Abstract
Developing elite hybrid rice varieties is one important objective of rice breeding programs. Several genes related to male sterilities, restores, and pollinators have been identified through map-based gene cloning within natural variations of rice. These identified genes are good targets for introducing genetic traits in molecular breeding. This study was conducted to breed elite hybrid lines with major genes related to hybrid traits and disease/insect resistance in 240 genetic resources and F1 hybrid combinations of rice. Molecular markers were reset for three major hybrid genes (S5, Rf3, Rf4) and thirteen disease/insect resistant genes (rice bacterial blight resistance genes Xa3, Xa4, xa5, Xa7, xa13, Xa21; blast resistance genes Pita, Pib, Pi5, Pii; brown planthopper resistant genes Bph18(t) and tungro virus resistance gene tsv1). Genotypes were then analyzed using molecular marker-assisted selection (MAS). Biological assay was then performed at the Red River Delta region in Vietnam using eleven F1 hybrid combinations and two control vatieties. Results showed that nine F1 hybrid combinations were highly resistant to rice bacterial blight and blast. Finally, eight F1 hybrid rice varieties with resistance to disease/insect were selected from eleven F1 hybrid combinations. Their characteristics such as agricultural traits and yields were then investigated. These F1 hybrid rice varieties developed with major genes related to hybrid traits and disease/insect resistant genes could be useful for hybrid breeding programs to achieve high yield with biotic and abiotic resistance.
-
Key words: Hybrid rice, Restorer line, Marker-assisted selection, Disease resistance, Insect resistance
INTRODUCTION
Rice (
Oryza sativa L.) is one of the most important crops in the world. It has been estimated that more than 414 million metric tons (MT) of milled rice are consumed in developing countries in 2018 (
Ikeda-Kawakatsu et al. 2012). According to USDA’s Economic Research Service (USDA-ERS), rice consumption in Africa is expected to reach 3,500 tons by 2028. Thus, it is essential to enhance rice productivity (
Thome et al. 2018). Most heterosis researches of F
1 hybrid rice have been focused on yield increase or related genes. Hybrid rice area is expanding in major rice-producting countries in Asia. Most of hybrid rice varieties have high yield and resistance to disease and insect such as bacterial blight, blast, and brown planthopper (
Viet 2008;
Dyah et al. 2013). Since 1970s, China has been conducting hybrid rice research, commercialization, and cultivation in 18 million hectares, comprising more than fifty percent of total national rice area. Meanwhile, hybrid rice varieties have yield increases of more than 30% compared to their parental inbred lines in Vietnam (
FAORAP 2014).
Hybrid rice can be produced using either a three-line or a two-line system. The former is derived from cytoplasmic male sterility (CMS) while the latter is derived from genic male sterility (GMS). Well-known GMS materials are photo-period-sensitive genic male sterile rice (PGMS) and temperature-sensitive genic male sterile rice (TGMS). The female parent of three-line hybrid rice is a GMS line and the male parent is a restorer line (
Li et al. 2007;
Lee et al. 2011;
Jo and Kim 2016). To breed various CGMS and restorer lines with backgrounds of Korean
japonica rice varieties, CGMS line BT-CMS was crossed with restorer line AR-3 (
Seo and Song 1993). In CMS line, one or more nuclear genes known as restorer-of-fertility (
Rf) genes can suppress the expression of aberrant mitochondrial CMS genes and restore viable pollen production (
Chase 2007). The CMS/Rf system has been an indispensable resource for commercial hybrid seed production (
Lin and Yuan 1980;
Chen and Liu 2014;
Bohra et al. 2016).
Jing et al. (2001) have found that
Rf4 locus in IR24 is flanked by RM171 (OSR33) and RM228 on the long arm of chromosome 10.
Rf3 is mapped on chromosome 1 and linked to RM1 about 1.9 cM (
Zhang et al. 1996;
He et al. 2002;
Balaji et al. 2012). Wide compatibility gene
S5 is known as one of the loci that can enhance wide compatibility during crossing made between
indica and
japonica lines. When sequences of indica (
S5i) and japonica (
S5j) are compared, two SNPs are identified in the coding region located at 1,010 bp [C/A] and 1,604 bp [C/T] downstream of the start codon (
Ouyang et al. 2009;
Sundaram et al. 2010).
Crop yield losses are caused by diseases and insect pests of tropical and temperate rice cultivation area in Asia, including bacterial blight (50–80%), blast (50–85%), tungro virus (5–10%), and brown planthopper (60%) (
Park et al. 2011;
Shin et al. 2011;
Fujita et al. 2013). About thirty major bacterial blight resistance genes have been identified in rice of many countries. Bacterial blight
R-genes are distributed across 9 of 12 chromosomes in rice (none on chromosome 1, 9, or 10). More than eight bacterial blight
R-genes are intensively clustered on chromosome 11 (
Jin et al. 2007;
Cheema et al. 2008;
Xia et al. 2012;
Zhang et al. 2015). Approximately 350 quantitative trait loci (QTLs) for resistance to rice blast have been identified and 23 of them have been molecularly characterized (
Yang et al. 2009;
Liu et al. 2010). Tungro virus resistance in Ultri Merah has been found to be under the control of a recessive gene (
tsv1) (
Lee et al. 2010). The gene was mapped on chromosome 7 where a gene encoding initiation factor (
eIF4G) was strongly associated with tungro virus resistance (
Lee et al. 2010). Rice brown planthopper resistance gene
Bph18(t) was identified by SSR marker RM463 and STR marker S15552 in an indica introgression line (
Myint et al. 2012). It expresses strong resistance to brown planthopper biotype of Korea (
Myint et al. 2012).
The objective of this study was to identify F1 hybrid rice with hybrid traits and genes related to disease/insect resistance using MAS and phenotypic selection for improving grain yield of F1 hybrid rice and developing excellent variety of hybrid rice for export.
MATERIALS AND METHODS
Plan meterials
We used a rice panel comprised of 240 genetic resources and F
1 hybrid combinations (
Table 1) provided by National Institute of Crop Science, Rural Development Administration (RDA), Republic of Korea. F
1 hybrid combinations were selected using MAS. They were applied for screening disease/insect resistance in the rice field of the Red River Delta region in Vietnam.
Total genomic DNA was extracted from fresh leaves of two-week-old rice seedlings using TissueLyser II system (QIAGEN, UK) with modified Cetyl Trimethyl Ammonium Bromide (CTAB) method as described previously by Cho et. al. (2007). DNA concentration was quantified using a spectrophotometer (NanoDropTM One, Thermo Fisher Scientific, USA). DNA solution was then diluted to a working concentration with distilled water and stored at −20°C until use.
Genotyping
Polymerase chain reaction (PCR) was performed using gene-specific primers reported in previous studies (
Song et al. 2016) and developed precisely in this study (
Table 2).
Control varieties were used to compare the size and presence/absence of amplicon between resistant and susceptible lines (
Table 3). Approximately 40 ng of genomic DNA was used in a 20 μL PCR reaction containing 2 μL of primer pairs (10 pmol/μL), 2.0 μL of 10 × PCR buffer, 1.6 μL of dNTP (2.5 mM), and 0.2 μL of
Taq DNA polymerase (5 unit/μL; Promega, USA). The reaction mixture was subjected to the following PCR conditions: initial denaturation at 94°C for 5 minutes, followed by 35 cycles of denaturation at 94°C for 30 seconds, annealing at 55–60°C for 30 seconds, and extension at 72°C for 45–60 seconds, and a final extension step at 72°C for 10 minutes. PCR amplified products of genes related to hybrid traits (
S5,
Rf4), bacterial blight (
Xa3,
Xa7,
xa13, and
Xa21), blast (
Pib,
Pita,
Pi5, and
Pii), and tungro virus (
tsv1) resistant genes were separated on 1.5–2.0% agarose gel and stained with ethidium bromide. PCR amplified products of
Rf3,
Xa4, and
xa5 were run on a fragment capillary gel electrophoresis system (Fragment analyzer, USA). Fragments were sized and scored using PROSize 2.0 software (Fragment analyzer, USA). Amplification of
Bph18(t) was performed on an Eco
TM Real-Time PCR System (Illumina, San Diego, CA, USA) according to the user guide manual. Allele callings of amplified fragments of hybrid lines and control varieties were based on their respective resistance and susceptible controls.
Resistance to disease and insect was assessed during summer season (2018) in an experimental field at Haihau District, Namdinh Province, Vietnam. To evaluate bacterial blight, rice blast, and brown planthopper resistance, we selected F
1 hybrid lines (
Table 4) and control varieties with hybrid related genes. A randomized complete block design with three replicates was used. Eleven F
1 hybrid lines and control varieties were sown on February 15, 2018 and transplanted a week later. Plot size was 1 m × 1 m. Row to row and plant to plant spacing were 20 cm × 20 cm. Three varieties, namely TN1, B40, and Tetep, were used as control varieties to compare resistant and susceptible lines. Disease/insect pest outbreaks and damages in the filed were screened three times (April 3, April 24, and May 4) to determine degree of resistance and susceptibility. Levels of resistance were bioassayed with a standard seed-box screening test (rice brown planthopper), sequential planting and nursery screening test (rice blast), and inoculation screening under a net house (bacterial blight) according to IRRI protocol for accurate determination.
Investigation of agronomic traits and yields was performed during rainy season at the same place as bioassay. A randomized complete block design with three replicates was used and eight F1 hybrid combinations and two control varieties were transplanted on June 29, 2018. Plot size was 4.8 m2. Row to row and plant to plant spacing were 20 cm × 20 cm. IIA838 and BC15 were used as control varieties to compare agronomic traits and yield performance. Measurement of plant height, tiller number, heading date, and ripening conditions was conducted during cultivation period. Yields were investigated from October 8 to October 18.
RESULTS
Development of F1 hybrid combinations with genes related to hybrid traits and disease/insect resistance using MAS
A total of 240 genetic resources and F
1 hybrid combinations were genotyped using PCR-based markers related to hybrid traits and disease/insect resistance genes based on fragment size differences and presence/absence (
Figs. 1,
2).
We examined the genotype of
S5 gene with Indel markers targeting sequences flanking the 136-bp deletion and SNP markers converting SNPs between
indica and
japonica alleles into PCR-based allele specific markers. S5-Indel amplified a 281-bp fragment with 136-bp deletion of intron in 18 hybrid breeding lines (Entry No. 111–128 in
Table 1) with wide-compatibility. The restorer of fertility gene
Rf3 was detected in 176 and 216 hybrid lines using microsatellite marker RF3-5 and RF3-10, respectively. The major gene
Rf4 was amplified in 96 and 125 hybrid lines using RF4-14 and M19280, respectively (
Supplementary Table S1).
We determined the presence of bacterial blight resistance genes (
Xa3,
Xa4,
xa5,
Xa7,
xa13,
Xa21), blast resistance genes (
Pita,
Pib,
Pi5,
Pii), a brown planthopper resistance gene (
Bph18(t)), and a tungro virus resistance gene (
tsv1) in genetic resources and F
1 hybrid combinations (
Supplementary Table S2). Among 240 genetic resources and F
1 hybrid combinations, bacterial blight genes
Xa3,
Xa4,
xa5,
Xa7,
xa13,
Xa21 were found in 20, 32, 36, 10, 3, and 4 F
1 hybrid combinations, respectively, while most lines (226 genetic resources) were detected to carry
Pib. We further surveyed the distribution of resistant genes to brown planthopper using High Resolution Melt (HRM) markers in hybrid rice breeding lines.
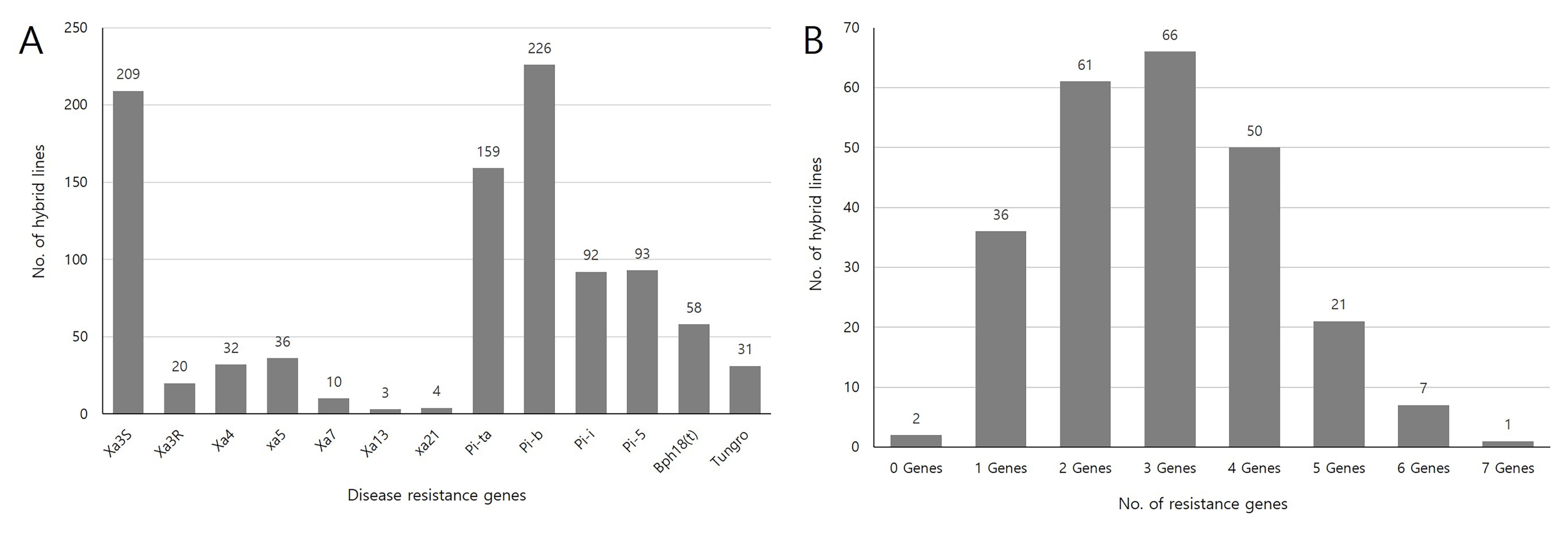
Bph18(t) was present in 24.1% of total genetic resources and F
1 hybrid combinations (
Fig. 3A). Distribution of different gene combination varied greatly among lines. Number of lines with three genes was found to be the highest (27.4%), followed by two-gene-containing lines (25%) and four-gene-containg lines (20.4%). Seven resistance genes combination,
Xa4 +
Xa5 +
Pi-ta +
Pib +
Pi5 +
Bph18(t) +
tsv1, was identified in
indica variety ‘Rumpe’ (
Fig. 3B,
Supplementary Table S3).
In this study, we selected eleven F
1 combinations having
S5+
Rf3+
Rf4 related to hybrid traits among 240 genetic resources and F
1 hybrid combinations. Field screening was performed to identify lines resistant to bacterial blight, blast, and brown planthopper. Eight F
1 hybrid combinations excluding KR0695H, KR0696H, and KR1444H showed resistance to bacterial blight. Meanwhile, in ten F
1 hybrid combinations excluding KR1487H, blast resistance genes were integrated with more than three other resistance genes. F
1 hybrid combinations containing brown planthopper and tungro virus resistance genes had fewer than two combinations. A F
1 hybrid combination, KR2116H, had the most resistance genes among eleven F
1 combinations (
Table 5).
To evaluate resistance to bacterial blight, blast, and brown planthopper, we performed field screen with the selected eleven F
1 combinations and control varieties in the Red Rever Delta region, Vietnam. F
1 hybrid combination KR1994H without a resistance gene of rice brown planthopper showed resistance to rice brown planthopper at 6 days after treatment compared to BTP 33 (resistant variety) but showed medium sensitivity to rice brown planthopper at 8 days after treatment. Most of F
1 hybrid combinations (except KR1994H) with two or most blast resistance genes were found to have very strong resistance to blast fungus. F
1 hybrid combination KR0203H showed the same level of resistance to bacterial blight as OM 1490 (susceptible variety). Eleven F
1 combinations were found to have strong resistance to bacterial blight (
Table 6).
We developed eight F
1 hybrid combinations selected from eleven F
1 combinations that accumulated genes related to hybrid traits and disease/insect resistance genes by genetic analysis and bioassay (
Table 7). Their agronimic traits were compared with control varieties. Heading date was the earilest in a control variety, IIA838 (May 12), while it was the latest in F
1 hybrid combinations KR1454H (May 19) and KR1354H (May 19). Heading days of KR0695H, KR0696H, KR1497H, and KR2116H were the same (May 15). Regarding plant height, KR0695H (122.6 cm) and KR0696H (122.9 cm) were the highest while KR1497H (111.6 cm) was the shortest. Panicle length of KR1354H (31.1 cm) was the longest, followed by that of KR0203H (29.4 cm) and KR2116H (29.0 cm). Panicle lengths of control varieties and F
1 hybrid combinations were slimilar (within 27–28 cm). Percent ripened grain was the hightest in KR1498H (88.5%) but the lowest in KR0695H (65.2%). Thousand grain weights of KR0203H (27.0 g) and KR0696H (27.0 g) were heavier than other F
1 combinations. KR0695H (11.8 Ton/ha) showed the hightest yield of rough rice among the eight F
1 combinations selected. KR1455H (9.6 Ton/ha) had lower yield while the rest had yield of 10–11 Ton/ha (
Table 7). F
1 hybrid combinations KR0695H (11.8 Ton/ha) and KR0696 (11.1 Ton/ha) showed the highest yield as seen in
Fig. 4.
DISCUSSION
Plant breeding based on MAS has become an important approach to ensure the development of crop varieties with durable resistance to diseases and insects and to accelerate the application of MAB (
Lau et al. 2015;
Hu et al. 2016). In this study, we conducted screening of hybrid traits and genes related to disease/insect resistance using molecular MAS. We also performed field screening of responses to diseases and insects for developing F
1 hybrid rice varieties with high yield.
Three hybrid genes (
S5,
Rf3,
Rf4) and thirteen disease/insect resistance genes (bacterial blight resistance genes,
Xa3,
Xa4,
xa5,
Xa7,
xa13,
Xa21; blast disease resistance genes,
Pi-ta,
Pib,
Pi5,
Pii; a leafhopper resistance gene,
Bph; tungro virus resistance gene,
tsv1) were analyzed for 240 genetic resources and F
1 hybrid combinations using molecular marker analysis. Most of these genetic resources showed correlation with
S5,
Rf3, and
Rf4 genes. The total number of combinations with diease/insect resistance genes was 72. All possible combinations were made by a maximum of seven resistance genes. The seven resistance gene combination,
Xa4 +
Xa5 +
Pi-ta +
Pib +
Pi5 +
Bph18(t) +
tsv1, was identified in
indica variety. Eleven F
1 combinations that accumulated useful genes related to hybrid traits were selected and used for field screening at the Red River Delta, Vietnam. As a result, they exhibited high resistance to bacterial blight and blast diseases. Nine F
1 combinations that accumuated F
1 hybrid genes,
S5,
Rf3, and
Rf4, and biotic stress resistance genes for bacterial blight, blast, and brown planthopper resistance were also integrated. Agronomic traits were compared with control varieties and the productivity of F
1 combinations was analyzed. KR1498H (88.5%) showed the highest in percent ripened grain, but KR0695H (65.2%) was the lowest. Thousand grain weights of KR0203H (27.0 g) and KR0696H (27.0 g) were heavier than those of other F
1 combinations. Among the eight selected hybrid combinations, KR0695H (11.8 Ton/ha) and KR0696 (11.1 Ton/ha) showed the hightest rough rice yield (
Fig. 4). These two hybrid rice combinations showed higher yield than those of Suweon Hybrid 1 (8.34 Ton/ha) and Suweon Hybrid 2 (7.34 Ton/ha) bred in Korea in 1990s and also much higher than those of ICGHR 2 (6.99 Ton/ha) and Hao uu 19 (6.75 Ton/ha) grown in Vietnam, implying that these two hybrid variety candidates would be competitive for future export.
Recently, damages by diseases and pests have increased due to climate change. As a result, rice production is falling into an unstable state. Accordingly, developing new varieties with high yield and durable resistance to disease and pest is indespensible. Philippines and China have developed varieties that integrate two, three, or more than four resistance genes using MAS. For example,
Xa21 resistance gene has been integrated into IR72, showing a widespread resistance (
Tu et al. 1998;
Singh et al. 2011). In South Korea, breeders have also developed countermeasures against mutant strain K3a by breeding a resistance variety Jinbaek which integrates Xa3 and xa5 genes (
Kim et al. 2009;
Shin et al. 2011). These varieties have diversified single-resistance genes and integrated main genes for stabilizing yield and durability of rice varieties.
The current direction of plant breeding is to select resistance lines by using molecular markers. By using molecular markers for selection, it can reduce the effort of screening resistance genes. Stable resistance genotypes can be selected at early generation so that its resistance can remain longer (
Peleman and Voort 2003). However, to confirm whether genes inserted or accumulated by MAS show resistance or not, it is nessary to screen selected plants by inoculation or in the field. To develop resistance varieties with useful genes and resistant genes into practically cultivated varieties, comprehensive considerations are needed through selection of phenotypes of various agronomic traits in the breeding process.
In this study, eight F1 combinations were superior F1 combinations that expressed useful genes with complex disease and pest resistance. This result could be useful for breeding F1 hybrid varieties with complex biotic resistance genes for future export.
Supplementary Information
ACKNOWLEDGEMENTS
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Golden Seed Project funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (213009-05-3-WT211).
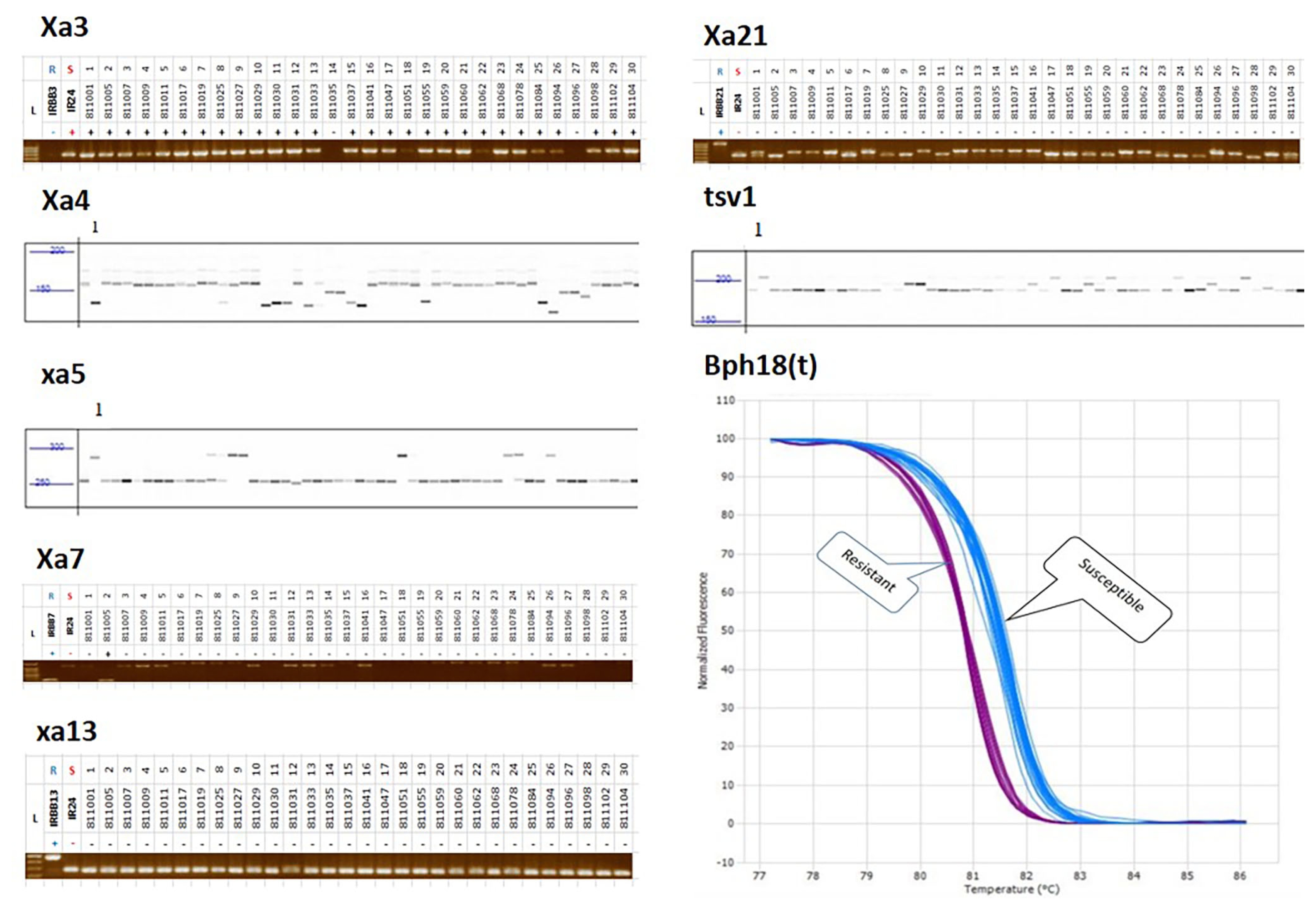
Fig. 1PCR products for genotyping with each marker linked to disease and insect resistance in breeding lines. S5, Rf4, Pita, Pib, Pii, and Pi5 show band patterns on agarose gel electrophoresis; Rf3 indicates size differentiation by fragment analyzer gel electrophoresis. L, R, S indicates ladder marker, resistant, and susceptible control variety, respectively.

Fig. 2PCR products for genotyping with each marker linked to hybried related genes and blast resistance genes in breeding lines. Xa3, Xa7, xa13, and Xa21 show band patterns on gel electrophoresis; xa4, xa5, and tsv1 indicate size differentiation by fragment analyzer gel electrophoresis. Bph18(t) shows HRM curve profiles of 240 genetic resources and F1 hybrid combinations. L, R, S indicates ladder marker, resistant, and susceptible control variety, respectively.
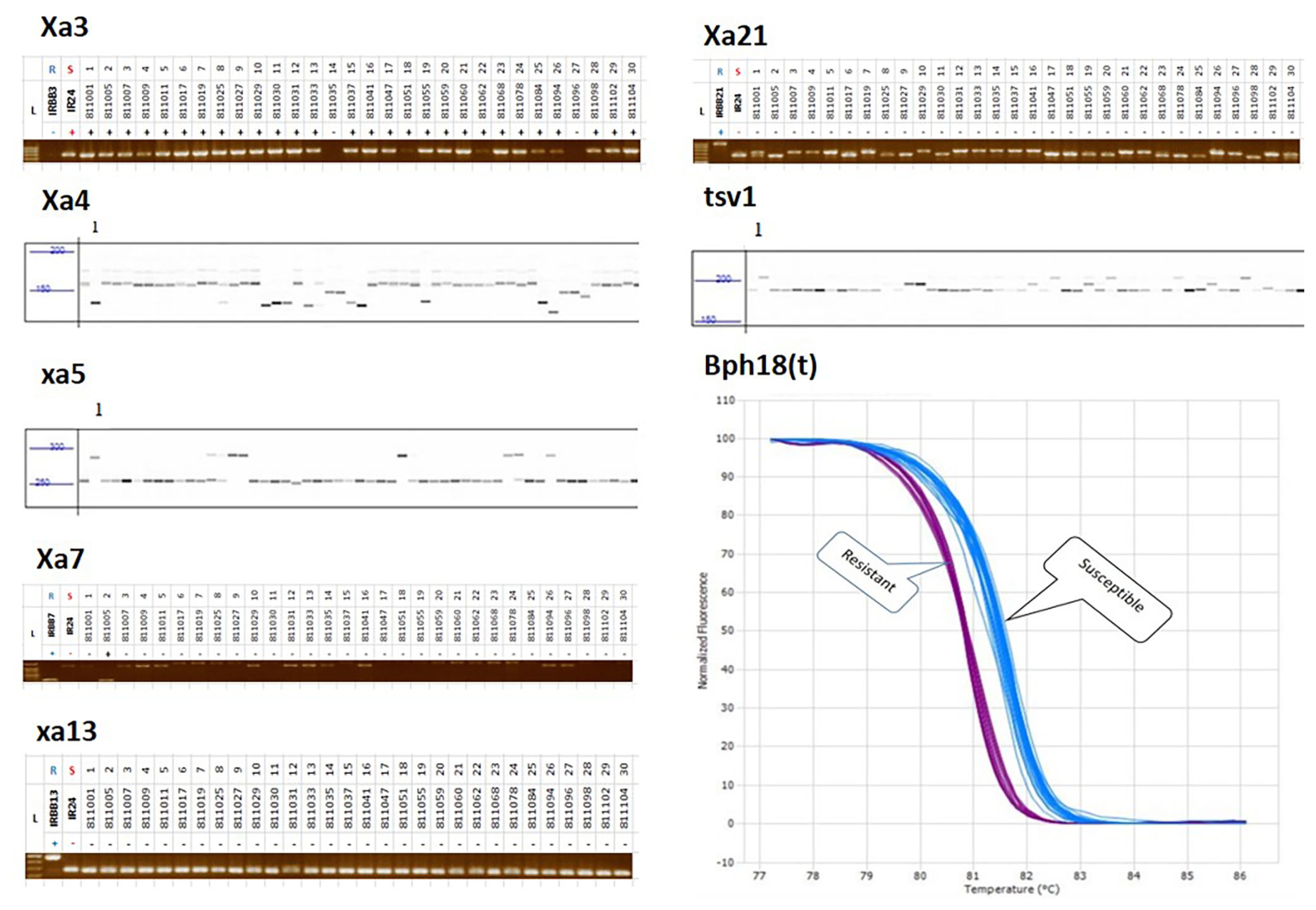
Fig. 3(A) Frequency distribution of disease and insect resistant genes among 240 genetic resources and F1 hybrid combinations, and (B) Frequency of rice lines having different numbers of resistant genes.
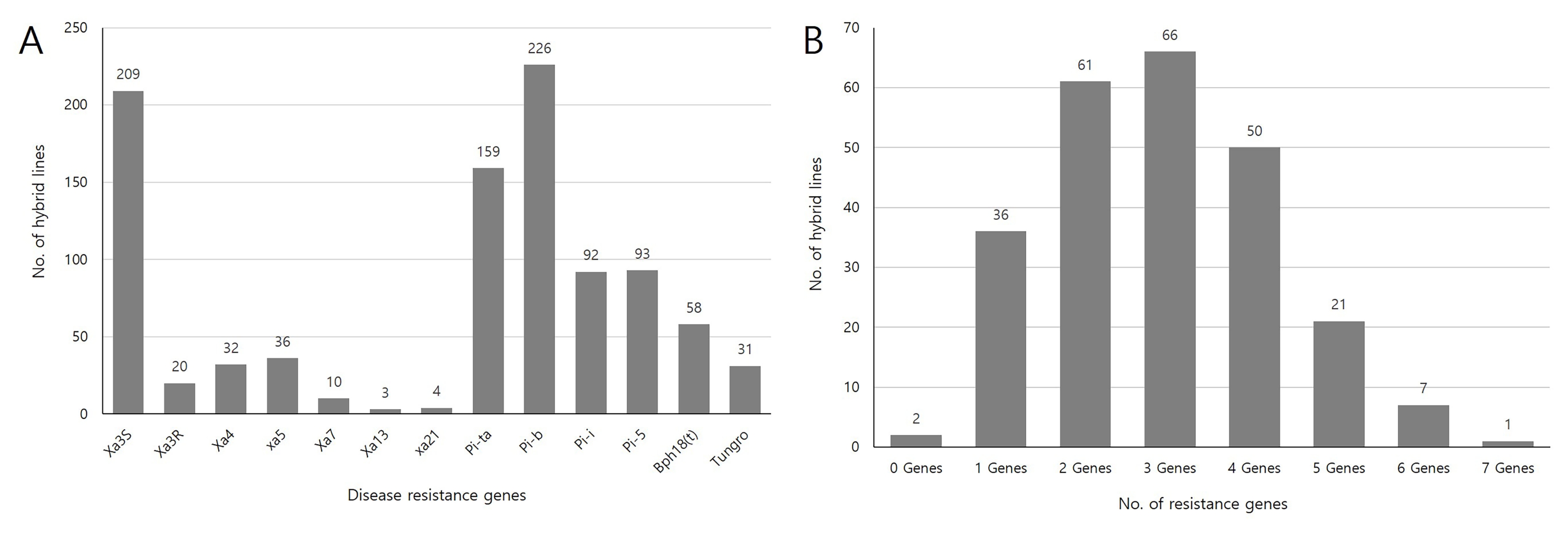
Fig. 4Panicle type and growth performance of F1 hybrid combinations, KR0695H, and KR0696H. (A) Panicle length and morphologies of KR0695H and KR0696H; (B) Growth performance of KR0695H and KR0696H at Hai Hau field in Vietnam.

Table 1List of 240 genetic resources and hybrid rice combinations used in this study.
Table 1
|
Entry No. |
Designation |
Cross/Origin |
Remark |
Source |
|
1 |
9019 |
China |
Restorer |
811001 |
|
2 |
Basmati 370 |
India |
Aromatic rice |
811005 |
|
3 |
Chiherang |
Cambodia |
Local variety |
811007 |
|
4 |
Cigeulis |
Cambodia |
Local variety |
811009 |
|
5 |
Ciliwung |
Cambodia |
Local variety |
811011 |
|
6 |
FFZ1 |
China |
Local variety |
811017 |
|
7 |
Giza178 |
Egypt |
Local variety |
811019 |
|
8 |
HHZ12-SAL2-Y3-Y1 |
China |
Breeding line |
811025 |
|
9 |
HHZ12-SAL8-Y1-Y2 |
China |
Breeding line |
811027 |
|
10 |
IR05N412 |
IR72875-94-3-3-2/IR73707-45-3-2-3 |
Breeding line, IRRI |
811029 |
|
11 |
IR05N412 |
IR72875-94-3-3-2/IR73707-45-3-2-3 |
Breeding line, IRRI |
811030 |
|
12 |
IR06A145 |
IR02A127/JANAKI |
Breeding line, IRRI |
811031 |
|
13 |
IR09N538 |
IRRI 132/PR 30138-35-2//IR04N114 |
Breeding line, IRRI |
811033 |
|
14 |
IR10A267 |
IR02A483/IRBB 60-1 |
Breeding line, IRRI |
811035 |
|
15 |
IR10N305-1 |
IRRI |
Breeding line, IRRI |
811037 |
|
16 |
IR78581-12-3-2-2-1 |
IRRI |
Breeding line, IRRI |
811041 |
|
17 |
IR98070-kB13-1-2-2-1-1-1 |
IR72903-131-1-2-3R/IR72998-78-1-3-2 R |
Restorer |
811047 |
|
18 |
IR98070-kB14-1-2-3-1-4-1 |
IR72903-131-1-2-3R/IR72998-78-1-3-2 R |
Restorer |
811051 |
|
19 |
IR98102-kB9-1-1-1-1-1-1 |
IR65622-151-2-2-2R/IR73885-1-4-3-2-1-10R |
Restorer |
811055 |
|
20 |
IR98102-kB9-1-1-3-1-1-B2-1 |
IR65622-151-2-2-2R/IR73885-1-4-3-2-1-10R |
Restorer |
811059 |
|
21 |
IR98229-9-2-1-k1-1-6-1-1 |
IR08A138/IR72998-93-3-3-2R |
Restorer |
811060 |
|
22 |
IR98229-24-1-1-k1-1-1-1-1 |
IR08A138/IR72998-93-3-3-2R |
Restorer |
811062 |
|
23 |
IR98241-24-2-1-k1-1-1-1 |
IR06N172/IR86612-38-2-2-1-1-1-1-1 |
Breeding line, IRRI |
811068 |
|
24 |
IR101872-46-1-K1-1-2-1 |
MingHui63/IR86590-22-2-2-1-3-1-1-1 |
Breeding line, IRRI |
811078 |
|
25 |
L24 |
China |
Local variety |
811084 |
|
26 |
NSIC 238 |
Philippines |
Local variety |
811094 |
|
27 |
KR3R |
MY3R(SSLR-12, Myanma Col. 2012) |
Local variety |
811096 |
|
28 |
OM52 |
Vietnam |
Local variety |
811098 |
|
29 |
TLR353 |
Vietnam |
Local variety |
811102 |
|
30 |
TLR363 |
Vietnam |
Local variety |
811104 |
|
31 |
Vietnam collection 1 |
Vietnam |
Local variety |
811106 |
|
32 |
WEED TOLERANT RICE 1-1 |
IRRI |
Breeding line, IRRI |
811108 |
|
33 |
Zhong419 |
China |
Local variety |
811110 |
|
34 |
6527 |
Unknown |
Local variety |
811113 |
|
35 |
Com. Collection 3 |
Com Col. (Cambodia Col. 2015) |
Local variety |
811114 |
|
36 |
HHZ1-Y4-Y1 |
HUANG-HUA-ZHAN*2/YUE-XIANG-ZHAN |
Breeding line, China |
811120 |
|
37 |
HUA564 |
China |
Breeding line |
811122 |
|
38 |
IR02A127 |
IR00A107/IR62243-41-1-3-3 |
Breeding line, IRRI |
811124 |
|
39 |
IR05N359 |
IR72158-11-5-2-3/Ir72903-121-2-1-2 |
Breeding line, IRRI |
811126 |
|
40 |
IR06A181 |
IR71718-59-1-2-3/IR72 |
Breeding line, IRRI |
811128 |
|
41 |
IR08N136 |
IR72967-12-2-3/PR 31090-33-2-1 |
Breeding line, IRRI |
811130 |
|
42 |
IR10K153 |
HR 24580-15-1/IR03K105 |
Breeding line, IRRI |
811132 |
|
43 |
IR11A303 |
IR04A427/IR72875-94-3-3-2 |
Breeding line, IRRI |
811134 |
|
44 |
IR11A334 |
IR04A427/IRRI 115 |
Breeding line, IRRI |
811136 |
|
45 |
Japonica 1 |
Philippines |
Local variety |
811138 |
|
46 |
SACG4 |
IRRI |
Local variety |
811150 |
|
47 |
SAGC-02 |
IRRI |
Local variety |
811152 |
|
48 |
ZH1 |
China |
Local variety |
811156 |
|
49 |
HHZ11-Y10-DT3-Y3 |
China |
Breeding line |
811158 |
|
50 |
HHZ5-DT1-DT1 |
China |
Breeding line |
811160 |
|
51 |
HHZ5-SAL12-DT3-Y2 |
China |
Breeding line |
811162 |
|
52 |
HHZ5-SAL8-DT2-SAL1 |
China |
Breeding line |
811164 |
|
53 |
HHZ8-SAL14-SAL1-SUB1 |
China |
Breeding line |
811166 |
|
54 |
HHZ8-SAL6-SAL3-SAL1 |
China |
Breeding line |
811168 |
|
55 |
HHZ8-SAL6-SAL3-Y1 |
China |
Breeding line |
811170 |
|
56 |
HHZ8-SAL6-SAL3-Y2 |
HUANG-HUA-ZHAN*2/PHALGUNA |
Breeding line |
811172 |
|
57 |
IR06M150 |
MEM BERANO/PADI ABANG GOGO |
Breeding line, IRRI |
811174 |
|
58 |
IR72 (IR72) |
IR19661-9-2-3-3/IR15795-199-3-3//IR9129-209-2-2-2-1 |
Advanced variety, IRRI |
811178 |
|
59 |
Teqing |
China |
Local variety |
811180 |
|
60 |
TME80518 |
TME 80518 |
Local variety |
811182 |
|
61 |
KR2R |
MR2R(Myanma Col. 2012) |
Restorer |
811184 |
|
62 |
A 69-1 |
IRRI |
Breeding line |
811186 |
|
63 |
BR 28-SalTol |
Bangladesh |
Breeding line |
811188 |
|
64 |
Chulsa |
Cambodia |
Local variety |
811190 |
|
65 |
IR04A395 |
IRRI |
Breeding line |
811192 |
|
66 |
IR07A234 |
NSIC RC 138/IRRI 123 |
Breeding line |
811194 |
|
67 |
IR10A 227 |
IR01A154/IR72870-19-2-2-3//Irri 123 |
Breeding line |
811196 |
|
68 |
IR65482-4-136 |
IRRI |
Breeding line |
811198 |
|
69 |
AN 424627 |
IRRI |
Local variety |
811200 |
|
70 |
BR 26 |
Bangladesh |
Local variety |
811202 |
|
71 |
Daerip H-R11-2-1-1-1 |
78/Daelipbyeo F1 |
Breeding line |
811204 |
|
72 |
IR64 Sub1 |
IRRI |
Breeding line |
811206 |
|
73 |
IR68897H-B24-B-1-1-2 |
IRRI |
Breeding line |
811208 |
|
74 |
IR09A228 |
PR29232-B-17-2-1-1/IR 64 |
Breeding line, |
811210 |
|
75 |
J.P.5-IR946-2-2-2/IR1635-1F |
IRRI |
Breeding line |
811212 |
|
76 |
IR97727-82-1-2-2 |
IRRI |
Breeding line |
811214 |
|
77 |
IR98073-3-1-1-K1-1 |
IR72903-131-1-2-3R/IR85485-106-B-B-1-1-1-1 |
Breeding line |
811216 |
|
78 |
IR98107-kB3-1-1-2-1-1 |
IR71604-4-1-4-4-4-2-2-2R/IR65622-151-2-2-2R |
Breeding line |
811218 |
|
79 |
IR98108-kB13-1-2-3-1-2-1 |
IRRI |
Breeding line |
811220 |
|
80 |
IR98161-2-1-1-k2-2-2 |
IR86409-3-1-1-1-1-1/IRBB66 |
Breeding line |
811222 |
|
81 |
IR98194-9-2-1-k1-1-1 |
IRRI |
Breeding line |
811224 |
|
82 |
IR101861-7-1-K1-1-1 |
MingHui63/IR03A550 |
Breeding line |
811226 |
|
83 |
IRRI 102 |
IR4215-301-2-2-6/BG90-2//IR19661-131-1-2 |
Advanced variety, IRRI |
811234 |
|
84 |
Jasponica Bulk Aroma4-1 |
Philippines |
Breeding line |
811236 |
|
85 |
Jasponica Bulk Aroma5-1 |
Philippines |
Breeding line |
811238 |
|
86 |
KCD1 |
IRRI |
Local variety |
811240 |
|
87 |
MY1H-R23-3-1-1-1-1 |
MY 1 A/R |
Breeding line |
811242 |
|
88 |
MY1H-R23-3-2-1-1-1 |
MY 1 A/R |
Breeding line |
811244 |
|
89 |
NSIC 222 |
Philippines |
Local variety |
811246 |
|
90 |
OM100411 |
Vietnam |
Local variety |
811248 |
|
91 |
OM10375 |
Vietnam |
Local variety |
811250 |
|
92 |
OM4900 |
Vietnam |
Local variety |
811252 |
|
93 |
OM7347 |
Vietnam |
Local variety |
811254 |
|
94 |
OM8108 |
Vietnam |
Local variety |
811256 |
|
95 |
OMCS 2012 |
Vietnam |
Local variety |
811258 |
|
96 |
Pearl riceH-R28-3-2-1-1 |
IRRI |
Breeding line |
811260 |
|
97 |
Pearl riceH-R52-2-1-1-1 |
IRRI |
Breeding line |
811262 |
|
98 |
PHB73H-R9-2-1-1-1 |
IRRI |
Breeding line |
811264 |
|
99 |
Phka Romeat |
Cambodia |
Local variety |
811266 |
|
100 |
Phka Rumchang |
Cambodia |
Local variety |
811268 |
|
101 |
Phka Rumchek |
Cambodia |
Local variety |
811270 |
|
102 |
Phka Rumdeng |
Cambodia |
Local variety |
811272 |
|
103 |
Phka Rumduol |
Cambodia |
Local variety |
811274 |
|
104 |
Popoul |
Cambodia |
Local variety |
811276 |
|
105 |
Rumpe |
Cambodia |
Local variety |
811278 |
|
106 |
S430 |
China |
Local variety |
811280 |
|
107 |
San pidao |
Cambodia |
Local variety |
811283 |
|
108 |
TH82H-R2-1-1-1-1-1 |
Vietnam |
Breeding line |
811284 |
|
109 |
TLR405 |
Vietnam |
- |
811286 |
|
110 |
TLR407 |
Vietnam |
- |
811288 |
|
111 |
WC467-2-1-1-1-2-2-1-1 |
(Milyang154/Norin PL9//Milyang154)/Milyang154 |
Breeding line |
811290 |
|
112 |
WC467-2-3-2-1-2-1-1-1 |
(Milyang154/Norin PL9//Milyang154)/Milyang154 |
Breeding line |
811292 |
|
113 |
WC468-2-1-3-1-2-3-1-1 |
(Milyang154/Norin PL9//Milyang154)/Milyang154 |
Breeding line |
811294 |
|
114 |
WC488-6-1-1-2-1-1-1-1 |
(Milyang23//Norin PL9/Dular///Milyang23)/Milyang23 |
Breeding line |
811296 |
|
115 |
WC488-6-1-1-2-1-3-1-1 |
(Milyang23//Norin PL9/Dular///Milyang23)/Milyang23 |
Breeding line |
811298 |
|
116 |
WC495-1-1-1-1-2-3-1-1 |
(Milyang160//Norin PL9/Dular///Areumbyeo)/Areumbyeo |
Breeding line |
811300 |
|
117 |
WC509-4-1-2-1-2-3-1-1 |
(Jangsungbyeo/Dular//Jangsungbyeo)/Jangsungbyeo |
Breeding line |
811302 |
|
118 |
WC540-2-1-3-1-1-1-1-1 |
(Milyang160/CPSLO 17//Areumbyeo)/Areumbyeo |
Breeding line |
811304 |
|
119 |
WC540-2-1-3-1-2-2-1-1 |
(Milyang160/CPSLO 17//Areumbyeo)/Areumbyeo |
Breeding line |
811308 |
|
120 |
WC540-2-3-3-1-2-3-1-1 |
(Milyang160/CPSLO 17//Areumbyeo)/Areumbyeo |
Breeding line |
811312 |
|
121 |
WC549-1-1-2-1-1-1-1-1 |
(Yongmunbyeo/CPSLO 17//Yongmunbyeo)/Yongmunbyeo |
Breeding line |
811314 |
|
122 |
WC570-2-1-3-1-1-1-1-1 |
(Samgangbyeo//Dular/Samgangbyeo///Samgangbyeo)/Samganbyeo |
Breeding line |
811316 |
|
123 |
WC634-1-1-2-1-2-1-1-1 |
(Yongjubyeo//N22/Yongjubyeo)/Yongjubyeo |
Breeding line |
811320 |
|
124 |
WC647-1-1-1-1-1-2-1-1 |
Ilpumbyeo/IR65600-96-1-2-2//Ilpumbyeo)/Ilpumbyeo |
Breeding line |
811324 |
|
125 |
WC962-1-2-1-1-1-1 |
02428-97-2/Areumbyeo//3*Yeonghaebyeo |
Breeding line |
811328 |
|
126 |
WC964-1-1-2-1-3-1-1-1 |
Sambaekbyeo/02428-97-1//3*Sambaekbyeo |
Breeding line |
811332 |
|
127 |
WC972-3-3-1-1-1-1 |
02428/3*Yongmunbyeo |
Breeding line |
811334 |
|
128 |
WC972-4-2-1-3-2-1 |
02428/3*Yongmunbyeo |
Breeding line |
811340 |
|
129 |
Corn rice |
China |
Local variety |
811375 |
|
130 |
Restorer 1-1 |
Unknown |
Breeding line |
811377 |
|
131 |
Restorer 2-1 |
Unknown |
Breeding line |
811379 |
|
132 |
Restorer 3-1 |
Unknown |
Breeding line |
811381 |
|
133 |
Indonesia col. 1(2016) |
Indonesia Col. (Cambodia Col. 2016) |
Local variety |
811383 |
|
134 |
Indonesia col. 2(2016) |
Indonesia Col. (Cambodia Col. 2016) |
Local variety |
811385 |
|
135 |
HYT 116H-3-1-3-2-1-1 |
HYT 116 H |
Breeding line |
811391 |
|
136 |
HYT 116H-17-2-1-1-2-1 |
HYT 116 H |
Breeding line |
811395 |
|
137 |
HYT 116H-31-2-3-1-1-1 |
HYT 116 H |
Breeding line |
811397 |
|
138 |
HYT 116H-46-1-1-1-1-1 |
HYT 116 H |
Breeding line |
811401 |
|
139 |
HYT 116H-50-1-1-2-1-1 |
HYT 116 H |
Breeding line |
811405 |
|
140 |
HYT 119H-18-2-2-2-1-1 |
HYT 119 H |
Breeding line |
811407 |
|
141 |
HYT 119H-18-3-2-2-2-2 |
HYT 119 H |
Breeding line |
811411 |
|
142 |
HYT 119H-21-1-3-2-2-1 |
HYT 119 H |
Breeding line |
811414 |
|
143 |
HYT 123H-13-3-3-1-2-1 |
HYT 123H |
Breeding line |
811416 |
|
144 |
HYT 124H-3-3-1-1-2-1 |
HYT 124H |
Breeding line |
811420 |
|
145 |
HYT 128H-8-2-2-2-2-1 |
HYT 128H |
Breeding line |
811426 |
|
146 |
HYT 128H-11-2-1-1-2-1 |
HYT 128H |
Breeding line |
811428 |
|
147 |
HYT 130H-3-2-3-1-1-1 |
HYT 130H |
Breeding line |
811432 |
|
148 |
CASH 1H-1-2-3-1-1-1 |
CASH 1H |
Breeding line |
811434 |
|
149 |
CASH 1H-4-3-2-1-1-1 |
CASH 1H |
Breeding line |
811440 |
|
150 |
CASH 1H-34-2-2-2-1-1 |
CASH 1H |
Breeding line |
811444 |
|
151 |
Hipa Jatim 1H-5-2-2-3-2-1 |
Hipa Jatim 1 H |
Breeding line |
811446 |
|
152 |
IR68897H-94 B-1-3-3-1-1 |
IR68897H |
Breeding line |
811448 |
|
153 |
IR101922-BK-KB-2-2-1 |
Maybelle/PSBRC80 |
Breeding line |
811452 |
|
154 |
IR101922-BK-KB-6-1-1 |
Maybelle/PSBRC80 |
Breeding line |
811456 |
|
155 |
IR101923-BK-KB-1-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86508-4-2-3-2-1-1-2-1 |
Breeding line |
811458 |
|
156 |
IR101923-BK-KB-3-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86508-4-2-3-2-1-1-2-1 |
Breeding line |
811460 |
|
157 |
IR101923-BK-KB-4-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86508-4-2-3-2-1-1-2-1 |
Breeding line |
811462 |
|
158 |
IR101923-BK-KB-4-3-1 |
IR86427-29-6-1-3-1-1-1-1/IR86508-4-2-3-2-1-1-2-1 |
Breeding line |
811464 |
|
159 |
IR101923-BK-KB-7-1-1 |
IR86427-29-6-1-3-1-1-1-1/IR86508-4-2-3-2-1-1-2-1 |
Breeding line |
811466 |
|
160 |
IR101924-BK-KB-2-3-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811468 |
|
161 |
IR101924-BK-KB-7-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811470 |
|
162 |
IR101924-BK-KB-10-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811472 |
|
163 |
IR101924-BK-KB-11-3-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811474 |
|
164 |
IR101924-BK-KB-14-1-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811476 |
|
165 |
IR101924-BK-KB-14-2-1 |
IR86427-29-6-1-3-1-1-1-1/IR86519-12-1-3-1-1-1-1-1 |
Breeding line |
811478 |
|
166 |
IR101933-BK-KB-2-1-1 |
IR86505-6-4-3-1-1-1-1-1/MingHui63 |
Breeding line |
811480 |
|
167 |
IR101937-BK-KB-3-2-1 |
IR86505-6-4-3-1-1-1-1-1/IR86612-26-1-1-1-1-2-1-1 |
Breeding line |
811482 |
|
168 |
IR101937-BK-KB-9-3-1 |
IR86505-6-4-3-1-1-1-1-1/IR86612-26-1-1-1-1-2-1-1 |
Breeding line |
811484 |
|
169 |
KR0301-B-12-1-1-1 |
OM 052/NSIC RC 238 |
Breeding line |
811485 |
|
170 |
KR0302-B-5-2-2-1 |
OM 052/Minghui 63 |
Breeding line |
811490 |
|
171 |
KR0302-B-5-3-1-1 |
OM 052/Minghui 63 |
Breeding line |
811492 |
|
172 |
KR0302-B-14-2-1-1 |
OM 052/Minghui 63 |
Breeding line |
811496 |
|
173 |
KR0302-B-17-1-3-1 |
OM 052/Minghui 63 |
Breeding line |
811498 |
|
174 |
KR0302-B-10-2-1-1 |
OM 052/Minghui 63 |
Breeding line |
811500 |
|
175 |
KR0302-B-10-2-2-1 |
OM 052/Minghui 63 |
Breeding line |
811502 |
|
176 |
KR0302-B-13-1-1-1 |
OM 052/Minghui 63 |
Breeding line |
811506 |
|
177 |
GR19-1-8-2-2-1 |
2A/3m164 |
Breeding line |
811510 |
|
178 |
GR20-1-3-1-1-1 |
2A/3m170 |
Breeding line |
811512 |
|
179 |
PHB 73H-1-2-3-1-2-1 |
PHB 73 |
Breeding line |
811514 |
|
180 |
PHB 73H-18-2-2-3-1-1 |
PHB 73 |
Breeding line |
811522 |
|
181 |
Matibay H-5-3-1-1-1-1 |
Matibay |
Breeding line |
811526 |
|
182 |
ABp H-20-2-1-1-1-1 |
Arize Bigante plusH |
Breeding line |
811532 |
|
183 |
ABp H-29-2-1-1-2-1 |
Arize Bigante plusH |
Breeding line |
811538 |
|
184 |
ABp H-33-2-3-2-1-1 |
Arize Bigante plusH |
Breeding line |
811540 |
|
185 |
Hipa Jatim 2H-1-3-3-1-2-1 |
Hipa Jatim 2 H |
Breeding line |
811542 |
|
186 |
HYT 106H-17-2-1-2-1-1 |
HYT 106H |
Breeding line |
811544 |
|
187 |
HYT 106H-17-2-2-3-2-1 |
HYT 106H |
Breeding line |
811548 |
|
188 |
HYT 106H-17-2-3-1-2-1 |
HYT 106H |
Breeding line |
811551 |
|
189 |
HYT 106H-17-3-1-1-1-1 |
HYT 106H |
Breeding line |
811559 |
|
190 |
HYT 106H-17-3-2-3-2-1 |
HYT 106H |
Breeding line |
811563 |
|
191 |
HYT 108H-10-2-3-1-3-1 |
HYT 108H |
Breeding line |
811565 |
|
192 |
Jasponica-4-1-1-2-2-1 |
SH 9 |
Breeding line |
811569 |
|
193 |
Jasponica-12-1-1-2-3-1 |
SH 9 |
Breeding line |
811575 |
|
194 |
Jasponica-14-3-1-1-1-1 |
SH 9 |
Breeding line |
811577 |
|
195 |
Jasponica-15-1-1-1-1-1 |
SH 9 |
Breeding line |
811583 |
|
196 |
Jasponica-26-1-1-1-3-1 |
SH 9 |
Breeding line |
811585 |
|
197 |
Jasponica-29-3-1-1-2-1 |
SH 9 |
Breeding line |
811587 |
|
198 |
Jasponica-40-3-1-1-2-1 |
SH 9 |
Breeding line |
811599 |
|
199 |
IR24 |
IRRI |
Advanced variety |
811601 |
|
200 |
IRBB1 |
IRRI |
Bacterial Blight |
811603 |
|
201 |
IRBB3 |
IRRI |
Bacterial Blight |
811605 |
|
202 |
IRBB7 |
IRRI |
Bacterial Blight |
811607 |
|
203 |
IRBB13 |
IRRI |
Bacterial Blight |
811609 |
|
204 |
IRBB55 |
IRRI |
Bacterial Blight |
811611 |
|
205 |
IRBB59 |
IRRI |
Bacterial Blight |
811613 |
|
206 |
Dasan |
Korea |
Local variety |
810801 |
|
207 |
Hanareum |
Korea |
Local variety |
810802 |
|
208 |
KR0203H |
Korea |
Hybrid rice F1 |
810803 |
|
209 |
KR0695H |
Korea |
Hybrid rice F1 |
810804 |
|
210 |
KR0696H |
Korea |
Hybrid rice F1 |
810805 |
|
211 |
KR1454H |
Korea |
Hybrid rice F1 |
810806 |
|
212 |
KR1455H |
Korea |
Hybrid rice F1 |
810807 |
|
213 |
KR1354H |
Korea |
Hybrid rice F1 |
810808 |
|
214 |
KR1497H |
Korea |
Hybrid rice F1 |
810809 |
|
215 |
KR1444H |
Korea |
Hybrid rice F1 |
810810 |
|
216 |
KR1487H |
Korea |
Hybrid rice F1 |
810811 |
|
217 |
KR1994H |
Korea |
Hybrid rice F1 |
810812 |
|
218 |
KR2116H |
Korea |
Hybrid rice F1 |
810813 |
|
219 |
KR2117H |
Korea |
Hybrid rice F1 |
810814 |
|
220 |
KR 1B |
Korea |
Maintainer |
812501 |
|
221 |
KR 1A |
Korea |
CGMS |
812503 |
|
222 |
KR 2B |
Korea |
Maintainer |
812526 |
|
223 |
KR 2A |
Korea |
CGMS |
812528 |
|
224 |
KR 211B |
Korea |
Maintainer |
812551 |
|
225 |
KR 211A |
Korea |
CGMS |
812553 |
|
226 |
MingHui 63 |
China |
Local variety, Restorer |
812571 |
|
227 |
IR75589-31-27-8-33S-5-1 |
IRRI |
Breeding line, TGMS |
812608 |
|
228 |
IR102100-KB5S2-1-4-1-1-1-1 |
IRRI |
Breeding line, TGMS |
812618 |
|
229 |
OM052 |
Vietnam |
Local variety, Restorer |
812636 |
|
230 |
IR102100-KB14-2S2-1-22-2-1-2 |
IRRI |
Breeding line, TGMS |
812718 |
|
231 |
IR98070-kB14-1-2-3-1-1 |
IRRI |
Breeding line |
812736 |
|
232 |
IR98229-2-2-1-K1-1-1 |
IRRI |
Breeding line |
812756 |
|
233 |
IR98229-9-2-1-K1-1-1 |
IRRI |
Breeding line |
812771 |
|
234 |
IR98229-9-2-1-K1-1-3 |
IRRI |
Breeding line |
812776 |
|
235 |
IR101861-7-1-K1-1-1 |
IRRI |
Breeding line |
812786 |
|
236 |
IR98241-24-2-1-k1-1-1 |
IRRI |
Breeding line |
812806 |
|
237 |
IR98102-kB9-1-1-3-1-1-1 |
IRRI |
Breeding line |
812821 |
|
238 |
IR102100-KB12S1-1-25-1-1-1-1 |
IRRI |
Breeding line, TGMS |
812823 |
|
239 |
IR98102-kB21-1-3-1-3-1 |
IRRI |
Breeding line |
812836 |
|
240 |
IR102452-KB3-1-2-2-1-1 |
IRRI |
Breeding line |
812851 |
Table 2Gene-specific PCR primers and their primer sequences used for analysis of related genes.
Table 2
|
Target |
Gene |
Marker name |
Type |
Primer sequence (5′→3′) |
Exspected size (bp) |
Reference |
|
Genes related to hybrid |
S5 |
S5-Indel |
Fw |
CCTACGTTTGACTGCCTGCCTG |
281/417 |
, Sundaram et al. 2010
|
|
|
|
Rv |
CTACACGCGGCTTCGGGAAAGC |
|
|
|
|
Indica-specific SNP |
Fw |
GACAGCAGCATCAACGACTTCC |
527 |
|
|
|
|
Rv |
TCGTCAGTGGGCAAGCAGTAGCTG |
|
|
|
|
Japonica-specific SNP |
Fw |
ACCCTGATATTCTGAGTTACAAGGCATTA |
325 |
|
|
|
|
Rv |
GCTCTTGATGTCCGGTGATACC |
|
|
|
Rf3 |
DRRM-RF3-5 |
Fw |
GATGGCACAGCTTCAGAACA |
120/134 |
Suresh et al. 2012 |
|
|
|
Rv |
CTAATTCTGGGCGAGCAAAG |
|
|
|
|
DRRM-RF3-10 |
Fw |
TCACCTCTTCCTGCTTCGAC |
180/195 |
|
|
|
|
Rv |
CTCCACCAGTGCAGGTTTTT |
|
|
|
Rf4 |
DRCG-RF4-14 |
Fw |
GCAATGCTTGTATTCAGCAAA |
845/885 |
, Tang et al. 2014
|
|
|
|
Rv |
TCCAGCTGTAAATCCGTCAA |
|
|
|
|
M19280 |
Fw |
ATCTGTCCAGACACCATTTTC |
147/141 |
|
|
|
|
Rv |
TCCACTGATGAGTGCATTG |
|
|
|
Blast |
Pita |
YL155/87 |
Fw |
AGCAGGTTATAAGCTAGGCC |
1042 |
, Jia et al. 2002, 2004
|
|
|
|
Rv |
CTACCAACAAGTTCATCAAA |
|
|
|
|
YL183/87 |
Fw |
AGCAGGTTATAAGCTAGCTAT |
|
|
|
|
|
Rv |
CTACCAACAAGTTCATCAAA |
|
|
|
Pib |
NSb |
Fw |
ATCAACTCTGCCACAAAATCC |
629 |
, Kwon et al. 2008
|
|
|
|
Rv |
CCCATATCACCACTTGTTCCCC |
|
|
|
Pi5 |
JJ817 |
Fw |
GATATGGTTGAAAAGCTAATCTCA |
1450 |
, Kwon et al. 2008
|
|
|
|
Rv |
ATCATTGTCCTTCATATTCAGAGT |
|
|
|
Pii |
JJ113 |
Fw |
GGATGATGTGATCTGCAGAG |
484 |
Jeon 2002 |
|
|
|
Rv |
CTCTTGGTGATCTTTGTTAC |
|
|
|
Bacterial blight (BB) |
Xa3 |
BB3-Sus |
Fw |
CGGAGCGACACAGCTATCAT |
743 |
, Hu et al. 2013
|
|
|
|
Rv |
CGTGAGGTTCCCTATGGCGATT |
|
|
|
|
BB3-Re |
Fw |
CCACAATGCCATGTCAGGTGGCATCCCTGCA |
255 |
, Hu et al. 2013
|
|
|
|
Rv |
AGGTGTTGGAGGATTGGCAT |
|
|
|
Xa4 |
RM224 |
Fw |
ATCGATCGATCTTCACGAGG |
150/120 |
, McCouch et al. 2002
|
|
|
|
Rv |
TGCTATAAAAGGCATTCGGG |
|
|
|
Xa5 |
RM122 |
Fw |
GCACTGCAACCATCAATGAATC |
236/232 |
, Chen et al. 1997
|
|
|
|
Rv |
CCTAGGAGAAACTAGCCGTCCA |
|
|
|
Xa7 |
M5 |
Fw |
CGATCTTACTGGCTCTGCAACTCTGT |
294/1170 |
, Porter et al. 2013
|
|
|
|
Rv |
GCATGTCTGTGTCGATTCGTCCGTACGA |
|
|
|
Xa13 |
Xa13prom |
Fw |
GGCCATGGCTCAGTGTTTAT |
1000/520 |
, Zhang et al. 1996; Singh et al. 2011
|
|
|
|
Rv |
GAGCTCCAGCTCTCCAAATG |
|
|
|
Xa21 |
pTA248 |
Fw |
AGACGCGGAAGGGTGGTTCCCGGA |
1000/750 |
, Huang et al. 1997
|
|
|
|
Rv |
AGACGCGGTAATCGAAAGATGAAA |
|
|
|
BPH |
Bph18(t) |
Bph18(t) |
SNP23 |
CGATGGATTACCCTATCACCT CAA |
HRM |
Developed in this study |
|
|
|
SNP24 |
AACCCTCTGCACACCATCGG |
|
|
|
Tungro virus |
tsv1 |
RM6152 |
Fw |
GAATTCACCGCTCTCCAGTC |
206 |
, Lee et al. 2010
|
|
|
|
Rv |
AGGAGGATCTCCTCCAGGAG |
|
|
Table 3List of control varieties used for identification of resistance genes.
Table 3
|
Gene |
Resistant controls |
Susceptible controls |
|
Xa3
|
IRBB3 |
IR24 |
|
Xa4
|
IRBB4 |
IR24 |
|
xa5
|
IRBB5 |
IR24 |
|
Xa7
|
IRBB7 |
IR24 |
|
xa13
|
IRBB13 |
IR24 |
|
Xa21
|
IRBB21 |
IR24 |
|
Pib
|
IRBL-b |
LTH |
|
Pita
|
IRBL-ta(K1), IRBL-ta(CT2), IRBL-ta2(Pi) |
LTH |
|
Pi5, Pii
|
Pi3, Pi5(t), Pii |
LTH |
|
Bph18(t)
|
Anmi, Anda |
IR24, Ilpum |
|
tsv1
|
Utri merah, N22 |
Nipponbare, TN1 |
Table 4List of eleven hybrid rice breeding lines and their cross combinations used for evaluating agronomic traits and screening disease/insect resistance in the Red River Delta in Vietnam.
Table 4
|
F1 entry |
Cross combination |
Remark |
|
KR0203H |
KR2A/Minghui 63 |
CGMS |
|
KR0695H |
KR1A/OM052 |
CGMS |
|
KR0696H |
KR2A/OM052 |
CGMS |
|
KR1454H |
KR1A/MY2R (= KR2R) |
CGMS |
|
KR1455H |
KR2A/MY2R (= KR2R) |
CGMS |
|
KR1354H |
KR2A/IR98070-KB14-1-2-3-1-1 |
CGMS |
|
KR1497H |
IR75589-31-27-8-33S/IR98229-9-2-1-k1-1 |
TGMS |
|
KR1444H |
IR75589-31-27-8-33S/IR102452-KB3-1-2-2-1-1 |
TGMS |
|
KR1487H |
HYT 108 S8-1-25-2/IR98102-kB9-1-1-3-1-1 |
TGMS |
|
KR1994H |
IR75589-31-27-8-33S/Minghui 63 |
TGMS |
|
KR2116H |
IR102100-KB12S2-1-19-1/IR98102-kB21-1-3-1-3-1 |
TGMS |
Table 5Identification patterns of disease and insect resistant genes in eleven hybrid rice combinations.
Table 5
|
Entry |
Bacterial Blight |
No. of R genes |
Blast |
No. of R genes |
Bph 18(t) |
tsv1 |
Total of R genes |
|
|
|
Xa3S |
Xa3R |
Xa4 |
xa5 |
Xa7 |
xa13 |
Xa21 |
Pi-ta |
Pib |
Pii |
Pi5 |
|
KR0203H |
+ |
− |
− |
+/− |
− |
− |
− |
0 |
+ |
− |
+ |
+ |
+ |
3 |
− |
+/− |
3 |
|
KR0695H |
− |
− |
− |
− |
− |
− |
− |
0 |
+ |
− |
+ |
+ |
+ |
3 |
− |
+/− |
3 |
|
KR0696H |
+ |
− |
− |
− |
− |
− |
− |
0 |
+ |
+ |
+ |
+ |
+ |
4 |
− |
+/− |
4 |
|
KR1454H |
+ |
− |
− |
− |
+ |
− |
− |
1 |
+ |
− |
+ |
+ |
+ |
3 |
− |
− |
4 |
|
KR1455H |
+ |
− |
− |
− |
+ |
− |
− |
1 |
+ |
− |
+ |
+ |
+ |
3 |
− |
− |
4 |
|
KR1354H |
+ |
+ |
− |
+/− |
− |
− |
− |
1 |
+ |
− |
+ |
+ |
+ |
3 |
− |
+/− |
4 |
|
KR1497H |
+ |
+ |
− |
− |
− |
− |
− |
1 |
+ |
+ |
+ |
+ |
+ |
4 |
− |
− |
5 |
|
KR1444H |
+ |
− |
− |
− |
− |
− |
− |
0 |
+ |
+ |
+ |
+ |
+ |
4 |
− |
− |
4 |
|
KR1487H |
+ |
− |
+/− |
− |
− |
− |
− |
0 |
+ |
− |
+ |
− |
+ |
2 |
− |
− |
2 |
|
KR1994H |
− |
− |
+/− |
− |
− |
− |
− |
0 |
− |
+ |
+ |
+ |
+ |
4 |
− |
− |
4 |
|
KR2116H |
+ |
+ |
+/− |
+/− |
− |
− |
− |
1 |
+ |
+ |
+ |
+ |
+ |
4 |
− |
+ |
6 |
Table 6Relative levels of resistance on hybrid rice combinations and control varieties upon inoculation separately by disease and insect in the Red River Delta region in Vietnam.
Table 6
|
Entryz)
|
Brown Planthopper |
Blast |
Bacterial Blight |
|
|
|
|
6 DARy)
|
8 DAR |
28 DAI |
35 DAIy)
|
42 DAI |
|
|
|
|
|
|
Score |
Reaction |
Score |
Reaction |
Score |
Reaction |
Score |
Reaction |
Score |
Reaction |
|
KR0203H |
6.0 |
MSx)
|
8.0 |
S |
0.0 |
HR |
1.0 |
HR |
5.0 |
MS |
|
KR0695H |
7.0 |
S |
9.0 |
HS |
0.0 |
HR |
0.0 |
HR |
1.0 |
HR |
|
KR0696H |
6.0 |
MS |
9.0 |
HS |
0.0 |
HR |
0.0 |
HR |
3.0 |
R |
|
KR1454H |
5.0 |
MS |
8.0 |
S |
0.0 |
HR |
0.0 |
HR |
3.0 |
R |
|
KR1455H |
6.0 |
MS |
8.0 |
S |
0.0 |
HR |
0.0 |
HR |
3.0 |
R |
|
KR1354H |
7.0 |
S |
8.0 |
S |
1.0 |
HR |
1.0 |
HR |
3.0 |
R |
|
KR1497H |
4.0 |
MR |
8.0 |
S |
0.0 |
HR |
0.0 |
HR |
1.0 |
HR |
|
KR1444H |
7.0 |
S |
9.0 |
HS |
0.0 |
HR |
0.0 |
HR |
3.0 |
R |
|
KR1487H |
4.0 |
MR |
9.0 |
HS |
1.5 |
HR |
2.0 |
HR |
3.0 |
R |
|
KR1994H |
3.0 |
R |
6.0 |
MS |
4.0 |
MR |
5.0 |
MS |
0.0 |
HR |
|
KR2116H |
5.0 |
MS |
9.0 |
HS |
0.0 |
HR |
1.0 |
HR |
1.0 |
HR |
|
BTP 33 |
1.0 |
R |
1.0 |
R |
- |
- |
- |
- |
- |
- |
|
TN 1 |
9.0 |
HS |
9.0 |
HS |
- |
- |
- |
- |
- |
- |
|
OM 1490 |
- |
- |
- |
- |
9.0 |
HS |
9.0 |
HS |
5.0 |
MS |
Table 7Agronomic traits and yields of F1 hybrid combinations with disease/insect resistantance genes introduced by MAS and control varieties.
Table 7
|
Entry |
HD (mm.dd) |
GP (day) |
PH (cm) |
PL (cm) |
PET (%) |
PRG (%) |
GW (g) |
Yield (tonne/ha) |
|
KR0203H |
5.18 |
121 |
114.5 |
29.4 |
64.8 |
78.1 |
27.0 |
10.1 |
|
KR0695H |
5.15 |
119 |
122.6 |
27.5 |
74.0 |
65.2 |
26.5 |
11.8 |
|
KR0696H |
5.15 |
119 |
122.9 |
27.9 |
75.3 |
71.8 |
27.0 |
11.1 |
|
KR1454H |
5.19 |
123 |
118.2 |
27.4 |
71.7 |
68.4 |
26.0 |
9.7 |
|
KR1455H |
5.18 |
122 |
114.4 |
27.4 |
72.6 |
73.8 |
26.4 |
9.6 |
|
KR1354H |
5.19 |
126 |
113.6 |
31.1 |
71.5 |
69.9 |
26.4 |
10.7 |
|
KR1497H |
5.15 |
119 |
111.6 |
28.3 |
76.2 |
88.5 |
26.2 |
10.0 |
|
KR2116H |
5.15 |
122 |
113.8 |
29.0 |
83.6 |
68.5 |
26.1 |
10.3 |
|
IIA838 |
5.12 |
113 |
118.3 |
28.1 |
79.3 |
90.4 |
30.3 |
14.6 |
|
BC15 |
5.16 |
115 |
112.8 |
28.1 |
69.8 |
74.3 |
22.1 |
9.1 |
References
- Balaji Suresh P, Srikanth B, Hemanth Kishore V, Subhakara Rao I, Vemireddy LR, Dharika N, et al. 2012. Fine mapping of Rf3 and Rf4 fertility restorer loci of WA-CMS of rice (Oryza sativa L.) and validation of the developed marker system for identification of restorer lines. Euphytica. 187: 421-435.
- Bohra A, Jha UC, Adhimoolam P, Bisht D, Singh NP. 2016. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 35: 967-993.
- Chase CD. 2007. Cytoplasmic male sterility: a window to the world of plant mitochondrial-nuclear interactions. Trends Genet. 23: 81-90.
- Cheema KK, Grewal NK, Vikal Y, Sharma R, Lore JS, Das A, et al. 2008. A novel bacterial blight resistance gene from Oryza nivara mapped to 38kb region on chromosome 4L and transferred to Oryza sativa L. Genet. Res. (Camb). 90: 397-407.
- Chen L, Liu Y. 2014. Male sterility and fertility restoration in crops. Annu Rev Plant Bio. 65: 579-606.
- Chen X, Temnykh S, Xu Y, Cho YG, McCouch SR. 1997. Development of a microsatellite framework map providing genomewide coverage in rice (Oryza sativa L.). Theor Appl Genet. 95: 553-567.
- Dyah RP, Kei M, Bambang HT. 2013. The dynamics of rice production in Indonesia 1961–2009. J Saudi Soc Agric Sci. 12: 27-37.
- FAORAP.2014. Hybrid rice development in Asia: Assessment of limitations and potential.
- Franz MN, Song JY, Ouk S, Rahimi S, Kwon SW, Kang KK, et al. 2016. Current applicable DNA markers for marker assisted breeding in abiotic and biotic stress tolerance in rice (Oryza sativa L.). Plant Breed Biotech. 4: 271-284.
- Fujita D, Kohli A, Horgan FG. 2013. Rice resistance to planthoppers and leafhoppers. Crit Rev Plant Sci. 32: 162-191.
- He GH, Wang WM, Liu GQ, Hou L, Xiao YH, Tang M, et al. 2002. Mapping of two fertility restoring gene for WA cytoplasmic male mterility in Minghui63 using SSR markers. Acta Genet Sin. 29: 798-802.
- Huang N, Angeles ER, Domingo J, Magpantay G, Singh S, Zhang G, et al. 1997. Pyramiding of bacterial blight resistance genes in rice: marker assisted selection using RFLP and PCR. Theor Appl Genet. 95: 313-320.
- Hu J, Xiao C, He Y. 2016. Recent progress on the genetics and molecular breeding of brown planthopper resistance in rice. Rice (N Y). 9: 30
- Hu YJ, Jeung JU, Kim SY, Park HS, Cho JH, Lee JY. 2013. Functional markers for bacterial blight resistance gene Xa3 in rice. Mol Breed. 31: 981-985.
- Ikeda-Kawakatsu K, Maekawa M, Izawa T, Itoh J, Nagato Y. 2012. ABERRANT PANICLE ORGANIZATION 2/RFL, the rice ortholog of Arabidopsis LEAFY, suppresses the transition from inflorescence meristem to floral meristem through interaction with APO1. Plant J. 69: 168-180.
- Jeon JS, Chen D, Yi GH, Wang GL, Ronald PC. 2003. Genetic and physical mapping of Pi5(t), a locus associated with broad-spectrum resistance to rice blast. Mol. Genet. Genomics. 269: 280-289.
- Jia Y, Wang Z, Singh P. 2002. Development of dominant rice blast Pi-ta resistance gene markers. Crop Sci. 42: 2145-2149.
- Jia Y, Redus M, Wang Z, Rutger JN. 2004. Development of a SNLP marker from the Pi-ta blast resistance gene by tri-primer PCR. Euphytica. 138: 97-105.
- Jin XW, Wang CL, Yang Q, Jiang QX, Fan YL, Liu GC, et al. 2007. Breeding of near-isogenic line CBB30 and molecular mapping of Xa30(t), a new resistance gene to bacterial blight in rice. Sci Agric Sin. 40: 1094-1100.
- Jing RC, Li XM, Yi P, Zhu YG. 2001. Mapping fertility-restoring genes of rice WA cytoplasmic male sterility using SSLP markers. Bot Bull Acad Sin. 42: 167-171.
- Jo YC, Kim SG. 2016. Efficient F1 variety development using CGMS system in onion (Allium cepa L.). Trends Agric Life Sci. 53: 81-85.
- Kim KY, Shin MS, Kim WJ, Mo YJ, Nam JK, Noh TH, et al. 2009. Effective combination of resistance genes against rice bacterial blight pathogen. Korean J Breed Sci. 41: 244-251.
- Kwon SW, Cho YC, Kim YG, Suh JP, Jeung JU, Roh JH. 2008. Development of near-isogenic Japonica rice lines with enhanced resistance to Magnaporthe grisea. Mol Cells. 25: 407-416.
- Lau WC, Rafii MY, Ismail MR, Puteh A, Latif MA, Ramli A. 2015. Review of functional markers for improving cooking, eating, and the nutritional qualities of rice. Front Plant Sci. 6: 832
- Lee JD, Do JW, Han JH, An CG, Kweon OY, Kim YK, et al. 2011. Allelism and molecular marker tests for genic male sterility in paprika cultivars. Korean J Hortic Sci Technol. 29: 130-134.
- Lee JH, Muhsin M, Atienza GA, Kwak DY, Kim SM, Leon TB. 2010. Single nucleotide polymorphisms in a gene for translation initiation factor (eIF4G) of rice (Oryza sativa) associated with resistance to Rice tungro spherical virus. Mol Plant Microbe Interact. 23: 29-38.
- Li S, Yang D, Zhu Y. 2007. Characterization and use of male sterility in hybrid rice breeding. J Integr Plant Biol. 49: 791-804.
- Lin SC, Yuan LP. 1980. Hybrid rice breeding in China. pp. 35-51. International Rice Research Institute.Innovative approaches to rice breeding. International Rice Research Institute. Manila, Philippines.
- Liu J, Wang X, Mitchell T, Hu Y, Liu X, Dai L, et al. 2010. Recent progress and understanding of the molecular mechanisms of the rice-Magnaporthe oryzae interaction. Mol Plant Pathol. 11: 419-427.
- McCouch SR, Teytelman L, Xu YB, Lobos KB, Clare K, Walton M. 2002. Development and mapping of 2,240 new SSR markers for rice (Oryza sativa L.). DNA Res. 9: 199-207.
- Myint KKM, Fujita D, Matsumura M, Sonoda T, Yoshimura A, Yasui H. 2012. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stål]) in the rice cultivar ADR52. Theor Appl Genet. 124: 495-504.
- Ouyang YD, Chen JJ, Ding JH, Zhang QF. 2009. Advances in the understanding of inter-subspecific hybrid sterility and wide compatibility in rice. Sci Bull. 54: 2332-2341.
- Park HS, Kim KY, Shin MS, Noh TH, Ha KY, Baek MG, et al. 2011. Breeding for japonica rice variety with resistance to bacterial blight in Korea. Korean J Breed Sci. 43: 44.
- Peleman JD, Voort JR. 2003. Breeding by design. Trends Plant Sci. 8: 330-334.
- Porter BW, Chittoor JM, Yano M, Sasaki T. 2003. Development and mapping of markers linked to the rice bacterial blight resistance gene Xa7. Crop Sci. 43: 1484-1492.
- Seo HS, Song YC. 1993. Breeding hybrid rice with good quality and high yield: I. Breeding of cytoplasmic-genetic male sterile and restorer lines with backgrounds of Korean rice varieties. Korean J Crop Sci. 38: 413-417.
- Shin MS, Kim KY, Park HS, Ko JK. 2011. Breeding for resistance to bacterial blight in rice. Korean J Breed Sci. 43: 241-251.
- Singh AK, Gopala Krishnan S, Singh VP, Prabhu KV, Mohapatra T, Singh NK. 2011. Marker assisted selection: a paradigm shift in Basmati breeding. Indian J Genet Pl Br. 71: 1-9.
- Song JY, Ouk S, Nogoy FM, Niño MC, Kwon SW, Ha WG, et al. 2016. Application and utilization of marker assisted selection for biotic stress resistance in hybrid rice (Oryza sativa L.). J Plant Biotechnol. 43: 317-331.
- Suh JP, Cho YC, Won YJ, Ahn EK, Baek MK, Kim MK, et al. 2015. Development of Resistant Gene-Pyramided Japonica Rice for Multiple Biotic Stresses Using Molecular Marker-Assisted Selection. Plant Breed Biotech. 3: 333-345.
- Sundaram RM, Sakthivel K, Hariprasad AS, Ramesha MS, Viraktamath BC, Neeraja CN, et al. 2010. Development and validation of a PCR-based functional marker system for the major wide-compatible gene locus S5 in rice. Mol Breed. 26: 719-727.
- Suresh PB, Srikanth B, Kishore VH, Rao IS, Vemireddy LR, Dharika N, et al. 2010. Fine mapping of Rf3 and Rf4 fertility restorer loci of WA-CMS of rice (Oryza sativa L.) and validation of the developed marker system for identification of restorer lines. Euphytica. 187: 421-435.
- Tang H, Luo D, Zhou D, Zhang Q, Tian D, Zheng X, et al. 2014. The ricer restorer Rf4 for wild-abortive cytoplasmic male sterility encodes a mitochondrial-localized PPR protein that functions in reduction of WA352 transcripts. Mol Plant. 7: 1-4.
- Thome K, Birgit M, Kamron D, Cheryl C. 2018. International Food Security Assessment, 2018–2028, GFA-29. Departmetn of Agriculture, Economic Research Service. U.S.
- Tu J, Ona I, Zhang Q, Mew TW, Khush GS, Datta SK. 1998. Transgenic rice variety ‘IR72’ with Xa21 is resistant to bacterial blight. Theor Appl Genet. 97: 31-36.
- Viet NV. 2008. National agricultural research system of Vietnam. Vietnam Academy of Agricultural Sciences. Vietnam.
- Xia C, Chen H, Zhu X. 2012. Identification, mapping, isolation of the genes resisting to bacterial blight and application in rice. Mol Plant Breed. 3: 121-131.
- Yang Q, Lin F, Wang L, Pan Q. 2009. Identification and mapping of Pi41, a major gene conferring resistance to rice blast in the Oryza sativa subsp. indica reference cultivar, 93-11. Theor Appl Genet. 118: 1027-1034.
- Zhang F, Zhuo DL, Huang LY, Wang WS, Xu JL, Vera Cruz C, et al. 2015. Xa39, a novel dominant gene conferring broad-spectrum resistance to Xanthomonas oryzae pv. oryzae in rice. Plant Pathol. 64: 568-575.
- Zhang G, Angeles ER, Abenes MLP, Khush GS. 1996. RAPD and RFLP mapping of the bacterial blight resistance gene Xa-13 in rice. Theor Appl Genet. 93: 65-70.