Abstract
Low temperature germinability (LTG) is an important trait for stand establishment in the direct-seeding method of rice cultivation. In temperate growing regions, water temperature during sowing season is frequently below 15°C resulting in poor crop establishment. The objective of this study was to select enhanced rice germplasm for low temperature germinability. Association of the phenotype for LTG with the genotype for qLTG3-1, a major QTL for LTG, in japonica rice accessions (n = 180) from East Asia was conducted. A highly significant association was observed between qLTG3-1 alleles and tolerant (χ2 = 56.617, P = 5.08E-13) or sensitive (χ2 = 32.844, P = 7.38E-08) accessions. Association of genotype and phenotype suggested the germplasm panel used in this study were closely associated with cold environment at the germination stage. Genotypic data from 32 SNP markers derived from previously reported LTG QTLs showed a mean polymorphism information content (PIC) of 0.248 and an average gene diversity of 0.307. A neighbor-joining tree clustered 23 tolerant accessions into one group and the genotypic data for four of the markers revealed that all of these accessions had the alternative allele for the qLTG3-1 and qLTG2-6 markers and the reference allele at the qLTG11-1 marker. Sensitive accessions clustered into one group and harbored the reference allele at the qLTG4-3 marker. Finally, ten accessions were selected after phenotypic and genotypic evaluation for LTG and these temperate japonica cultivars or breeding lines may contribute to the breeding of rice varieties for direct-seeding systems.
-
Key words: Low temperature germinability, Rice, Selection, SNP, Marker
INTRODUCTION
The direct-seeding of rice is becoming more widespread in many Asian countries as traditional transplanting is very labor-intensive and increases the cost of rice culture. Success of the direct-seeding method greatly depends on the control of weeds and good crop establishment which results from early stage vigor (i.e., germination and seedling growth). Low temperature germinability (LTG) is an important trait for crop establishment. In temperate rice growing areas such as Korea, northern China, Japan and high altitude regions in the tropics and subtropics, water temperature at sowing season is frequently below 15°C. In South Korea for example, the average lowest temperature in May during the period 1981–2010 in the central region of Suwon was 12°C (Korea Meteorological Administration). This unfavorable temperature causes poor establishment and non-uniform maturation as the optimum temperature for rice germination is between 25 to 35°C. Development of enhanced LTG varieties would address this constraint of the direct-seeding method and should lead to improved crop establishment and decreases in the associated yield loss.
Previously, we performed phenotypic and genotypic evaluation for low temperature germinability in a germplasm panel of 180
japonica rice accessions from temperate areas of East Asia (South Korea, Japan, China and North Korea). The germplasm showed significant variation in seed germination rate and vigor under low temperature conditions (
Hyun et al. 2015). Among the four groups, accessions from Japan and N. Korea showed higher germination rates than those from S. Korea and China. A high proportion (73%) of the low-temperature tolerant (i.e., strong LTG) accessions carried the Italica Livorno (IL) allele for
qLTG3-1 compared to the sensitive (i.e., weak LTG) accessions where Hayamasari (HY) allele was present in 70% of those germplasm. The LTG phenotyping was consistent with the genotyping for
qLTG3-1, which indicated that the germplasm from Japan and N. Korea had a higher occurrence of the IL allele than those from China and S. Korea.
Fujino and Iwata (2011) examined the distribution of
qLTG3-1 alleles for Hokkaido rice germplasm and found that cultivars harboring the IL allele (80.8%) showed more vigorous low-temperature germinability than those with the HY allele (30.3%). Similar results were reported by
Challam et al. (2013) who carried out a survey of the IL and HY allele in 65 diverse Indian rice accessions. However, several landraces from regions which do not experience low temperatures had the IL allele, consistent with the idea that this gene may not specifically address low-temperature germination (
Fujino et al. 2008) and suggesting that additional markers are needed for evaluation of LTG in diverse rice germplasm.
Single nucleotide polymorphisms (SNPs) currently have become the most popular genetic markers because they are stable, efficient and cost-effective for automation (
Rafalski 2002;
Duran et al. 2009;
Edwards and Batley 2010) and widely used in many breeding and research programs for applications such as the assessment and selection of germplasm for target traits. Recently, we developed four SNP markers for LTG from previously reported LTG QTLs located on chromosome 2, 4 and 11 and using sequencing data from 60 germplasm accessions (
Hyun et al. 2015). The markers,
qLTG2-6,
qLTG4-3,
qLTG4b-1 and
qLTG11-1, were strongly associated with LTG and their
R2 values were 0.175, 0.091, 0.190 and 0.083, respectively (
Hyun et al. 2015). The comparison of the genotypes for the each of 25 tolerant and sensitive accessions for these four marker loci revealed that in tolerant (strong LTG) accessions the alternative allele is prevalent among the
qLTG2-6 and
qLTG4-3 loci while the reference allele occurs more frequently at
qLTG4b-1 and
qLTG11-1 loci. In the sensitive (weak LTG) group, only the reference allele was observed at the
qLTG4-3 locus and most of the alleles were of the reference and alternative type at the
qLTG2-6 and
qLTG4b-1 loci, respectively (
Hyun et al. 2015). It is suggested that the genotypes for these markers may be useful reference for screening of LTG with use in combination with the
qLTG3-1 marker in
temperate japonica rice germplasm.
In this study, we have performed association of LTG phenotype with the qLTG3-1 alleles using the data previously examined in a germplasm panel of 180 temperate japonica rice accessions. Also, genotypic assessment for SNP markers designed from regions harbored LTG QTLs was conducted for determining of genetic distance among the accessions. We generated neighbor-joining tree and identified clustered groups. Comparison of the genotypes for four SNP markers was carried out in tolerant and sensitive accessions. Tolerant germplasm were selected based on the clustering and genotype patterns and these may be used as a reference for screening and selecting germplasm for using in breeding program for direct-seeding varieties.
MATERIALS AND METHODS
Plant Materials and LTG evaluation
One hundred and eighty
temperate japonica rice germ-plasm, consisting of advanced breeding lines and modern varieties conserved at the National Agrobiodiversity Center (Jeonju), were selected for evaluation of LTG. The germplasm was consisted of 63 accessions from North Korea, 58 from China, 20 from Japan, and 39 from South Korea (
Table 1). LTG evaluation of the germplasm was reported previously (
Hyun et al. 2015).
To determine the association between the allele distribution of
qLTG3-1 and the LTG phenotype, the frequencies with which
qLTG3-1 alleles were associated with LTG phenotype (i.e., tolerant, moderate and sensitive) were calculated using the previously reported data (
Hyun et al. 2015). The allelic frequencies were also examined with regard to the country of origin of the accessions (N. Korea, Japan, China and S. Korea) to identify the association of these alleles with the country of origin. Significant differences in allele frequencies within the country of origin were evaluated using the χ
2 test.
As described previously (
Hyun et al. 2015), 32 SNP markers were selected from regions reported to harbor LTG QTLs and from the sequencing of 60 germplasm accessions. The SNP marker data from the evaluation of the 180 germplasm (
Hyun et al. 2015) were analyzed using PowerMarker version 3.25 software (
Liu and Muse 2005). Major allele frequency, gene diversity and polymorphism information content (PIC) values for each SNP marker were calculated. The PIC value described by
Bostein et al. (1980) and modified by
Anderson et al. (1993) was defined as follows:
where p
ij is the frequency of the jth allele for marker i, and summation extends over n alleles. A neighbor-joining tree for evaluation of genetic distance between the accessions was constructed using Nei distance (
Nei and Takezaki 1983) and MEGA version 6.0 software (
Tamura et al. 2013) was used to visualize the tree.
One hundred and forty one germplasm except S. Korea accessions were assessed for agronomic characters at Iksan (N. Korea accessions) and Namyang (Japan and China accessions), Korea in 2012. Germinated seeds were sown in May 1 and May 5 and 4-week-old seedlings were then transplanted in June 1 and June 5 at Namyang and Iksan, respectively. A total of 25 seedlings for each accession were transplanted with spacing of 30 cm × 15 cm between the two plants. Agronomic traits such as days to heading, plant height and panicle length were characterized at maturity and 1,000-grain weight was measured after seeds harvest and cleaning. All traits were recorded in replicates following the standard protocol (
RDA 2012).
RESULTS
Association of qLTG3-1 allele with LTG phenotype
There was a highly significant association of the IL allele with LTG tolerant accessions (χ
2 = 56.617, P = 5.08E-13) and the HY allele with sensitive accessions (χ
2 = 32.844, P = 7.38E-08). No significant association was observed in moderate accessions (
Table 2). Association of genotype and phenotype by the origin indicates that all germplasm were significantly associated with
qLTG3-1 alleles. In particular, the HY and IL alleles exhibited a highly significant association with germplasm from S. Korea and Japan, respectively, which supports our previous findings that germination rate of S. Korea and Japan accessions was the lowest and highest among all groups (
Hyun et al. 2015).
A total of 31 markers derived from chromosome 2, 4, 7, 9 and 11, where 2 or more QTLs were overlapped, were used for genotyping and
qLTG3-1 was used as a control. The statistical values for each marker with 180 germplasm are given in
Table 3. The average major allele frequency was 0.773, ranging from 0.508 to 0.966. The average gene diversity was 0.307, ranging from 0.065 to 0.499. The average polymorphism information content (PIC) was 0.248, ranging from 0.063 to 0.375.
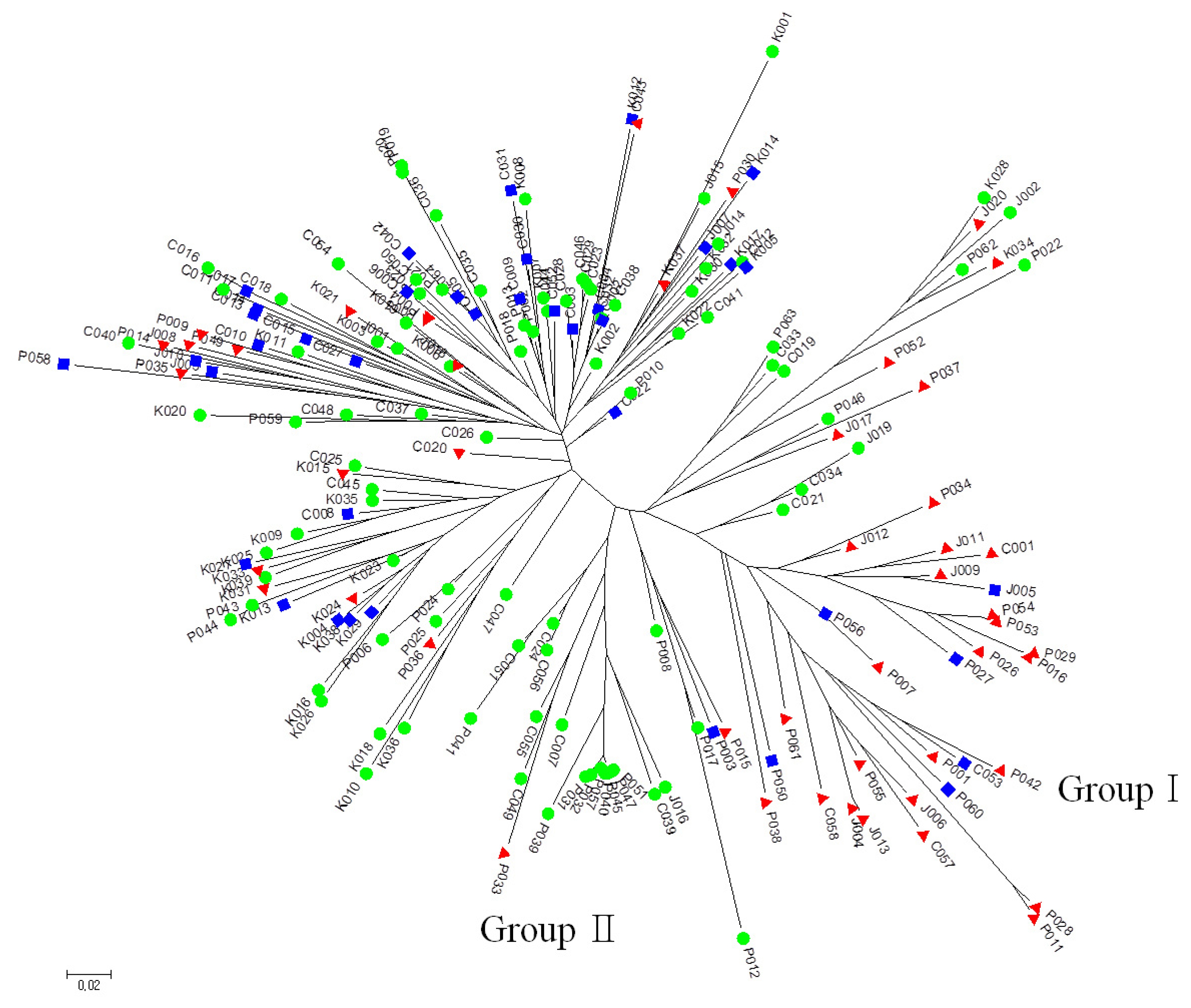
The SNP genotype data for 180 germplasm across the 32 SNP loci were used for generating a neighbor-joining tree based on Nei’s genetic distance (
Fig. 1). It revealed that 23 tolerant and 6 moderate accessions were clustered together (Group I) consisting of 18 accessions from N. Korea, 7 Japan, and 4 China. Two of the accessions from China (C057, C058) showed the highest germination rate and germination vigor score. Seventeen sensitive accessions were clustered into Group II, which also contained one tolerant accession. There were no other groups clustered by LTG phenotype (i.e., tolerant, moderate and sensitive) or germplasm origin. Of the 29 accessions in Group I, 28 accessions had the IL allele for
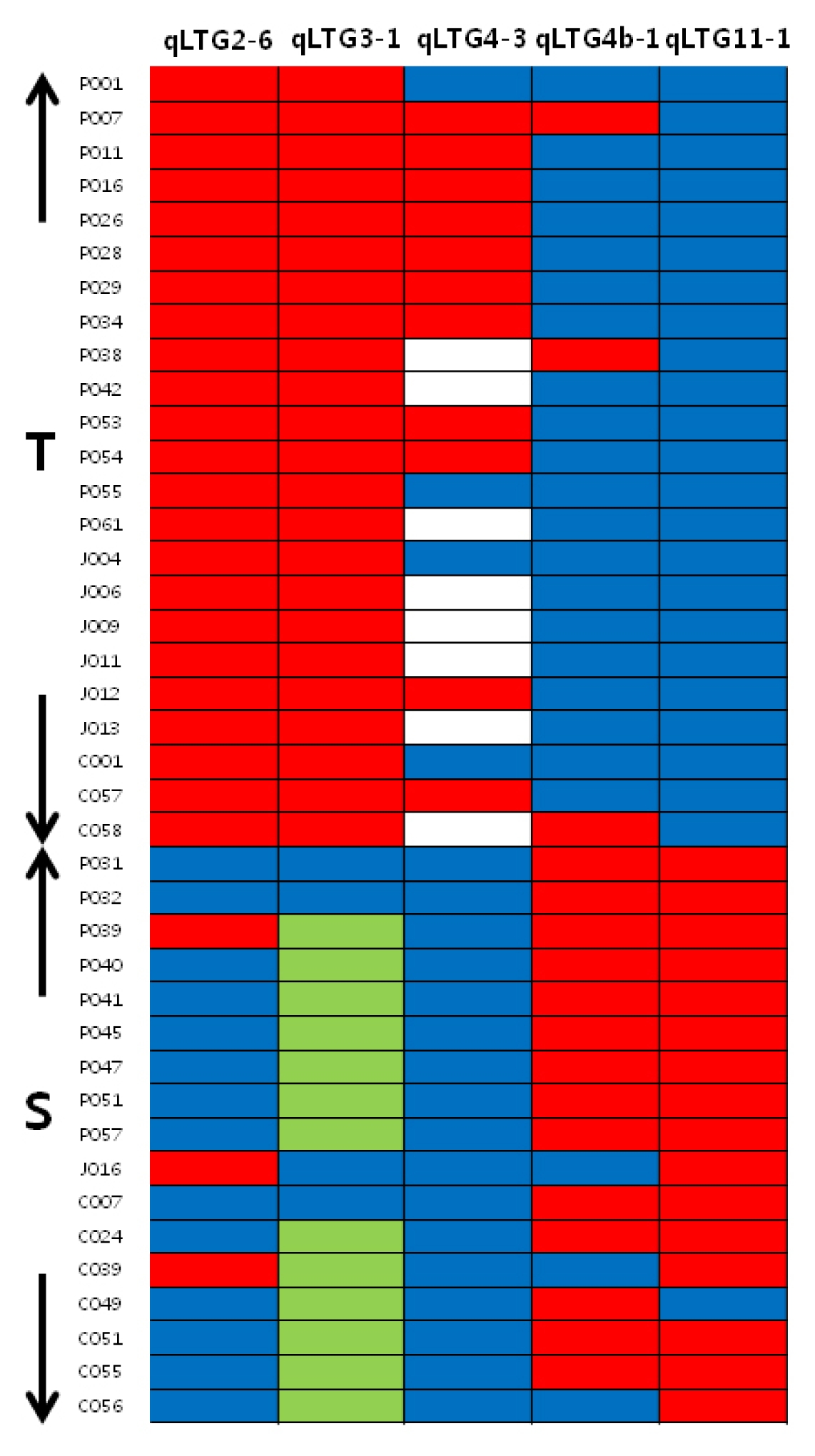
qLTG3-1 marker and 12 N. Korea accessions exhibited vigorous growth during the seedling stage. Based on the cluster analysis result, the comparison of the genotype between 23 tolerant and 17 sensitive accessions at five loci revealed that tolerant accessions had same allele type in
qLTG2-6,
qLTG3-1 and
qLTG11-1 (
Fig. 2). Twenty of the tolerant accessions also had the same allele type in
qLTG4b-1. In the sensitive group, most alleles were of the reference type for the
qLTG2-6 locus and the alternative type at the
qLTG4b-1 and
qLTG11-1 locus. All alleles were the reference type at the
qLTG4-3 locus (
Fig. 2).
A total of ten germplasm including 6 accessions from N. Korea, 2 from Japan, and 2 from China were selected for LTG based on the phenotypic and genotypic data (
Table 4). They are clustered into Group I and have the same alleles at
qLTG2-6,
qLTG3-1, and
qLTG11-1 loci. All selected accessions showed high germination rate of over 85% under low temperature conditions. The two accessions from China, W-14 and W-15, were exceptional, showing a 100% germination rate. Agronomic traits of these accessions were previously characterized in 2012 at Iksan (N. Korea accessions) and Namyang (Japan and China accessions). Among the accessions selected for strong LTG, days to heading ranged from a minimum of 54 days for DPKR 036S 11685 to a maximum of 74 days for Hosakan and plant height from minimum of 40 cm for Tokachikuromomi to maximum of 97 cm for DPKR 036S 11721 which also had the longest panicle length (24 cm). Relatively short panicle lengths (15 cm) were observed in three accessions, Tokachikuromomi, W-14 and W-15. Panicles from the variety Hosakan, which was the second shortest accession, were of moderate length (20 cm) among these accessions. The N. Korea accession DPKR 036S 11713 showed the highest 1,000-grain weight of 41.1 g. Another N. Korea accession DPKR 036S 432 also exhibited a high 1,000-grain weight of 37.5 g. Germplasm from N. Korea showed higher 1,000-grain weight than those from Japan and China. The mean 1,000-grain weight was 32.1 g for N. Korea accessions and 27.1 g and 25.4 g for Japan and China, respectively.
DISCUSSION
Low temperature germinability is an important trait for breeding rice varieties for direct-seeding production systems. Phenotypic and genotypic evaluation for LTG was previously performed on 180
temperate japonica rice accessions originating from S. Korea, China, Japan, and N. Korea. Most of these germplasm were modern varieties or advanced breeding lines which are suitable materials for use in breeding programs. In our previous study, genotyping for
qLTG3-1 alleles revealed 73% of tolerant and 70% of sensitive germplasm harbored IL and HY alleles, respectively (
Hyun et al. 2015). In this study, our analysis showed that both alleles were significantly associated with tolerant (χ
2 = 56.617, P = 5.08E-13) and sensitive (χ
2 = 32.844, P = 7.38E-08) phenotypes (
Table 2).
Challam et al. (2013) also showed the association of
qLTG3-1 genotype with LTG phenotype in diverse rice germplasm from the India, which the insertion allele (IL or NB) and the deletion allele (HY) were associated with tolerance (χ
2 = 2.812, P = 0.094) and susceptibility (χ
2 = 5.196, P = 0.019), respectively. The genotype-phenotype association for LTG in the
temperate japonica germplasm used in this study was much more significant than that observed in diverse Indian germplasm, suggesting that the genotypes of 180 germplasm panel originated from East Asia were more closely associated with cold environment at germination stage. In Indian germplasm, the IL allele was detected in the varieties or landraces which were not grown or developed in low temperature environments (
Challam et al. 2013). However, rice varieties growing in East Asia are exposed to low temperature at germination stage and breeders have selected tolerance varieties or breeding lines to this stress artificially. This process may lead to higher association of the
qLTG3-1 genotype with LTG phenotype in this region than in India. It was revealed that
qLTG3-1 alleles contributed to the tolerance for low temperature germinability among rice cultivars from Hokkaido and the selection pressure on this gene was focused during rice breeding program in this region (
Fujino and Iwata, 2011), suggesting that
qLTG3-1 gene is a perfect marker for LTG in
temperate japonica rice cultivars or landraces. However, two accessions, Milyang260 and Josaengheugchal, carrying the IL allele were identified as sensitive to LTG in our previous study. These Tongil-type varieties are derived from the cross of
japonica and
indica subspecies, indicating that new markers are needed to evaluate LTG in diverse germplasm panels (
Hyun et al. 2015).
A total of 32 SNP markers selected from previously reported QTLs for LTG were used for genotyping 180 germplasm (
Hyun et al. 2015). In this study, the statistical values for these markers were examined. The polymorphic information content (PIC) values were shown to range from 0.248 to 0.375 (
Table 3). PIC values from our study using SNP markers were relatively lower than those from other reports using SSR or STS markers. The average PIC value of 0.74, ranging from 0.17 to 0.92 was reported in the world collections of rice, and the average PIC value of 0.50, ranging from 0.02 to 0.88 in the US collection (
Xu et al. 2005).
Agrama and Eizenga (2008) reported that the highest PIC value of 0.78 was shown in the wild relatives (
Oryza spp.) represented by ten different species, while international and US cultivars had values of 0.57 and 0.39, respectively. The PIC value of a rice collection in China ranged from 0.0466 to 0.7216 with a mean of 0.4214 (
Jin et al. 2010). The average gene diversity in this study was 0.307, ranging from 0.065 to 0.499 (
Table 3). The international and US cultivars had an average gene diversity values of 0.61 and 0.43, respectively (
Agrama and Eizenga 2008). The China accessions had a value of 0.4736 (
Jin et al. 2010).
Gonzaga et al. (2015) reported that the maximum PIC value of SSRs (0.563–0.830) was higher than those of SNPs (0.490–0.580) which is due to the multi-allelic nature of SSR polymorphism. The average PIC values of SNP markers ranged from 0.289 to 0.382 (
Gonzaga et al. 2015), which is similar value to our result. It is suggested that lower PIC values we observed might result from a narrow germplasm panel consisting of East Asia
temperate japonica or use of SNP markers.
The neighbor-joining tree showed that most of accessions were clustered with mixed phenotypes, while 23 tolerant accessions were grouped with several moderate gemplasm (Group I) and 17 sensitive accessions were clustered into a different group (Group II) (
Fig. 1). All accessions except one in Group I had the IL allele. The comparison of the genotype pattern between these groups using four newly developed markers for LTG (
Hyun et al. 2015) revealed that all tolerant accessions carried the alternative allele for
qLTG2-6 and
qLTG3-1 and the reference allele for
qLTG11-1 loci, while only the alternative allele was identified in all sensitive accessions for
qLTG4-3 locus (
Fig. 2).
qLTG2-6 was derived from the region containing the QTLs
qLTG-2-1 (
Ji et al. 2008) and
qGR-2 (
Ji et al. 2009), which was expressed mainly during the early germination period (
Ji et al. 2009). Several QTLs for LTG were identified as surrounding the
qLTG11-1 locus including
qLTG-11 (
Miura et al. 2001),
qLTG-11 (
Ji et al. 2008), and
qLTG11.2 (
Wang et al. 2011). The
R2 value of
qLTG11-1 was 0.083 (
Hyun et al. 2015) and respective
R2 value of
qLTG-11 (
Miura et al. 2001),
qLTG-11 (
Ji et al. 2008), and
qLTG11.2 was 0.114, 0.127/0.109 (Nanjing/Lingshui) and 0.085. Also QTL
qLTG11 was identified in the same chromosomal location explaining 27.3% of the total phenotypic variance (
Iwata and Fujino 2010), suggesting that this locus was stably expressed for low temperature germinability on chromosome 11. Interestingly, QTL
qSES11 for seedling establishment (SES) was detected with the
R2 value of 0.256 in the same chromosomal region (
Iwata et al. 2010). SES is a useful trait for direct-seeding method in rice, especially under low-temperature conditions (
Iwata et al. 2010).
qLTG4-3 was derived from the QTLs
qLTG-4-1 (
Miura et al. 2001) and
qGR-4 (
Wang et al. 2009). The
R2 values of
qLTG4-3 and
qLTG-4-1 were 0.091 and 0.086, respectively. Three
R2 values were reported for
qGR-4: 0.168 (18 days), 0.117 (20 days) and 0.091 (23 days). Other two markers,
qLTG2-6 and
qLTG4b-1, increased the accuracy in predicting sensitive germplasm in combination with
qLTG3-1 (
Hyun et al. 2015).
Although we have evaluated the phenotypic variation in 180
temperate japonica rice germplasm previously, all accessions classified as tolerant (n=48) could not be used for breeding materials as there was no consideration for genotypic evaluation data. For example, two accessions identified as strong LTG for phenotyping carried the HY allele, associated with weak LTG, for genotyping (
Hyun et al. 2015). Based on the clustering and genotype patterns, a total of 10 germplasm were selected for using in further genetic study and breeding program. Among these accessions, four were early maturity (DPKR 036S 11685, Tokachikuromomi, W-14 and W-15). Five accessions from N. Korea except DPKR 036S 11685 were medium maturity and Hosakan was medium-late maturity. The average germination rate of the 12 Korean direct-seeding varieties developed from 1996 (Dongan) to 2007 (Hwanggeumnodeul) was 63.4% (
Ko et al. 2011), which was much lower than that of accessions selected in our study where it was 94.2%.
Ko et al. (2011) suggested that lower germination rate of the direct-seeding varieties was because develop of the varieties has been focused on enhanced lodging tolerant and high yielding trait and not low temperature germinability. Also, they insisted that selection and breeding for tolerant LTG varieties is needed for elevation of seedling establishment in wet direct-seeding system (
Ko et al. 2011). In terms of seedling establishment, selected accessions demonstrated high germination vigor with average of 2.1 (
Hyun et al. 2015). Besides germination rate, germination vigor is an important trait for breeding direct-seeding varieties. Faster seedling growth can lead to increased crop establishment ratio and enhanced survival ratio in competition with weeds. Therefore, when germplasm are evaluated and screened for the LTG trait, germination vigor should be considered together with germination rate.
In this study, we identified a significant association between the qLTG3-1 alleles and low temperature germinability in temperate japonica rice germplasm. The qLTG3-1, which is the only gene cloned to date for the LTG trait, proved to be a useful marker for screening of LTG in the germplasm examined in this study, which consisted mostly of modern varieties or advanced breeding lines. In addition, genotyping result performed with SNP markers clustered tolerant germplasm into one group based on genetic distance, which resulted in the confirmation of the genotype pattern for increasing the detection of tolerant accessions for direct-use in breeding program. Ten accessions selected in this study through the phenotypic and genotypic evaluation for LTG, therefore, will be useful for breeding of direct-seeding varieties in temperate japonica rice.
ACKNOWLEDGMENTS
This work was supported by the “Research Program for Agricultural Science & Technology Development (Project No. PJ010871)”, Rural Development Administration, Republic of Korea.
Fig. 1Neighbor-joining tree of SNP data using Nei’s distance. Red triangle, tolerant; Blue square, moderate; Green circle, sensitive.
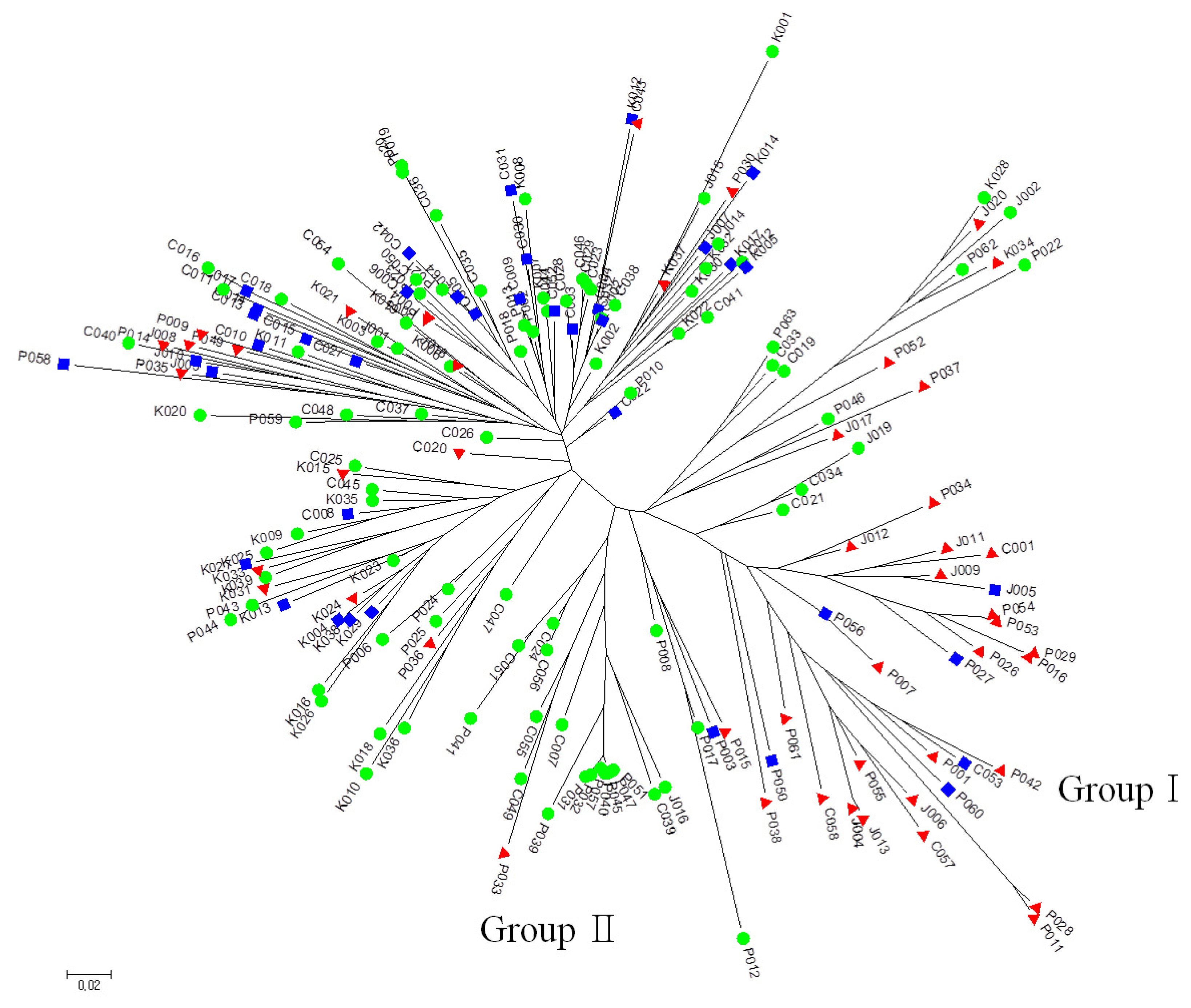
Fig. 2Genotype of five loci from 23 tolerant and 17 sensitive accessions. blue reference allele, red alternative allele, green Hayamasari allele (71bp deletion), blank missing data.
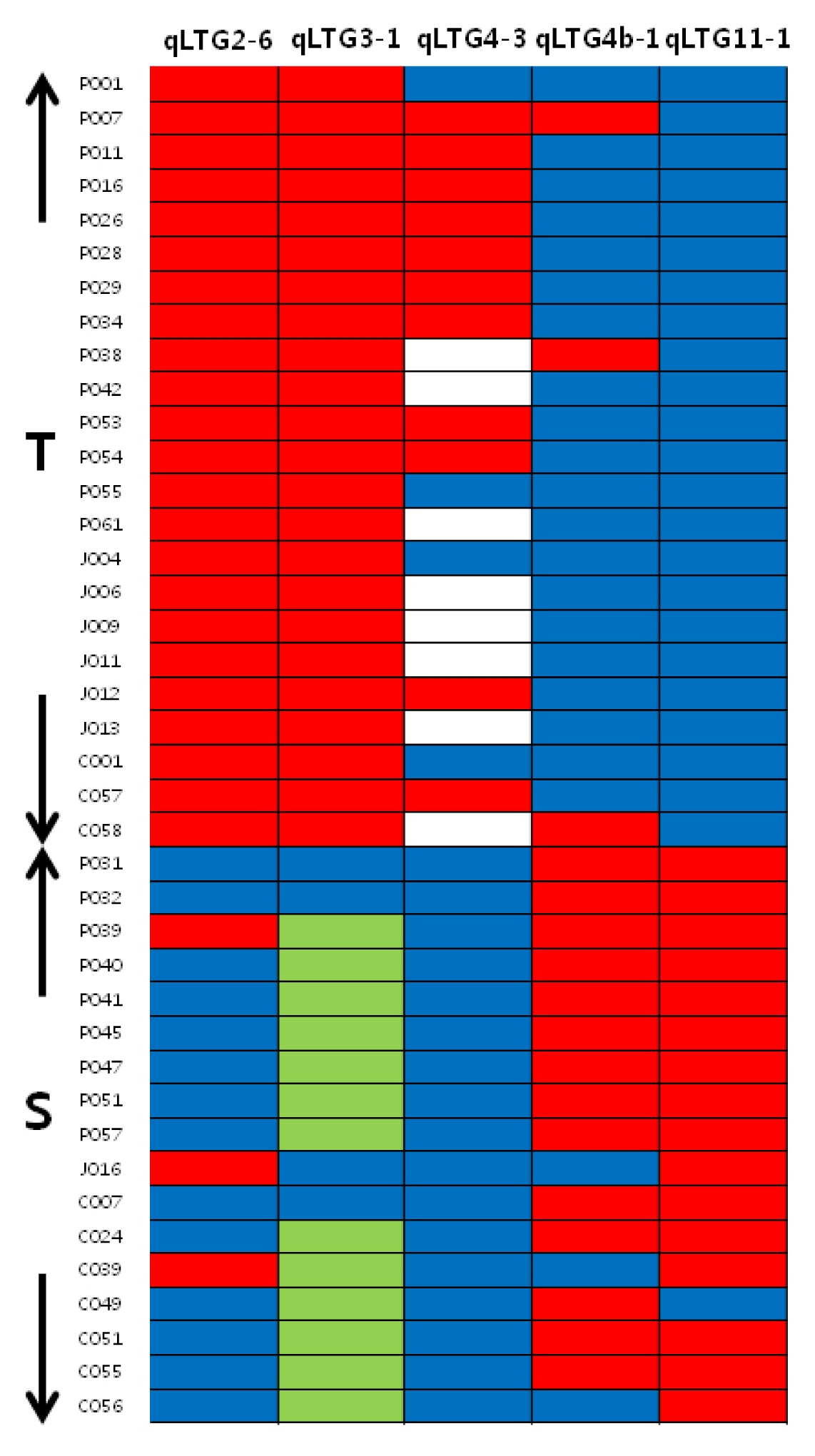
Table 1Germplasm list used in this study.
Table 1
|
No. |
Name |
Origin |
|
1 |
DPKR 036M 11767 |
PRK |
|
2 |
DPKR 036M 1177 |
PRK |
|
3 |
DPKR 036M 1404 |
PRK |
|
4 |
DPKR 036M 2090 |
PRK |
|
5 |
DPKR 036M 2137 |
PRK |
|
6 |
DPKR 036M 3983 |
PRK |
|
7 |
DPKR 036S 432 |
PRK |
|
8 |
DPKR 036S 443 |
PRK |
|
9 |
DPKR 036S 446 |
PRK |
|
10 |
DPKR 036S 454 |
PRK |
|
11 |
DPKR 036S 89 |
PRK |
|
12 |
DPKR 036M 1087 |
PRK |
|
13 |
DPKR 036M 1147 |
PRK |
|
14 |
DPKR 036M 1400 |
PRK |
|
15 |
DPKR 036M 1407 |
PRK |
|
16 |
DPKR 036M 1610 |
PRK |
|
17 |
DPKR 036M 1654 |
PRK |
|
18 |
DPKR 036M 2114 |
PRK |
|
19 |
DPKR 036M 2186 |
PRK |
|
20 |
DPKR 036M 2188 |
PRK |
|
21 |
DPKR 036M 238 |
PRK |
|
22 |
DPKR 036M 299 |
PRK |
|
23 |
DPKR 036M 680 |
PRK |
|
24 |
DPKR 036M 694 |
PRK |
|
25 |
DPKR 036M 971 |
PRK |
|
26 |
DPKR 036S 10288 |
PRK |
|
27 |
DPKR 036S 11511 |
PRK |
|
28 |
DPKR 036S 11727 |
PRK |
|
29 |
DPKR 036S 11685 |
PRK |
|
30 |
DPKR 036S 10296 |
PRK |
|
31 |
DPKR 036S 10309 |
PRK |
|
32 |
DPKR 036S 10312 |
PRK |
|
33 |
DPKR 036S 10322 |
PRK |
|
34 |
DPKR 036S 10332 |
PRK |
|
35 |
DPKR 036S 10336 |
PRK |
|
36 |
DPKR 036S 10338 |
PRK |
|
37 |
DPKR 036S 10370 |
PRK |
|
38 |
DPKR 036S 10454 |
PRK |
|
39 |
DPKR 036S 10467 |
PRK |
|
40 |
DPKR 036S 10469 |
PRK |
|
41 |
DPKR 036S 10487 |
PRK |
|
42 |
DPKR 036S 10491 |
PRK |
|
43 |
DPKR 036S 10500 |
PRK |
|
44 |
DPKR 036S 10503 |
PRK |
|
45 |
DPKR 036S 10512 |
PRK |
|
46 |
DPKR 036S 11487 |
PRK |
|
47 |
DPKR 036S 11510 |
PRK |
|
48 |
DPKR 036S 11539 |
PRK |
|
49 |
DPKR 036S 11556 |
PRK |
|
50 |
DPKR 036S 11666 |
PRK |
|
51 |
DPKR 036S 11687 |
PRK |
|
52 |
DPKR 036S 11712 |
PRK |
|
53 |
DPKR 036S 11713 |
PRK |
|
54 |
DPKR 036S 11721 |
PRK |
|
55 |
DPKR 036S 11744 |
PRK |
|
56 |
DPKR 036S 11750 |
PRK |
|
57 |
DPKR 036S 11752 |
PRK |
|
58 |
DPKR 036S 11765 |
PRK |
|
59 |
DPKR 036S 11781 |
PRK |
|
60 |
DPKR 036S 11782 |
PRK |
|
61 |
DPKR 036S 1221 |
PRK |
|
62 |
DPKR 036S 127 |
PRK |
|
63 |
DPKR 036S 1706 |
PRK |
|
64 |
Dajiaoba |
JPN |
|
65 |
Mogumoguaoba |
JPN |
|
66 |
Hatsumurasaki |
JPN |
|
67 |
Kokushokuto-2 |
JPN |
|
68 |
Kuroke |
JPN |
|
69 |
Kurouruchi |
JPN |
|
70 |
Motsurewase |
JPN |
|
71 |
Norin 20 |
JPN |
|
72 |
Hosakan |
JPN |
|
73 |
Sasaminori |
JPN |
|
74 |
Tokachikuromomi |
JPN |
|
75 |
Norin 9 |
JPN |
|
76 |
Kokushokuto |
JPN |
|
77 |
Nagayama 77402 spontaneous mutant |
JPN |
|
78 |
410750 |
JPN |
|
79 |
thick culm dwarf |
JPN |
|
80 |
A-107 |
JPN |
|
81 |
AC-34 |
JPN |
|
82 |
Daobei 47 |
JPN |
|
83 |
Shang yu 697 |
JPN |
|
84 |
Hao Hai Huan (Acc.66984) |
CHN |
|
85 |
Jiudao 55 |
CHN |
|
86 |
Jiudao 60 |
CHN |
|
87 |
Jiudao 39 |
CHN |
|
88 |
Tongyu 223 |
CHN |
|
89 |
Tongyu 221 |
CHN |
|
90 |
Jijing 504 |
CHN |
|
91 |
Jijing 91 |
CHN |
|
92 |
Jijing 801 |
CHN |
|
93 |
Ji 08-156 |
CHN |
|
94 |
Nan jing 47 |
CHN |
|
95 |
Wu yu jing 23 |
CHN |
|
96 |
Chang jing 08-2 |
CHN |
|
97 |
Chang jing 09-8 |
CHN |
|
98 |
Su jing 8 |
CHN |
|
99 |
Xiushui 12 |
CHN |
|
100 |
Xiushui 114 |
CHN |
|
101 |
Xiushui 128 |
CHN |
|
102 |
Hong mi |
CHN |
|
103 |
Pin 6 |
CHN |
|
104 |
Long hua 05-0501 |
CHN |
|
105 |
Xichang li |
CHN |
|
106 |
Sui 02-6159 |
CHN |
|
107 |
He xuan 03-13 |
CHN |
|
108 |
Ha 99-245 |
CHN |
|
109 |
Long yang 2083 |
CHN |
|
110 |
Song 5119 |
CHN |
|
111 |
Jin lang 1 |
CHN |
|
112 |
Xiu 02-6173 |
CHN |
|
113 |
Mu 2002-1235 |
CHN |
|
114 |
Bei 02-20 |
CHN |
|
115 |
Long dun 301-12 |
CHN |
|
116 |
Long pin 02-1 |
CHN |
|
117 |
Long D99709 |
CHN |
|
118 |
Ji 2005-2 |
CHN |
|
119 |
Ji 2005-8 |
CHN |
|
120 |
Ji 2005-13 |
CHN |
|
121 |
Ji 2006-2348 |
CHN |
|
122 |
Qing 02-11 |
CHN |
|
123 |
Nonglin 20 |
CHN |
|
124 |
Long jiao 92-66 |
CHN |
|
125 |
Song 9023 |
CHN |
|
126 |
Ji hei jing |
CHN |
|
127 |
Ji nong da 3 |
CHN |
|
128 |
Dong nong 91-11 |
CHN |
|
129 |
Tong yu 139 |
CHN |
|
130 |
Sasanisikix2302 |
CHN |
|
131 |
Song 98-10 |
CHN |
|
132 |
Yan jing 105 |
CHN |
|
133 |
Te you 12 |
CHN |
|
134 |
Fuyuan 3 |
CHN |
|
135 |
Te zhan 2 |
CHN |
|
136 |
Eluosi dao |
CHN |
|
137 |
Dun nei dao |
CHN |
|
138 |
Ruan xiang jing |
CHN |
|
139 |
Long dun 97-1 |
CHN |
|
140 |
W-14 |
CHN |
|
141 |
W-15 |
CHN |
|
142 |
IKSAN558 |
KOR |
|
143 |
Heugjinju Byeo |
KOR |
|
144 |
Nokyang |
KOR |
|
145 |
IKSAN541 |
KOR |
|
146 |
Suweon 546 |
KOR |
|
147 |
Hwaseong Byeo |
KOR |
|
148 |
Jungan Byeo |
KOR |
|
149 |
Suweon 541 |
KOR |
|
150 |
Milyang 237 |
KOR |
|
151 |
Ilpum Byeo |
KOR |
|
152 |
Josaengheugchal |
KOR |
|
153 |
Jinbu Byeo |
KOR |
|
154 |
Iksan 527 |
KOR |
|
155 |
Seopyeong |
KOR |
|
156 |
Milyang 234 |
KOR |
|
157 |
Milyang 276 |
KOR |
|
158 |
Sobi Byeo |
KOR |
|
159 |
Heugkwang Byeo |
KOR |
|
160 |
Suweon 540 |
KOR |
|
161 |
Suweon 525 |
KOR |
|
162 |
Hongjinju Byeo |
KOR |
|
163 |
Joun |
KOR |
|
164 |
Cheolweon 84 |
KOR |
|
165 |
Dami |
KOR |
|
166 |
Dongan Byeo |
KOR |
|
167 |
Suwon 563 |
KOR |
|
168 |
Daelip Byeo 1 |
KOR |
|
169 |
Milyang 260 |
KOR |
|
170 |
Dongjin Byeo |
KOR |
|
171 |
Milyang 256 |
KOR |
|
172 |
Suweon 530 |
KOR |
|
173 |
Gopum |
KOR |
|
174 |
Hwaseon Chal Byeo |
KOR |
|
175 |
Chilsung |
KOR |
|
176 |
Suweon 544 |
KOR |
|
177 |
Odae Byeo |
KOR |
|
178 |
Iksan 555 |
KOR |
|
179 |
Suwon 555 |
KOR |
|
180 |
Milyang 282 |
KOR |
Table 2Distribution of
qLTG3-1 allele and its association with phenotype and country of origin. Phenotyping and
qLTG3-1 genotyping were previously described (
Hyun et al. 2015).
Table 2
|
Phenotype |
Origin |
|
|
|
|
Tolerant |
Moderate |
Sensitive |
S. Korea |
China |
Japan |
N. Korea |
Total |
Ave. GRz)
|
|
ILy)
|
35 |
10 |
3 |
4 |
8 |
12 |
24 |
48 |
76.2 |
|
NB |
11 |
16 |
25 |
20 |
16 |
6 |
10 |
52 |
37.3 |
|
HY |
2 |
13 |
65 |
15 |
34 |
2 |
29 |
80 |
17.6 |
|
χ2
|
56.617 |
3.087 |
32.844 |
11.022 |
6.261 |
13.681 |
6.816 |
|
|
|
P |
5.08E-13***
|
0.214 |
7.38E-08***
|
0.004**
|
0.044*
|
0.001**
|
0.033*
|
|
|
Table 3Statistical values for SNP markers.
Table 3
|
Marker |
MAFz)
|
Gene Diversity |
PICy)
|
Marker |
MAF |
Gene Diversity |
PIC |
|
qLTG2-1
|
0.508 |
0.50 |
0.375 |
qLTG7a-3
|
0.681 |
0.435 |
0.340 |
|
qLTG2-2
|
0.750 |
0.375 |
0.305 |
qLTG7b-1
|
0.967 |
0.064 |
0.062 |
|
qLTG2-3
|
0.517 |
0.499 |
0.375 |
qLTG7b-2
|
0.602 |
0.479 |
0.364 |
|
qLTG2-5
|
0.802 |
0.318 |
0.267 |
qLTG7b-3b
|
0.944 |
0.107 |
0.101 |
|
qLTG2-6
|
0.656 |
0.451 |
0.349 |
qLTG7b-5c
|
0.684 |
0.433 |
0.339 |
|
qLTG3-1
|
0.525 |
0.499 |
0.374 |
qLTG7b-7
|
0.936 |
0.120 |
0.112 |
|
qLTG4-1
|
0.831 |
0.280 |
0.241 |
qLTG7b-9
|
0.892 |
0.193 |
0.175 |
|
qLTG4-3
|
0.859 |
0.242 |
0.213 |
qLTG7c-1
|
0.874 |
0.220 |
0.196 |
|
qLTG4-4
|
0.612 |
0.475 |
0.362 |
qLTG9-1
|
0.944 |
0.105 |
0.100 |
|
qLTG4-5
|
0.956 |
0.085 |
0.081 |
qLTG9-2
|
0.843 |
0.265 |
0.230 |
|
qLTG4-6
|
0.578 |
0.488 |
0.369 |
qLTG9-4
|
0.619 |
0.472 |
0.360 |
|
qLTG4-7
|
0.950 |
0.095 |
0.090 |
qLTG9-6
|
0.966 |
0.065 |
0.063 |
|
qLTG4-9
|
0.876 |
0.217 |
0.193 |
qLTG11-1
|
0.693 |
0.426 |
0.335 |
|
qLTG4b-1
|
0.696 |
0.424 |
0.334 |
qLTG11-2
|
0.750 |
0.375 |
0.305 |
|
qLTG7a-1
|
0.964 |
0.070 |
0.067 |
qLTG11-3
|
0.761 |
0.364 |
0.298 |
|
qLTG7a-2
|
0.856 |
0.247 |
0.217 |
qLTG11-4
|
0.654 |
0.453 |
0.350 |
|
|
|
|
Mean |
0.773 |
0.307 |
0.248 |
Table 4Germination rate and agronomic traits for selected strong LTG accessions.
Table 4
|
ID |
Accession Name |
IT No. |
Originz)
|
GRy) (%) |
DTHx) (d) |
PHw) (cm) |
PLv) (cm) |
1,000-GWu) (g) |
|
P007 |
DPKR 036S 432 |
276730 |
PRK |
85 |
61 |
72 |
23 |
37.5 |
|
P016 |
DPKR 036M 1610 |
276898 |
PRK |
86.7 |
65 |
91 |
21 |
26.6 |
|
P026 |
DPKR 036S 10288 |
276967 |
PRK |
93.3 |
56 |
89 |
20 |
29.5 |
|
P029 |
DPKR 036S 11685 |
277036 |
PRK |
91.7 |
54 |
80 |
18 |
26.1 |
|
P054 |
DPKR 036S 11713 |
277271 |
PRK |
96.7 |
57 |
85 |
20 |
41.1 |
|
P055 |
DPKR 036S 11721 |
277274 |
PRK |
95 |
57 |
97 |
24 |
31.6 |
|
J009 |
Hosakan |
275325 |
JPN |
95 |
74 |
55 |
20 |
28.9 |
|
J011 |
Tokachikuromomi |
275327 |
JPN |
98.3 |
59 |
40 |
15 |
25.2 |
|
C057 |
W-14 |
277478 |
CHN |
100 |
56 |
70 |
15 |
25.6 |
|
C058 |
W-15 |
277479 |
CHN |
100 |
59 |
70 |
15 |
25.1 |
References
- Agrama HA, Eizenga GC. 2008. Molecular diversity and genome-wide linkage disequilibrium patterns in a worldwide collection of Oryza sativa and its wild relatives. Euphytica. 160: 339-355.
- Anderson JA, Churchill GA, Sutrique JE, Tanksley SD, Sorrels ME. 1993. Optimizing parental selection for genetic linkage maps. Genome. 36: 181-186.
- Bostein D, White RL, Skolnick M, Davis RW. 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet. 32: 314-331.
- Challam C, Kharshing GA, Yumnam JS, Rai M, Tyagi W. 2013. Association of qLTG3–1 with germination stage cold tolerance in diverse rice germplasm from the Indian subcontinent. Plant Genet Resour-C. 11: 206-211.
- Duran C, Appleby N, Clark T, Wood D, Imelfort M, Batley J, Edwards D. 2009. AutoSNPdb: an annotated single nucleotide polymorphism database for crop plants. Nucl Acids Res. 37: D951-D953.
- Edwards D, Batley J. 2010. Plant genome sequencing: applications for crop improvement. Plant Biotech J. 8: 2-9.
- Fujino K, Sekiguchi H, Sato T, Kiuchi H, Nonoue Y, Takeuchi Y, Ando T, Lin SY, Yano M. 2004. Mapping of quantitative trait loci controlling low-temperature germinability in rice (Oryza sativa L). Theor Appl Genet. 108: 794-799.
- Fujino K, Sekiguchi H, Matsuda Y, Sugimoto K, Ono K, Yano M. 2008. Molecular identification of a major quantitative trait locus, qLTG3-1, controlling low-temperature germinability in rice. Proc Natl Acad Sci USA. 105: 12623-12628.
- Fujino K, Iwata N. 2011. Selection for low-temperature germinability on the short arm of chromosome 3 in rice cultivars adapted to Hokkaido, Japan. Theor Appl Genet. 123: 1089-1097.
- Fujino K, Sekiguchi H. 2011. Origins of functional nucleotide polymorphisms in a major quantitative trait locus, qLTG3-1, controlling low-temperature germinability in rice. Plant Mol Biol. 75: 1-10.
- Gonzaga ZJ, Aslam K, Septiningsih EM, Collard BCY. 2015. Evaluation of SSR and SNP markers for molecular breeding in rice. Plant Breed Biotech. 3: 139-152.
- Han LZ, Zhang YY, Qiao YL, Cao GL, Zhang SY, Kim JH, Koh HJ. 2006. Genetic and QTL analysis for low-temperature vigor of germination in rice. Acta Genet Sin. 33: 998-1006.
- Hyun DY, Lee GA, Kang MJ, Burkart-Waco D, Kim SI, Kim JY, Lee MC, Gwag JG, Kim YG, Tai TH. 2015. Development of low-temperature germinability markers for evaluation of rice (Oryza sativa L.) germplasm. Mol Breed. 35: 104
- Iwata N, Fujino K. 2010. Genetic effects of major QTLs controlling low-temperature germinability in different genetic backgrounds in rice (Oryza sativa L). Genome. 53: 763-768.
- Iwata N, Shinada H, Kiuch H, Sato T, Fujino K. 2010. Mapping of QTLs controlling seedling establishment using a direct seeding method in rice. Breed Sci. 60: 353-360.
- Ji SL, Jiang L, Wang YH, Liu SJ, Liu X, Zhai HQ, Atsushi Y, Wan JM. 2008. QTL and epistasis for low temperature germinability in rice. Acta Agron Sin. 34: 551-556.
- Ji SL, Jiang L, Wang YH, Zhang WW, Liu X, Liu SJ, Chen LM, Zhai HQ, Wan JM. 2009. Quantitative trait loci mapping and stability for low temperature germination ability of rice. Plant Breeding. 128: 387-392.
- Jiang L, Liu SJ, Hou MY, Tang JY, Chen LM, Zhai HQ, Wan JM. 2006. Analysis of QTLs for seed low temperature germinability and anoxia germinablity in rice (Oryza sativa L). Field Crops Res. 98: 68-75.
- Jin L, Lu Y, Xiao P, Sun M, Corke H, Bao JS. 2010. Genetic diversity and population structure of a diverse set of rice germplasm for association mapping. Theor Appl Genet. 121: 475-487.
- KMA. 2015. http://www.kma.go.kr/weather/climate/average_30years. Korea Meteorological Administration
- Ko JC, Baek MK, Kim WJ, Shon JY, Ha KY, Kang HJ, Shin MS, Ko JK. 2011. Analysis of low temperature germination ratio in rice cultivars for breeding of direct seeding variety. Korean J Intl Agri. 23: 89-94.
- Li LF, Liu X, Xie K, Wang YH, Liu F, Lin QY, Wang WY, Yang CY, Lu BY, Liu SJ, Chen LM, Jiang L, Wan JM. 2013. qLTG-9, a stable quantitative trait locus for low-temperature germination in rice (Oryza sativa L). Theor Appl Genet. 126: 2313-2322.
- Liu K, Muse SV. 2005. PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics. 21: 2128-2129.
- Miura K, Lin SY, Yano M, Nagamine T. 2001. Mapping quantitative trait loci controlling low temperature germinability in rice (Oryza sativa L). Breed Sci. 51: 293-299.
- Nei M, Takezaki N. 1983. Estimation of genetic distances and phylogenetic trees from DNA anlysis. Proc 5th World Cong Genet Appl Livstock Prod. 21: 405-412.
- Rafalski A. 2002. Applications of single nucleotide polymorphisms in crop genetics. Curr Opin Plant Biol. 5: 94-100.
- RDA.2012. Standard protocol of research method and analysis. Rural Development Administration. Suwon: pp. 316-334.
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 30: 2725-2729.
- Teng S, Zeng DL, Qian Q, Kunihifo Y, Huang DN, Zhu LH. 2001. QTL analysis of rice low temperature germinability. Chin Sci Bull. 46: 1800-1803.
- Wang ZF, Wang JF, Wang FH, Bao YM, Wu YY, Zhang HS. 2009. Genetic control of germination ability under cold stress in rice. Rice Sci. 16: 173-180.
- Wang ZF, Wang FH, Zhou R, Wang JF, Zhang HS. 2011. Identification of quantitative trait loci for cold tolerance during the germination and seedling stages in rice (Oryza sativa L). Euphytica. 181: 405-413.
- Xu Y, Beachell H, McCouch SR. 2005. A marker-based approach to broadening the genetic base of rice in the USA. Crop Sci. 44: 1947-1959.