Abstract
Temperatures that extend beyond normal levels of tolerance cause severe stress to plants, especially during the reproductive and grain filling/ripening stages. Heat stress leads to serious yield losses in many crop plants, including rice (Oryza sativa). In view of the current scenario of global climate change, frequent fluctuations and a significant increase in average temperatures will pose challenges to protecting those yields. Therefore, elucidating the molecular mechanisms that make crop plants more tolerant of heat, particularly in organs at the reproductive stage, is of utmost importance. Precise molecular information will be helpful for the manipulation and exploration of relevant genes for use in crop improvement programs. In this review, we highlight recent progress in research on the molecular responses to high temperatures in pollen and seed and provide a perspective on the development of heat tolerance in rice cultivars. The responsible mechanism is a very complex phenomenon that involves several biochemical and physiological changes, molecular responses, and a series of signal transductions. Improving our understanding requires detailed knowledge at various omics levels. Recent technological advancements have accelerated genomics, transcriptomics, and proteomics studies in rice, a model crop plant. Here, we discuss those technological and omics approaches being taken to investigate the heat tolerance mechanism, particularly in rice. In addition, we address the tools being used to identify key genes and QTLs that can then be utilized for molecular breeding and biotechnology.
-
Key words: Heat stress tolerance, Pollen development, Rice, Seed development, Spikelet fertility
INTRODUCTION
Rice (
Oryza sativa) is a staple food crop worldwide, upon which almost half of the population depends. To meet growing demands by consumers there as well as in other parts of the world, annual grain yields must increase by 0.6 to 0.9% until 2050 (
Carriger and Vallee 2007). However, it is difficult to combat the challenges associated with situations such as an ever-changing climate, decreases in the amount of agricultural land available per capita, and greater frequencies of biotic and abiotic stresses. The primary concern is global warming and unpredictable temperature fluctuations. Conventional rice-growing areas feature relatively higher day and night temperatures (28/22°C) that are more strongly influenced by climate change (
Prasad et al. 2006). A mathematical model has projected a 2.0 to 4.5°C increase in the global temperature by the end of the century (
IPCC 2007). Those predictions are even more intimidating for rice-growing regions such as Asia, the Middle East, and Africa. If the average temperature were to rise by 3 to 4°C in those areas that would lead to a 15 to 35% reduction in crop yields (
Ortiz et al. 2008).
High-temperature stress (HTS) alters several physiological and metabolic processes in cells. The vulnerability of plants to HTS differs according to developmental stage, but it affects both vegetative and reproductive phases to some extent. Those effects depend upon species and genotype, and an ample amount of inter- or intraspecific variations can occur (
Sakata and Higashitani 2008). Various morphological responses have been observed, e.g., wounding and aging of leaves; inhibition of leaf, stem, root, and shoot growth; a reduction in seed germination rates; and unevenness in seed maturation. Many species undergo structural changes to their hypocotyls and floral organs to avoid the consequences of high temperatures (
Tian et al. 2009). Likewise, HTS affects the total assimilation rate in the shoot, which then contributes to a decrease in whole-plant dry weight (
Wahid et al. 2007). Other physiological adjustments include the production of reactive oxygen species, increased water loss, greater membrane permeability, and decreased photosynthesis. Although these physiological effects have been evaluated in some plants (
Wahid et al. 2007;
Bokszczanin et al. 2013;
Mathur et al. 2014), HTS responses in rice have not received much focus, and only a few studies have examined these responses at the flowering stage or in reproductive organs (
Jagadish et al. 2010b;
Rang et al. 2010).
Here, we describe the recent progress made in research that addresses these high-temperature influences during the reproductive and grain-filling/ripening stages in rice. We summarize the current understanding about the molecular and genetic events involved in male and female organ development under HTS, while presenting omics-scale studies and advanced breeding strategies being used to improve HTS tolerance. Key candidate genes, including transcription factors (TFs), and the molecular pathways involved in this mechanism, particularly during the reproductive stages, are also discussed, along with genetic engineering and molecular breeding approaches that are being applied to produce rice that is tolerant of high temperatures.
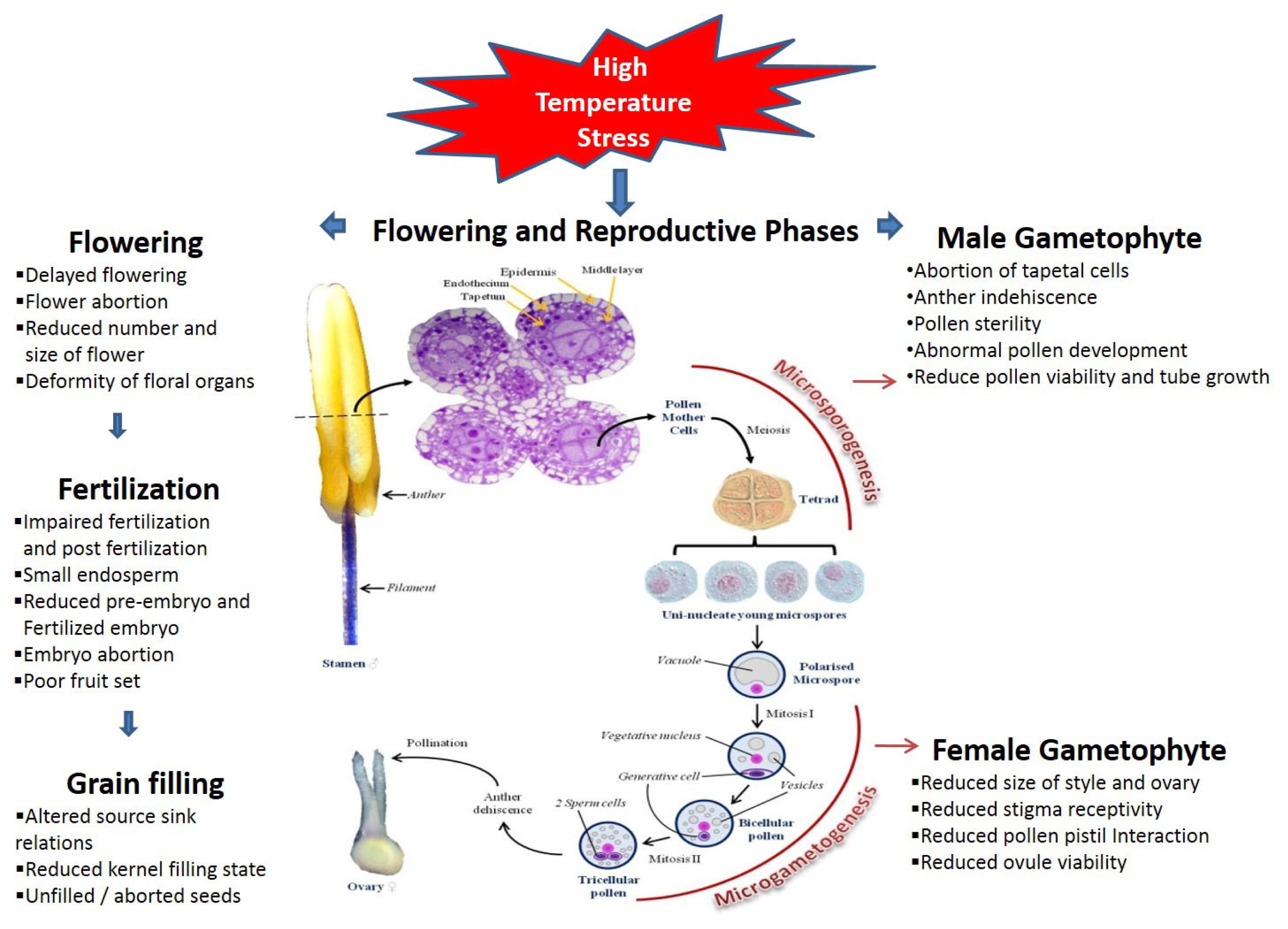
Effect of high-temperature stress on pollen and anther development
In flowering plants, sexual reproduction usually requires flowers that produce male and female gametophytes. This process can be restricted because formation of the male gametophyte (pollen) is more sensitive to the growing environment (
Ahmed et al. 1992;
Peet et al. 1998;
Kim et al. 2001;
Rieu et al. 2017). Pollen develops inside locules from diploid pollen mother cells that undergo cell division (meiosis) to generate four haploid microspores in the locular fluid. Each microspore then asymmetrically divides to form a vegetative cell (VC) and generative cell (GC) during pollen mitosis I. Small GCs are subsequently engulfed within the cytoplasm of the larger VC and further cell division occurs, i.e., mitosis (pollen mitosis II), to form two sperm cells (
McCormick 2004). During this process, the tapetum is generated from the innermost anther wall (
Goldberg et al. 1993). The tapetum plays an important role at the early microspore stage, providing the nutrients, carbohydrates, lipids, and enzymes necessary for synthesis of the outer pollen wall. This process is closely associated with microspore development, and the tapetum degenerates soon after the microspore is released from the tetrad. Therefore, proper tapetal functioning and its timely degradation are crucial components in pollen development (
Rieu et al. 2017).
Abiotic-stress conditions such as HTS have a severe impact on pollen and anther development. Various reproduction-related events affected by heat stress are shown in
Fig. 1. Analysis of tapetum sensitivity, based on transcriptome studies of rice, has shown how the pattern of expression by heat-repressed genes can vary in tapetal tissue (
Endo et al. 2009). Pollen tends to be more sensitive than the female gametophyte to HTS across plant species, including chickpea (
Cicer arietinum) (
Clarke and Siddique 2004;
Devasirvatham et al. 2012,
2013;
Kaushal et al. 2013), rice (
Sakata and Higashitani 2008;
Wassmann et al. 2009), tomato (
Lycopersicon esculentum) (
Giorno et al. 2013), maize (
Zea mays) (
Herrero and Johnson 1980), and
Vigna unguiculata (
Ahmed et al. 1992). When compared with other cereal crops such as maize and wheat (
Triticum aestivum), rice is relatively more sensitive to HTS (> 35°C), mainly during the stages of flowering (
Prasad et al. 2006;
Jagadish et al. 2007,
2008,
2010b) and gametogenesis (
Jagadish et al. 2013). Under such high temperatures, microspores are aborted and pollen viability is reduced during anthesis, subsequently causing spikelet sterility (
Cheng et al. 2009;
Mohammed and Tarpley 2009a,
2009b,
2010). The relationship between heat stress and spikelet sterility has been reported in rice (
Nguyen et al. 2013;
Oort et al. 2014) and other crop species (
Oshino 2007;
Sakata et al. 2008). As demonstrated in several investigations, HTS can severely alter the morphology, structure, and functions of male gametophytes. Because pollen quality is the most crucial factor in determining crop yields during periods of heat stress (
Firon et al. 2006;
Rang et al. 2010), the results of analyses on pollen viability and spikelet fertility under elevated temperatures can help improve our understanding of how heat tolerance is conferred in rice.
As part of the reproductive process, pistils, the maternal organ, form in the flower, providing nutrition and protection to the female gametophyte while also supporting the male gametophyte. Although this organ is considered more tolerant of HTS than the male gametophyte (
Hedhly et al. 2011), the effect of such stress on the sporopytic tissue of the pistil has not been widely studied. Nevertheless, high temperatures influence the pistil both pre-and post-pollination as well as during the stages of embryo development, silique formation, and seed set. Species can vary in their sensitivity to heat, based on genotype and the temperatures to which they are exposed. In crops such as
Prunus armeniaca (
Rodrigo and Herrero 2002), HTS causes shorter styles and abnormal ovary growth. Similarly, post-pollination defects in the female gametophyte have been reported in chickpea (
Srinivasan et al. 1999), and in rapeseed (
Brassica napus L.) (
Young et al. 2004). In many species, the impact of HTS can be more detrimental on microsporogenesis than on macrosporogenesis, and the receptivity of pollen on stigmas can be reduced, leading to failed fertilization (
Srinivasan et al. 1999). For
Prunus persica L., elevated temperatures mean that pollen germination and pollen tube growth cannot be supported (
Hedhly et al. 2005). Reports have been made about ovary defects in wheat (
Saini et al. 1983) and in Arabidopsis, where HTS is associated with a reduction in the number of ovules as well as more abortions (
Whittle et al. 2009). In rapeseed, increasing the growing temperature from 26°C to 32°C is linked with defects of the embryo sac (
Polowick and Sawhney 1988). Elevated temperatures cause pistil hyperplasia in rice, which leads to spikelet sterility and stamen hypoplasia (
Takeoka et al. 1991).
Both monocotyledonous and dicotyledonous plants are affected by HTS (
Herrero and Johnson 1980;
Saini et al. 1983;
Carlson 1990;
Nuttal et al. 1992;
Morrison 1993;
Peet et al. 1998;
Angadi et al. 2000;
Sato et al. 2002). Heat stress that occurs during the period of seed development can decrease grain size for many cereal crops (
Nagato and Ebata 1960;
Hunter et al. 1977;
Savin et al. 1996). In rice, the intricate process of reproductive differentiation and degeneration has three genetic components: diploid maternal tissues (e.g., pericarp, testa, and nucellus), triploid endosperm, and diploid embryos (
Wu et al. 2016a,
2016b). There, the early stage of seed development is highly sensitive to heat stress (
Folsom et al. 2014). Because the endosperm accounts for most of the rice caryopsis, any defect in it can affect the final seed size. Formation of the rice endosperm requires distinct developmental processes, i.e., syncytial nuclear division, cellularization, differentiation, and maturation (
Olsen 2001;
Sabelli and Larkins 2009;
Wu et al. 2016a). Shrunken endosperm results from a noticeable reduction in starch synthesis (
Lee et al. 2007). The duration of the syncytial phase and the timing of cellularization are critical because they are correlated with the extent of nuclear proliferation and may influence grain filling and grain weight (
Brown et al. 1996; Mitzutani
et al. 2010). Heat stress is linked with diverse types of chalky rice grains, including white-black, white-belly basal white, and milky-white (
Wada et al. 2015).
Folsom et al. (2014) have demonstrated that initiation of endosperm cellularization is controlled by a rice Polycomb Repressive Complex2 member, Fertilization-Independent Endosperm 1 (
OsFIE1), and can be influenced by HTS. This gene is maternally expressed and endosperm-imprinted (
Zhang et al. 2012). Its expression increases after fertilization due to demethylation, which is associated with the initiation of endosperm cellularization (
Folsom et al. 2014;
Chen et al. 2016). When developing seeds are exposed to elevated temperatures, changes occur in both the methylation level of the 5’ coding region of
OsFIE1 and its H3K9me methylation mark (
Folsom et al. 2014). These reports suggest that
OsFIE1 is a potential component involved in regulating seed enlargement under heat stress. Other genes, such as rice
AGL36 and the orthologs of
Pheres1,
OsMADS82, and
OsMADS87, are epigenetically repressed by
OsFIE1.
Chen et al. (2016) have identified putative imprinted genes during the endosperm transition from syncytium to the cellularization stage in young heat-stressed seeds. Such conditions promote a mis-timed release of imprinting from a subset of seed-specific genes in rice, thereby leading to the thermal resilience of rice during reproductive development. Indeed, one heat-sensitive imprinted gene,
OsMADS87, a rice homolog of the Arabidopsis endosperm-specific
PHE1 (
Köhler et al. 2005), directly regulates rice seed size (
Chen et al. 2016).
When regions in diverse climates experience ever-changing temperatures during the reproductive stage, seed development and fruit set are altered. Although HTS can reduce seed production in various species, it remains unclear whether this is an outcome of decreased flowering, fruit abortion, or disruptions in fertilization or post-fertilization events. These impacts of HTS can be examined through approaches that utilize microarrays, RNA sequencing (RNA-Seq), or screening of genes that are expressed during the post-fertilization process. The results of such research can be used to improve our understanding of the heat-stress response in developing rice seeds and the impact of HTS on crop yields.
Functional characterization of rice genes that confer tolerance to high-temperature stress
Because the rice genome is well-annotated, it provides a basis for comparative genomics among crop species. Many genes have been identified in rice through multiple approaches, including microarrays and RNA-Seq, proteomics data, protein–protein interactions, and genome-wide gene-indexed mutant populations (
Chandran and Jung 2014). These data are helpful for predicting gene functions. The information made publicly available from genetic analyses, gene-indexed mutants, or transgenic plants have enabled us to learn the functions of more than 1,000 genes in rice (
Yamamoto et al. 2012). Further progress can be made based on high-throughput omics data. For example, one important resource for functional genomics is provided by the International Rice Functional Genomics Consortium (IRFGC), which has generated gene-indexed mutants that can facilitate functional annotations for most rice genes. All of the information about functionally characterized genes can be accessed using the Overview of functionally characterized Genes in Rice Online database, or OGRO (
Yamamoto et al. 2012;
http://qtaro.abr.affrc.go.jp/ogro).
Table 1 presents the genes that have been identified as responsible for HTS tolerance, based on functional characterization and validation through overexpression analysis, mutant screening, and transgenic approaches.
Several studies involving expression profiling have identified genes that are induced by environmental stresses (
Rabbani et al. 2003;
Bray 2004;
Yamaguchi-Shinozaki and Shinozaki 2006;
Zhang et al. 2012). Using micro-arrays, gene expression at the transcriptome level has been monitored in the reproductive and ripening tissues of heat-sensitive ‘Nipponbare’ rice and the heat-tolerant ‘996’ (
Endo et al. 2009;
Zhang et al. 2012). Furthermore, RNA-Seq has been conducted to examine the reproductive tissues during anthesis for heat-tolerant ‘N22’ rice and the sensitive cultivars ‘IR64’ and ‘6264’ (
González-Schain et al. 2016).
Endo et al. (2009) have found that, under HTS conditions, the expression of more than 1,400 genes is significantly changed in the anthers from ‘Nipponbare’ plants. Similarly,
Zhang et al. (2012) have described almost 2,500 heat-responsive genes in young florets during meiosis.
González-Schain et al. (2016) have reported that, among 630 heat-responsive genes, a relatively small number (37) are core heat-responsive. Under combined drought and heat stress, 20 genes are up-regulated in pollinated rice pistils (
Li et al. 2015a).
Table 2 shows the heat-responsive genes expressed in rice reproductive and ripening tissues that can potentially be used to improve HTS tolerance in that crop. The large number of differentially expressed genes (DEGs) suggested by expression profiling will be useful in future studies that can expand our understanding of the overall molecular responses and probable pathways involved in the stress response. For example,
Endo et al. (2009),
Zhang et al. (2012), and
González-Schain et al. (2016) have determined that TFs are over-represented among the DEGs that respond to heat stress during the reproductive stage in rice. Genes that regulate the pathways for sugar and amino acid metabolism in rice also have important roles in heat-stress tolerance (
Li et al. 2015a). Therefore, these recent reports of numerous DEGs in rice can help us interpret the interactions and possible involvement of these key reproduction-related genes that might also confer HTS tolerance.
Chalkiness and smaller-than-normal grains are typical of rice exposed to heat stress during the stage of caryopsis maturation. Such symptoms have been reported in transgenic and mutant rice with decreased or deficient expression of various starch synthesis-related genes, including those for a sucrose transporter (
SUT1), invertase (
CIN2), UDP-glucose pyrophosphorylase (
UGPase), ADP-glucose pyrophosphorylase (
AGPS), soluble starch synthase (
SSIIIa), and branching enzyme (
BEIIb) (
Nishi et al. 2001;
Kawagoe et al. 2005;
Fujita et al. 2007;
Wang et al. 2008;
Woo et al. 2008;
Ito et al. 2009). Transcriptomic profiling has revealed that high temperatures down-regulate the expression of those genes as well as various others involved in sucrose import/degradation and starch biosynthesis while up-regulating genes for starch degradation (
Yamakawa et al. 2007;
Yamakawa and Hakata 2010). Transgenic rice plants that over-express amylase genes also display the chalky endosperm phenotype, suggesting that premature autolysis of starch granules is one reason that this phenotype occurs for rice grains that ripen under high temperatures (
Asatsuma et al. 2006). Examination of the rice grain proteome under HTS has revealed differential expression (up- or down-regulation by more than 2-fold) for 61 genes implicated in diverse metabolic and physiological functions, such as protein synthesis, redox homeostasis, lipid metabolism, and starch biosynthesis/degradation (
Kaneko et al. 2016). In addition to proteins related to starch metabolism, storage proteins also accumulate, accounting for up to 10% of the total dry matter and affecting the physicochemical characteristics of rice flour and brewing quality that are also differentially affected by heat stress. Meanwhile, accumulations of prolamin and globulin, but not of glutelin and albumin, are reduced under high temperatures (
Yamakawa et al. 2007;
Lin et al. 2010). Enhanced expression of small heat shock proteins or redox homeostasis-related genes under high temperatures is strongly correlated with the occurrence of chalky grains, as demonstrated in studies of various conventional cultivars and transgenic lines of rice (
Lin et al. 2005,
2014;
Liu et al. 2010,
2011). Metabolomic analysis of the developing caryopsis has indicated that high temperatures decrease the levels of glucose, fructose, most sugar phosphates, and organic acids involved in glycolysis/gluconeogenesis and the TCA cycle while increasing the amounts of sucrose, UDP-glucose, and ADP-glucose (
Yamakawa and Hakata 2010). Such metabolomic changes also closely parallel the inhibition of starch accumulations and transcriptomic alterations in the developing caryopsis under high temperature (
Yamakawa et al. 2007;
Yamakawa and Hakata 2010).
Research with genetic and transcriptome profiling has shown that the high temperature-induced decline in grain weight and quality is a very complex phenomenon that requires the interactions of various metabolic pathways, including those for sugar import/degradation, starch synthesis/degradation, storage protein synthesis, and maintenance of homeostasis. Several candidate genes involved in the mechanism for heat-stress tolerance have already been identified in many plant species. To elucidate these multifaceted effects of high temperature on the various metabolic and physiological processes of filling grains, researchers must still investigate how such stress can crosstalk with the developmental program of seed ripening or metabolic regulation when the caryopsis is maturing.
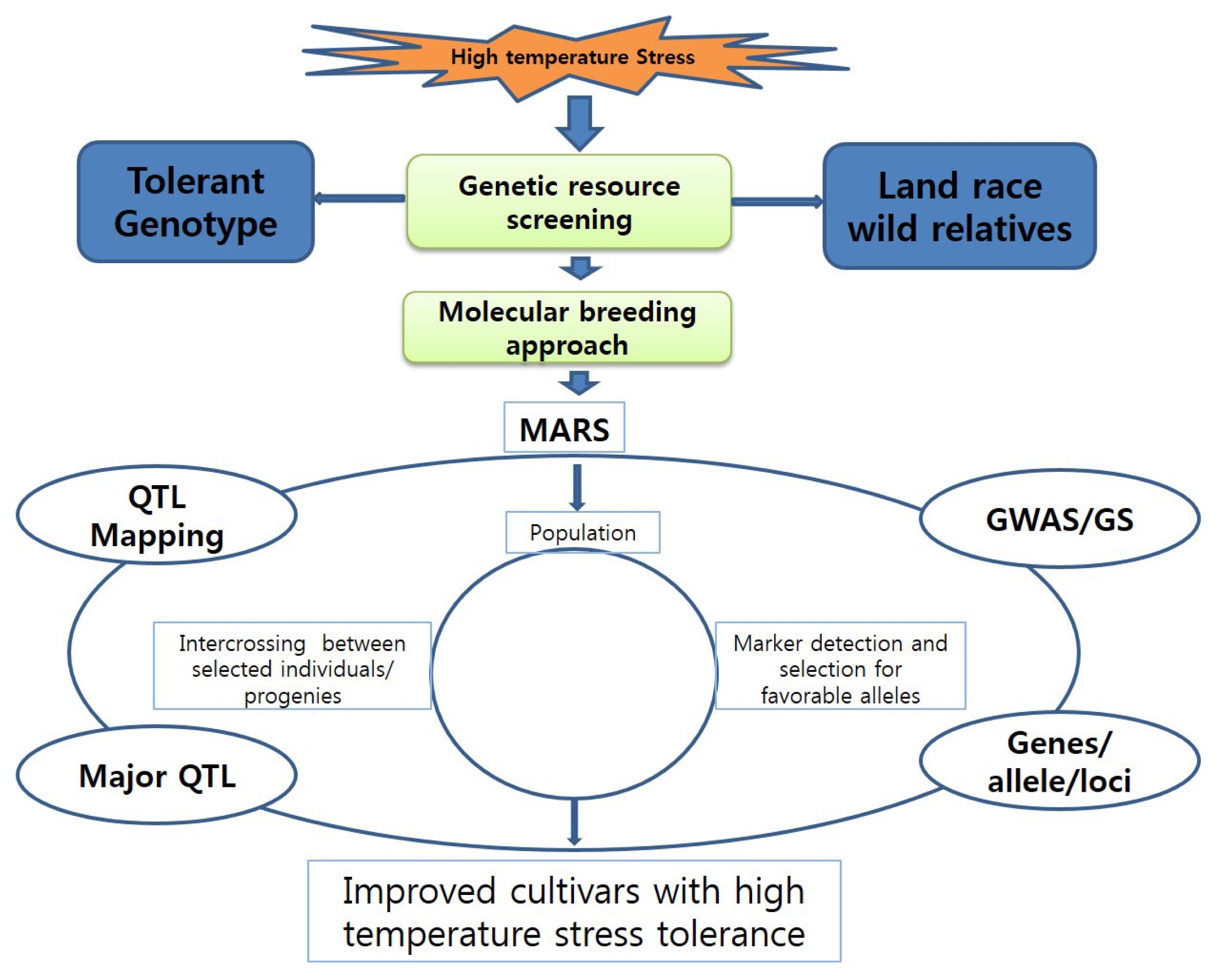
Breeding strategy for introducing tolerance to high temperatures
Breeding approaches involve the improvement of wild species to produce elite or domesticated breeding lines that introduce novel alleles into desired crops. High-temperature tolerance is a polygenic and complex trait highly influenced by environmental variations, but the genetics behind such tolerance is less understood (
Wahid et al. 2007;
Collins et al. 2008;
Ainsworth and Ort 2010). Due to uncontrolled environmental factors and the influence of additional biotic stresses, it can be difficult to select for high-temperature tolerance through conventional breeding. Therefore, better methods are needed for conducting more accurate greenhouse experiments. To gain further understanding, rice researchers in the past decade have looked to other tools for identifying the genes and quantitative trait loci (QTLs) associated with HTS tolerance. The availability of the well-annotated genome sequence (IRGSP 2005) has enabled significant advances to be made in this relatively new genomics field. Marker assisted recurrent selection (MARS) and genomic selection (GS) are proving efficient for the development of novel cultivars in many crops (
Varshney et al. 2009). Each tool has a potential advantage over marker-assisted back-crossing (MABC), which requires prior knowledge about the genetic markers associated with genes, QTLs, or alleles that affect whole-plant stress tolerance or contributing mechanisms. Both MARS and GS can also handle the problem of stacking minor alleles and QTLs. Although MARS breeding is widely used in the private sector (
Ribaut et al. 2010), its application has not been reported in the public sector. The second most comprehensive approach for predicting the breeding value of an individual within a breeding population has been the GS method (
Meuwissen et al. 2001). However,
Collins et al. (2008) have demonstrated that relying upon complex traits, e.g., heat-stress tolerance, for crop improvement is not effective mainly due to genotype × environment and gene–gene (i.e., epistasis) interactions, which frequently result in low breeding efficiency. In general, monogenic or less complex traits are easier to predict with GS tools while traits such as stress responses are more complex and harder to estimate by the genomic estimated breeding value, or GEBV. When heat-stress tolerance, which has a quantitative hereditary characteristic, is the focus, then MARS seems to be one of the most efficient methods for plant breeding. After using MARS,
Jain et al. (2014) have described improvements in drought and heat tolerance in wheat, while International Maize and Wheat Improvement Center (
http://www.cimmyt.org) have started utilizing this approach in maize, and
Spindel et al. (2015) have made strong advancements via MARS in rice.
Efforts to achieve HTS tolerance are still in the early phases.
Fig. 2 presents several general strategies that are being explored. The complexity of this trait makes it difficult to obtain suitable breeding material using inbred lines and commercial hybrids. Progress might be accelerated by identifying and characterizing the genetic resources for HTS and screening a large population that covers various developmental stages (
Wahid et al. 2007). An advanced molecular breeding approach would also be advantageous because it could provide data that would contribute significantly to the development of HTS-tolerant rice plants.
Abundant information is available regarding the physiological and metabolic traits that define HTS tolerance. However, little is known about the responsible mechanism(s). With the goal of improving crop yields under various stress conditions, two standard biotechnological approaches have been taken: development of transgenic plants and advanced molecular breeding.
Transgenics tools have become more efficient for introducing novel traits for biotic and abiotic stresses while also annotating candidate genes for molecular functions and regulation.
Tables 1–
3 show the genes, TFs, and QTLs already identified in rice through various means. Genetic engineering can also be applied to manipulate genes from other species that might be transferred to an heterologous system.
Katiyar-Agarwal et al. (2003) have demonstrated enhanced heat tolerance in a transgenic line of ‘Pusa Basmati 1’ rice by over-expressing an Athsp101 protein. Their transgenic plants are able to survive being exposed to severe heat stress, i.e., 45 to 50°C, and they show vigorous growth during subsequent recovery at 28°C.
Shen et al. (2015) have reported that overexpression of the
Arabidopsis thaliana receptor-like kinase
ERECTA (
ER) improves thermotolerance in
Arabidopsis, rice, and tomato. A transgenic rice line that over-expresses
ER has 55 to 70% higher seed set when compared with the non-transformed control line. Similarly,
Li et al. (2015b) have indicated that a natural allele of a proteasome alpha 2 subunit gene can contribute to thermotolerance and adaptation by African rice species.
Fang et al. (2015) have described a stress-responsive NAC TF,
SNAC3, that confers heat and drought tolerance in rice. Furthermore, when that gene is knocked out, the transgenics become susceptible to HTS. However, despite these reports of transgenic heat-tolerant rice, less effort has been applied in that research area when compared with the efforts made to develop lines that are salt-, cold-, or drought-tolerant.
Heterologous expression of genes in transgenic plants is highly influenced by the choice of promoter used to regulate the transgene. Several well-characterized promoters that provide ubiquitous or tissue-specific gene expression are available for rice.
Jeong and Jung (2015) have reviewed techniques that utilize a tissue-specific promoter rather than constitutive and spatio-temporal expression to exploit an effect of the transgene on transgenic plants. However, such approaches have not been widely applied in crop biotechnology and require further evaluation if they are to be successful in crop improvement. Genetic engineering provides great promise for enhancing plant tolerance to emerging abiotic stresses, and it can have a significant impact on the manipulation of innate genetic potential. In rice, this technology is still in the early phase but might possibly be used to transfer multiple genes for abiotic-stress tolerance. Additional knowledge about tolerance modules, including the identification of tissue-specific promoters and responsive genes, might enable the transformation of rice plants with multiple genes. Only a few field tests with transgenic rice have been reported. Before conducting such studies under field conditions, researchers must have a clear understanding of the genetic, biochemical, metabolic, and physiological mechanisms by which plants achieve HTS tolerance. Moreover, an advanced molecular breeding approach has a great advantage over the transgenic approach because it is universally accepted as a technology that can contribute significantly to the development of HTS-tolerant rice plants.
Because of predicted global climate changes, an appropriate strategy is urgently needed to combat the losses that will occur due to high-temperature stress. Rice is a very important crop for supporting food security. Plants are much more sensitive to high temperatures when exposed during the reproductive stage. Production of functional and viable pollen that complete the pollination process is a crucial aspect to ensuring natural fertility and stable crop yields. Understanding the complexity of regulatory molecular mechanisms involved in these processes can be eased through omics-based studies that lead to the identification of many key genes, TFs, and QTLs. Genome-wide analyses that rely upon microarrays or next generation technologies such as RNA-Seq have started to provide molecular information on candidate genes involved in heat tolerance, which can then be further characterized in depth. Additional efforts should be made to generate integrated data from omics, metabolic, and proteomics analyses. Heat stress is a polygenic characteristic that is often defined based upon complex traits such as yield under stress. Therefore, introgression of genes, TFs, and QTLs through conventional breeding is not feasible when developing an HTS-tolerant cultivar. Instead, alternative strategies such as advanced molecular breeding or genetic engineering are necessary if we are to develop rice plants that are tolerant to high temperatures.
ACKNOWLEDGEMENTS
This work was supported in part by grants from the Next-Generation BioGreen 21 Program (Agricultural Biotechnology Research Center. Project No. 01369001 to SKP) and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2016R1D1A1A09919568 to KHJ).
Fig. 1Effect of high temperature on flowering and reproductive phases.
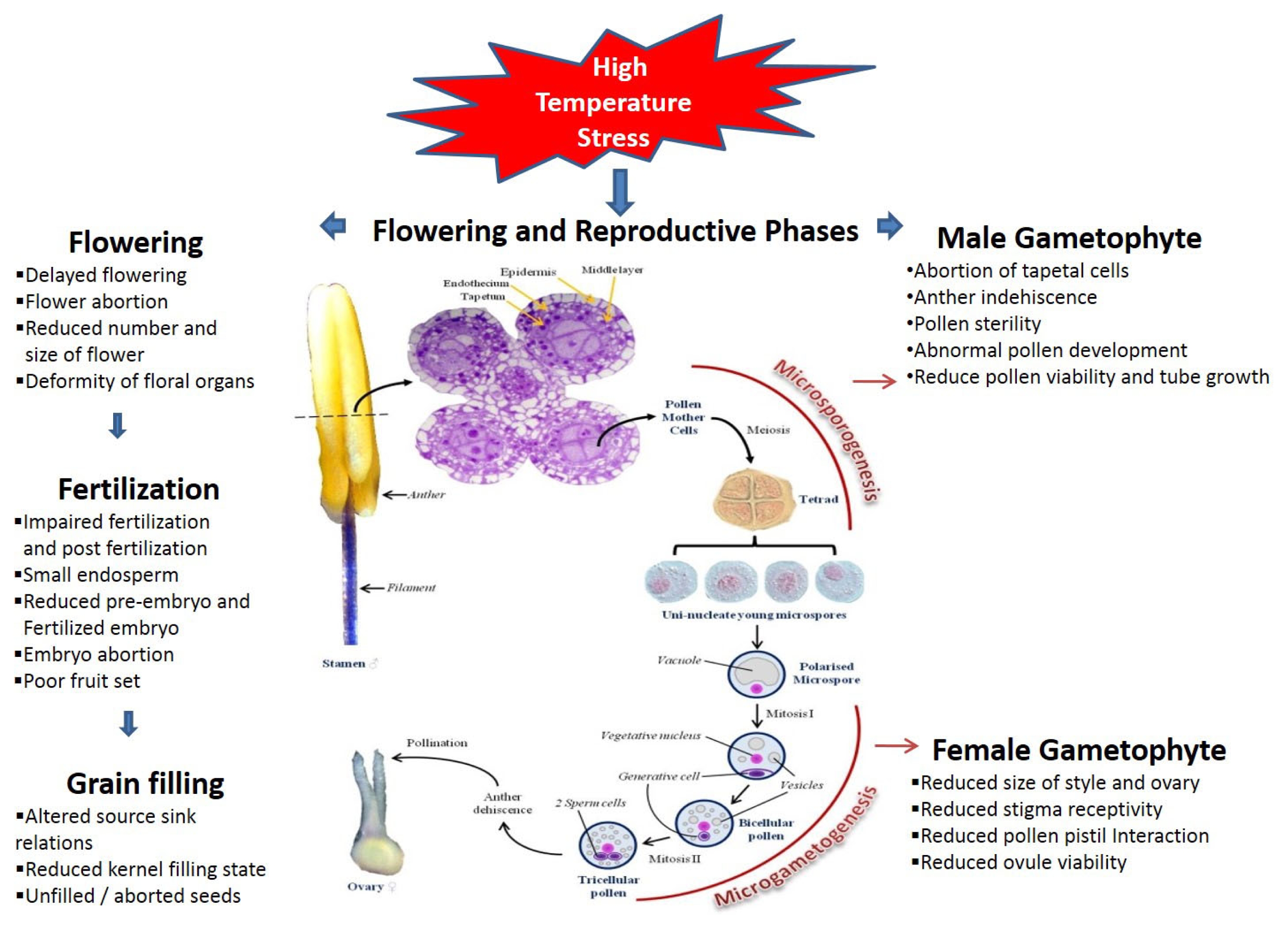
Fig. 2General strategies used to breed HTS-tolerant plants.
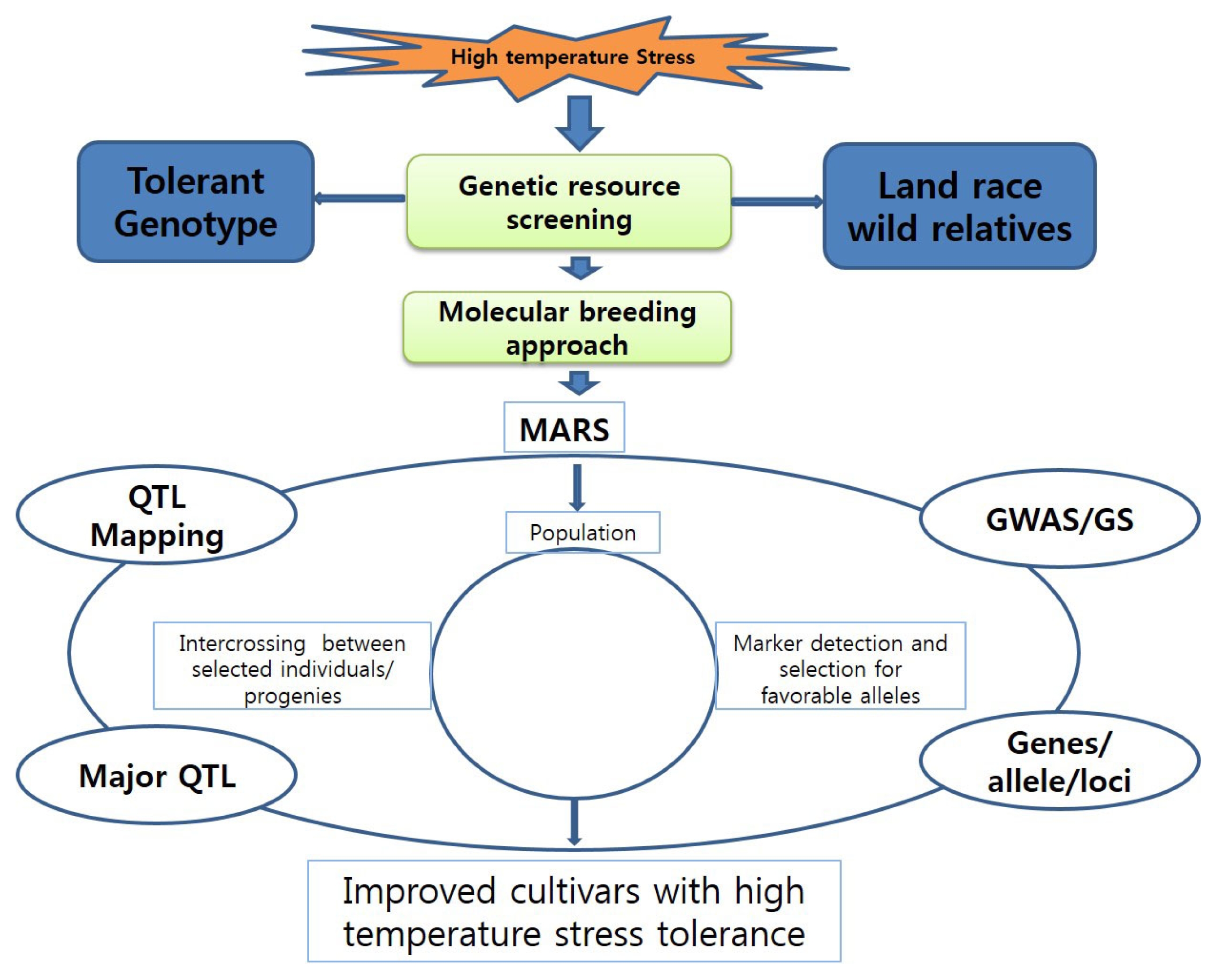
Table 1Summary of recent researches in rice for increased high temperature stress tolerance.
Table 1
|
Gene |
Gene symbol |
Plant species |
Methods |
Known functions |
Reference |
|
Glycogen synthase kinase3-like gene 1 |
OsGSK1 |
Rice |
Mutant |
Cold, drought, salinity and heat tolerance |
, Koh et al. 2007
|
|
Oswrky11 |
OsWRKY11 |
Rice |
Overexpression |
Drought and heat tolerance |
, Wu et al. 2009
|
|
Sedoheptulose-1,7-bisphosph atase |
SBPase |
Rice |
Overexpression |
Photosynthetic ability under high temperature condition |
, Feng et al. 2007
|
|
Mitochondrial heat shock protein 70 |
mtHsp70 |
Rice |
Overexpression |
Heat and redox induced programmed cell death in protoplast |
Qi et al. 2010 |
|
Small heat-shock protein17.7 |
sHSP17.7 |
Rice |
Overexpression |
Heat and UV tolerance |
, Murakami et al. 2004
|
|
Hsf binding protein 2 |
OsHSBP2 |
Rice |
Knockdown Overexpression |
Heat shock tolerance |
, Rana et al. 2012
|
|
Hsf binding protein 1 |
OsHSBP1 |
Rice |
Knockdown Overexpression |
Heat shock tolerance. |
, Rana et al. 2012
|
|
Osmyb55 |
OsMYB55 |
Rice |
Overexpression |
Growth under high temperature |
, El-kereamy et al. 2012
|
|
Rice carbon catabolite repressor 4(ccr4)-associated factor 1g |
OsCAF1G |
Rice |
Others |
ABA, heat stress tolerance |
, Chou et al. 2014
|
|
Rice carbon catabolite repressor 4(ccr4)-associated factor 1h |
OsCAF1H |
Rice |
Others |
Heat stress tolerance |
, Chou et al. 2014
|
|
Small heat shock proteins (shsps) oshsp16.9 |
HSP 16.9 |
Rice |
Overexpression |
Drought and cold, salt and heat tolerance |
Jung et al. 2014 |
|
A 2 subunit 26s proteasome |
TT1 (Os03g0387100) |
Rice |
Overexpression |
Heat stress tolerance |
, Li et al. 2015b
|
|
Manganese superoxide dismutase |
SOD 1 (MSD1) |
Rice |
Overexpression |
Heat stress tolerance |
, Shiraya et al. 2015
|
|
Spl7 |
HSFA4d |
Rice |
Overexpression |
Heat stress tolerance |
, Yamanouchi et al. 2002
|
|
Receptor-like kinase erecta |
ERECTA |
Rice |
Overexpression |
Heat stress tolerance |
, Shen et al. 2015
|
|
Hsp101 |
AtHsp101 |
A. thaliana
|
Overexpression |
Heat stress tolerance |
, Katiyar-Agarw al et al. 2003
|
|
Fatty acid desaturase |
FAD7 |
A. thaliana
|
Overexpression |
Heat stress tolerance |
, Sohn et al. 2007
|
|
Polycomb group protein |
FIE1 |
A. thaliana
|
Overexpression |
Heat stress tolerance |
, Folsom et al. 2014
|
|
Stress-responsive nac gene |
SNAC3 |
Rice |
Overexpression |
Heat stress tolerance |
, Fang et al. 2015
|
Table 2Summary of heat stress-responsive core genes in reproductive tissues of rice.
Table 2
|
MSU-LOCUS ID |
RAPDB-LOCUS ID |
Description |
References |
|
Transcription factors |
|
LOC_Os02g52190 |
Os02g0759000 |
Helix-loop-helix DNA binding Protein |
, Mittal et al. 2012
|
|
LOC_Os09g35790z)
|
Os09g0526600 |
Hsfb2c |
, Mittal et al. 2012
|
|
LOC_Os07g48550 |
Os07g0684800 |
No apical meristem protein |
, Gonzalez-Schain et al. 2016
|
|
Chaperones |
|
LOC_Os01g08860 |
Os01g0184100 |
Hsp20/alpha family protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os03g16860 |
Os03g0276500 |
Dnak family protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os01g42190z)
|
Os01g0606900 |
Heat shock protein dnaj |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os02g52150z)
|
Os02g0758000 |
Heat shock 22 kda protein, mitochondrial precursor |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os02g54140z)
|
Os02g0782500 |
Hsp20/alpha crystallin family protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os03g14180z)
|
Os03g0245800 |
Hsp20/alpha crystallin family protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os03g16040z)
|
Os03g0267200 |
Hsp20/alpha crystallin family protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os04g01740z)
|
Os04g0107900 |
Heat shock protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os04g36750z)
|
Os04g0445100 |
Sp20/alpha crystallin family protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os04g45480z)
|
Os04g0538000 |
Heat shock protein STI |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os05g44340z)
|
Os05g0519700 |
Osclpb-cyt |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os06g09560z)
|
Os06g0195800 |
Heat shock protein dnaj |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os11g13980z)
|
Os11g0244200 |
Hsp20/alpha crystallin family protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
Transporters |
|
LOC_Os01g17214 |
Os01g0279400 |
Major facilitator superfamily antiporter |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os02g50680z)
|
Os02g0740300 |
AAA-type atpase family protein |
, Mittal et al. 2012
|
|
LOC_Os03g24870 |
Os03g0363600 |
Transporter family protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os03g46440 |
Os03g0667100 |
BTBA4 - Bric-a-Brac, Tramtrack, Broad Complex BTB domain with Ankyrin repeat region |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os06g39260z)
|
Os06g0593100 |
Solute carrier family 35 member B1 |
, Mittal et al. 2012
|
|
Cell Wall Modification |
|
LOC_Os01g24710 |
Os01g0348900 |
Jacalin-like lectin domain containing protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os02g44108 |
Os02g0658800 |
Expansin precursor |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os09g36060 |
Os09g0530200 |
Endoglucanase |
, Gonzalez-Schain et al. 2016
|
|
None |
Os11g0702100 |
Similar to Class III chitinase homologue (oschib3hh) (Fragment) |
, Gonzalez-Schain et al. 2016
|
|
Metabolism |
|
LOC_Os01g07530z)
|
Os01g0170000 |
Uncharacterized glycosyltransferase |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os01g27360z)
|
Os01g0371200 |
Glutathione S-transferase |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os03g59430z)
|
Os03g0808900 |
Uncharacterized glycosyltransferase |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os06g10510 |
Os06g0206900 |
Oxidoreductase transition metal ion binding protein |
, Gonzalez-Schain et al. 2016
|
|
Calcium |
|
LOC_Os01g55270z)
|
Os01g0757500 |
SGS domain containing protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os01g59530 |
Os01g0810300 |
Oscml1 - Calmodulin-related calcium sensor protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os02g15930z)
|
Os02g0259900 |
BAG6-like |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
RNA |
|
LOC_Os02g40900 |
Os02g0622500 |
RNA recognition motif containing protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os03g21160 |
Os03g0329200 |
RNA-binding zinc finger protein |
, Gonzalez-Schain et al. 2016
|
|
Others |
|
LOC_Os03g16460z)
|
Os03g0271400 |
Expressed protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
|
LOC_Os03g17790 |
Os03g0286900 |
OsRCI2-5 - Putative low temperature and saltresponsive protein |
, Gonzalez-Schain et al. 2016
|
|
LOC_Os11g31740z)
|
Os11g0518600 |
Expressed protein |
, Mittal et al. 2012; Gonzalez-Schain et al. 2016
|
Table 3Compilation of mapped QTLs and major genes tested for high temperature stress resistance.
Table 3
|
Trait |
Plant species |
QTL population |
Chromosome |
Reference |
|
Photosynthetic rate and heat tolerance tillering and heading |
Rice |
IR64 × Azucena |
6 |
, Cao et al. 2003
|
|
Heat-tolerance at grain filling |
Rice |
Nipponbare/Kasalath//Nipponbare |
3 |
, Zhu et al. 2005
|
|
Thermo-tolerance of amylose content and gel consistency |
Rice |
Nipponbare/Kasalath//Nipponbare |
9 |
, Zhu et al. 2006
|
|
High temperatures associated with white-back kernels during ripening |
Rice |
Hana-echizen × Niigata-wase |
4 |
, Tabata et al. 2007
|
|
White-back and basal-white kernels under high temperaturestress |
Rice |
Hana-echizen × Niigata-wase |
2 |
, Kobayashi et al. 2007
|
|
Teat tolerance of the tassel period |
Rice |
RIL (Zhongyouzao No. 8 × Fengjin) |
3 |
, Zhang et al. 2008
|
|
Filled grains per panicle and grain yield |
Rice |
F2 (996 × 4628) |
2 |
, Zhang et al. et al. 2009
|
|
High temperature Tolerance at seeding stage |
Rice |
F1 and F2 (HT54 × HT13) |
1 |
, Wei et al. 2013
|
|
Heat response in seedling stage. |
Rice |
Introgressed line YIL106 (Teqing × O. rufipogon) |
5 |
, Lei et al. 2013
|
|
Filled grains per panicle, unfilled grain percentage |
Rice |
BC2F2 (OM5930 × N22) |
4+2+1+1+1 |
, Buu et al. 2014
|
|
Heat tolerance in spikelet fertility |
Rice |
Bala × Azucena |
1+2+8 |
, Jagadish et al. 2010a
|
|
Heat tolerance in spikelet fertility |
Rice |
996 × 4628 |
2 |
, Xiao et al. 2011
|
|
Heat tolerance in spikelet fertility |
Rice |
IR64 × N22 |
2 |
, Ye et al. 2012
|
|
Heat tolerance during grain development |
Rice |
Tohoku-168 × Kokoromachi |
1 |
, Shirasawa et al. 2013
|
|
Anthers heat tolerance |
Rice |
(Nipponbare × Kasalath) × Nipponbare |
4 |
, Tazib et al. 2015
|
|
Spikelet sterility escape from heat stress |
Rice |
O. officinalis introgression lines |
1 |
, Hirabayashi et al. 2015
|
|
Heat tolerance in spikelet fertility |
Rice |
IR64 × N22 |
1 |
, Ye et al. 2015a,b
|
|
Heat tolerance in grain ripening |
Rice |
Tsukushiroman’ × ‘Chikushi 52’ |
6 |
, Wada et al. 2015
|
References
- Ahmed FE, Hall AE, DeMason DA. 1992. Heat injury during floral development in cowpea (Vigna unguiculata, Fabaceae). Am J Bot. 79: 784-791.
- Ainsworth EA, Ort DR. 2010. How do we improve crop production in a warming world? Plant Physiol. 54: 526-530.
- Ainsworth EA, Ort DR. 2010. How do we improve crop production in a warming world? Plant Physiol. 54: 526-530.
- Angadi SV, Cutforth HW, Miller PR, McConkey BG, Entz MH, Brandt SA, et al. 2000. Response of three Brassica species to high temperature stress during reproductive growth. Can J Plant Sci. 80: 693-701.
- Asatsuma S, Sawada C, Kitajima A, Asakura T, Mitsui T. 2006. Alpha-amylase affects starch accumulation in the rice grain. J Appl Glycosci. 53: 187-192.
- Bokszczanin KL, Fragkostefanakis S. Solanaceae Pollen Thermotolerance Initial Training Network C2013. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front Plant Sci. 4: 315
- Bray EA. 2004. Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. J Exp Bot. 55: 2331-2341.
- Brown RC, Lemmon BE, Olsen OA. 1996. Development of the endosperm in rice Oryza sativa L.: cellularization. J Plant Res. 109: 301-313.
- Buu BC, Pham TTH, Bui PT, Tran TN, Nguyen VH, Nguyen TP, et al. 2014. Quantitative trait loci associated with heat tolerance in rice (Oryza sativa L.). Plant Breed Biotechnol. 2: 14-24.
- Cao L, Zhao J, Zhan X, Li D, He L, Cheng S. 2003. Mapping QTLs for heat tolerance and correlation between heat tolerance and photosynthetic rate in rice. Chin J Rice Sci. 17: 223-227.
- Carlson RE. 1990. Heat stress, plant-available soil moisture, and corn yields in Iowa: a short and long-term view. J Prod Agric. 3: 293-297.
- Carriger S, Vallee D. 2007. More crop per drop. Rice Today. 6: 10-13.
- Chandran AKN, Jung KH. 2014. Resources for systems biology in rice. J Plant Biol. 57: 80-92.
- Chen C, Begcy K, Liu K, Folsom JJ, Wang Z, Zhang C, et al. 2016. Heat stress yields a unique MADS box transcription factor in determining seed size and thermal sensitivity. Plant Physiol. 171: 606-22.
- Cheng W, Sakai H, Yagi K, Hasegawa T. 2009. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric For Meteorol. 149: 51-58.
- Chinnusamy V, Zhu J, Zhu JK. 2007. Cold stress regulation of gene expression in plants. Trends Plant Sci. 12: 444-451.
- Chou WL, Huang LF, Fang JC, Yeh CH, Hong CY, Wu SJ, et al. 2014. Divergence of the expression and subcellular localization of CCR4-associated factor 1 CAF1 deadenylase proteins in Oryza sativa. Plant Mol Biol. 85: 443-458.
- Clarke HJ, Siddique KHM. 2004. Response of chickpea genotypes to low temperature stress during reproductive development. Field Crops Res. 90: 323-334.
- Collins NC, Tardieu F, Tuberosa R. 2008. Quantitative trait loci and crop performance under abiotic stress: Where do we stand? Plant Physiol. 147: 469-486.
- De Storme N, Geelen D. 2014. The impact of environmental stress on male reproductive development in plants: biological processes and molecular mechanisms. Plant Cell Environ. 371: 1-18.
- Devasirvatham V, Gaur PM, Mallikarjuna N, Raju TN, Trethowan RM, Tan DKY. 2013. Reproductive biology of chickpea response to heat stress in the field is associated with the performance in controlled environments. Field Crops Res. 142: 9-19.
- Devasirvatham V, Gaur PM, Mallikarjuna N, Tokachichu RN, Trethowan RM, Tan DKY. 2012. Effect of high temperature on the reproductive development of chickpea genotypes under controlled environments. Funct Plant Biol. 39: 1009-1018.
- El-Kereamy A, Bi YM, Ranathunge K, Beatty PH, Good AG, Rothstein SJ. 2012. The rice R2R3-MYB transcription factor OsMYB55 is involved in the tolerance to high temperature and modulates amino acid metabolism. PLoS ONE. 7: e52030
- Endo M, Tsuchiya T, Hamada K, Kawamura S, Yano K, Ohshima M, et al. 2009. High temperatures cause male sterility in rice plants with transcriptional alterations during pollen development. Plant Cell Physiol. 50: 1911-1922.
- Fang Y, Liao K, Du H, Xu Y, Song H, Li X, Xiong L. 2015. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J Exp Bot. 66: 6803-6817.
- Feng L, Wang K, Li Y, Tan Y, Kong J, Li H, et al. 2007. Overexpression of SBPase enhances photosynthesis against high temperature stress in transgenic rice plants. Plant Cell Rep. 26: 1635-1646.
- Firon N, Shaked R, Peet MM, Pharr DM, Zamski E, Rosenfeld K, et al. 2006. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci Hort. 109: 212-217.
- Folsom JJ, Begcy K, Hao X, Wang D, Walia H. 2014. Rice fertilization-independent endosperm1 regulates seed size under heat stress by controlling early endosperm development. Plant Physiol. 165: 238-248.
- Fujita N, Yoshida M, Kondo T, Saito K, Utsumi Y, Tokunaga T, et al. 2007. Characterization of SSIIIa-deficient mutants of rice: the function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm. Plant Physiol. 144: 2009-2023.
- Giorno F, Wolters-Arts M, Mariani C, Rieu I. 2013. Ensuring reproduction at high temperatures: the heat stress response during anther and pollen development. Plants. 2: 489-506.
- Goldberg RB, Beals TP, Sanders PM. 1993. Anther development: basic principles and practical applications. Plant Cell. 5: 1217-1229.
- Gonzalez-Schain N, Dreni L, Lawas LM, Galbiati M, Colombo L, Heuer S, et al. 2016. Genome-wide transcriptome analysis during anthesis reveals new insights into the molecular basis of heat stress responses in tolerant and sensitive rice varieties. Plant Cell Physiol. 57: 57-68.
- Hedhly A, Hormaza JI, Herrero M. 2005. The effect of temperature on pollen germination, pollen tube growth and stigmatic receptivity in peach. Plant Biol. 7: 476-483.
- Hedhly A. 2011. Sensitivity of flowering plant gametophytes to temperature fluctuations. Environ Exp Bot. 74: 9-16.
- Herrero MP, Johnson RR. 1980. High temperature stress and pollen viability of maize. Crop Sci. 20: 796-800.
- Hirabayashi H, Sasaki K, Kambe T, Gannaban RB, Miras MA, Mendioro MS, et al. 2015. qEMF3, a novel QTL for the early morning flowering trait from wild rice, Oryza officinalis, to mitigate heat stress damage at flowering in rice, O sativa. J Exp Bot. 66: 1227-1236.
- Hunter RB, Tollenaar M, Breuer CM. 1977. Effect of photoperiod and temperature on vegetative and reproductive growth of a Zea mays hybrid. Can J Plant Sci. 57: 1127-1133.
- Iba K. 2002. Acclimative response to temperature stress in higher plants: approaches of gene engineering for temperature tolerance. Annu Rev Plant Biol. 53: 225-245.
- Intergovernmental Panel on Climate Change (IPCC).2007. Climate Change 2007: Impacts, adaptation and vulnerability, 869–883. Parry ML, Canziani OF, Palutikof JP, van der Linden PJ, Hanson CE, editors. Contribution of Working Group II to Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press. Cambridge, UK.
- Ito S, Hara T, Kawanami Y, Watanabe T, Thiraporn K, Ohtake N, Sueyoshi K, et al. 2009. Carbon and nitrogen transporter during grain filling in rice under high-temperature conditions. J Agron Crop Sci. 195: 368-376.
- Jagadish SVK, Cairns JE, Lafitte R, Wheeler TR, Price AH, Craufurd PQ. 2010a. Genetic analysis of heat tolerance at anthesis in rice. Crop Sci. 50: 1633-1641.
- Jagadish SVK, Craufurd PQ, Shi W, Oane R. 2013. A phenotypic marker for quantifying heat stress impact during micro sporogenesis in rice (Oryza sativa L.). Funct Plant Biol. 41: 48-55.
- Jagadish SVK, Craufurd PQ, Wheeler TR. 2007. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J Exp Bot. 58: 1627-1635.
- Jagadish SVK, Craufurd PQ, Wheeler TR. 2008. Phenotyping parents of mapping populations of rice for heat tolerance during anthesis. Crop Sci. 48: 1140-1146.
- Jagadish SVK, Muthurajan R, Oane R, Wheeler TR, Heuer S, Bennett J, et al. 2010b. Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.). J Exp Bot. 61: 143-156.
- Jain N, Singh GP, Singh PK, Ramya P, Krishna H, Ramya KT, et al. 2014. Molecular approaches for wheat improvement under drought and heat stress. Indian J Genet Plant Breed. 744: 578-583.
- Jeong HJ, Jung KH. 2015. Rice tissue-specific promoters and condition-dependent promoters for effective translational application. J Integr Plant Biol. 5711: 913-24.
- Kaneko K, Sasak M, Kuribayashi N, Suzuki H, Sasuga Y, Shiraya T, et al. 2016. Proteomic and glycomic characterization of rice chalky grains produced under moderate and high-temperature conditions in field system. Rice. 9: 26
- Katiyar-Agarwal S, Agarwal M, Grover A. 2003. Heat-tolerant basmati rice engineered by over-expression of hsp101. Plant Mol Biol. 51: 677
- Kaushal N, Awasthi R, Gupta K, Gaur P, Siddique KHM, Nayyar H, et al. 2013. Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct Plant Biol. 40: 1334-349.
- Kawagoe Y, Kubo A, Satoh H, Takaiwa F, Nakamura Y. 2005. Roles of isoamylase and ADP-glucose pyrophosphorylase in starch granule synthesis in rice endosperm. Plant J. 42: 164-174.
- Kim SY, Hong CB, Lee I. 2001. Heat shock stress causes stage specific male sterility in Arabidopsis thaliana. J Plant Res. 114: 301-307.
- Kobayashi A, Genliang B, Shenghai Y, Tomita K. 2007. Detection of quantitative trait loci for white-back and basal-white kernels under high temperature stress in japonica rice varieties. Breed Sci. 57: 107-116.
- Koh S, Lee SC, Kim MK, Koh JH, Lee S, An G, et al. 2007. T-DNA tagged knockout mutation of rice OsGSK1, an orthologue of Arabidopsis BIN2, with enhanced tolerance to various abiotic stresses. Plant Mol Biol. 65: 453-466.
- Köhler C, Page DR, Gagliardini V, Grossniklaus U. 2005. The Arabidopsis thaliana MEDEA Polycomb group protein controls expression of PHERES1 by parental imprinting. Nat Genet. 37: 28-30.
- Kotak S, Larkindale J, Lee U, von Koskull-Doring P, Vierling E, Scharf KD. 2007. Complexity of the heat stress response in plants. Curr Opin Plant Biol. 10: 310-316.
- Lee SK, Hwang SK, Han M, Eom JS, Kang HG, Han Y, et al. 2007. Identification of the ADP-glucose pyrophosphorylase isoforms essential for starch synthesis in the leaf and seed endosperm of rice Oryza sativa L. Plant Mol Biol. 65: 531-546.
- Lei D, Tan L, Liu F, Chen L, Sun C. 2013. Identification of heat-sensitive QTL derived from common wild rice (Oryza rufipogon Griff.). Plant Sci. 201: 121-127.
- Li X, Lawas LM, Malo R, Glaubitz U, Erban A, Mauleon R, et al. 2015a. Metabolic and transcriptomic signatures of rice floral organs reveal sugar starvation as a factor in reproductive failure under heat and drought stress. Plant Cell Environ. 38: 2171-2192.
- Li XM, Chao DY, Wu Y, Huang X, Chen K, Cui LG, et al. 2015b. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nat Genet. 47: 827-833.
- Lin CJ, Li CY, Lin SK, Yang FH, Huang JJ, Liu YH, et al. 2010. Influences of high temperature during grain filling on the accumulation of storage proteins and grain quality in rice (Oryza sativa L.). J Agric Food Chem. 58: 10545-10552.
- Lin SK, Chang MC, Tsai YG, Lur HS. 2005. Proteomic analysis of the expression of proteins related to rice quality during caryopsis development and the effect of high temperature on expression. Proteomics. 5: 2140-2156.
- Lin Z, Zhang X, Yang X, Li G, Tang S, Wang S, et al. 2014. Proteomic analysis of proteins related to rice grain chalkiness using iTRAQ and a novel comparison system based on a notched-belly mutant with white-belly. BMC Plant Biol. 14: 163
- Liu X, Guo T, Wan X, Wang H, Zhu M, Li A, et al. 2010. Transcriptome analysis of grain-filling caryopses reveals involvement of multiple regulatory pathways in chalky grain formation in rice. BMC Genomics. 11: 730
- Liu X, Wan X, Ma X, Wan J. 2011. Dissecting the genetic basis for the effect of rice chalkiness, amylose content, protein content, and rapid viscosity analyzer profile characteristics on the eating quality of cooked rice using the chromosome segment substitution line population across eight environments. Genome. 54: 64-80.
- Mathur S, Agrawal D, Jajoo A. 2014. Photosynthesis: limitations in response to high temperature stress. J Photochem Photobiol B. 137: 116-126.
- McCormick S. 2004. Control of male gametophyte development. Plant Cell. 16: S142-S153.
- Meuwissen TH, Hayes BJ, Goddard ME. 2001. Prediction of total genetic value using genome-wide dense marker maps. Genetics. 157: 1819-1829.
- Mittal D, Madhyastha DA, Grover A. 2012. Genome-wide transcriptional profiles during temperature and oxidative stress reveal coordinated expression patterns and overlapping regulons in rice. PLoS ONE. 77: e40899
- Mizutani M, Ohta D. 2010. Diversification of P450 genes during land plant evolution. Annu Rev Plant Biol. 61: pp. 291-315.
- Mohammed A-R, Tarpley L. 2009a. Impact of high nighttime temperature on respiration, membrane stability, antioxidant capacity, and yield of rice plants. Crop Sci. 49: 313-322.
- Mohammed AR, Tarpley L. 2009b. High nighttime temperatures affect rice productivity through altered pollen germination and spikelet fertility. Agri Forest Meteorol. 149: 999-1008.
- Mohammed AR, Tarpley L. 2010. Effects of high night temperature and spikelet position on yield-related parameters of rice Oryza sativa L. plants. Eur J Agron. 33: 117-123.
- Morrison MJ. 1993. Heat stress during reproduction in summer rape. Can J Bot. 71: 303-308.
- Murakami T, Matsuba S, Funatsuki H, Kawaguchi K, Saruyama H, Tanida M, et al. 2004. Over-expression of a small heat shock protein, sHSP17. 7, confers both heat tolerance and UV-B resistance to rice plants. Mol Breed. 13: 165-175.
- Nagato K, Ebata M. 1960. Effects of temperature in the ripening periods upon the development and qualities of lowland rice kernels. Proc Crop Sci Soc Jpn. 28: 275-278.
- Nakagawa H, Horie T, Matsui T. 2002. Effects of climate change on rice production and adaptive technologies. pp. 635-657. Mew TW, Brar DS, Peng S, Dawe D, Hardy B, editors. Rice science: innovations and impact for livelihood. IRRI. Los Baños, Philippines.
- Nguyen DN, Lee KJ, Kim DI, Anh NT, Lee BW. 2013. Modeling and validation of high-temperature induced spikelet sterility in rice. Field Crops Res. 156: 293-302.
- Nishi A, Nakamura Y, Tanaka N, Satoh H. 2001. Biochemical and genetic analysis of the effects of amylase-extender mutation in rice endosperm. Plant Physiol. 127: 459-472.
- Nuttal WF, Moulin AP, Townley-Smith LJ. 1992. Yield response of canola to nitrogen, phosphorus, precipitation, and temperature. Agr J. 84: 765-768.
- Olsen OA. 2001. Endosperm development: cellularization and cell fate specification. Annu Rev Plant Physiol Plant Mol Biol. 52: 233-267.
- Oort PAJ, Saito K, Zwart SJ, Shrestha S. 2014. A simple model for simulating heat 25 induced sterility in rice as a function of flowering time and transpirational cooling. Field Crops Res. 156: 303-312.
- Ortiz R, Sayre KD, Govaerts B, Gupta R, Subbarao GV, Ban T, et al. 2008. Climate change: can wheat beat the heat? Agr Ecosyst Environ. 126: 46-58.
- Oshino T, Abiko M, Saito R, Ichiishi E, Endo M, Kawagishi-Kobayashi M, et al. 2007. Premature progression of anther early developmental programs accompanied by comprehensive alterations in transcription during high-temperature injury in barely plants. Mol Genet Genom. 278: 31-42.
- Peet MM, Sato S, Gardner RG. 1998. Comparing heat stress effects on male-fertile and male-sterile tomatoes. Plant Cell Environ. 21: 225-231.
- Polowick PL, Sawhney VK. 1988. High-temperature-induced male and female sterility in canola (Brassica napus L.). Ann Bot. 62: 83-86.
- Prasad PVV, Boote KJ, Allen LH, Sheehy JE, Thomas JMG. 2006. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 95: 398-411.
- Qi Y, Wang H, Zou H, Liu Y, Wang Y, Zhang W. 2011. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 585: 231-239.
- Rabbani MA, Maruyama K, Abe H, Khan MA, Katsura K, Ito Y, et al. 2003. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol. 133: 1755-1767.
- Rana RM, Dong S, Tang H, Ahmad F, Zhang H. 2012. Functional analysis of OsHSBP1 and OsHSBP2 revealed their involvement in the heat shock response in rice (Oryza sativa L.). J Exp Bot. 63: 6003-6016.
- Rang ZW, Jagadish SVK, Zhoua QM, Craufurd PQ, Heuer S. 2010. Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ Exp Bot. 70: 58-65.
- Ribaut JM, Vicente MC, Delannay X. 2010. Molecular breeding in developing countries: challenges and perspectives. Curr Opin Plant Biol. 13: 1-6.
- Rieu I, Twell D, Firon N. 2017. Pollen development at high temperature: from acclimation to collapse. Plant Physiol. 173: 1967-1976.
- Rodrigo J, Herrero M. 2002. Effects of pre-blossom temperatures on flower development and fruit set in apricot. Sci Hortic. 92: 125-135.
- Sabelli PA, Larkins BA. 2009. The development of endosperm in grasses. Plant Physiol. 149: 14-26.
- Saini HS, Sedgley M, Aspinall D. 1983. Effect of heat stress during floral development on pollen tube growth and ovary anatomy in wheat (Triticum aestivum L.). Aus J Plant Physiol. 10: 137-144.
- Sakata T, Higashitani A. 2008. Male sterility accompanied with abnormal anther development in plants genes and environmental stresses with special reference to high temperature injury. Int J Plant Dev Biol. 2: 42-45.
- Sato S, Peet MM, Thomas JF. 2002. Determining critical pre and post anthesis periods and physiological processes in Lycopersicon esculentum Mill exposed to moderately elevated temperatures. J Exp Bot. 53: 1187-1195.
- Savin R, Stone PJ, Nicolas ME. 1996. Responses of grain growth and malting quality of barley to short periods of high temperature in field studies using portable chambers. Aust J Agrit Res. 47: 465-477.
- Shen H, Zhong X, Zhao F, Wang Y, Yan B, Li Q, et al. 2015. Overexpression of receptor-like kinase ERECTA improves thermotolerance in rice and tomato. Nat Biotechnol. 33: 996-1003.
- Shirasawa K, Sekii T, Ogihara Y, Yamada T, Shirasawa S, Kishitani S, et al. 2013. Identification of the chromosomal region responsible for high-temperature stress tolerance during the grain-filling period in rice. Mol Breed. 32: 223-232.
- Shiraya T, Mori T, Maruyama T, Sasaki M, Takamatsu T, Oikawa K, et al. 2015. Golgi/plastid-type manganese superoxide dismutase involved in heat-stress tolerance during grain filling of rice. Plant Biotechnol J. 13: 1251-1263.
- Sohn SO, Back K. 2007. Transgenic rice tolerant to high temperature with elevated contents of dienoic fatty acids. Biol Plantarum. 51: 340-342.
- Spindel J, Begum H, Akdemir D, Virk P, Collard B, Redoña E, et al. 2015. Genomic selection and association mapping in rice (Oryza sativa): effect of trait genetic architecture, training population composition, marker number and statistical model on accuracy of rice genomic selection in elite, tropical rice breeding lines. PLoS Genet. 11: e1004982
- Srinivasan A, Saxena NP, Johansen C. 1999. Cold tolerance during early reproductive growth of chickpea (Cicer arietinum L.): genetic variation in gamete development and function. Field Crops Res. 60: 209-222.
- Tabata M, Hirabayashi H, Takeuchi Y, Ando I, Iida Y, Ohsawa R. 2007. Mapping of quantitative trait loci for the occurrence of white-back kernels associated with high temperatures during the ripening period of rice (Oryza sativa L.). Breeding Sci. 57: 47-52.
- Takeoka Y, Hiroi K, Kitano H, Wada T. 1991. Pistil hyperplasia in rice spikelets as affected by heat stress. Plant Reprod. 4: 39-43.
- Tazib T, Kobayashi Y, Koyama H, Matsui T. 2015. QTL analyses for anther length and dehiscence at flowering as traits for the tolerance of extreme temperatures in rice (Oryza sativa L.). Euphytica. 203: 629-642.
- Tian J, Belanger FC, Huang B. 2009. Identification of heat stress-responsive genes in heat-adapted thermal Agrostisscabra by suppression subtractive hybridization. J Plant Physiol. 166: 588-601.
- Varshney RK, Dubey A. 2009. Novel genomic tools and modern genetic and breeding approaches for crop improvement. J Plant Biochem Biotech. 18: 127-138.
- Wada T, Miyahara K, Sonoda JY, Tsukaguchi T, Miyazaki M, Tsubone M, et al. 2015. Detection of QTLs for white-back and basal-white grains caused by high temperature during ripening period in japonica rice. Breeding Sci. 65: 216-225.
- Wahid A, Gelani S, Ashraf M, Foolad MR. 2007. Heat tolerance in plants: an overview. Environ Exp Bot. 61: 199-233.
- Wang E, Wang J, Zhu X, Hao W, Wang L, Li Q, et al. 2008. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet. 40: 1370-1374.
- Wassmann R, Jagadish SVK, Heuer S, Ismail A, Redona E, Serraj R, et al. 2009. Climate change affecting rice production: the physiological and agronomic basis for adaptation strategies. Adv Agron. 101: 59-122.
- Wei H, Liu J, Wang Y, Huang N, Zhang X, Wang L, et al. 2013. A dominant major locus in chromosome 9 of rice (Oryza sativa L.) confers tolerance to 48°C high temperature at seedling stage. J Hered. 104: 287-294.
- Whittle CA, Otto SP, Johnston MO, Krochko JE. 2009. Adaptive epigenetic memory of ancestral temperature regime in Arabidopsis thaliana. Botany. 87: 650-657.
- Woo MO, Ham TH, Ji HS, Choi MS, Jiang W, Chu SH, et al. 2008. Inactivation of the UGPase1 gene causes genic male sterility and endosperm chalkiness in rice (Oryza sativa L.). Plant J. 54: 190-204.
- Wu X, Liu J, Li D, Liu CM. 2016a. Rice caryopsis development II: Dynamic changes in the endosperm. J Integr Plant Biol. 58: 786-798.
- Wu X, Liu J, Li D, Liu CM. 2016b. Rice caryopsis development I: Dynamic changes in different cell layers. J Integr Plant Biol. 58: 772-785.
- Wu X, Shiroto Y, Kishitani S, Ito Y, Toriyama K. 2009. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 28: 21-30.
- Xiao YH, Pan Y, Luo LH, Deng HB, Zhang GL, Tang WB, et al. 2011. Quantitative trait loci associated with pollen fertility under high temperature stress at flowering stage in rice (Oryza sativa). Rice Sci. 18: 204-209.
- Yamaguchi-Shinozaki K, Shinozaki K. 2006. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu Rev Plant Biol. 57: 781-803.
- Yamakawa H, Hakata M. 2010. Atlas of rice grain filling-related metabolism under high temperature: joint analysis of metabolome and transcriptome demonstrated inhibition of starch accumulation and induction of amino acid accumulation. Plant Cell Physiol. 51: 795-809.
- Yamakawa H, Hirose T, Kuroda M, Yamaguchi T. 2007. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 144: 258-277.
- Yamamoto E, Yonemaru JI, Yamamoto T, Yano M. 2012. OGRO: the overview of functionally characterized genes in rice online database. Rice. 5: 1-10.
- Yamanouchi U, Yano M, Lin H, Ashikari M, Yamada K. 2002. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA. 99: 7530-7535.
- Ye C, Argayoso MA, Redoña ED, Sierra SN, Laza MA, Dilla CJ, et al. 2012. Mapping QTL for heat tolerance at flowering stage in rice using SNP markers. Plant Breed. 131: 33-41.
- Ye C, Tenorio FA, Argayoso MA, Laza MA, Koh H, Redoña ED, et al. 2015a. Identifying and confirming quantitative trait loci associated with heat tolerance at flowering stage in different rice populations. BMC Genet. 16: 41
- Ye C, Tenorio FA, Redoña ED, Morales-Cortezano PS, Cabrega GA, Jagadish KS, et al. 2015b. Fine-mapping and validating qHTSF4.1 to increase spikelet fertility under heat stress at flowering in rice. Theor Appl Genet. 128: 1507-1517.
- Yoshida S, Satake T, Mackill DS. 1981. High temperature stress in rice. IRRI Research Paper series. IRRI. Manila, Philippines: 67: pp. 1-15.
- Young LW, Wilen RW, Bonham-Smith PC. 2004. High temperature stress of Brassica napus during flowering reduces micro- and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J Exp Bot. 55: 485-495.
- Zhang GL, Chen LY, Xiao GY, Xiao YH, Chen XB. 2009. Bulked segregant analysis to detect QTL related to heat tolerance in rice (Oryza sativa L.) using SSR markers. Agr Sci China. 8: 482-487.
- Zhang T, Yang L, Jiang K, Huang M, Sun Q, Chen W, et al. 2008. QTL mapping for heat tolerance of the tassel period of rice. Mol Plant Breed. 6: 867-873.
- Zhang XW, Li JP, Liu AL, Zou J, Zhou XY, Xiang JH, et al. 2012. Expression profile in rice panicle: insights into heat response mechanism at reproductive stage. PLoS ONE. 7: e49652
- Zhu C, Jiang L, Zhang W, Wang C, Zhai H, Wan J, et al. 2006. Identifying QTLs for thermo--tolerance of amylose content and gel consistency in rice. Chin J Rice Sci. 20: 248-252.
- Zhu CL, Ying-hui X, Chun-ming W, Ling J, et al. 2005. Mapping QTL for heat-tolerance at grain filling stage in rice. Rice Sci. 12: 33-38.
- Zinn KE, Tunc-Ozdemir M, Harper JF. 2010. Temperature stress and plant sexual reproduction: uncovering the weakest links. J Exp Bot. 61: 1959-1968.