985. Genetic control of anthocyanin production on Perilla frutescens
. Shoyakugaku Zasshi. 39: 228-231.
- Lee JK, Nitta M, Kim NS, Park CH, Yoon KM, Shin YB, et al. 2002. Genetic diversity of Perilla and related weedy types in Korea determined by AFLP analyses. Crop Sci.. 42: 2161-2166.
- Lee JK, Ohnishi O. 2001. Geographical differentiation of morphological characters among Perilla crops and their weedy types in. East Asia. Breed. Sci.. 51: 247-255.
- Lee JK, Ohnishi O. 2003. Genetic relationships among cultivated types of Perilla frutescens and their weedy types in East Asia revealed by AFLP markers. Genet. Resour. Crop Evol.. 50: 65-74.
- Li HL. 1969. The vegetables of ancient. China. Econ. Bot.. 23: 235-260.
- Liu YX, Zhang WM. 1998. Classification and resources distribution of Perilla
. Chin. Wild Plant Resour.. 17: 1-4.
- Liu YX, Zhang WM, Qian XS. 1996. Research and utilization of Perilla
. Wild Plant Resour.. 3: 24-27.
- Ma SJ, Lee JK. 2017a. Morphological variation of two cultivated types of Perilla crop from different areas of. China. Hortic. Sci. Technol.. 35: 510-522.
- Ma SJ, Lee JK. 2017b. Morphological variation of Perilla crop and their weedy types from northern and southern areas of China. Genet. Mol. Res.. 16: gmr16039853.
- Ma SJ, Sa KJ, Hong TK, Lee JK. 2019. Genetic diversity and population structure analysis in Perilla crop and their weedy types from northern and southern areas of China based on simple sequence repeat (SSRs). Genes Genom.. 41: 267-281.
- Makino T, Hara H, Tuyama T, Fumio M. 1961. Makino's new illustrated flora of Japan. Hokuryukan. Tokyo, Japan: pp. 1060.
- Nagai I. 1935. On "Shiso" and "Egoma" (in Japanese). Agriculture and Horticulture. 10: 2265-2273.
- Nitta M, Lee JK, Ohnishi O. 2003. Asian Perilla crops and their weedy forms: their cultivation, utilization and genetic relationships. Econ. Bot.. 57: 245-253.
- Nitta M, Ohnishi O. 1999. Genetic relationships among two Perilla crops, shiso and egoma, and the weedy type revealed by RAPD markers. Genes Genet. Syst.. 74: 43-48.
- Nitta M. 2001. Origin Perilla crops and their weedy type. Kyoto University. Kyoto, Japan, 78. Thesis.
- Sa KJ, Choi SH, Ueno M, Park KC, Park YJ, Ma KH, et al. 2013. Identification of genetic variations of cultivated and weedy types of Perilla species in Korea and Japan using morphological and SSR markers. Genes Genom.. 35: 649-659.
- Sa KJ, Hong TK, Park DH, Lee JK. 2018. Variation in seed germination according to the storage period after harvest in the accessions of Perilla species. Korean J. Crop Sci.. 63: 48-56.
- Sa KJ, Kim JA, Lee JK. 2012. Comparison of seed characteristics between the cultivated and the weedy types of Perilla species. Hort. Environ. Biote.. 53: 310-315.
- Schwanitz F. 1966. The origin of cultivated plants. Harvard University Press. Cambridge, Massachusetts, U.S.A.: pp. 175.
- Wyatt R, Antonovics J. 1981. Butterfly weed re-revisited: Spatial and temporal patterns of leaf shape variation in Asclepias tuberosa
. Evolution. 35: 529-542.
- Yamane Y. 1950. Cytogenetic studies in Perilla and coleus. I. Chromosome numbers (in Japanese). Jpn. Genet.. 25: 220.
- Zeven AC, Wet JMJ. 1982. Dictionary of cultivated plants and their regions of diversity. Centre for Agricultural Publishing and Documentation. Wageningen, the Netherlands.
- Zhang X, Wu W, Zheng YL, Chen L, Qianrong C. 2009. Essential oil variations in different Perilla L. accessions: chemotaxonomic implications. Plant Syst. Evol.. 281: 1-10.