Abstract
This study was initiated to identify quantitative trait loci (QTL) involved in root system development by measuring root length and weight of rice seedlings grown in hydroponic culture condition. Growth conditions for estimating the root elongation were set up to supply NH4+ as a single nitrogen source. Four rice accessions and 40 introgression lines (ILs) derived from a cross between ‘Ilpumbyeo’, a temperate japonica variety, and ‘Moroberekan’, a tropical japonica accession, were used to detect QTL for two root traits of seedlings grown in 0, 250 and 500 μM NH4+. Seven chromosomal regions were found to be involved in root elongation. The qRW6 QTL detected on the long-arm of chromosome 6 promoted root weight and the ‘Moroberekan’ allele at this QTL was associated with enhanced root weight at 250 μM NH4+ concentration. The Moroberekan alleles were beneficial at all loci except for qRW4 at which the Ilpumbyeo allele increased root weight under 0, 250 and 500 μM NH4+. A field trial was conducted for selected four ILs to know whether the introgressed Moroberekan segments harboring the beneficial QTL were associated with effects on other agronomic traits. No significant difference was detected between four ILs and Ilpumbyeo for days to heading and culm length. Two lines showed significant difference from Ilpumbyeo in spikelets per panicle and 1,000 grain weight, and the difference might be attributed to the Moroberekan segment on chromosome 6. Our QTL results indicate that the Moroberekan introgression on chromosome 6 flanked by RM162-RM3430 has the potential to enhance root traits and yield potential in the temperate japonica background.
-
Key words: Rice, Introgression lines, NH4+ response, Root elongation, QTL
INTRODUCTION
Rice is one of the most important crops for almost 50% of the world’s population. Dramatic increase in the production and use of fertilizer N in rice-growing countries has occurred during the past decades (
Craswell and Vlek 1983). The usual cultivation of rice includes major changes in the N supply form as the seedling plants are initially raised in a nursery bed and subsequently transferred to the flooded paddy culture system (
Patil et al. 2013). Even though N fertilizer increases plant yield, plants consume much less than half of the fertilizers applied (
Frink et al. 1999;
Socolow 1999) while a majority of N fertilizers were lost to the atmosphere or leached into groundwater, lakes and rivers, which causes severe adverse effects to the environment.
Nitrogen (N) status strongly affects the photosynthetic rate in crop. Rice evolved with greater capacity to assimilate ammonium ion (NH
4+) compared with other plant species (
Yamaya and Oaks 2004). Studies have shown NH
4+ can improve the capacity to tolerate water stress in rice in comparison with NO
3− (
Guo et al. 2007). However, NH
4+ frequently reaches levels in soils that affect plant growth negatively. These negative effects manifest in stunted root growth, yield depression, and chlorosis of leaves (
Britto and Kronzucker 2002;
Balkos et al. 2010). High NH
4+ mainly inhibited the development of total root length and root area while the formation of lateral roots was not affected (
Zhao et al. 2014). Nitrogen deficiency delayed both vegetative and reproductive phenological development, slightly reduced leaf emergence rate, and strongly diminished leaf expands. Therefore, it is important to find ways to enhance nitrogen use efficiency (NUE) in rice.
Root morphological and physiological traits are controlled by QTL. Root system architecture (RSA) is generally defined by length, weight, number, thickness and density of primary, lateral and adventitious roots (
Obara et al. 2010) but RSA study is difficult to handle in soil condition. To overcome these limitations, many of the root studies grow rice seedlings in hydroponic system using diverse mapping populations (
Obara et al. 2010;
Ogawa et al. 2014). The results suggest that QTLs for root length uncovered under hydroponics may be predictive of enhanced grain yield under field conditions (
Obara et al. 2011). This approach has been widely used to detect QTLs associated with improved root systems in both stressed and non-stressed rice fields (
Price et al. 1997;
Shimizu et al. 2004). However, QTL studies for root traits using mapping populations from crosses between
japonica accessions are limited.
In this study, we established growth condition to reliably evaluate rice seedling traits grown in hydroponic condition to identify chromosomal regions associated with root traits under different NH4+ levels using introgression lines (ILs) derived from a cross between two japonica rice accessions ‘Ilpumbyeo’ and ‘Moroberekan’.
MATERIALS AND METHODS
Plant materials
Four rice accessions ‘Ilpumbyeo’, ‘Moroberekan’, ‘Hwaseongbyeo’ and ‘
O. rufipogon’ (IRGC 105491) were used for parental screening. In the previous research, 117 BC
3F
5 lines were developed from a cross between Ilpumbyeo and Moroberekan (
Kim et al. 2009). From the 117 BC
3F
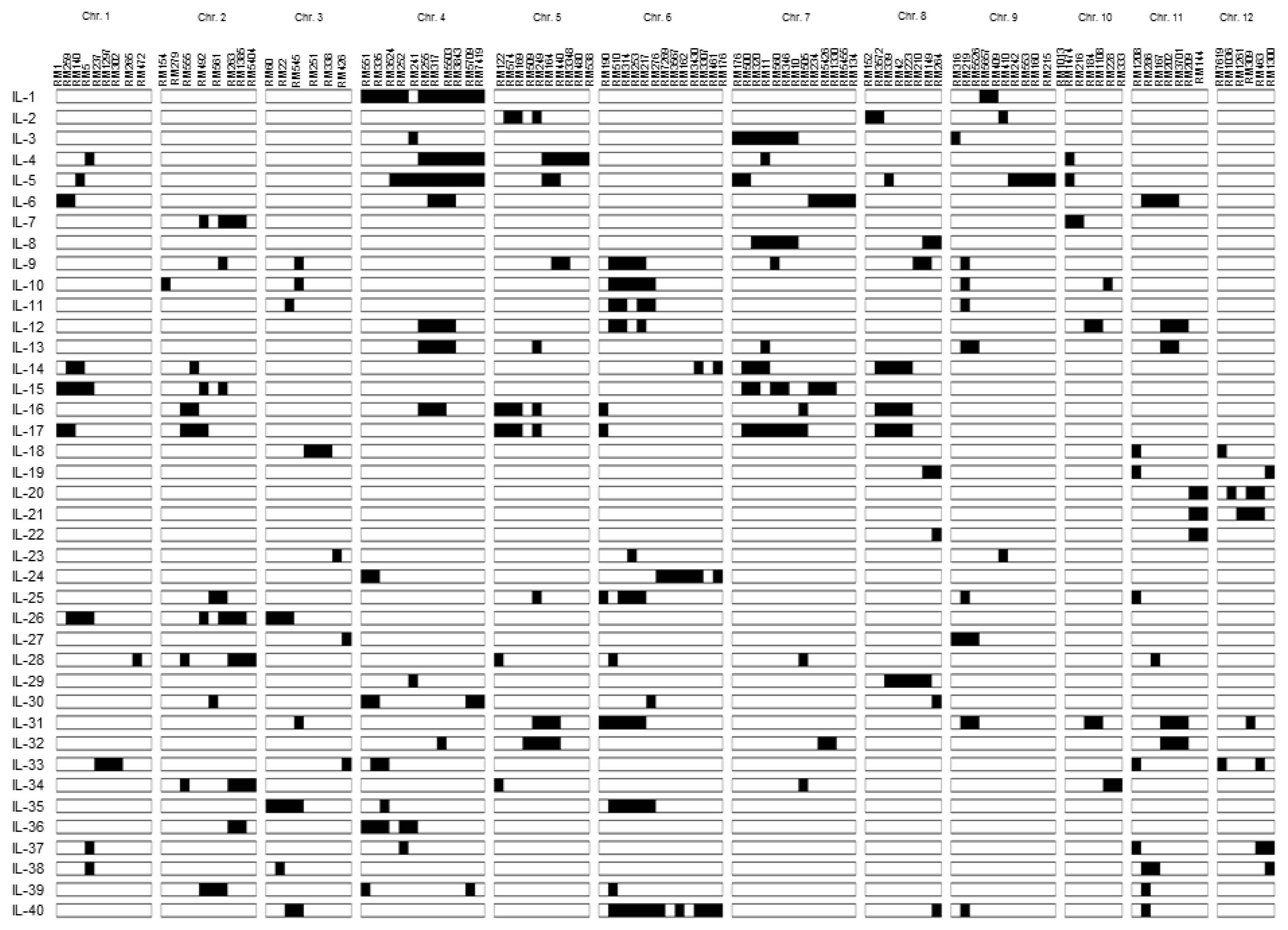
5, 40 ILs (introgression lines) covering the whole Moroberekan chromosome in the genetic background of Ilpumbyeo were selected and used in this study (
Fig. 1,
Kim et al. 2009).
A field trial was conducted to compare selected four ILs with Ilpumbyeo for agronomic traits. Plots for all genotypes were laid out in a completely randomized block design with two replications, one row per line, 25 plants per each row in 2013. Four phenotypic traits, days to heading (DTH), culm length (CL), 1,000 grain weight in brown rice (GW) and spikelets per panicle (SPP) were measured. The middle 3 plants from each line were chosen for the evaluation and average number of the measurements was used for the phenotype of each line for the four traits: DTH, CN, GW, and SPP. Evaluation of the traits followed the method as described in
Kim et al. (2014)
All seeds were pre-selected with NaCl solution (d= 1.113) and processed by three-step of sterilization (
Obara et al. 2010). First, the seeds were soaked for 10 min in distilled water at 60°C and then washed for 20 min with running tap water. Second, the seeds were soaked for 30 sec in a 70% (v/v) ethanol solution and they were washed for 2 min with distilled water. Finally, the seeds were soaked for 20 min in a 1% (v/v) sodium hypochlorite solution and then washed four times with distilled water.
Seeds were kept for germination at 30°C, in dark conditions for 2 days in an incubator. After the tip of the plumule had barely emerged, 50 seeds were placed between moist paper towels and then gently rolled. Rolled papers were then transferred to plastic boxes (456 × 296 × 112 mm). Rolled seedlings were grown in a chamber with a 14-hr light at 30°C/10h dark at 28°C, 70% relative humidity. 7 days after imbibition, uniform seedlings established in moist paper towels were selected and moved in a water culture tube with a sponge. The seedlings were grown in 0, 250, and 500 μM NH
4Cl dissolved in 10L half strength of Yoshida solution (
Yoshida et al. 1976). The seedlings were grown in a chamber with a 14-h light at 28°C/10-h dark at 25°C, 70% relative humidity. The experiments were conducted in a completely randomized design with two replications for each treatment. When NH
4Cl was applied to rice plants as a single source of nitrogen, the pH of the nutrient solutions shifted to the acidic region, around pH 3.0–4.0. To maintain pH, the nutrient solution was replaced every three days to maintain pH 5.5~5.6. 10 days after being treated. Seedling root lengths were measured with a ruler, and root fresh weight (RW) was measured with a scale.
The differences among mean values for seedling root traits (4 accessions) were statistically analyzed by one-way ANOVA (Tukey’s test with Minitab 6.0) (
Ryan et al. 1992). Previously, 134 SSR markers were mapped for the 117 BC
3F
5 lines and these data were used for detecting QTLs in 40 ILs (
Kim et al. 2009). Single point analysis (SPA) was performed to determine the effect of each marker on each trait using Q-gene program (
Nelson 1997). In SPA, QTL was declared if the phenotype was associated with a marker locus at
P<0.005 or with two adjacent marker loci at
P<0.01.
RESULTS
Variation of root traits in response to NH4+ supply among four rice accessions
Root length and weight of ‘Ilpumbyeo’, ‘Hwaseongbyeo’, ‘Moroberekan’ and ‘
O. rufipogon’ plants grown in hydroponic culture with three NH
4+ concentrations was measured as an indicator of genetic variation (
Table 1). Four rice accessions were grown for 11 days. Moroberekan and
O. rufipogon showed significantly higher values in root length and weight than Ilpumbyeo and Hwaseongbyeo (
P<0.05) at all NH
4+ levels. At 250 μM Hwaseong showed the shorter roots than Ilpumbyeo. Moroberekan and
O. rufipogon were not significantly different in root length and weight at all NH
4+ levels. Ilpumbyeo displayed better growth for roor length and weight at 250 μM NH
4+ than the other two NH
4+ levels. Hwaseongbyeo performed poor showing shortest roots of 14.3 cm at 250 μM NH
4+ whereas no difference was observed for root weight between 250 and 500 μM levels.
Seedlings of 40 ILs and their parental lines were grown in hydroponic culture conditions, supplemented with 0, 250, and 500 μM NH
4+. Frequency distributions of phenotypes of RL and RW of the 40 ILs are shown in
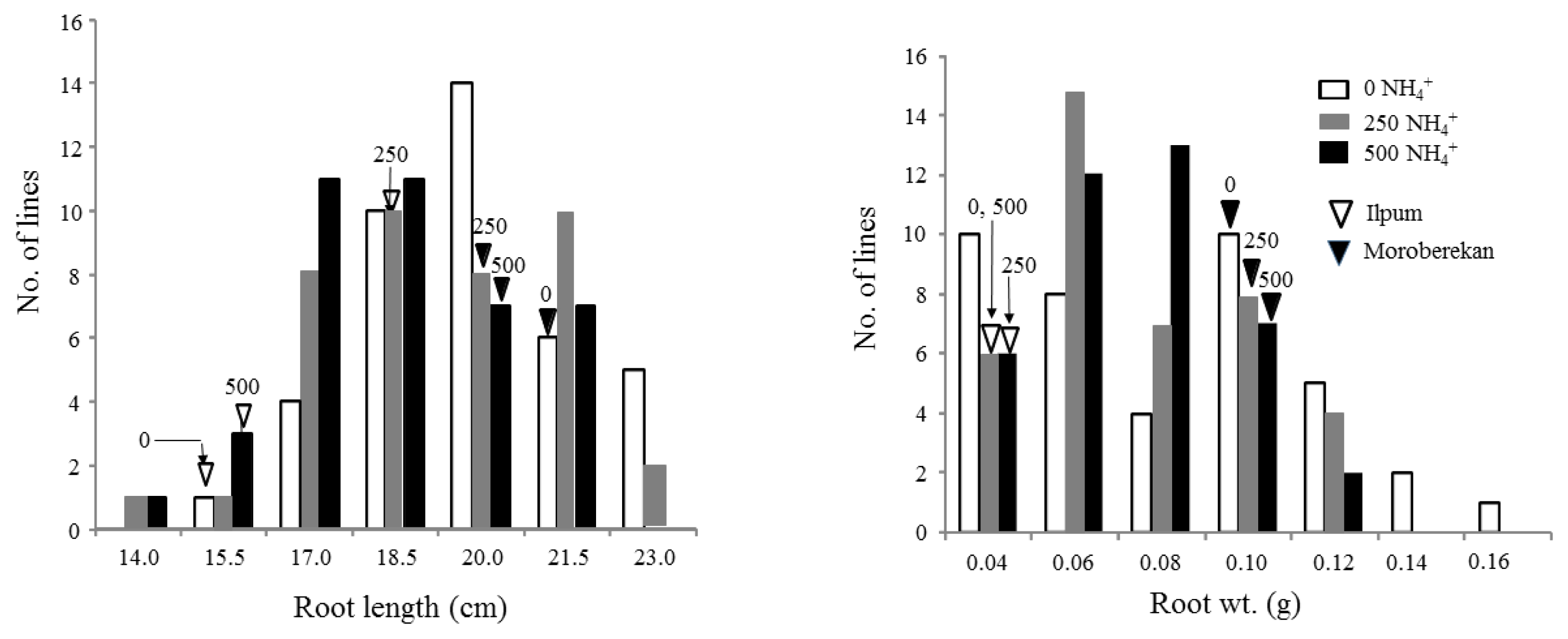
Fig. 2. RL value showed bimodal distributions at three NH
4+ levels and ranged from 14.0 to 23.2 cm at 250 μM NH
4+. RW also showed bimodal distributions. Transgressive lines with phenotypic values larger than Moroberekan were observed for RL and RW.
The root length of Ilpumbyeo and Moroberekan were 15.5 ± 1.8 and 20.4 ± 2.7 cm under the 0 μM NH4+ level, 18.8 ± 2.1 and 19.9 ± 2.8 cm under the 250 μM NH4+ level and 15.7 ± 2.0 and 19.5 ± 2.4 cm under the 500 μM NH4+ level, indicating that genetic variation of root length in response to exogenous NH4+ concentration exists between the parental lines in hydroponic condition.
The root weight of Ilpumbyeo and Moroberekan were 0.05 ± 0.02 and 0.09 ± 0.03 g under the 0 μM NH4+ level, 0.06 ± 0.02 and 0.10 ± 0.03 g under the 250 μM NH4+ level and 0.05 ± 0.01 and 0.10 ± 0.03 g under the 500 μM NH4+ level, indicating genetic variation of root weight.
QTL analysis
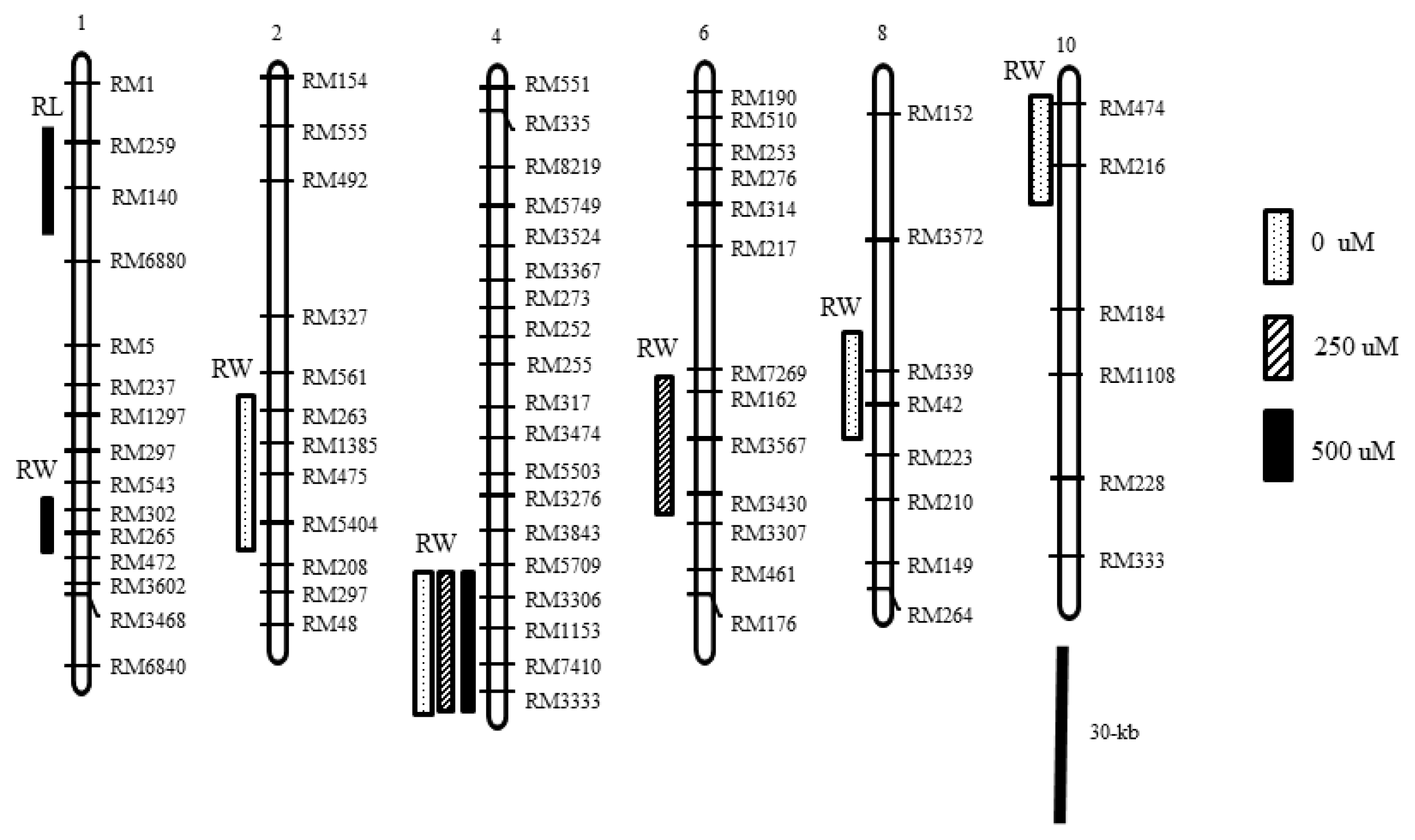
Seven QTLs distributed on six chromosomes (1, 2, 4, 6, 8, and 10) were detected in this study (
Table 2, and
Fig. 3). One QTL,
qRW4 was detected under 0, 250 and 500 NH
4+ levels among seven QTLs. The other QTLs include three QTLs (qRW2, qRW8, and qRL10)under 0 μM NH
4+, one QTL (qRW6)under 250 μM NH
4+ level, 2 QTLs (qRL1 and qRW1) detected under 500 μM NH
4+ level for RL and RW.
Two QTL of root length (RL) were detected on chromosomes 1 and 10. One QTL, qRL1 was detected at the marker interval of RM259-RM140 under 500 μM NH4+ level, and the other QTL, qRL10 was detected near the marker interval of RM474-RM216 under 0 μM NH4+ level. The phenotypic variance and LOD peak of two QTL were 31.4% and 3.3 and 17.7% and 1.7, respectively. Moroberekan alleles increased root length at two loci.
Five QTL of root weight (RW) were detected on chromosomes 1, 2, 4, 6 and 8. qRW1 on chromosome 1 was detected near the marker RM302-RM265 under 500 μM NH4+ level. qRW2 on chromosome 2 was detected near the marker interval RM263-RM5404 under the 0 μM NH4+ level. The QTL on chromosome 4, qRW4 was detected at the marker interval of RM7410-RM3333 at three NH4+ levels and at this locus the Ilpumbyeo allele increased root weight. The qRW4 QTL explained 26.6–27.7% of the phenotype variance for root weight (data not shown). qRW6 on chromosome 6 was detected near the marker RM7269-RM3430 under 250 μM NH4+ level. qRW8 on chromosome 8 was detected under the 0 μM NH4+ level.
Agronomic traits of selected ILs
To determine whether the introgressed segments harboring the beneficial QTL were associated with effects on other yield component and grain traits, a field trial was conducted using seeds from selected four ILs, and the parent, Ilpumbyeo (
Table 3). These four lines performed better than Ilpumbyeo in root weight and length in varying NH
4+ levels and were phenotypically similar to Ilpumbyeo. No significant difference was detected between four ILs and Ilpumbyeo for days to heading and culm length. Two lines, IL14 and IL24 showed significant difference from Ilpumbyeo for spikelets per panicle and 1,000 grain weight, and the difference might be attributed to the Moroberekan segment on chromosome 6 (
Kim et al. 2014).
DISCUSSION
Nitrogen (N) is essential for plant growth and development (
Su et al. 2005). Even though N fertilizer increases plant yield, nitrogen use efficiency of plants is generally low (
Frink et al. 1999). Rice plants mainly depend on NH
4+ that is the major form of nitrogen in paddy fields (
Yamaya et al. 2004). Excessive NH
4+ inhibits the growth of most crop species (
Roosta et al. 2008). Several studies have been conducted to identify QTL for root traits associated with different NH
4+ concentrations using various mapping populations (
Obara et al. 2010;
Obara et al. 2011; Chun
et al. 2014;
Ogawa et al. 2014). However, report on root traits using ILs derived from a cross between
japonica accessions is lacking.
We evaluated four rice accessions under different NH
4+, and Moroberekan and
O. rufipogon consistently displayed enhanced RL and RW compared with the other two temperate
japonica genotypes, Ilpumbyeo and Hwaseongbyeo. Two temperate
japonica accessions at 250 μM experienced the maximum growth compared to 0 and 500 μM NH
4+ levels. Moroberekan and
O. rufipogon were not significantly different for two root traits in various NH
4+ concentrations. Therefore, Moroberekan and
O. rufipogon were classified as NH
4+ less-sensitive.
Ogawa et al. (2013) also reported that
O. rufipogon constitutively elongated seminal roots under 5, 50, and 500 μM NH
4+ concentrations being not NH
4+ sensitive. These results also confirm the previous results that Moroberekan and
O. rufipogon possess beneficial alleles for root architectural traits in breeding programs to enhance
O. sativa. Moroberekan has been previously used as a donor of many useful agronomical traits, such as resistance to drought or blast disease (
Lilley et al. 1996;
Ju et al. 2004;
Kang et al. 2011).
O. rufipogon has also been utilized as a source of biotic and abiotic stress tolerance (
Brar and Khush 1997) and as a source of yield-enhancing alleles for
O. sativa (
Imai et al. 2013). Because
O. rufipogon can be easily crossed with
O. sativa, this progenitor species represents a valuable source of useful traits for rice improvement, including root architectural traits.
To identify and map QTL associated with root traits, we evaluated 40 ILs from a cross between Ilpumbyeo and Moroberekan and detected a total of 7 QTLs. When the chromosomal locations of the QTL detected in this study were compared with those of the previously detected QTL (
www.gramene.org), several overlapping loci were observed.
qRW1 near the marker RM302-RM265 on chromosome 1 was colocated with a QTL for seminal root elongation by
Obara et al. (2010 and
2011), which was detected in rice seedlings grown in hydrophonic condition using two different mapping populations derived from Koshihikari × Kasalath and Taichung 65 ×
O. glaberrima.
qRW1 was also colocalized with a major QTL for seminal root elongation under 500 μM NH
4+ using a CSSL population from a cross between ‘Curinga’ and
O. rufipogon (
Ogawa et al. 2014) and a QTL for shoot dry weight, and plant dry weight (
Wei et al. 2012). In our study, a
qRW1 introgression from Moroberekan in the Ilpumbyeo genetic background was associated with higher root weight at 500 μM NH
4+ level. The
qRW4 QTL on chromosome 4 shared the similar chromosomal regions related to root elongation (
Obara et al. 2010) and to nitrogen-use and nitrogen-deficiency tolerance (
Wei et al. 2012).
qRW6 detected in 250 μM NH
4+ level in our study shared the similar chromosomal region with a QTL,
qRL6.1 for seminal root elongation in a wide range of NH
4+ concentrations (
Obara et al. 2010).
qRL6.1 was narrowed down to a 336 kb-long region in the genome sequence and the ‘Kasalath’ allele on
qRL6.1 was associated with enhanced root elongation in the ‘Nipponbare’ genetic background. This region also harbors a QTL for maximum root length of seedlings grown in soil culture and a QTL for maximum root length of plants grown in low nitrogen conditions (
MacMillan et al. 2006). The QTL,
qRW8 shared the similar location with a QTL for nitrogen response (
Cho et al. 2007;
Obara et al. 2010;
Zhao et al. 2014).
qRL10 on chromosome 10 under 0 μM NH
4+ level detected in RM474-RM216 was colocated with QTL for relative biomass and nitrogen response (Lian
et al. 2005;
Wei et al. 2012). The QTL,
qRW8 shared the similar location with a QTL for nitrogen response (
Cho et al. 2007;
Obara et al. 2010;
Zhao et al. 2014).
However, it is very difficult to conclude whether these QTL are allelic or not because these QTL were mapped by QTL analysis, not by fine-mapping as was conducted for
qRL6.1 by
Obara et al. (2010). Additional fine-mapping will be required to conclude an allelic relationship among these QTL loci. It is noteworthy that the Moroberekan introgression near RM3430-RM20580 harbors a QTL cluster associated with grain weight and spikelets per panicle in the Ilpumbyeo genetic background (
Kim et al. 2014). Our QTL results indicate that the Moroberekan introgression on chromosome 6 flanked by RM162-RM3430 has the potential to enhance root traits and yield potential in the temperate
japonica background. All the Moroberekan alleles were associated with an increase in root length and weight except for
qRW4. Although Moroberekan had higher root weight than Ilpumbyeo, the Ilpumbyeo allele at
qRW4 was favorable in terms of root weight. This result is similar with the study of
Obara et al. (2010) in which a QTL for root length was detected on chromosome 4 using CSSLs and the Kasalath allele at this QTL locus inhibited root elongation. Frequency distribution indicated the existence of several lines showing transgressive segregation for RL and RW at 3 NH
4+ levels and this might be explained by this QTL due to a combination of favorable alleles from both parents. Further analyses for validating effect of
qRW4 and
qRW6 on root growth are underway.
In conclusion, this work is only a starting point for characterizing the genetic basis of rice growth in different NH4+ levels. A few lines with beneficial QTLs for root length and weight from Moroberekan in this study would be useful in developing lines with root system development in the breeding program.
ACKNOWLEDGEMENTS
This work was supported by the Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ906910 to S.N.A), Rural Development Administration, Republic of Korea.
Fig. 1Graphical genotypes of 40 introgression lines used in this study. Black boxes indicat Moroberekan introgressions in the Ilpumbyeo background. Markers are shown on each chromosome.
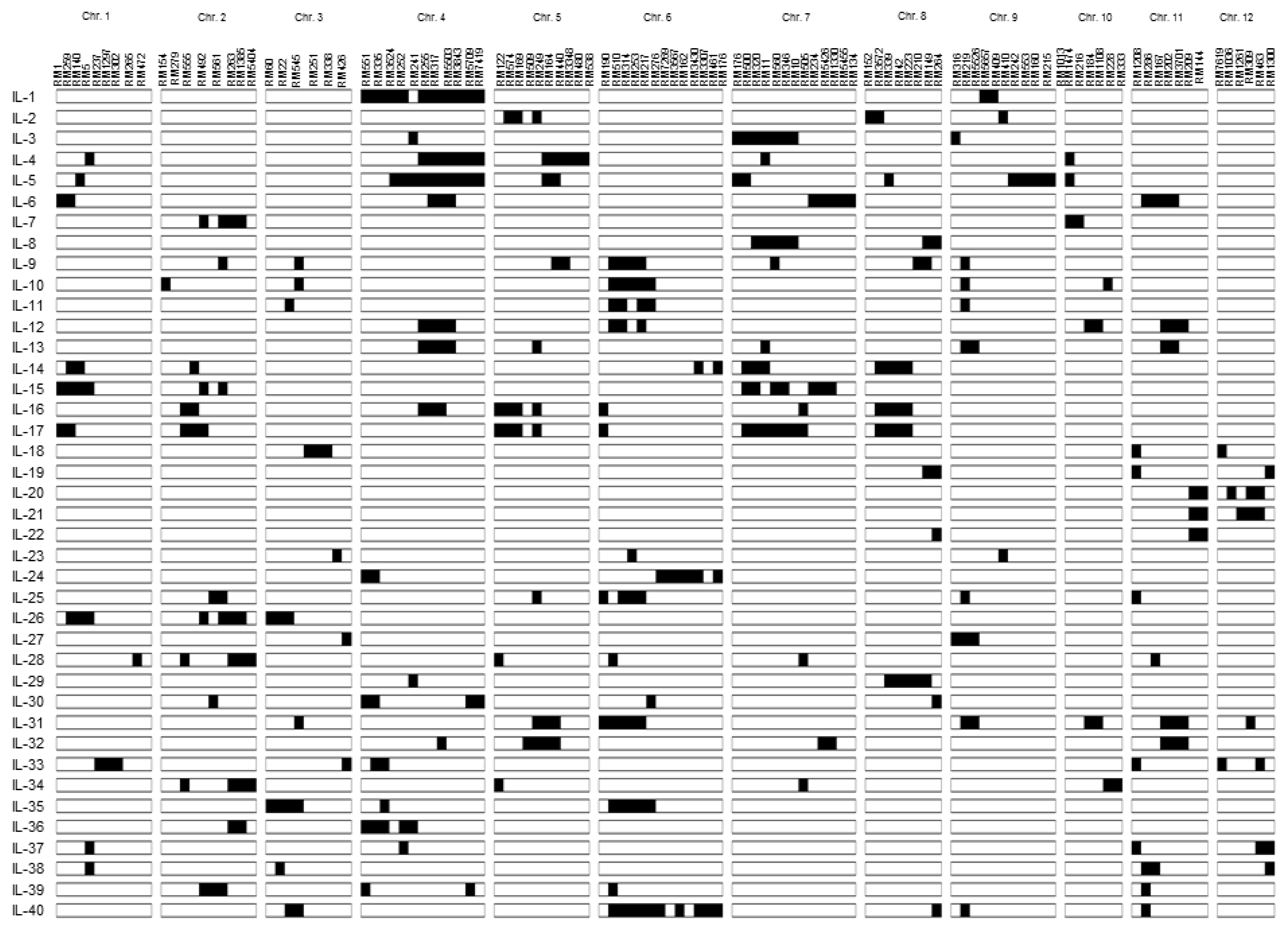
Fig. 2Distribution of root length and weight in 40 ILs with the parental lines grown in hydroponic condition under 0, 250, and 500 μM NH4+.
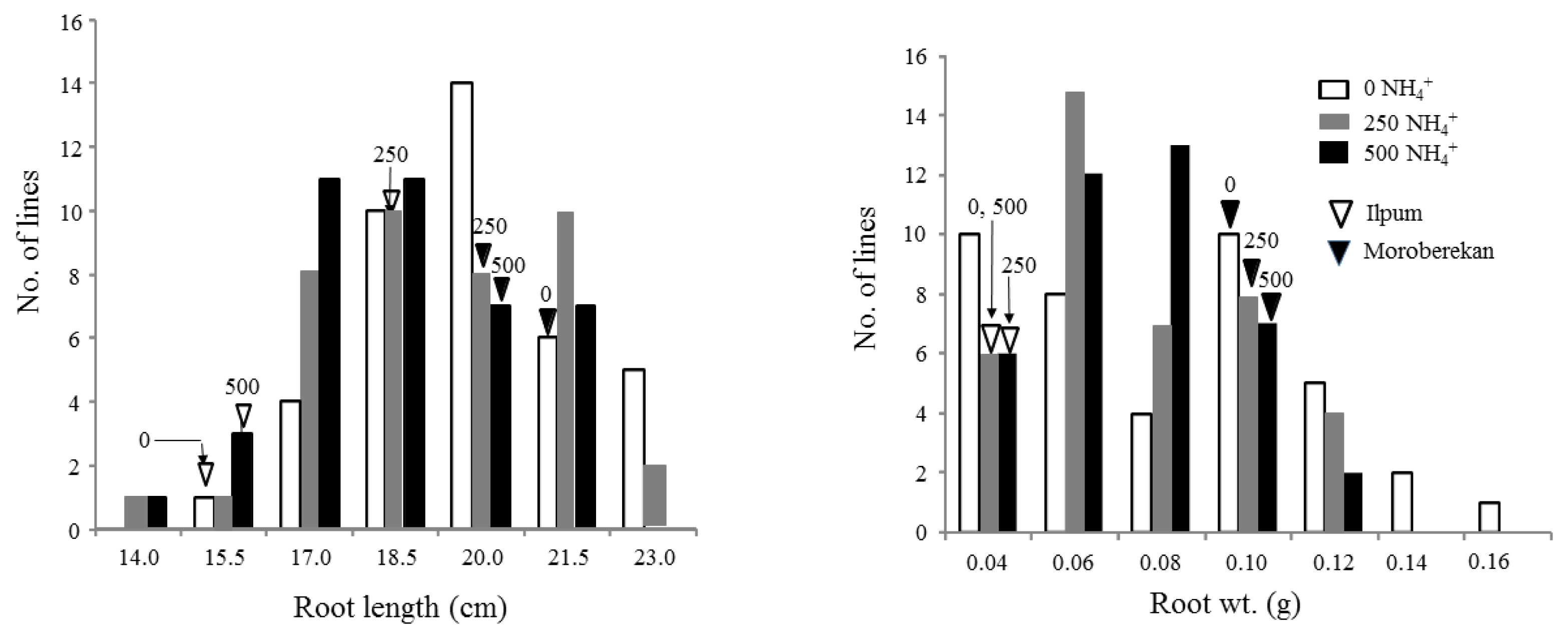
Fig. 3Chromosomal regions affecting root length and weight of seedlings grown in hydroponic culture. Open, slashed and closed columns on the left side of the chromosomes 1, 2, 4, 6, 8 and 10 indicate the most likely chromosomal regions of the QTL for root length and weight in 0, 250 and 500 μM NH4+, respectively.
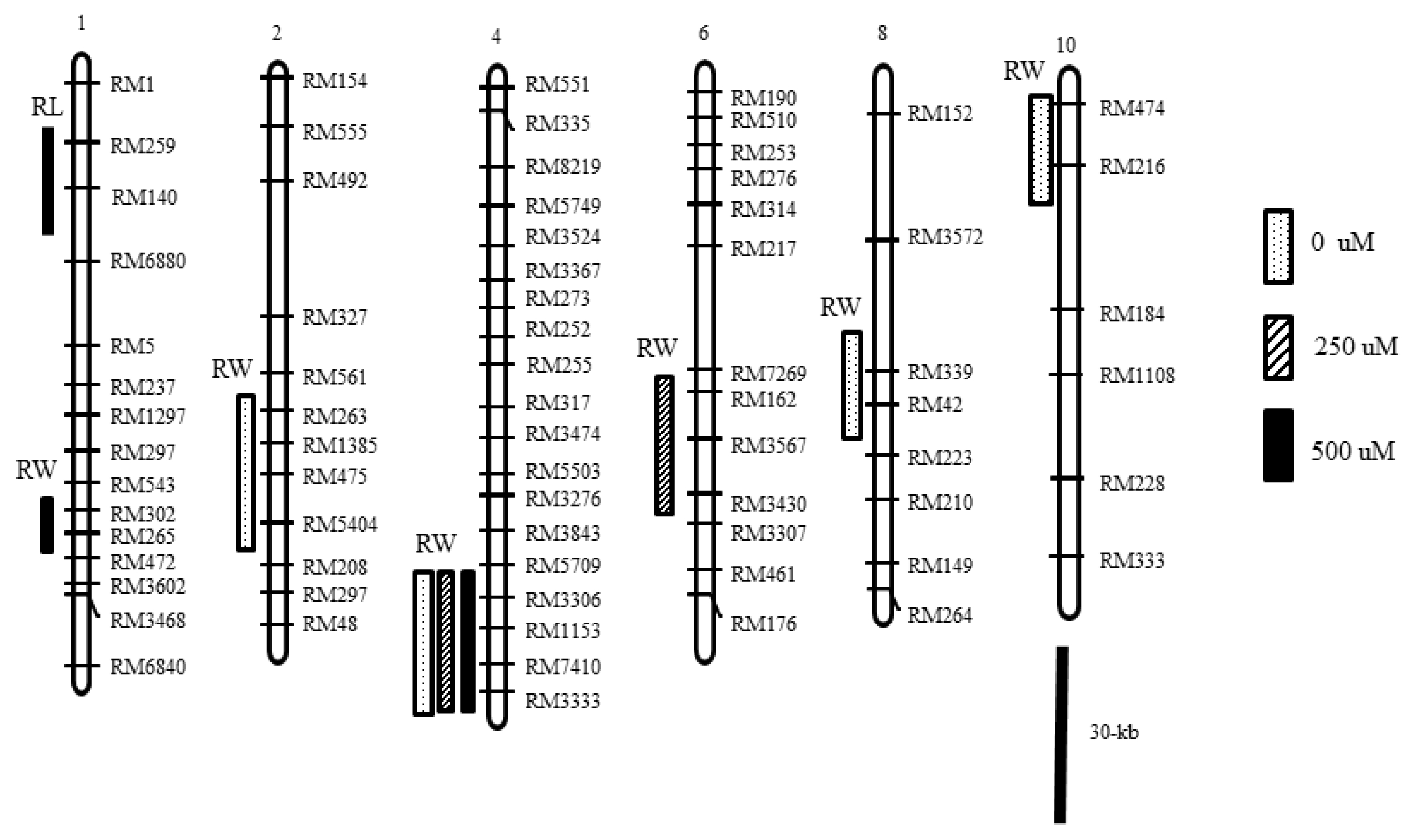
Table 1Comparison of two root traits among 4 accessions grown in hydroponic conditions at three different NH4+ levels.
Table 1
|
Traitz)
|
NH4+ (μM) |
Ilpumbyeo |
Moroberekan |
Hwaseongbyeo |
O. rufipogon
|
|
0 |
15.5 ± 1.8 by),bx)
|
20.4 ± 2.7 a,a |
15.9 ± 2.6 a,b |
21.2 ± 2.2 a,a |
|
RL |
250 |
18.8 ± 2.1 a,a |
19.9 ± 2.8 a,a |
17.1 ± 2.9 a,b |
20.3 ± 2.8 a,a |
|
500 |
15.7 ± 2.0 b,b |
19.5 ± 2.4 a,a |
14.3 ± 2.4 b,b |
20.7 ± 1.4 a,a |
|
|
0 |
0.05 ± 0.01 b,b |
0.09 ± 0.01 a,a |
0.05 ± 0.01 b,b |
0.11 ± 0.01 a,a |
|
RW |
250 |
0.06 ± 0.01 a,b |
0.10 ± 0.01a,a |
0.06 ± 0.02 a,b |
0.11 ± 0.01 a,a |
|
500 |
0.05 ± 0.01 b,b |
0.10 ± 0.00 a,a |
0.07 ± 0.02 a,b |
0.11 ± 0.01 a,a |
Table 2Characteristics of the QTLs detected in the 40 ILs.
Table 2
|
Chr. |
Level |
QTL |
Marker |
Trait |
P
|
Trait |
meansz)
|
|
|
II |
MM |
|
1 |
500 |
qRL1
|
RM259 - RM140 |
RL |
0.01 |
17.6(33)y)
|
18.8(4) |
|
1 |
500 |
qRW1
|
RM302 - RM265 |
RW |
0.01 |
0.10(38) |
0.13(2) |
|
2 |
0 |
qRW2
|
RM263 - RM5404 |
RW |
0.01 |
0.11(35) |
0.14(5) |
|
4 |
0 |
qRW4
|
RM7410 - RM3333 |
RW |
0.01 |
0.12(30) |
0.08(7) |
|
4 |
250 |
qRW4
|
RM7410 - RM3333 |
RW |
0.01 |
0.11(30) |
0.09(7) |
|
4 |
500 |
qRW4
|
RM7410 - RM3333 |
RW |
0.01 |
0.11(30) |
0.09(7) |
|
6 |
250 |
qRW6
|
RM7269 - RM3430 |
RW |
0.005 |
0.09(37) |
0.14(3) |
|
8 |
0 |
qRW8
|
RM339 - RM42 |
RW |
0.01 |
0.10(35) |
0.13(5) |
|
10 |
0 |
qRL10
|
RM474 - RM216 |
RL |
0.01 |
18.8(36) |
20.8(4) |
Table 3Comparison of agronomic traits of four lines with Ilpum.
Table 3
|
Line |
DTH (days) |
CL (cm) |
SPP (no.) |
GW (g) |
RL (cm) |
RW (g) |
Moroberekan introgression segments |
|
|
No. |
Chr. |
|
IL7 |
115az)
|
84a |
122b |
21.2b |
18.2a |
0.087b |
3 |
2, 10 (qRW10) |
|
IL12 |
116a |
81a |
145a |
23.4a |
15.4b |
0.142a |
4 |
4, 6, 10 (qRW10), 11 |
|
IL14 |
117a |
85a |
125b |
22.2b |
16.7a |
0.110a |
5 |
1(qRL1), 2, 6(qRW6), 7, 8(qRW8) |
|
IL24 |
114a |
85a |
142a |
23.1a |
16.6a |
0.107a |
2 |
4, 6 (qRW6) |
|
Ilpum |
116a |
84a |
128b |
21.8b |
14.6b |
0.090b |
|
|
References
REFFERENCES
- Balkos KD, Britto DT, Kronzucker HJ. 2010. Optimization of ammonium acquisition and metabolism by potassium in rice (Oryza sativa L. cv. IR-72). Plant, Cell & Environment. 33: 23-34.
- Brar DS, Khush GS. 1997. Alien introgression in rice. Plant Mol Biol. 35: 35-47.
- Britto DT, Kronzucker HJ. 2002. NH4+ toxicity in higher plants: a critical review. J of Plant Physiology. 159: 567-584.
- Chen G, Guo S, Kronzucker HJ, Shi W. 2013. Nitrogen use efficiency (NUE) in rice links to NH4+ toxicity and futile NH4+ cycling in roots. Plant and Soil. 369: 351-363.
- Cho Y, Jiang W, Chin J, Piao Z, Cho Y, McCouch SR, Koh HJ. 2007. Identification of QTLs associated with physiological nitrogen use efficiency in rice. Mol Cells. 23: 72-79.
- Craswell ET, Vlek PLG. 1983. Fate of fertilizer nitrogen applied to wetland rice. Freney JR, Simpson JR, editors. Gaseous Loss of Nitrogen from Plant-Soil Systems. Springer. Netherlands: pp. 237-264.
- Fernández-Crespo E, Camañes G, García-Agustín P. 2012. Ammonium enhances resistance to salinity stress in citrus plants. Journal of Plant Physiology. 169: 1183-1191.
- Frink CR, Waggoner PE, Ausubel JH. 1999. Nitrogen fertilizer: retrospect and prospect. Proc Natl Aca Sci. 96: 1175-1180.
- Guo S, Chen G, Zhou Y, Shen Q. 2007. Ammonium nutrition increases photosynthesis rate under water stress at early development stage of rice (Oryza sativa L). Plant and Soil. 296: 115-124.
- Imai I, Kimball JA, Conway B, Yeater KM, McCouch S, McClung A. 2013. Validation of yield-enhancing QTLs from a low-yielding wild ancestor of rice. Mol Breed. 32: 101-120.
- Ju H, Jeong O, Choi K, Yoon D, Xie X, Ahn S, Jeong E. 2004. Evaluation of Agronomic Traits of an Ilpumbyeobyeo× Moroberekan BC3F2Population. Korean J of Breeding. 36: 338-344.
- Kang JW, Ju H, Yang P, Ahn SN. 2011. Mapping QTLs for drought tolerance using an introgression line population from a cross between Ilpumbyeobyeo and Moroberekan in rice. CNU Journal of Agricultural Sci. 38: 199-204.
- Kim DM, Ju HG, Kwon TR, Oh CS, Ahn SN. 2009. Mapping QTLs for salt tolerance in an introgression line population between Japonica cultivars in rice. J of Crop Sci and Biotec. 12: 121-128.
- Kim DM, Lee HS, Kwon SJ, Fabreag ME, Kang JW, Yun YT, Chung CT, Ahn SN. 2014. High-density mapping of quantitative trait loci for grain-weight and spikelet number in rice. Rice. 7: 14
- Lilley JM, Ludlow MM, McCouch SR, O’Toole JC. 1996. Locating QTL for osmotic adjustment and dehydration tolerance in rice. J Exper Botany. 47: 1427-1436.
- MacMillan K, Emrich K, Piepho HP, Mullins CE, Price AH. 2006. Assessing the importance of genotype × environment interaction for root traits in rice using a mapping population II: conventional QTL analysis. Theor Appl Genet. 113: 953-964.
- Nelson JC. 1997. QGENE: software for marker-based genomic analysis and breeding. Molecular Breeding. 3(3): 239-245.
- Obara M, Takeda T, Hayakawa T, Yamaya T. 2011. Mapping quantitative trait loci controlling root length in rice seedlings grown with low or sufficient supply using backcross recombinant lines derived from a cross between Oryza sativa L. and Oryza glaberrima Steud. Soil Sci and Plant Nutrition. 57: 80-92.
- Obara M, Tamura W, Ebitani T, Yano M, Sato T, Yamaya T. 2010. Fine-mapping of qRL6.1, a major QTL for root length of rice seedlings grown under a wide range of NH4+ concentrations in hydroponic conditions. Theor Appl Genet. 21: 535-547.
- Ogawa S, Selvaraj MG, Fernando AJ, Lorieux M, Ishitani M, McCouch S, Arbelaez JD. 2013. N- and P-mediated seminal root elongation response in rice seedlings. Plant and Soil. 375: 303-315.
- Ogawa S, Valencia MO, Ishitani M, Selvaraj MG. 2014. Root system architecture variation in response to different NH4+ concentrations and its association with nitrogen-deficient tolerance traits in rice. Acta Physiologiae Plantarum. 36: 2361-2372.
- Patil J, Miller AJ, Gaur H. 2013. Effect of nitrogen supply form on the invasion of rice roots by the root-knot nematode, Meloidogyne graminicola. Nematology. 15: 483-492.
- Price A, Tomos A, Virk D. 1997. Genetic dissection of root growth in rice (Oryza sativa L.) I: A hydroponic screen. Theor Appl Genet. 95: 132-142.
- Roosta H, Schjoerring JK. 2008. Root carbon enrichment alleviates ammonium toxicity in cucumber plants. J of Plant Nutrition. 31: 941-958.
- Ryan BF, Joiner BL, Ryan TA. Minitab handbook. PWS-KENT. Boston, MA: 1992.
- Shimizu A, Yanagihara S, Kawasaki S, Ikehashi H. 2004. Phosphorus deficiency-induced root elongation and its QTL in rice (Oryza sativa L. Theor Appl Genet. 109: 1361-1368.
- Socolow RH. 1999. Nitrogen management and the future of food: lessons from the management of energy and carbon. Proc Natl Acad Sci. 96: 6001-6008.
- Steele KA, Price AH, Shashidhar HE, Witcombe JR. 2006. Marker-assisted selection to introgress rice QTLs controlling root traits into an Indian upland rice variety. Theor Appl Genet. 112: 208-221.
- Su C, Yin B, Zhu Z, Shen Q. 2005. Gaseous loss of nitrogen from fields and wet deposition of atmospheric nitrogen and their environmental effects. Soils. 37: 113-120.
- Wei D, Cui K, Ye G, Pan J, Xiang J, Huang J, Nie L. 2012. QTL mapping for nitrogen-use efficiency and nitrogen-deficiency tolerance traits in rice. Plant and Soil. 359: 281-295.
- Xu C, Li X, Xue Y, Huang Y, Gao J, Xing YZ. 2004. Comparison of quantitative trait loci controlling seedling characteristics at two seedling stages using rice recombinant inbred lines. Theor Appl Genet. 109: 640-647.
- Yamaya T, Oaks A. 2004. Metabolic regulation of ammonium uptake and assimilation. Amancio S, Stulen I, editors. Nitrogen acquisition and assimilation in higher plants. Kluwer. Dordrecht: pp. 35-63.
- Yoshida S, Forno DA, Cock JH, Gomez KA. 1976. Laboratory manual for physiological studies of rice. 3rd edn. IRRI. Manila.
- Zhao C, Zhou L, Zhang Y, Zhu Z, Chen T, Zhao Q, Yao S, Yu X, Wang C. 2014. QTL mapping for seedling traits associated with low-nitrogen tolerance using a set of advanced backcross introgression lines of rice. Plant Breeding. 133: 189-195.