Abstract
×Brassicoraphanus (AARR, 2n = 38) is a synthetic intergeneric allopolyploid between Brassica rapa L. ssp. pekinensis (AA, 2n = 20) and Raphanus sativus L. var. rafiphera (RR, 2n = 18). Abnormalities in meiosis are main causes for infertility, especially in recent intergeneric allopolyploids. Several ×Brassicoraphanus lines showing varied fertility rates were produced previously, but no cytogenetic data specifying the reasons for infertility have been reported. In this study, we performed cytogenetic analyses in BB4, BB6, BB12, and BB50 lines to evaluate their chromosomal composition and behavior during meiosis. The four lines had relatively small chromosomes, ranging in length from 1.82 to 5.72 μm. BB6, BB12, and BB50 have euploid chromosome number of 2n = 38, whereas BB4 is an aneuploid with 2n − 1 = 37. Fluorescent in situ hybridization karyotype analysis by using 5S/45S rDNA revealed 5/7, 6/7, 5/5 and 5/5 pairs in BB4, BB6, BB12 and BB50, respectively. Genomic in situ hybridization analysis on cells in prophase I revealed varying frequencies of tetravalent pairing and sticky, ring, rod, and laggard chromosomes across the lines, which were more abundant in BB4 and BB6. Unlike BB4 and BB6, both BB12 and BB50 are known to have relatively higher seed fertility and uniform plant morphology. The varied degrees of chromosomal pairing stability during meiosis could explain the different fertility rates among the four ×Brassicoraphanus lines in this study. These data might facilitate breeding programs of ×Brassicoraphanus and further cytogenomic analyses.
-
Key words: Intergeneric hybrid, ×Brassicoraphanus, Cytogenetic study
INTRODUCTION
In recent years, breeding of
Brassica and
Raphanus species has received considerable attention (
Namai et al. 1980;
Prakash et al. 2009) because of their agricultural importance worldwide as major crops as fodders, edible vegetables, and functional compounds sources (
Lee et al. 2017).
Brassica rapa (Chinese cabbage, 2
n = 20) and
Raphanus sativus (big root radish, 2
n = 18) produce economically important shoot and root parts, respectively, especially in eastern Asia, where they are the main ingredients of staple foods like kimchi (
Kim et al. 2000). While either shoot or root are utilized in either
B. rapa and
R. sativus, breeding for a species with combined shoot and root features of both species would optimize utility of plant parts. Thus, previous breeding efforts have initiated this program (
Lee et al. 2002).
Morphological and cytological methods have been used to select synthetic hybrid species between the two genomes of
Brassica and
Raphanus (
Dolstra 1982;
Kato and Tokumasu 1983). A newly synthesized intergeneric allotetraploid, ×
Brassicoraphanus, was developed by crossing
B. rapa L. ssp.
pekinensis and
R. sativus (
Lee et al. 1999;
Lee et al. 2002;
Lee et al. 2017); however it was found to be unstable with low seed fertility and uniformity. The progenies from these unstable materials were stabilized using microspore mutagenesis by using
N-nitroso-
N-methyl urethane (NMU) (
Lee et al. 2011;
Lee et al. 2017). The same procedure involving the use of NMU was applied to develop another allotetraploid hybrid between
B. rapa L. ssp.
pekinensis and
R. sativus L. var.
rafiphera, and the F1 hybrid was crossed with a mutagen-induced stabilized line to produce different lines of ×
Brassicoraphanus.
Recently, a FISH karyotype analysis of ×
Brassicoraphanus line BB#5 (
Belandres et al. 2015) and a conventional Giemsa-staining karyotype analysis of line BB4 were conducted (
Lim et al. 2012), however no cytogenetic data on the developed hybrid lines is available. In this study, we analyzed the chromosomal distribution and localization of 5S and 45S rDNA on four synthetic hybrid lines and constructed a dual-color FISH karyotype with both markers to identify the chromosome complement of each inbred line. Moreover, meiotic chromosome pairing patterns were analyzed to discriminate both parental genomes,
Brassica and
Raphanus, by using GISH method.
MATERIALS AND METHODS
Plant materials
The floral buds and seeds of the ×Brassicoraphanus lines BB4, BB6, BB12, and BB50 were kindly provided by Soo-Seong Lee who developed the intergeneric hybrids in the Bio-Breeding Institute, Ansung, Korea. The floral buds were collected from the adult plants of each line and the seeds were sent for this experiment after harvesting from each line. The seeds were germinated in Petri dishes with moist filter paper at 25°C for 48 hours. Root tips (length, 2 cm) were harvested and pretreated with 2 mM 8-hydroxyquinoline at 18°C for 5 hours, fixed in aceto-ethanol (1:3 v/v) solution for 2 to 24 hours, and then stored in 70% ethanol until use. Floral buds were fixed in the same solution for 24 hours and stored in 70% ethanol at −20°C.
Chromosome and probe preparation for FISH and GISH
Mitotic and meiotic chromosomes, rDNA probes, FISH and GISH procedures were done following the same procedures of previous study (
Belandres et al. 2015).
Homologous chromosomes were identified based on the FISH karyotype of ×
Brassicoraphanus line BB5 (
Belandres et al. 2015). Meiotic pairing configurations were analyzed in late prophase I. The varying degree of meiotic abnormalities were examined in 500 pollen mother cells (PMCs).
RESULTS
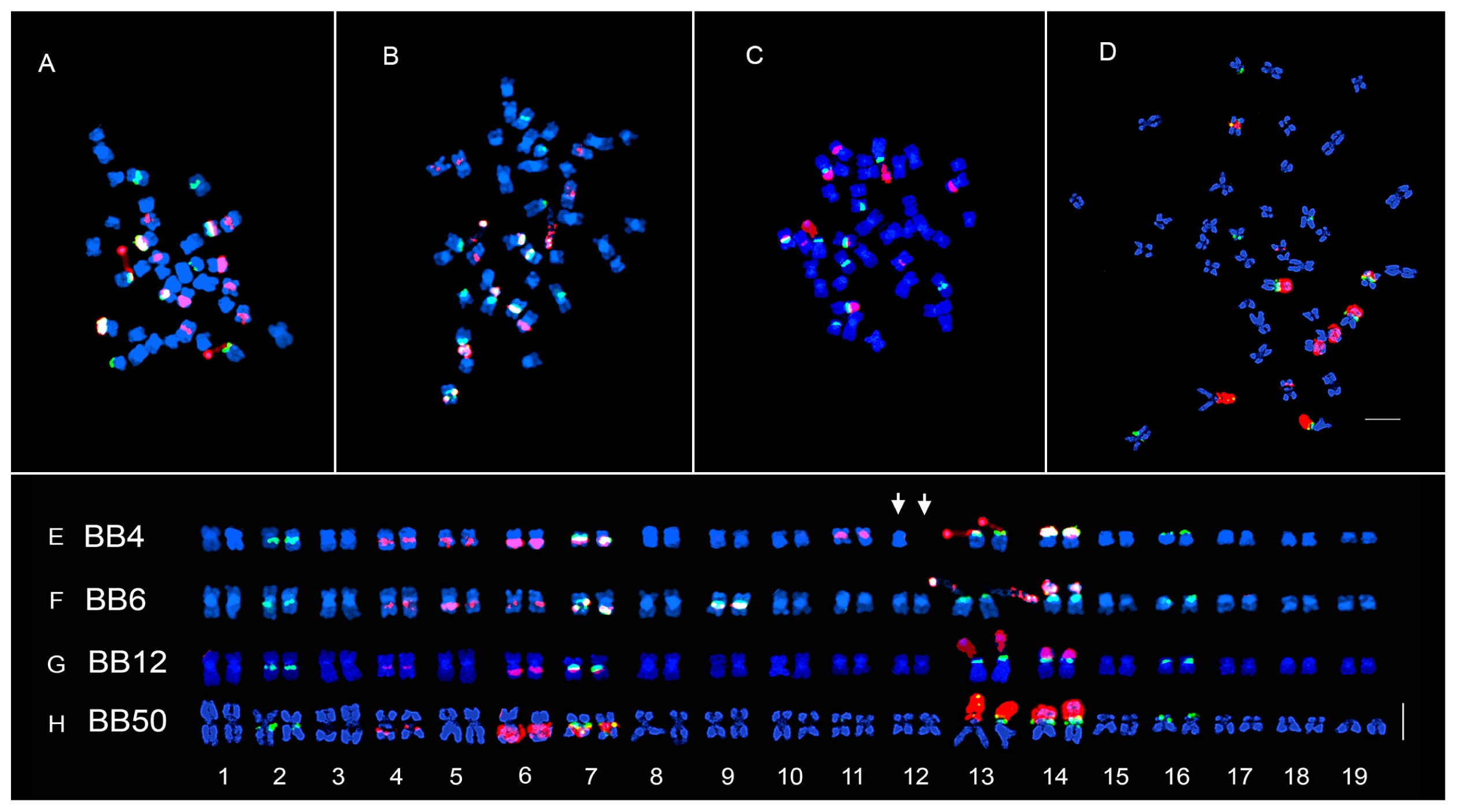
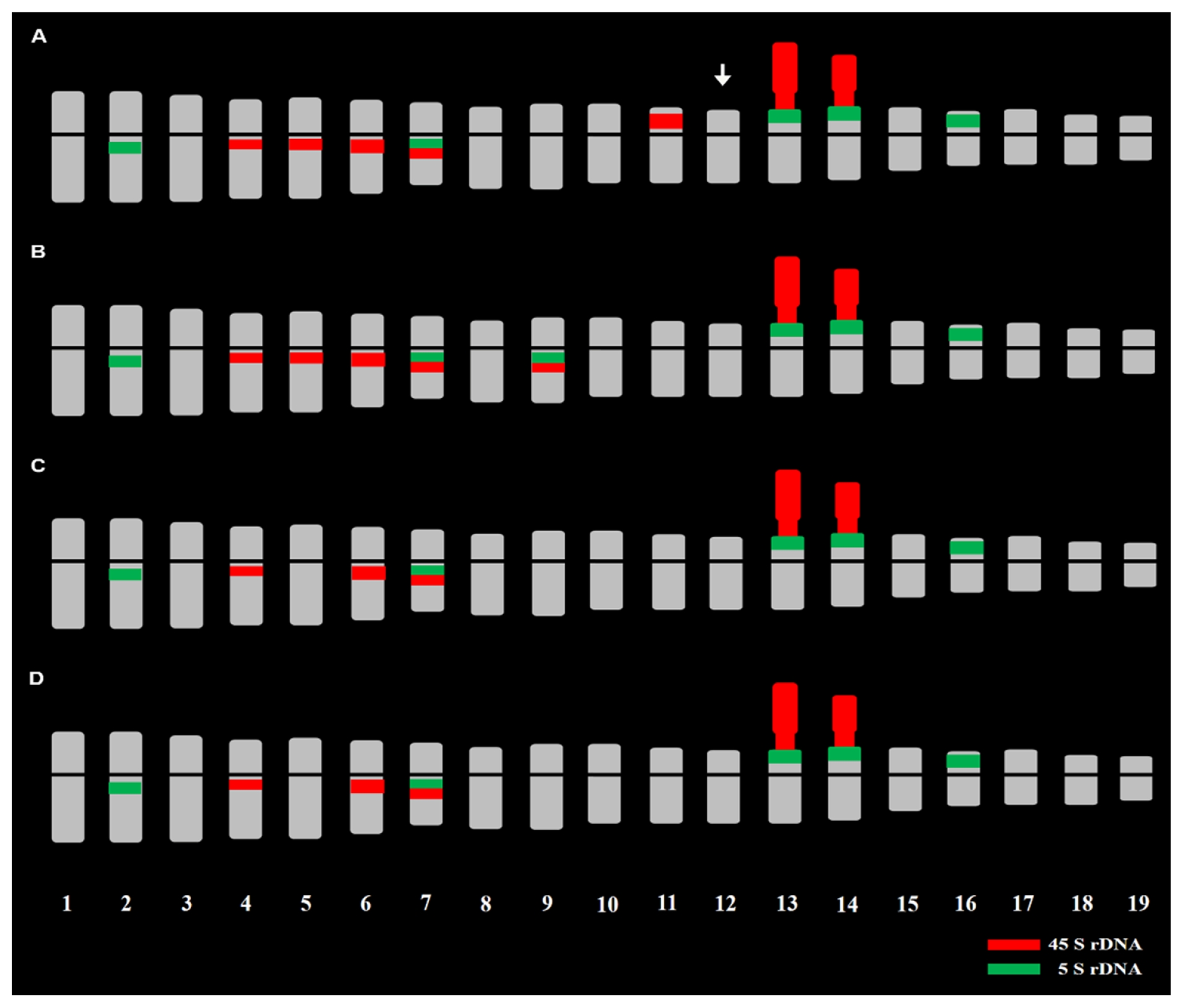
rDNA repeats
The fluorescence
in situ hybridization (FISH) karyotype of ×
Brassicoraphanus lines with 5S/45S rDNA revealed 5/7, 6/7, 5/5, and 5/5 pairs in BB4, BB6, BB12, and BB50, respectively (
Figs. 1 and
2,
Table 1). The 5S rDNA loci were observed in the para-centromeric region of the long arms of chromosomes 2 and 7, terminal part of the short arms of chromosome 13 and 14 being juxtaposed to the satellite chromosome, and on the terminal part of the short arm of chromosome 16. However, in BB6, another pair of 5S rDNA loci was observed in the para-centromeric region of the long arm of chromosome 9. The 45S rDNA loci pattern of BB12 and BB50 was identical, whereas two more pairs were observed for BB4 and BB6 (
Figs. 1 and
2,
Table 1). The 45S rDNA loci were observed at the paracentromeric region of the long arms of chromosomes 4, 6, and 7 and on the nuclear organizer region (NOR) regions of chromosome 13 and 14. However, two additional pairs of 45S rDNA loci were observed in the para-centromeric region of the long arms of chromosome 5 and 9 in BB6 and the para-centromeric region of the long arms of chromosome 5 and near the short arm of chromosome 11 in BB4.
When compared with the FISH karyotype of the stable line BB5, some of the 45S rDNA loci were absent, particularly in chromosome 8 of BB4, chromosomes 8 and 11 of BB6, and chromosomes 5, 8, and 11 of BB12 and BB50. However, the five pairs of 5S rDNA loci were all present in the four lines. In addition, a distinct pair of co-localized 5S and 45S rDNA loci was observed only in chromosome 9 of BB6, which was even absent in BB5.
FISH karyotype
BB4 was aneuploidy with a somatic chromosome number of 2
n − 1 = 37 (
Fig. 1A, E), with lengths ranging from 1.82 to 3.45 μm and a total length of 49.64 μm (
Table 1). The chromosome complement comprised fifteen metacentric and four submetacentric chromosomes. Chromosome 12 of BB4 has a missing copy of its homolog, resulting in a low total chromosome length compared to that for the other lines (
Fig. 1E, white arrows). BB6 somatic chromosome number was 2n = 38 (
Fig. 1B and F), with lengths ranging from 2.31 to 4.43 μm and a total length of 61.39 μm (
Table 1). The chromosome complement comprised fifteen metacentric and four submetacentric chromosomes. BB12 somatic chromosome number was 2
n = 38, (
Fig. 1C and G), with chromosome lengths ranging from 2.58 to 4.78 μm and a total length of 68.02 μm (
Table 1). The chromosome complement comprised eighteen metacentric and one submetacentric chromosomes. BB50 chromosome number was 2
n = 38 (
Fig. 1D and H), with lengths ranging from 2.40 to 5.72 μm and a total length of 74.34 μm (
Table 1). The chromosome complement comprised twelve metacentric and seven submetacentric chromosomes (
Fig. 2).
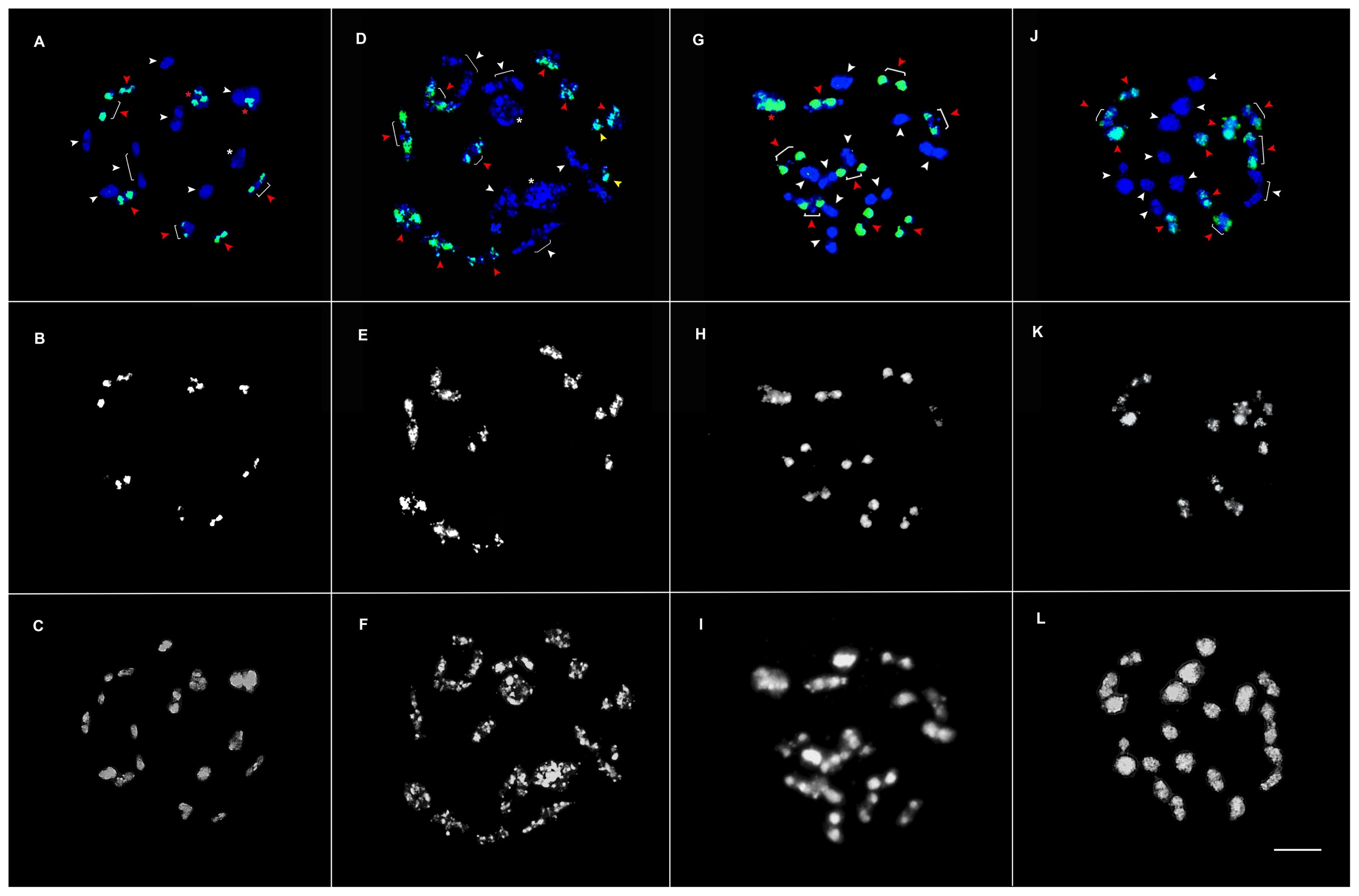
GISH analysis was done to identify chromosomes from each parent,
B. rapa (
Fig. 3B–K, raw) and
R. sativus (
Fig. 3A–J, DAPI-blue) on the PMCs at prophase I of meiosis in the four lines (
Fig. 3). In meiotic pairing, nineteen bivalent chromosome pairs were observed in BB50 (
Fig. 3J–L), consisting of ten and nine bivalent chromosome pairs of
B. rapa (
Fig. 3J, red arrowheads, K-raw) and
R. sativus, respectively (
Fig. 3J, white arrowheads), whereas the rest of the lines BB4, BB6, and BB12 consisted of combined bivalent and tetravalent chromosome pairs from both the genomes (
Fig. 3A–I). For instance, BB4 has six bivalent (
Fig. 3A, red arrowheads) and two tetravalent chromosome pairs of
B. rapa (
Fig. 3A, red asterisk, B-raw), and seven bivalent (
Fig. 3A, white arrowheads) and one tetravalent chromosome pairs from
R. sativus (
Fig. 3A, white asterisk); BB6 has nine bivalent (
Fig. 3D, red arrowheads) and two univalent chromosome pairs of
B. rapa (
Fig. 3D, yellow arrowheads, E-raw) and five bivalent (
Fig. 3D, white arrowheads) and two tetravalent chromosome pairs from
R. sativus (
Fig. 3D, white asterisk); BB12 has eight bivalent (
Fig. 3G, red arrowheads) and one tetravalent chromosome pairs from
B. rapa (
Fig. 3G, red asterisk, H-raw) and nine bivalent chromosome pairs of
R. sativus (
Fig. 3G, white arrowheads).
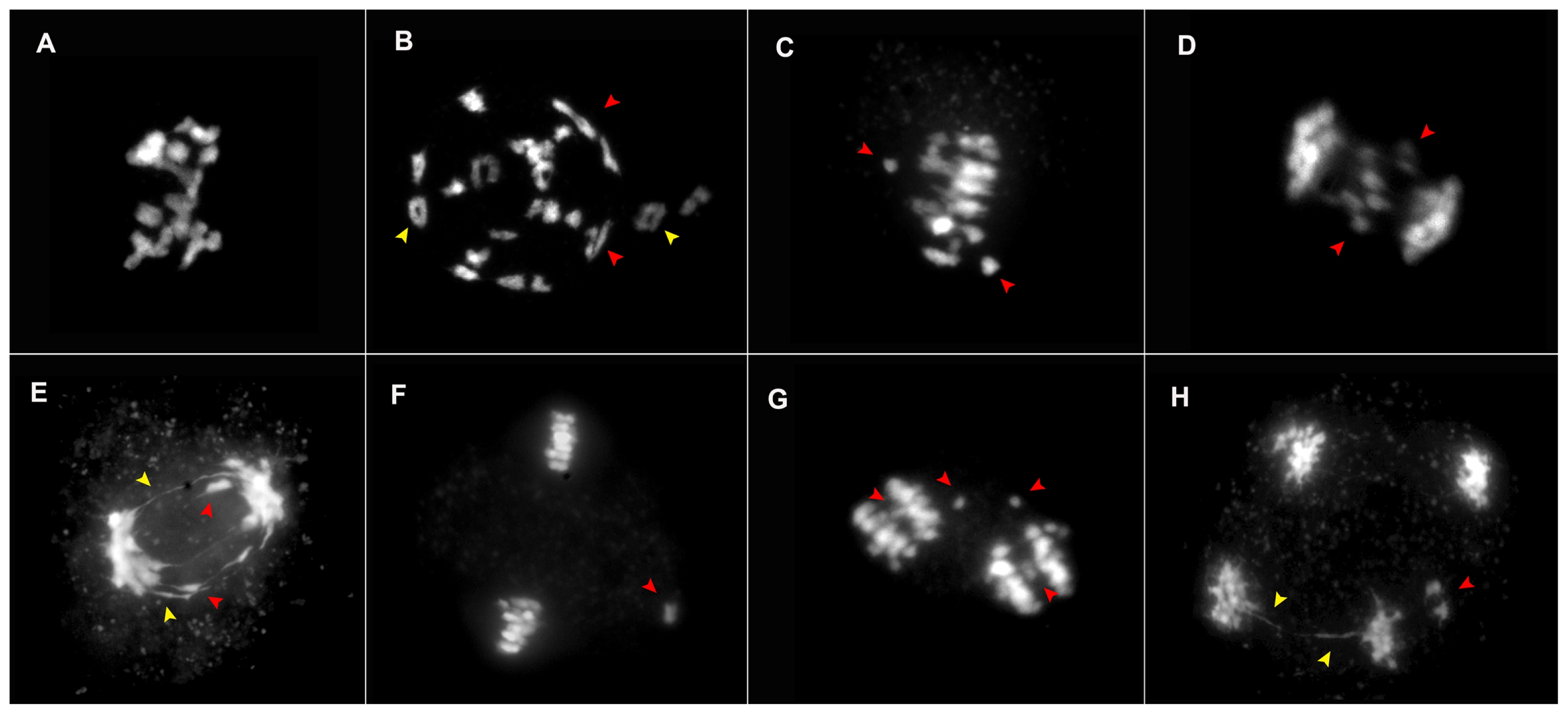
Meiotic abnormalities were noted in all lines from different stages of meiosis, although all the lines were not equally affected. The irregularities observed include chromosome stickiness (
Fig. 4A), rings (
Fig. 4B, yellow arrowheads), rods (
Fig. 4B, red arrowheads), laggard chromosomes (
Fig. 4C–G, red arrowheads), and bridge fragmentation (
Fig. 4E, yellow arrowheads). In addition, micronuclei (
Fig. 4H, red arrowheads) were identified in telophase II due to acentric chromosome fragments or lagging chromosomes that were not incorporated into one of the daughter nuclei during cell division. The unstable lines BB4 and BB6 showed high frequencies of meiotic abnormalities, which accounted for 57% (287) and 34% (169), respectively, compared to 12% (60) and below in the stable lines BB50 and BB12. High degree of laggard chromosomes in metaphase I was observed, which accounted for 31% (154), followed by bridge fragmentation in anaphase I for 10% (49) in BB4, whereas laggards in metaphase I and II accounted for 13% (65) and 7% (36) in BB6, examined using 500 PMCs (
Table 2).
DISCUSSION
Intergeneric allopolyploids between
Brassica and
Raphanus have remained unstable despite numerous attempts to improve their chromosomal pairing stability. However,
Lee et al. (2011) successfully developed stable progenies, ×
Brassicoraphanus, through induced mutation by using microspore culture, which became a commercial vegetable crop. In this study, we assessed the somatic metaphase chromosome karyotype by fluorescence
in situ hybridization (FISH), meiotic pairing behavior by genomic
in situ hybridization (GISH) and meiotic abnormalities in four lines of ×
Brassicoraphanus.
In this study, we observed varying numbers of 5S and 45S rDNA loci within and among the several lines of ×
Brassicoraphanus. However, two pairs of nucleolar organizers of
Brassica and
Raphanus were present in all lines. These factors might have contributed to the concerted changes in multiple tandem repeats (
Eickbush and Eickbush 2007), in which the common mechanisms involve unequal crossing-over and gene conversion (
Dover 1989;
Shaked et al. 2001). In addition, these changes in rDNA distribution can be attributed to genomic rearrangements that include rDNA loci exchanges, fragment loss, and homoeologous exchange, as reported previously (
Pontes et al. 2004;
Gaeta et al. 2007;
Lim et al. 2008). Major karyotypic changes such as aneuploidy can result in alterations and irregularities in somatic and meiotic cell division. Meiotic mechanisms inducing aneuploidy might include non-homologous recombination, asynapsis, loss of recombination and chromosome segregation defects in meiosis (
Cai and Xu 2007;
Ravi et al. 2011). In general, allopolyploids show genetic and phenotypic changes because of chromosomal rearrangement, gene dosage effects, unbalanced parental genome contributions, existence of different genomes in the nucleus, and variations in chromosome number (
Leitch and Leitch 2008). In a synthetic hybrid, several chromosomal variations and abnormal meiosis were observed, implying that the genetic instability of the hybrid polyploid was due to abnormal chromosome number and structure, which were attributed to aberrant meiosis (
Fujii and Ohmido 2011). In the present study, the unstable line BB4 was aneuploid with 2
n −1 = 37, showing a high degree of meiotic irregularities and nondisjunction and anaphase chromosome lagging could be the two mechanisms responsible for chromosome loss or aneuploidy. The nuclear envelope was formed with a laggard might form a micronucleus.
Brassicaceae interspecific hybrid subgenomes were successfully identified using GISH method (
Wang et al. 2006;
Yao et al. 2010;
Howell and Armstrong 2013). Moreover, the parental subgenomes,
Brassica and
Raphanus, were clearly discriminated in somatic metaphase (
Lim et al. 2012) and meiotic chromosomes (
Belandres et al. 2015) of the hybrid ×
Brassicoraphanus. In this study, we discriminated ten and nine pairs of
Brassica and
Raphanus bivalent chromosomes in BB50, and the remaining lines BB4, BB6, and BB12 consisted of the combined univalent, bivalent, and tetravalent chromosome pairs of both parental sub-genomes. Genomic reports showed that a high degree of synteny is shared between
B. rapa and
R. sativus genomes (
Kitashiba et al. 2014;
Moghe and Shiu 2014). This might imply that multivalent chromosome pairing between
B. rapa and
R. sativus chromosomes might be attributed to the possible pairing of nonhomologous chromosomes to the homeologous region at the early stage of meiosis. In addition, multivalent formation can be generated because of chromosomal changes due to abnormal segregation in meiosis (
Comai 2005;
Leitch and Leitch 2008;
Fujii and Ohmido 2011), which could increase the chance of chromosomal rearrangement such as translocation and deletion over two nonhomologous chromosomes during segment exchange, leading to genome instability and infertility of hybrid allopolyploid plants.
Different levels of meiotic irregularities are possible in allopolyploid because of segmental interchange during homeologous recombination influenced by the progenitor genotype and/or their combination (
Szadkowski et al. 2010;
Grandont et al. 2014). We observed several chromosomal abnormalities in different stages of meiosis. Chromosome stickiness is characterized by an intense clumping of chromosomes during any phase of the cell cycle (
Rao et al. 1990;
Kiihl et al. 2011). This might have been caused by environmental factors such as ionizing radiation, temperature, and soil elements (
Mendes-Bonato et al. 2001). Ring chromosomes might form in cells following genetic damage by mutagens where both ends of the chromosome were often missed, enabling the broken ends to fuse together to form a ring. Laggard chromosomes may either be excluded or rejoined forming daughter nuclei later than the other chromosomes (
De Storme and Mason 2014). This is common in hybrids and new allopolyploids might go through unequal interaction of centromeres and the loss of centromeric histone H3 (CenH3) (
Ravi and Chan 2010), causing subsequent chromosome loss and uniparental genome elimination (
Sanei et al. 2011). In some cases, incorrect chromosome segregation can lead to bridge and bridge fragmentations (
Kiihl et al. 2011) and can be attributed to chromatin breakage and fusion or breakage due to the failure of chiasma termination that leads to subsequent lagging at anaphase I (
Saylor and Smith 1966).
Advanced molecular cytogenetic studies including FISH and GISH using chromosome-specific DNA markers and comparative studies related to genome sequencing are necessary to elucidate more accurate genome structures of the hybrid lines. Our results might be useful for further genomic study of ×Brassicoraphanus lines and their improvement as new promising breeding varieties.
ACKNOWLEDGEMENTS
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through the Agri- Bio Industry Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA; 117045-3).
Fig. 1Dual color FISH metaphase of the four inbred lines of ×Brassicoraphanus. Three mitotic metaphase spreads of lines BB6 (B, F), BB12 (C, G), BB50 (D, H) showing chromosome number 2n = 38, and BB4 (A, E, arrow), 2n − 1 = 37. The 5S and 45S rDNA loci are shown as green and red signals, respectively. FISH metaphase chromosomes were arranged in decreasing order according to the length and morphology of each chromosome (E–H). Line BB4 has an incomplete chromosome number indicated by an arrow (E). Scale bar, 5 μm.
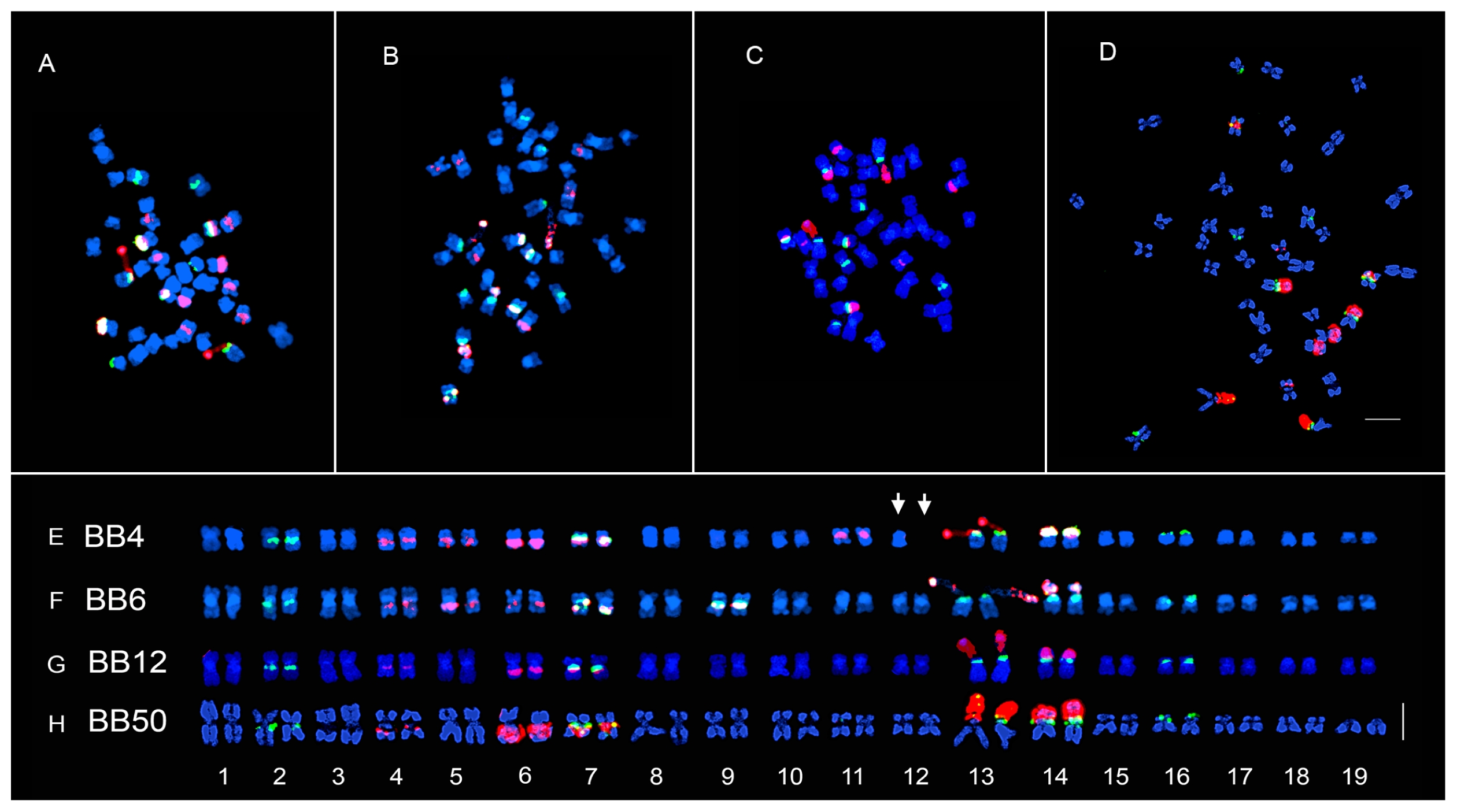
Fig. 2FISH karyotypic idiograms showing chromosome number, length, morphology, and 5s and 45s rDNAs signals in ×
Brassicoraphanus inbred lines, BB4 (2n − 1 = 37, A) with missing chromosomes indicated by an arrow, BB6 (2n = 38, B), BB12 (2n = 38, C), and BB50 (2n = 38, D). The 5S and 45S rDNA loci are shown as green and red signals, respectively. The chromosomes were arranged according to the length, morphology, and rDNAs distribution described by
Belandres et al. (2015).
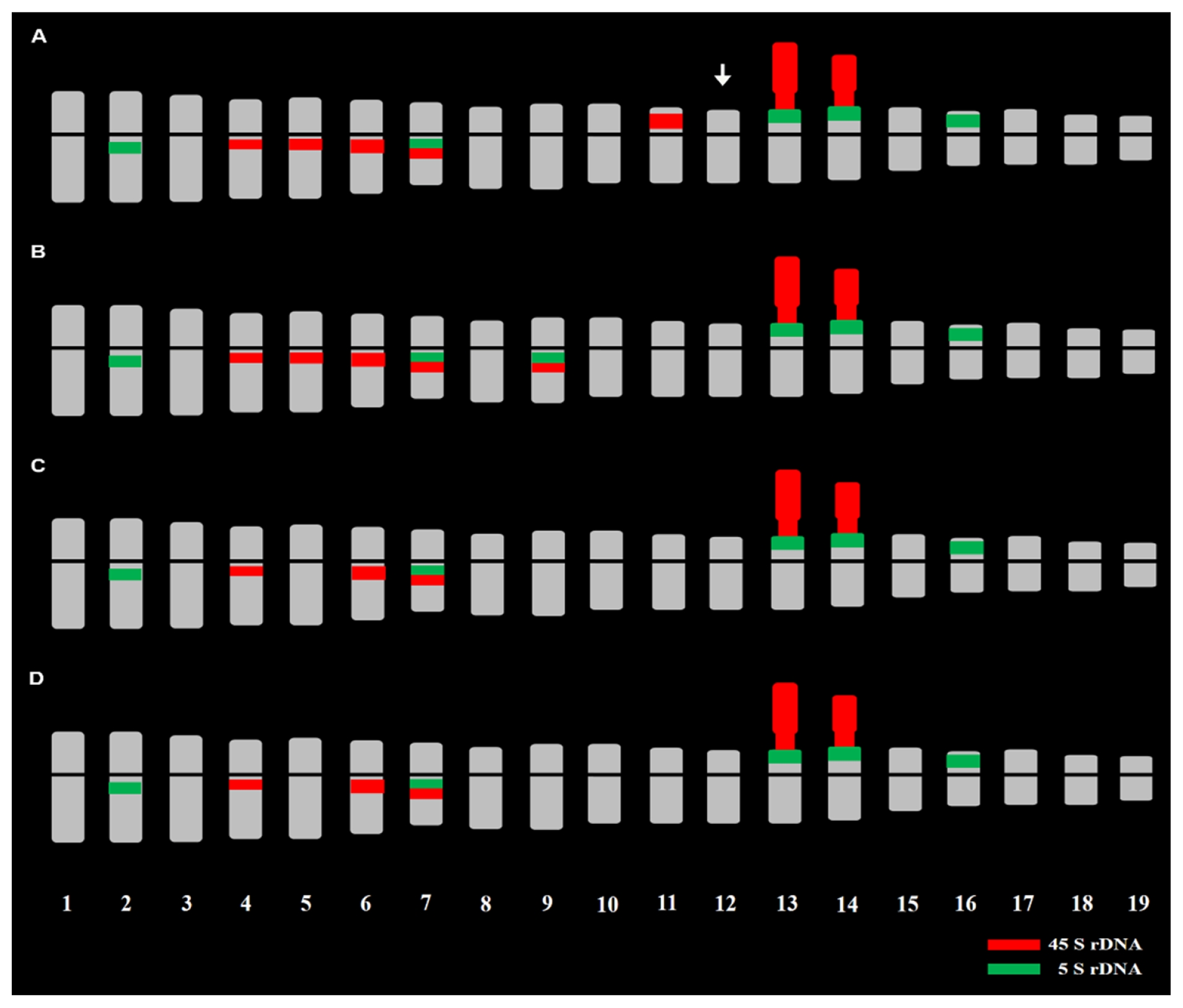
Fig. 3Meiotic chromosome pairing in prophase I of ×Brassicoraphanus inbred lines BB4 (A–C), BB6 (D–F), BB12 (G–I), and BB50 (J–L). Nineteen complete bivalents were observed in BB50, as shown in (J), 10 of which are of B. rapa (green fluorescence, K- raw) and nine are of R. sativus chromosomes (DAPI, blue), whereas the remaining lines show multivalent and univalent chromosome pairing. In the panels above, white arrowheads and white asterisks indicate R. sativus chromosomes; red arrowheads, red asterisks, and the raw images in the middle panels are of B. rapa; yellow arrowheads indicate univalent chromosomes; and asterisks (red and white) indicate tetravalent chromosomes. The bottom panels are raw DAPI images of the four lines. Scale bar, 5 μm.
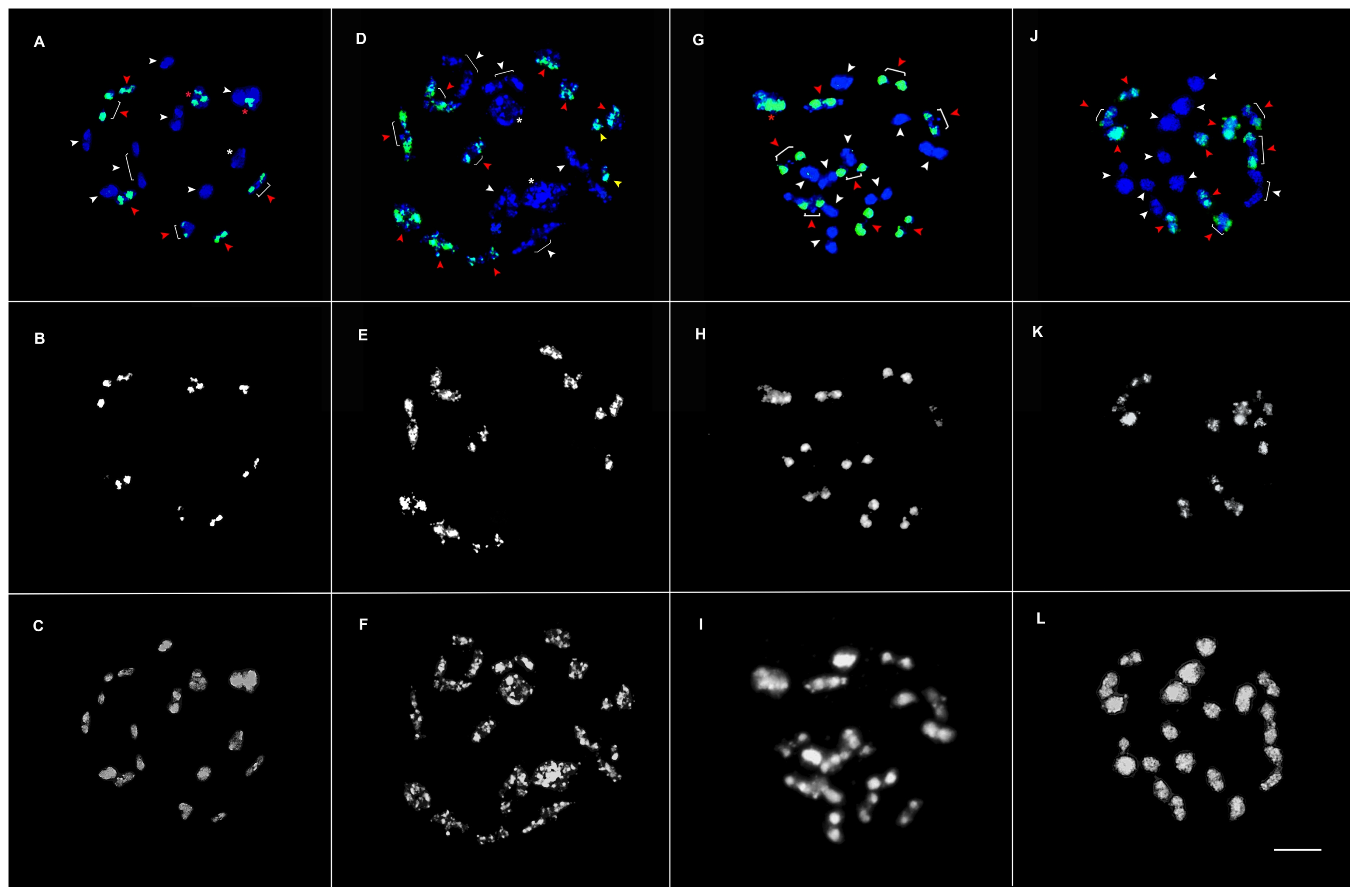
Fig. 4Meiotic abnormalities observed in the four lines of ×Brassicoraphanus. (A–H) DAPI raw images of abnormal cells observed including sticky chromosomes (A), ring (yellow arrows) and rod (red arrows) (B), lagging chromosomes in metaphase I/II and anaphase I/II (C–G, red arrows), bridge fragmentation in anaphase I and telophase II (E, H, yellow arrows) with two micronuclei (H, red arrow), all of which contributes to the characteristic increase in aneuploidy and sterility.
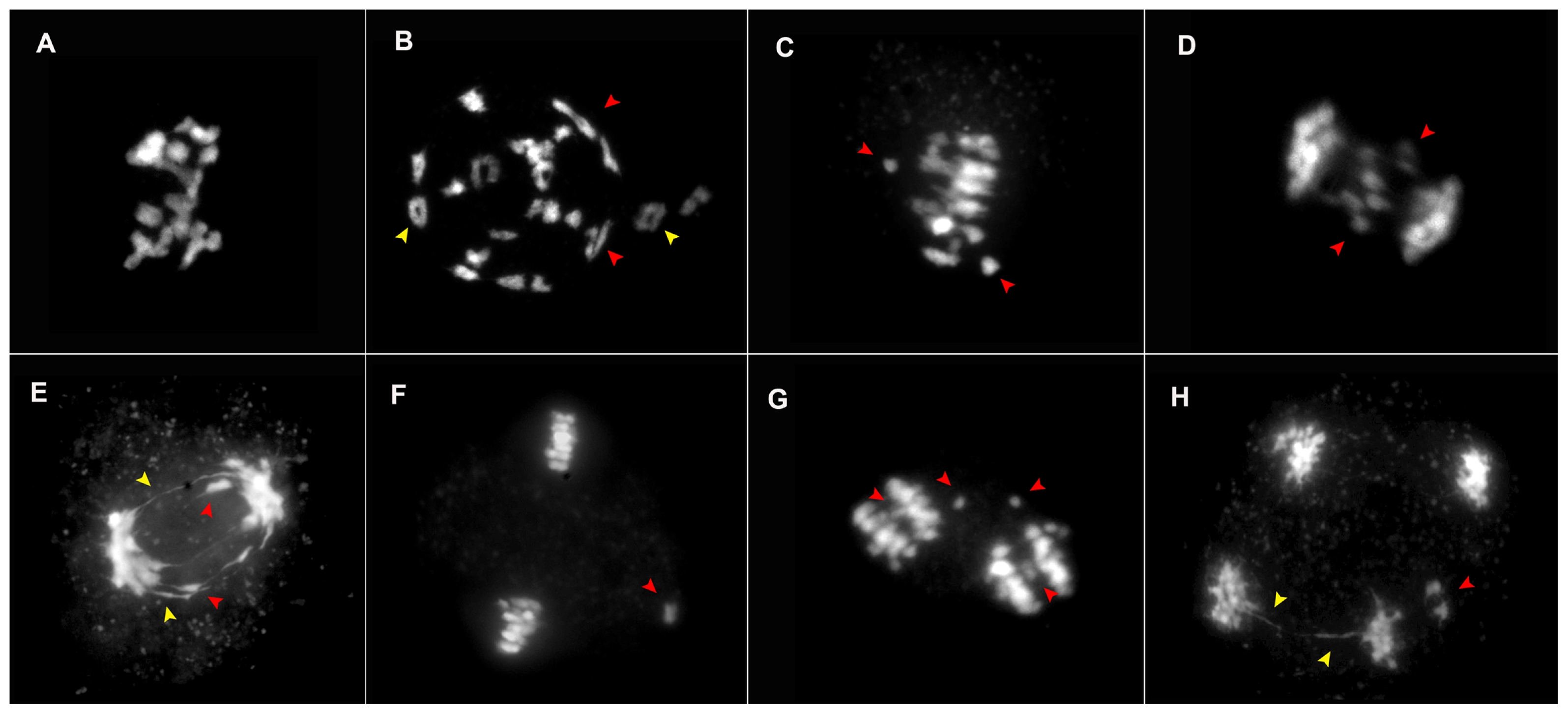
Table 1Summary of FISH karyotype analyses of intergeneric hybrid × Brassicoraphanus lines BB4, BB6, BB12, and BB50.
Table 1
|
Inbred lines |
Chr. no. (2n) |
Chr. length (μm) |
rDNA signals |
Karyotype formula (2n) |
|
|
|
Shortest |
Longest |
Total |
5S |
45S |
|
BB4 |
37 |
1.82 |
3.45 |
49.69 |
5 |
7 |
2m + 2m + 2sm + 2m + 2m + 2m + 2m + 2m + 2m + 2sm + 2m + 2m + 2smz) + 2mz) + 2m + 2sm + 2m + 2m + 2m |
|
BB6 |
38 |
2.31 |
4.43 |
61.39 |
6 |
7 |
2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2sm + 2sm + 2smz) + 2mz) + 2sm + 2m + 2m + 2m + 2m |
|
BB12 |
38 |
2.58 |
4.78 |
68.02 |
5 |
5 |
2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2smz) + 2mz) + 2m + 2m + 2m + 2m + 2m |
|
BB50 |
38 |
2.40 |
5.72 |
74.34 |
5 |
5 |
2m + 2m + 2m + 2m + 2sm + 2m + 2m + 2m + 2m + 2m + 2m + 2m + 2smz) + 2mz) + 2sm + 2sm + 2sm + 2sm + 2sm |
Table 2The frequency of meiotic abnormalities in different meiotic stages of the four inbred lines of ×Brassicoraphanus.
Table 2
|
Stages |
No. of abnormal PMCs |
|
|
BB4 |
BB6 |
BB12 |
BB50 |
|
Prophase I (diakinesis) |
|
Sticky chromosomes |
35 |
28 |
4 |
6 |
|
Rod/ring chromosomes |
18 |
7 |
8 |
0 |
|
Laggard chromosomes |
22 |
5 |
12 |
5 |
|
Metaphase I |
|
Laggard chromosomes |
154 |
65 |
15 |
17 |
|
Anaphase I |
|
Bridge fragmentation |
49 |
14 |
5 |
27 |
|
Metaphase II |
|
Laggard chromosomes |
8 |
36 |
6 |
5 |
|
Anaphase II |
|
Bridge fragmentation |
1 |
9 |
0 |
0 |
Total meiotic abnormality (%)
(per 500 PMCs) |
287 (57%) |
169 (34%) |
50 (10%) |
60 (12%) |
References
- Belandres HR, Waminal NE, Hwang YJ, Park BS, Lee SS, Huh JH. 2015. FISH karyotype and GISH meiotic pairing analyses of a stable intergeneric hybrid xBrassicoraphanus line BB#5. Korean J of Hortic Sci Technol. 33: 83-92.
- Bennett ST, Kenton AY, Bennett MD. 1992. Genomic in situ hybridization reveals the allopolyploid nature of Milium montianum (Gramineae). Chromosoma. 101: 420-424.
- Cai X, Xu SS. 2007. Meiosis-driven genome variation in plants. Curr Genomics. 8: 151-161.
- Cao W. 2003. Cytogenetic and molecular genetic evidence on evolution of genus Triticum. Plant Genome: Biodiversity Evolution. 1: 223-247.
- Capdeville G, Souza Júnior MT, Szinay D, Diniz LEC, Wijnker E, Swennen R. 2008. The potential of high-resolution BAC-FISH in banana breeding. Euphytica. 166: 431-443.
- Cheng B, Heneen W, Chen B. 1995. Mitotic karyotypes of Brassica campestris and Brassica alboglabra and identification of the B. alboglabra chromosome in an addition line. Genome. 38: 313-319.
- Comai L. 2005. The advantages and disadvantages of being polyploid. Nat Rev Genet. 6: 836-846.
- De Storme N, Mason A. 2014. Plant speciation through chromosome instability and ploidy change: cellular mechanisms, molecular factors and evolutionary relevance. Curr Plant Biol. 1: 10-33.
- Devi J, Ko J, Seo B. 2005. FISH and GISH: Modern cytogenetic techniques. Indian J Biotechnol. 4: 307-315.
- Dolstra O. 1982. Synthesis and fertility of xBrassicoraphanus and ways of transferring Raphanus chracters to Brassica. Agric Res Rep. 917: 1-90.
- Dover GA. 1989. Linkage disequilibrium and molecular drive in the rDNA gene family. Genetics. 122: 249-252.
- Drouin G, De Sá MM. 1995. The concerted evolution of 5S ribosomal genes linked to the repeat units of other multigene families. Mol Biol Evol. 12: 481-493.
- Eickbush TH, Eickbush DG. 2007. Finely orchestrated movements: evolution of the ribosomal RNA genes. Genetics. 175: 477-485.
- Fujii K, Ohmido N. 2011. Stable progeny production of the amphidiploid resynthesized Brassica napus cv. Hanakkori, a newly bred vegetable. Theor Appl Genet. 123: 1433-1443.
- Fukui K. 2005. Recent development of image analysis methods in plant chromosome research. Cytogenet Genome Res. 109: 83-89.
- Gaeta RT, Pires JC, Iniguez-Luy F, Leon E, Osborn TC. 2007. Genomic changes in resynthesized Brassica napus and their effect on gene expression and phenotype. Plant Cell. 19: 3403-3417.
- Grandont L, Cuñado N, Coriton O, Huteau V, Eber F, Chèvre AM. 2014. Homoeologous chromosome sorting and progression of meiotic recombination in Brassica napus: Ploidy does matter! Plant Cell. 26: 1448-1463.
- Hasterok R, Wolny E, Hosiawa M, Kowalczyk M, Kulak-Ksiazczyk S, Ksiazczyk T. 2006. Comparative analysis of rDNA distribution in chromosomes of various species of Brassicaceae. Ann Bot. 97: 205-216.
- Howell EC, Armstrong S. 2013. Using sequential fluorescence and genomic in situ hybridization (FISH and GISH) to distinguish the A and C genomes in Brassica napus. Methods Mol Biol. 990: 25-34.
- Hwang Y-J, Yu H-J, Mun J-H, Bok K, Park B-S, Lim KBJKJOHS. 2012. Centromere repeat DNA originated from Brassica rapa is detected in the centromere region of Raphanus sativus chromosomes. Korean J Hortic Sci. 30: 751-756.
- Hwang Y, Kim H, Kwon S, Yang T, Ko H, Park B. 2009. Karyotype analysis of three Brassica species using five different repetitive DNA markers by fluorescence in situ hybridization. Korean J Hortic Sci. 27: 456-463.
- Jellen E, Gill B, Cox T. 1994. Genomic in situ hybridization differentiates between A/D-and C-genome chromatin and detects intergenomic translocations in polyploid oat species (genus Avena). Genome. 37: 613-618.
- Kato A, Lamb JC, Birchler JA. 2004. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc Natl Acad Sci U S A. 101: 13554-13559.
- Kato M, Tokumasu S. 1983. The stabilization of chromosome numbers and the maintenance of euploidy in Brassicoraphanus. Euphytica. 32: 415-423.
- Kenton A, Parokonny AS, Gleba YY, Bennett MD. 1993. Characterization of the Nicotiana tabacum L. genome by molecular cytogenetics. Mol Gen Genet. 240: 159-169.
- Kiihl PRP, Pereira ARA, Godoy SMD, Stenzel NMC, Risso-Pascotto C. 2011. Chromosome stickiness during meiotic behavior analysis of Passiflora serrato-digitata L. (PassifloraCEAE). Ciênc Rural. 41: 1018-1023.
- Kim YJ, Pak WS, Koo KH, Kim MR, Jang JJ. 2000. Inhibitory effect of Baechu Kimchi (Chinese Cabbage Kimch,) and Kakduki (Radish Kimchi) on diethylnitrosamine and D-galactosamine induced hepatocardnogenesis. Food Sci Biotechnol. 9: 89-94.
- Kitashiba H, Li F, Hirakawa H, Kawanabe T, Zou Z, Hasegawa Y. 2014. Draft sequences of the radish (Raphanus sativus L.) genome. DNA Res. 21: 481-490.
- Koo DH, Hong CP, Batley J, Chung YS, Edwards D, Bang JW. 2011. Rapid divergence of repetitive DNAs in Brassica relatives. Genomics. 97: 173-185.
- Lee S-S, Choi W-J, Woo JG. 2002. Development of a new vegetable crop in xBrassicoraphanus by hybridization of Brassica campestris and Raphanus sativus. Korean J Hortic Sci. 43: 693-698.
- Lee S-S, Hwang BH, Kim TY, Yang J, Han NR, Kim J. 2017. Developing stable cultivar through microspore mutagenesis in xBrassicoraphanus koranhort, inter-generic allopolyploid between Brassica rapa and Raphanus sativus. Am J Plant Sci. 8: 1345-1356.
- Lee S-S, Kim J-K, Chun W. 1999. Breeding a fertile intergeneric allotetraploid plant between heading Chinese cabbage and korean radish. Korean J Hortic Sci. 17: 653-653.
- Lee S-S, Lee S-A, Yang J, Kim J. 2011. Developing stable progenies of xBrassicoraphanus, an intergeneric allopolyploid between Brassica rapa and Raphanus sativus, through induced mutation using microspore culture. Theor Appl Genet. 122: 885-891.
- Leitch A, Leitch I. 2008. Genomic plasticity and the diversity of polyploid plants. Science. 320: 481-483.
- Lim K-B, De Jong H, Yang T-J, Park J-Y, Kwon S-J, Kim JS. 2005. Characterization of rDNAs and tandem repeats in the heterochromatin of Brassica rapa. Mol Cells. 19: 436-444.
- Lim KB, Yang TJ, Hwang YJ, Kim JS, Park JY, Kwon SJ. 2007. Characterization of the centromere and peri-centromere retrotransposons in Brassica rapa and their distribution in related Brassica species. Plant J. 49: 173-183.
- Lim KY, Soltis DE, Soltis PS, Tate J, Matyasek R, Srubarova H. 2008. Rapid chromosome evolution in recently formed polyploids in Tragopogon (Asteraceae). PLoS One. 3: e3353
- Lim SJ, Lee SS, Bang JW. 2012. Karyotype and genomic in situ hybridization pattern in xBrassicoraphanus, an intergeneric hybrid between Brassica campestris ssp pekinensis and Raphanus sativus. Plant Biotechnol Rep. 6: 107-112.
- Lou Q, Iovene M, Spooner DM, Buell CR, Jiang J. 2010. Evolution of chromosome 6 of Solanum species revealed by comparative fluorescence in situ hybridization mapping. Chromosoma. 119: 435-442.
- Maluszynska J, Heslop-Harrison J. 1993. Physical mapping of rDNA loci in Brassica species. Genome. 36: 774-781.
- Mendes-Bonato AB, Pagliarini MS, Do Valle CB, Penteado MIDO. 2001. A severe case of chromosome stickiness in pollen mother cells of Brachiaria brizantha (Hochst.) Stapf (Gramineae). Cytologia. 66: 287-291.
- Moghe GD, Shiu SH. 2014. The causes and molecular consequences of polyploidy in flowering plants. Ann N Y Acad Sci. 1320: 16-34.
- Namai H, Sarashima M, Hosoda T. 1980. Interspecific and intergeneric hybridization breeding in Japan. Tsunoda S, Hinata K, Gómez-Campo C, editors. Brasscia crops and wild alles Biology and breeding. Japan Scientific Societies Press. Tokyo: pp. 191-204.
- Olin-Fatih M, Heneen W. 1992. C-banded karyotypes of Brassica campestris, B. oleracea, and B. napus. Genome. 35: 583-589.
- Olin-Fatih M. 1994. A new method for differential staining of Brassica metaphase chromosomes, and karyotypes of B. campestris, B. oleracea, and B. napus. Hereditas. 120: 253-259.
- Park HM, Jeon EJ, Waminal NE, Shin KS, Kweon SJ, Park BS. 2010. Detection of transgenes in three genetically modified rice lines by fluorescence in situ hybridization. Genes & Genomics. 32: 527-531.
- Pendinen G, Spooner DM, Jiang J, Gavrilenko T. 2012. Genomic in situ hybridization reveals both auto-and allopolyploid origins of different North and Central American hexaploid potato (Solanum sect. Petota) species. Genome. 55: 407-415.
- Pontes O, Neves N, Silva M, Lewis MS, Madlung A, Comai L. 2004. Chromosomal locus rearrangements are a rapid response to formation of the allotetraploid Arabidopsis suecica genome. Proc Natl Acad Sci U S A. 101: 18240-18245.
- Prakash S, Bhat SR, Quiros CF, Kirti PB, Chopra VL. 2009. Brassica and its close allies: cytogenetics and evolution. Janick J, editor. Plant Breeding Reviews. John Wiley & Sons, Inc.
- Rao P, Ranganadham P, Nirmala A. 1990. Behaviour of a ‘sticky-desynaptic’mutant in pearl millet. Genetica. 81: 221-227.
- Ravi M, Chan SWL. 2010. Haploid plants produced by centromere-mediated genome elimination. Nature. 464: 615-618.
- Ravi M, Shibata F, Ramahi JS, Nagaki K, Chen C, Murata M. 2011. Meiosis-specific loading of the centromere-specific histone CENH3 in Arabidopsis thaliana. PLoS Genet. 7: e1002121
- Sanei M, Pickering R, Kumke K, Nasuda S, Houben A. 2011. Loss of centromeric histone H3 (CENH3) from centromeres precedes uniparental chromosome elimination in interspecific barley hybrids. Proc Natl Acad Sci U S A. 108: E498-E505.
- Saylor LC, Smith BW. 1966. Meiotic irregularity in species and interspecific hybrids of Pinus. Am J Bot. 53: 453-468.
- Shaked H, Kashkush K, Ozkan H, Feldman M, Levy AA. 2001. Sequence elimination and cytosine methylation are rapid and reproducible responses of the genome to wide hybridization and allopolyploidy in wheat. Plant Cell. 13: 1749-1759.
- Singh M, Kumar R, Nagpure N, Kushwaha B, Gond I, Lakra W. 2009. Chromosomal localization of 18S and 5S rDNA using FISH in the genus Tor (Pisces, Cyprinidae). Genetica. 137: 245-252.
- Szadkowski E, Eber F, Huteau V, Lode M, Huneau C, Belcram H. 2010. The first meiosis of resynthesized Brassica napus, a genome blender. New Phytol. 186: 102-112.
- Vasconcelos S, De Souza AA, Gusmão CLS, Milani M, Benko-Iseppon AM, Brasileiro-Vidal AC. 2010. Heterochromatin and rDNA 5S and 45S sites as reliable cytogenetic markers for castor bean (Ricinus communis, Euphorbiaceae). Micron. 41: 746-753.
- Wang Y, Sonntag K, Rudloff E, Wehling P, Snowdon RJ. 2006. GISH analysis of disomic Brassica napus-Crambe abyssinica chromosome addition lines produced by microspore culture from monosomic addition lines. Plant Cell Rep. 25: 35-40.
- Xiong Z, Pires JC. 2011. Karyotype and identification of all homoeologous chromosomes of allopolyploid Brassica napus and its diploid progenitors. Genetics. 187: 37-49.
- Yang Q, Hanson L, Bennett M, Leitch I. 1999. Genome structure and evolution in the allohexaploid weed Avena fatua L.(Poaceae). Genome. 42: 512-518.
- Yao X-C, Ge X-H, Chen J-P, Li Z-Y. 2010. Intra-and intergenomic relationships in interspecific hybrids between Brassica (B. rapa, B. napus) and a wild species B. maurorum as revealed by genomic in situ hybridization (GISH). Euphytica. 173: 113-120.