Abstract
The allelic variations at the Rht-1, Vrn-1 and Ppd-1 of 410 Korean wheat cultivars, including 111 Korean experimental lines, 238 Korean landraces and 61 North Korean collections, were investigated to provide the information of plant height and heading date and to elucidate the relationship between those traits and allelic variation of these genes because earliness is major consideration in Korean wheat production. All Korean wheats displayed vrn-A1 and Ppd-A1b alleles, while Rht-B1a, Rht-D1a, vrn-B1, Vrn-D1, Ppd-B1b and Ppd-D1a alleles were also predominantly found. Most Korean wheats carried both Rht-B1a and Rht-D1a alleles, both vrn-B1 and Vrn-D1 alleles, or both Ppd-B1b and Ppd-D1a alleles. The Rht-B1a, vrn-D1, Ppd-B1b and Ppd-D1b alleles were found to exhibit longer culm and spike length than their counterpart alleles. The Rht-B1a allele also showed longer spike length than Rht-B1b. Vrn-B1b and vrn-D1 alleles exhibited longer days to heading date than their counterpart alleles at the Vrn-B1 and Vrn-D1 loci. Lines carrying both Rht-B1b and Rht-D1b alleles displayed shorter culm and longer spike length and days to heading date than any other combination of alleles at the Rht-B1 and Rht-D1 loci. In contrast, lines carrying both Ppd-B1b and Ppd-D1b alleles exhibited longer culm and spike length than any other combination of alleles at the Ppd-B1 and Ppd-D1 loci.
-
Key words: Korean wheat, Allele, Rht-1, Vrn-1, Ppd-1
INTRODUCTION
Semi-dwarfing genes, especially
Rht-B1b and
Rht-D1b, have been used as the main components of the green revolution, as these genes are insensitive to exogenous gibberellic acid (GA3,
Tosovic-Maric et al. 2008). The
Rht-B1 and
Rht-D1 loci located on chromosome 4B and 4D, respectively, encode DELLA proteins, for which polymorphisms result in different alleles which have different effects on plant height (
Börner et al. 1996). These semi-dwarfing genes have influenced the reduction of plant height and increased grain yield of wheat grown in favorable climatic conditions, albeit the grain yield was found to be reduced under high temperature and drought conditions (
Flintham et al. 1997).
The flowering time in wheat, though complex in character, is mainly controlled by vernalization (
Vrn) and photoperiod (
Ppd) response (
Kamran et al. 2014). These response genes hasten or delay flowering in response to environmental stimuli, by which floral initiation is determined through interactions with growth temperatures (
Kamran et al. 2014). The
Vrn-1 and
Ppd-1 alleles include three orthologous genes at each locus, and are located on the homoeologous group 5 and 2 chromosomes, respectively (
Kamran et al. 2014). The
Vrn-1 alleles are known to have differential effects on flowering time, plant height and various yield components (
Stelmakh 1998). Spring habit
Vrn-A1a allele is predominant in wheat cultivars grown at high latitudes, while the
Vrn-D1 allele is predominantly found in cultivars from Asia and in Japanese and Chinese landraces, with particular prevalence in sub-tropical regions where the growing season is longer (
Kamran et al. 2014). Photoperiod insensitive alleles,
Ppd-A1a,
Ppd-B1a and
Ppd-D1a, were observed to positively affect the yield when wheat was grown in summer under hot and dry conditions, because these alleles displayed earlier flowering date and larger grain than the photoperiod sensitive alleles (
Snape et al. 2001).
The wheat consumed in Korean has mostly been imported from Australia, Canada and the United States, though the per capita wheat consumption increased to 32.3 kg in 2013 (
MAF 2014). The Korean government has recently been pushed to increase wheat production up to 200,000 MT by 2020, which was targeted to increase the country’s self-sufficiency rate from its current level of 1.2% to 10%. Korean wheat has typically been cultivated by double-cropping with rice, as the time of the wheat harvest is overlapped with the rainy seasons and the time for rice transplanting in Korea. Therefore, earliness, as well as grain yield, have been the most important criteria considered in Korean wheat breeding programs. Korean wheat breeders have been focusing on achieving improved grain yield through the selection of wheat lines with reduced plant height and shortened days to flowering since the 1970s. Consequently, the Korean wheat cultivars have narrow genetic variations in semi-dwarf genes, growth habits and photoperiod response (
Cho et al. 2015a,
b). Therefore, the present study was conducted to investigate the distribution of the
Rht-1,
Vrn-1 and
Ppd-1 alleles in Korean experimental lines and landraces using molecular markers, and to elucidate the relationship between agronomic traits and allelic variation of these genes.
MATERIALS AND METHODS
Plant materials
A total of 410 Korean experimental lines and landraces, including 111 Korean experimental lines, 238 Korean landraces and 61 North Korean collections, were kindly provided by the National Agrobiodiversity Center, National Academy of Agricultural Science of the Rural Development Administration (Suwon, Korea). Korean experimental lines contained wheat lines developed from 1950s to 2010s in National Institute of Crop Science (NICS), Rural Development Administration (RDA). Korean landraces and North Korean collections mainly collected in the past 1950s. These lines were sown in the Upland Crop Experimental Farm of the NICS, RDA in 2013 on 50% clay loam soil. The seeds were sown on October 25 with each plot consisting of three 2-m rows spaced 25 cm apart which were performed in randomized complete blocks with 3 replicates, and the plots were combine-harvested on June 20. Fertilizer was applied at a ratio of 5:7:5kg/10a (N:P:K) before sowing, and the plots were stringently controlled for weeds, insects and disease. No supplemental irrigation was applied.
Agronomic traits
Agronomic traits were recorded as follows: days to heading date as the number of days from last October 25 to exertion of the spike from the stem in 50 % of a plot; culm length as the length from ground level to the base of the spike; and spike length as the length of the spike excluding the awn. Culm length and spike length were determined from a sample of 10 main tillers randomly selected from each plot at maturity.
PCR conditions
Five seeds of each line were grown in a temperature-controlled greenhouse to analyze the allelic variations of
Rht-B1,
Rht-D1,
Vrn-A1,
Vrn-B1,
Ppd-A1,
Ppd-B1 and
Ppd-D1. Two weeks after germination, single leaves taken from five plants of each cultivar were collected, bulked and snap-frozen in liquid nitrogen, and then stored at −80°C until needed. All plants were kept in the greenhouse for analysis of allelic variations. Genomic DNA was extracted from young leaf tissue (100 mg) using a genomic DNA prep kit (Solgent Co., Korea) according to the manufacturer’s instructions. PCR for
Rht-B1 and
Rht-D1 was carried out using allele-specific markers according to the methods of
Ellis et al. (2002) and
Zhang et al. (2006). The primers for
Vrn-1 allele-specific markers and PCR followed the methods of
Yan et al. (2004),
Fu et al. (2005),
Santra et al. (2009),
Zhang et al. (2012) and
Chen et al. (2013). For PCR of
Ppd-1, the allele-specific markers and methods from
Beales et al. (2007) and
Bentley et al. (2011) were employed. Amplified PCR fragments were separated on 1.5% agarose gels, stained with ethidium bromide, and visualized using UV light.
Statistical analysis of the data was performed by SAS software (SAS Institute, NC, USA) using Fisher’s least significant difference test (LSD), analysis of variance (ANOVA) and pair-wise t-test. Analysis of variance was conducted using the general linear model (GLM) procedure. The GLM procedure was used for the randomized complete block (RCB) analysis. Sources of variation in the model were considered to be fixed effects. PROC GLM was used to estimate the relative contribution of the tested loci for the measured traits of Korean wheats. Pearson correlations between pairs of traits were determined from the least square means of wheat cultivars for each year separately, and over all environments, using the Corr procedure.
RESULTS
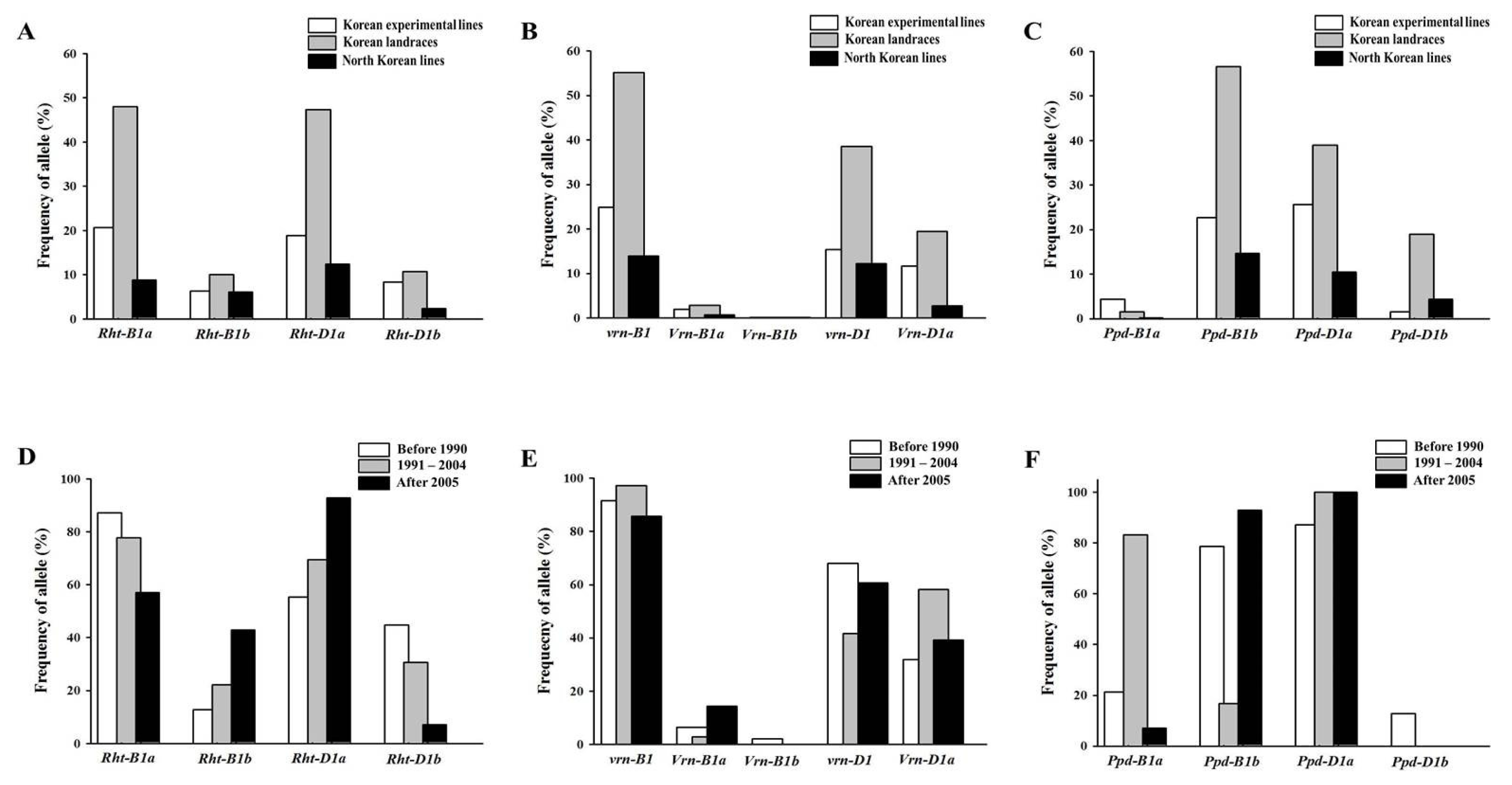
Genetic variations at Rht-1, Vrn-1 and Ppd-1 loci
Genetic variations at the
Rht-1,
Vrn-1 and
Ppd-1 loci in the Korean wheats are shown in
Fig. 1 and
2, and summarized in
Table 1. Thirteen types of allelic composition were identified among the 410 Korean wheats. Two types of alleles were found at the
Rht-B1, Rht-D1, Vrn-D1, Ppd-B1 and
Ppd-D1 loci, and three types of alleles found at the
Vrn-B1 locus, while all of the Korean wheats tested displayed
vrn-A1 and
Ppd-A1b alleles. The Korean wheats showed higher frequency of
Rht-B1a,
Rht-D1a,
vrn-B1,
Vrn-D1,
Ppd-B1b and
Ppd-D1a alleles (77.6, 78.5, 93.9, 66.1, 93.9 and 75.1%, respectively) than their counterpart alleles at the same loci. Among the 111 Korean experimental lines tested, 85 (76.6%) carried
Rht-B1a and 77 (69.4%) displayed
Rht-D1a at the
Rht-B1 and
Rht-D1 loci, respectively. The
vrn-B1,
Ppd-B1b and
Ppd-D1a alleles were predominantly found at the
Vrn-B1 and
Ppd-1 loci (91.9, 83.8 and 94.6%, respectively). The Korean experimental lines showed similar frequency of
Vrn-D1 and
Vrn-D1a alleles (56.8 and 43.2%, respectively). The
Rht-D1a,
vrn-B1,
Vrn-D1 and
Ppd-B1b alleles (83.6, 93.4, 82.0 and 98.4%, respectively) were found to be dominant in the 61 North Korean collections. In contrast, the North Korean collections showed similar frequencies of the
Rht-B1a and
Rht-B1b alleles (59.0 and 41.0%, respectively), while the frequencies of the
Ppd-D1a and
Ppd-D1b alleles (70.5 and 29.5%, respectively) were similar to those of Korean landraces.
Korean experimental lines released after 2005 exhibited lower frequency of the Rht-B1a allele (57.1%) and higher frequency of the Rht-D1a allele (92.9%) than the experimental lines released before 2004 (> 77.8% and < 69.4%, respectively). However, the experimental lines mainly possessed the vrn-B1 allele (> 85.7%) regardless of the year released. The frequency of the Vrn-B1a allele increased in lines released after 2005, while only one line, Suwon229, had the Vrn-B1b allele. The Korean experimental lines released between 1991 and 2004 displayed higher frequency of the Vrn-D1a (58.3%) and Ppd-B1a (83.3%) alleles than the experimental lines released before 1990 or after 2005 (< 39.3% and < 21.3%, respectively). Only two lines released after 2005, Iksan311 and Iksan313, contained the Ppd-B1a allele. All Korean experimental lines were found to possess the Ppd-D1a allele, except six lines released before 1990.
The frequency of lines carrying both the Rht-B1a and Rht-D1a alleles was higher (56.8%) than that of any other combination (< 22.0%). In particular, those alleles were dominantly found in Korean landraces (64.7%). In contrast, the experimental lines showed higher frequency of both Rht-B1a and Rht-D1b alleles (30.6%) than the other lines, while North Korean collections exhibited a higher frequency of both Rht-B1b and Rht-D1a alleles (37.7%). Only two North Korean collections, Seuyuk28 and Seuyuk44, carried both the Rht-B1b and Rht-D1b alleles. Most of the Korean wheats carried either both the vrn-B1 and Vrn-D1 alleles (61.0%), or both the vrn-B1 and Vrn-D1a alleles (32.9%). Only 20 lines were found to carry both the Vrn-B1a and Vrn-D1 alleles, while three carried both the Vrn-B1a and Vrn-D1a alleles. One North Korean collection, IT215959, displayed both the Vrn-B1b and vrn-D1 alleles, while one Korean experimental line, Suwon229, carried both the Vrn-B1b and Vrn-D1a alleles. Though about 70% of Korean wheats carried both the Ppd-B1b and Ppd-D1a alleles, only 6.1% carried both the Ppd-B1a and Ppd-D1a alleles. Only one of the North Korean collections, IT246394, and 6 Korean landraces carried both the Ppd-B1a and Ppd-D1a alleles. Six of the Korean experimental lines displayed both the Ppd-B1b and Ppd-D1b alleles.
Relationship between allelic variations and agronomic traits
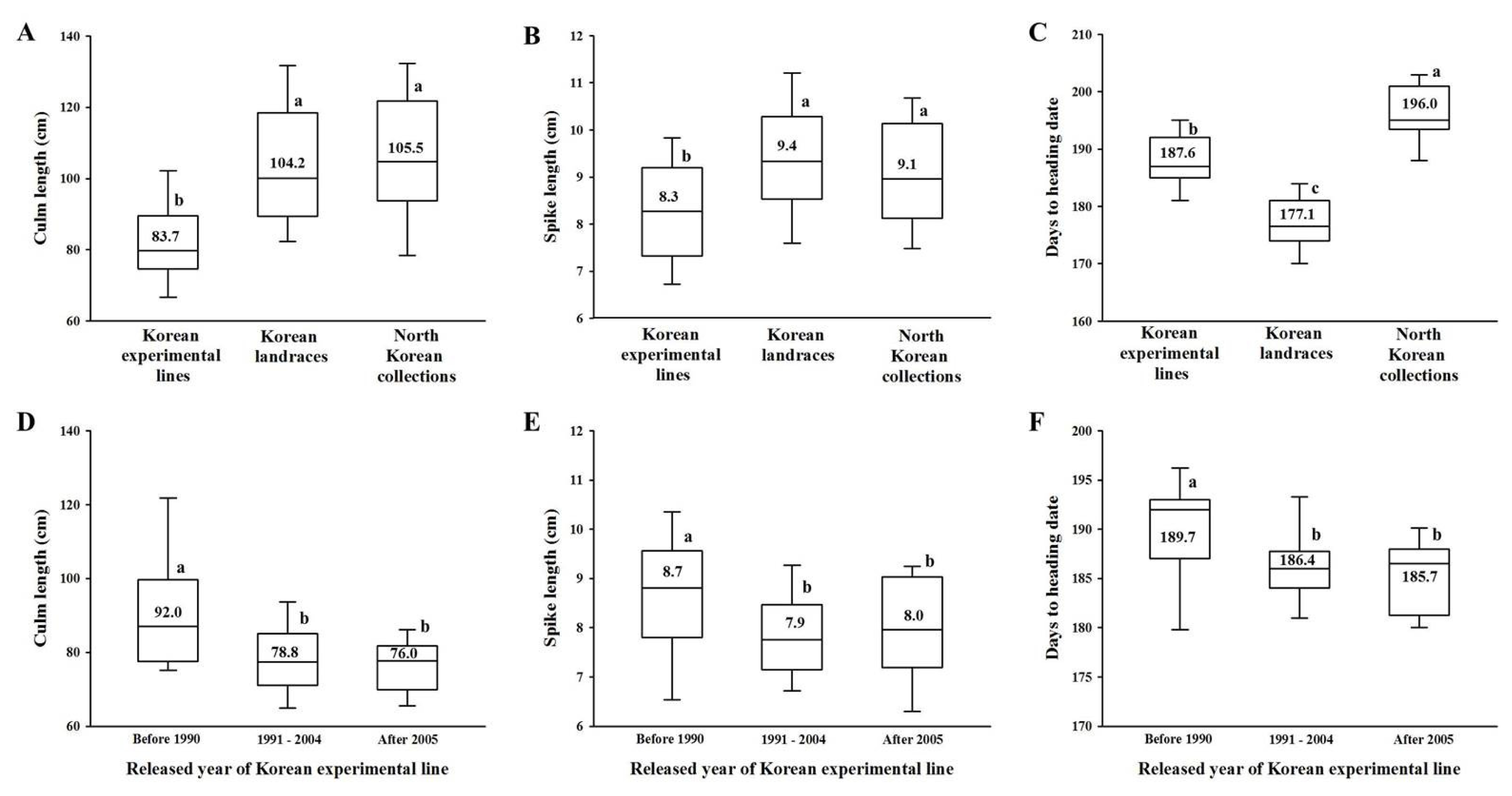
The ranges of the culm and spike length, and days to heading date of 410 Korean wheats are shown in
Fig. 3. The mean values of the agronomic traits were 98.8 cm for culm length, 9.1 cm for spike length and 182.8 days for days to heading date. The culm and spike length, and days to heading date of the Korean wheats ranged from 48.8 – 161.7 cm, 4.9 – 13.3 cm and 162.0 – 207.0 days, respectively. The Korean experimental lines showed shorter culm and spike length (83.7 and 8.3 cm, respectively) than the Korean landraces (104.2 and 9.4 cm, respectively) and North Korean collections (105.5 and 9.1 cm, respectively). Regarding days to heading date, the experimental lines showed intermediate values (187.6) compared to the Korean landraces (177.1) and North Korean collections (196.0). Of the 111 experimental lines, those released before 1990 displayed longer culm and spike length, as well as days to heading date (92.0 cm, 8.7 cm and 189.7 days, respectively) than those released after 1991 (< 78.8 cm, < 8.0 cm and < 186.4 days, respectively). The culm lengths of Korean experimental lines released after 2005 were shorter than those of lines released between 1991 and 2004, though no significant differences were observed in spike length and days to heading date between those lines. Culm length positively correlated with spike length in the 410 Korean wheats and 111 Korean experimental lines (r = 0.444*** and r = 0.363***, respectively), while culm length also displayed a positive correlation with days to heading date in the 111 experimental lines (r = 0.258**).
Korean wheats with allelic variation at the
Rht-1,
Vrn-1 and
Ppd-1 loci exhibited significant differences in culm and spike length, as well as days to heading date (
Table 1). Those carrying the
Rht-B1a allele exhibited longer culm and spike length (101.4 and 9.2 cm, respectively) than those with
Rht-B1b (90.0 and 8.7 cm, respectively). Those carrying the
Rht-D1a allele also displayed longer culm length (102.3 cm) than those with the
Rht-D1b allele (86.3 cm), but no significant difference was observed in spike length. There were also no significant differences of days to heading date in the Korean wheats according to allelic variations at the
Rht-B1 and
Rht-D1 loci. Two Korean experimental lines released before 1990, Seuyuk28 and Seuyuk28, which carried the
Rht-B1b and
Rht-D1b alleles displayed shorter culm length (82.0 cm) but longer spike length and days to heading date (10.5 cm and 194.5 days, respectively) than those with the combined alleles of both
Rht-B1 and
Rht-D1 (> 87.5 cm, > 9.2 cm and < 184.9 days, respectively). One Korean experimental line and one North Korean collection, Suwon 229 and IT215959, which carried the
Vrn-B1b allele showed longer days to heading date (193.5) than the other Korean wheats carrying the
vrn-B1 and
Vrn-B1a allele (182.7 and 183.1, respectively). However, significant differences of culm and spike length among Korean wheats were not observed according to allelic variations at the
Vrn-B1 locus. Korean wheats carrying the
Vrn-D1a allele showed shorter culm and spike length (95.0 and 8.7 cm, respectively), as well as shorter days to heading date (180.5) than those carrying the
vrn-D1 allele (100.8 cm, 9.3 cm and 183.9 days, respectively). However, no significant differences of culm and spike length, as well as days to heading date, were found among the Korean wheats with a combination of both
Vrn-B1 and
Vrn-D1 alleles. Korean wheats carrying the
Ppd-B1b or
Ppd-D1b allele displayed longer culm and spike length than those with the
Ppd-B1a or
Ppd-D1a allele, though no significant differences in days to heading date were observed. Korean wheats carrying both the
Ppd-B1b and
Ppd-D1b alleles also showed longer culm and spike length than those with both the
Ppd-B1a and
Ppd-D1a or
Ppd-B1b and
Ppd-D1a alleles.
DISCUSSION
Korean wheat cultivars, as well as Chinese and Japan wheats, were found to display low frequency of the
Rht-B1b allele and high frequency of the
Rht-D1b allele (
Yamada 1990;
Zhang et al. 2006;
Cho et al. 2015a). Korean experimental lines also exhibited similar frequency of
Rht-B1 and
Rht-D1 alleles as Korean wheat cultivars (
Cho et al. 2015a). Only two North Korean collections showed both
Rht-B1b and
Rht-D1b alleles, which was similar frequency as previous reports (
Yamada 1990;
Zhang et al. 2006;
Tosovic-Maric et al. 2008). The frequencies of
Rht-B1b and
Rht-D1b alleles increased when compared with the landraces and leading cultivars in China (
Zhang et al. 2006). Korean experimental lines showed higher frequency of
Rht-B1b and
Rht-D1b alleles than Korean landraces and North Korean collections. Therefore, the Korean experimental lines displayed shorter culm length than the others. The
Rht-B1b and
Rht-D1b alleles were reported to reduce plant height by 15% and increase yield by 24% (
Flintham et al. 1997). The experimental lines released after 2005 showed higher frequency of the
Rht-B1b allele than those released before 2004, but the frequency of the
Rht-D1b allele was decreased from 44.7% to 7.1%. In German wheat cultivars, those carrying the
Rht-D1b were reported to be more susceptible to Fusarium head blight (
Knopf et al. 2008). However, no significant difference in the culm and spike length were found, although significant difference in the frequency of the
Rht-D1b allele was observed. The effects of the
Rht-B1b and
Rht-D1b alleles on plant height and grain yield are known to be influenced by the growth conditions (
Flintham et al. 1997;
Korzun et al. 1998). However, the
Rht8 dwarfing gene, which also reduced plant height by around 10%, has higher resistance to environments (
Worland and Law 1986;
Börner et al. 1993;
Worland et al. 1998). Therefore, allelic variations of Korean wheats at the
Rht8 locus and other dwarfing genes should be evaluated to elucidate adaptations for improving the grain yield, including reducing the height for lodging resistance and increasing the disease resistance to factors such as Fusarium head blight.
Allelic variations at the
Vrn-1 loci have differential effects on the flowering time, plant height and grain yield (
Stelmakh 1998). Seven different
Vrn-1 alleles were reported in spring wheats with facultative growth habits:
Vrn-A1a,
Vrn-A1b and
Vrn-A1c at the
Vrn-A1 locus;
Vrn-B1a and
Vrn-B1b at the
Vrn-B1 locus; and
Vrn-D1a and
Vrn-D1b at the
Vrn-D1 locus (
Kamran et al. 2014). Herein, six different alleles were found in the 410 Korean wheats, all of which possessed the
vrn-A1 allele, while three types of alleles were found at the
Vrn-B1 locus and two types at the
Vrn-D1 locus. About 60% of the Korean wheats possessed the
vrn-A1, vrn-B1 and
vrn-D1 alleles, which was a higher frequency than in Korean wheat cultivars (
Cho et al. 2015b). The
Vrn-B1a and
Vrn-B1b alleles were rarely found in the Korean lines, and no significant differences were observed in the frequency of
Vrn-B1 alleles among the 410 whetas tested. However, the North Korean wheat collections showed higher frequency of the
vrn-D1 allele and lower frequency of the
Vrn-D1a allele than the Korean experimental lines and landraces. Conversely, the frequency of the
Vrn-D1a allele increased in the Korean experimental lines compared to Korean landraces and North Korean collection. The
vrn-A1 allele was found to have moderate occurrence in Chinese wheats, while the
Vrn-A1a and
Vrn-B1a alleles appeared frequently in Canadian spring wheats, the
Vrn-A1b or
Vrn-A1c alleles have rarely been observed, and the
Vrn-B1c allele was found to be highly prevalent in Russian spring wheats (
Iqbal et al. 2007;
Zhang et al. 2008;
Shcherban et al. 2012;
Kamran et al. 2014). While the
Vrn-D1a allele was found to be highly prevalent in Indian spring wheats, none of the Canadian spring wheats displayed the
vrn-D1 allele, and only one sample among Russian spring wheats had the
vrn-D1 allele (
Iqbal et al. 2007;
Shcherban et al. 2012;
Singh et al. 2013). The Korean experimental lines released after 2005 showed different frequencies of the
vrn-B1 and
Vrn-B1a alleles, but exhibited similar frequency of the
Vrn-D1 alleles compared to wheats released before 1990. The
Vrn-A1 allele has the strongest effect on inhibiting the vernalization requirement, followed by the
Vrn-D1 and
Vrn-B1 alleles (
Goncharov 2004). Facultative growth habits or the partial vernalization requirement of
Vrn-D1 plays a key role in determining the wider adaptability of wheat cultivars (
Kamran et al., 2014). Recently, more attention has been called to the development of facultative or spring wheat in Korea due to water logging during the sowing season in late October, frequently resulting in reduced growing area. Therefore, wheat lines which could be harvested in late summer after planting in the early spring, and introduction of the
Vrn-A1a or
Vrn-A1b alleles should be considered for the development of facultative or spring wheat. The
Vrn-B1b and
vrn-D1 alleles had to be involved longer days to heading date in the 410 Korean wheats. The heading date of wheat is known to be determined 70 – 75 % by the growing habits and 20 – 25% by the photoperiod response (
Stelmakh 1998). The rainy season has generally been overlapped with the harvest time of wheat in the southern part of Korea, which is closely related to the occurrence of pre-harvest sprouting at the maturation stage. Therefore, earliness has been a major consideration for wheat growing of Korea, and increase of the genetic diversity at the
Vrn-1 alleles, as well as investigation of other
Vrn alleles should be considered in Korean wheat breeding programs.
Photoperiod response plays a key role in accelerating or delaying the heading time under field conditions in vernalized wheats, for which the
Ppd-1 genes are located on the homoeologous group 2 chromosomes (
Kamran et al. 2014). The
Ppd-1a alleles are known to confer insensitivity in response to the photoperiod, while the
Ppd-1b alleles are related to sensitive response (
Snape et al. 2001;
Kamran et al. 2014). Photoperiod insensitive cultivars were also preferred in the southern regions of Europe and North America, whereas cultivars in the northern regions were generally photo sensitive (
Worland et al. 1994;
Dyck et al. 2004). Wheats carrying
Ppd-1a alleles displayed earlier flowering date and larger grain yield than those carrying the
Ppd-1b alleles in Southern and Eastern Europe, wherein the summer conditions were hot and dry (
Snape et al. 2001).
Ppd-D1 is the most photoperiod insensitive locus, followed by
Ppd-B1 (
Worland 1996). All of the 410 Korean wheats tested were found to have the
Ppd-A1b allele, while higher frequency of the
Ppd-B1b allele than the
Ppd-B1a allele was observed, in spite of the high frequency of the
Ppd-D1a allele. In addition, the Korean experimental lines showed higher frequency of the
Ppd-B1b allele than the Korean landraces and North Korean collections. Furthermore, the experimental lines released between 1991 and 2004 showed higher frequency of the
Ppd-B1b allele than the others. The
Ppd-D1a allele is also wide spread in most of the European, Canadian and Chinese insensitive cultivars (
Kamran et al. 2014). All of the Korean wheat cultivars also possessed the
Ppd-A1b and
Ppd-D1a alleles, and about 90% of cultivars exhibited the
Ppd-B1b allele (
Cho et al. 2015b). Almost all of the improved cultivars released in China after 1970 possessed the
Ppd-D1a allele, whereas below 50% of Chinese landraces had this allele (
Yang et al. 2009). The pleiotropic influence of the
Ppd-D1 gene on the yield potential of the major wheat growing regions of Europe could be involved in reducing the flowering time, plant height and number of spikelets per spike (
Snape et al. 2001;
Foulkes et al. 2004;
Kamran et al. 2014). Almost all high-yield improved cultivars are insensitive to photoperiod, while all landraces in North Africa, Mediterranean regions and Asia are sensitive, because selection of the photoperiod insensitive trait has enhanced the adaptability of the improved cultivars to a wide range of environments in comparison with the photoperiod-sensitive landraces in these regions (
Kamran et al. 2014). Herein, no significant difference in days to heading date according to the allelic variation at
Ppd-1 loci was observed, although wheats with the
Ppd-B1b or
Ppd-D1b allele showed longer culm and spike length than those with the
Ppd-B1a or
Ppd-D1a allele. The
Ppd-B1b allele, when combined with
Vrn-D1b alleles, could be involved in shortening the days to maturation and increasing the grain yield in Korean wheat cultivars (
Cho et al. 2015b). The introduction of the
Ppd-A1a allele and increase of the frequency of the
Ppd-B1a allele should be considered for reducing the flowering time of Korean wheats, and further investigation is also needed to elucidate the relationship between specific alleles combined with
Vrn-1 and
Ppd-1 alleles and the major agronomic traits in Korean wheat breeding programs.
ACKNOWLEDGEMENTS
This work was carried out with the support of the “Cooperative Research Program for Agriculture Science & Technology Development” (Project title: Improvement of crop characteristics of supernumerary spike of wheat for stable production and incensement of yield, Project No. PJ009220), Rural Development Administration, Republic of Korea.
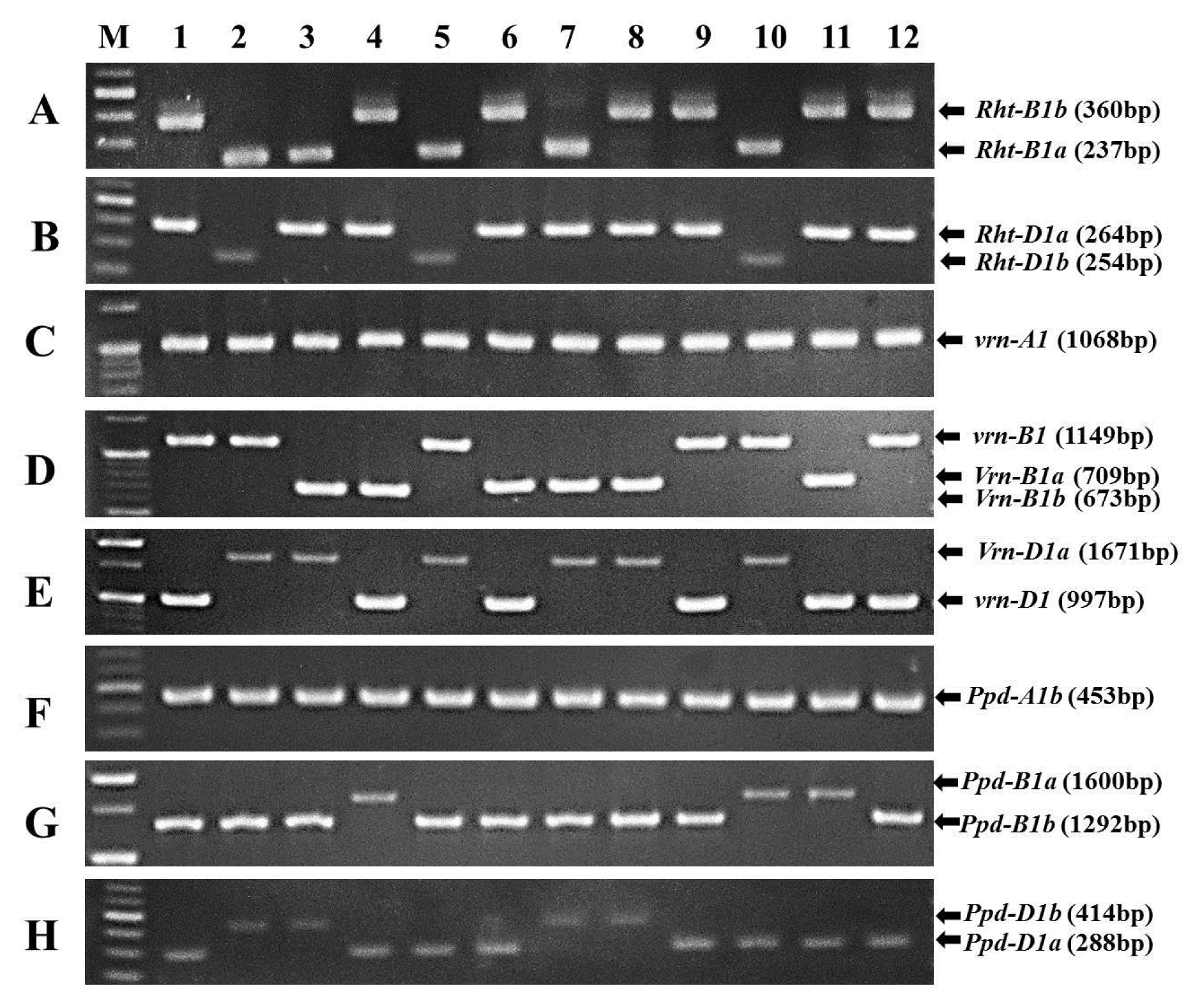
Fig. 1PCR patterns of Rht-B1 (A), Rht-D1 (B), Vrn-A1 (C), Vrn-B1 (D), Vrn-D1 (E), Ppd-A1 (F), Ppd-B1 (G) and Ppd-D1 (H) alleles in Korean wheats. M, molecular size marker.
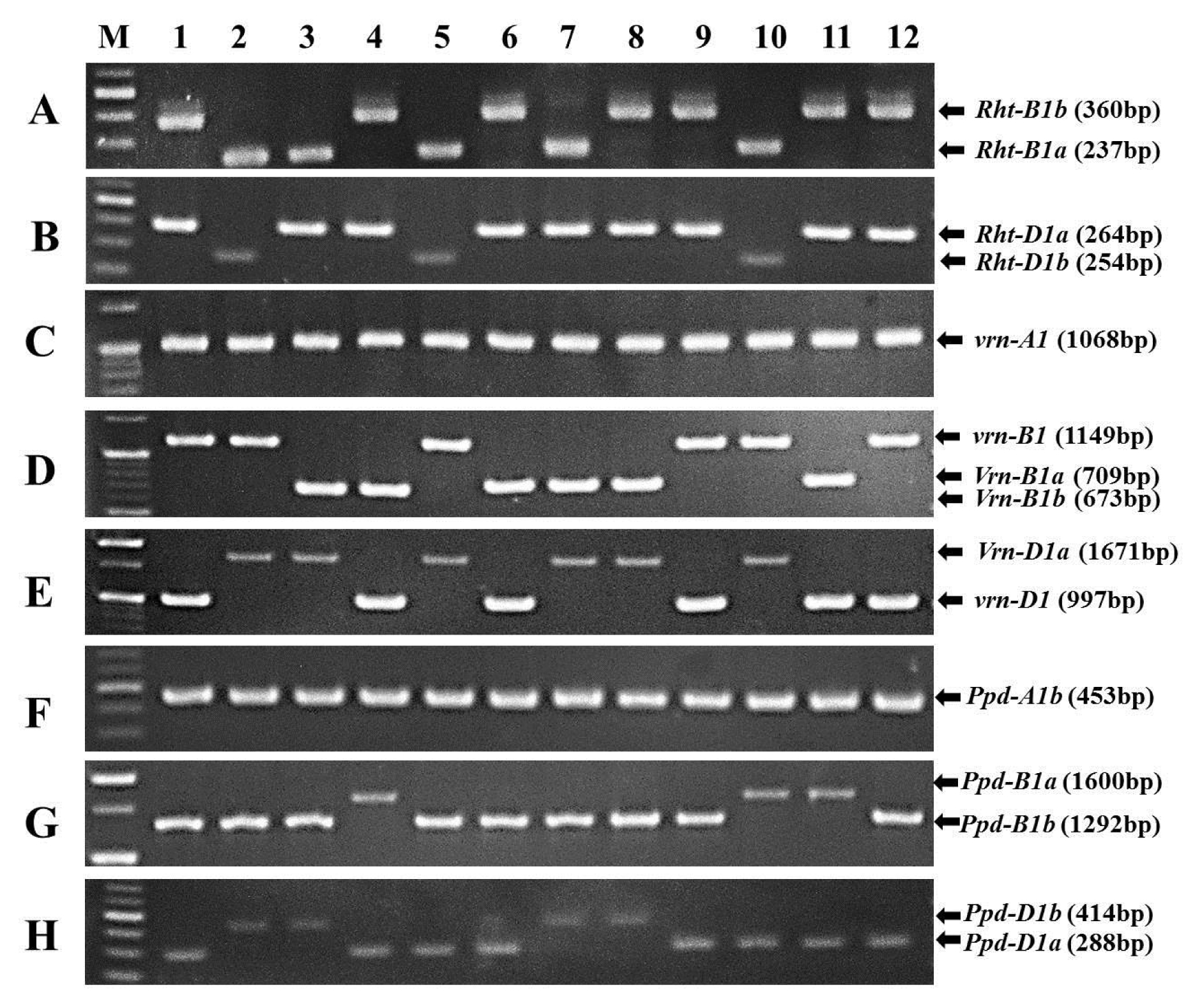
Fig. 2Frequency of Rht-1 (A), Vrn-1 (B) and Ppd-1 (C) alleles in 410 Korean wheats, and frequency of Rht-1 (D), Vrn-1 (E) and Ppd-1 (F) alleles in 111 Korean experimental lines according to the year released.
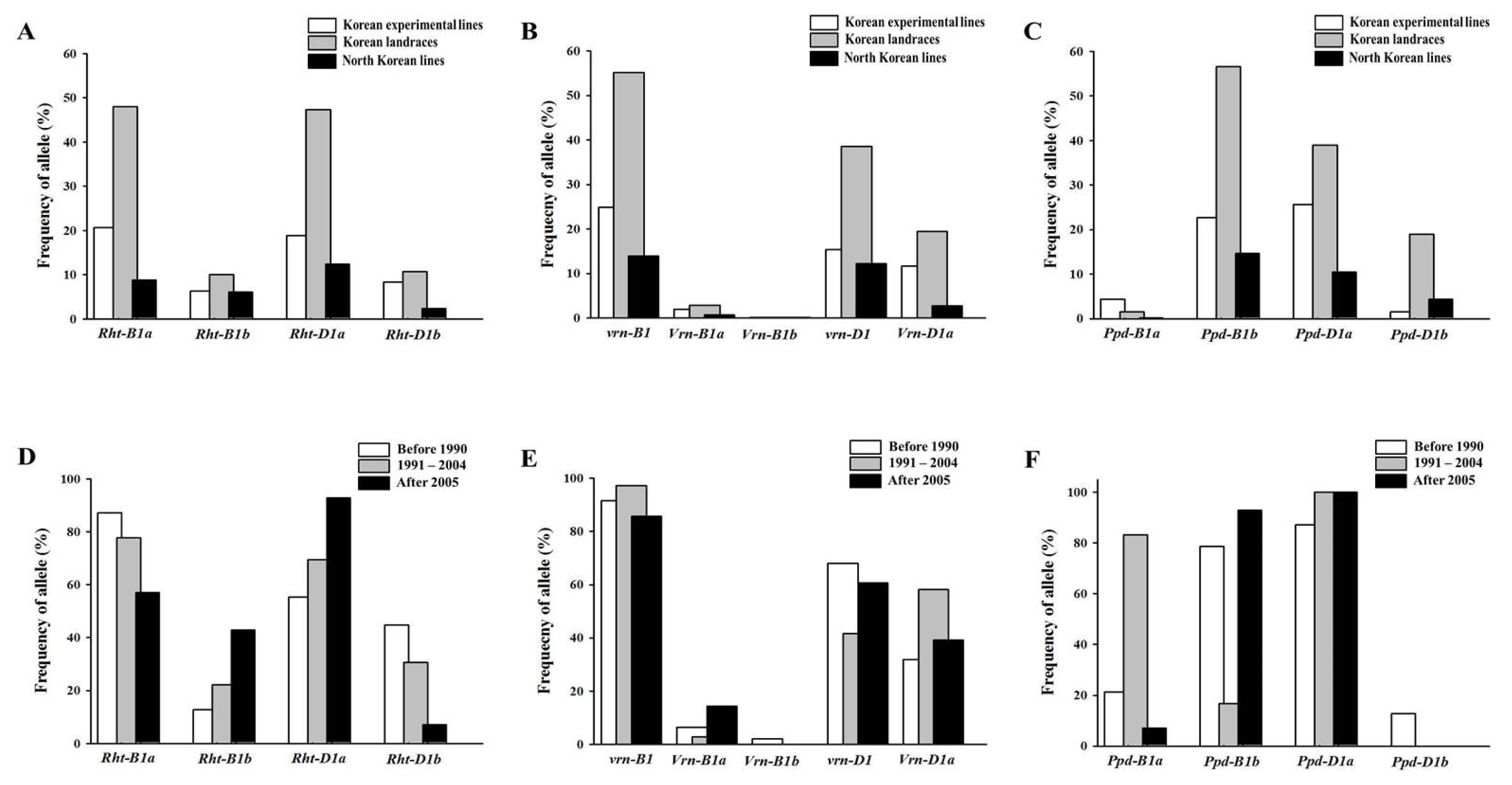
Fig. 3Frequency of culm length (A), spike length (B) and days to heading date (C) alleles in 410 Korean wheats, and frequency of culm length (D), spike length (E) and days to heading date (F) alleles in 111 Korean experimental lines according to the year released. Bars indicates ranges of Korean wheats.
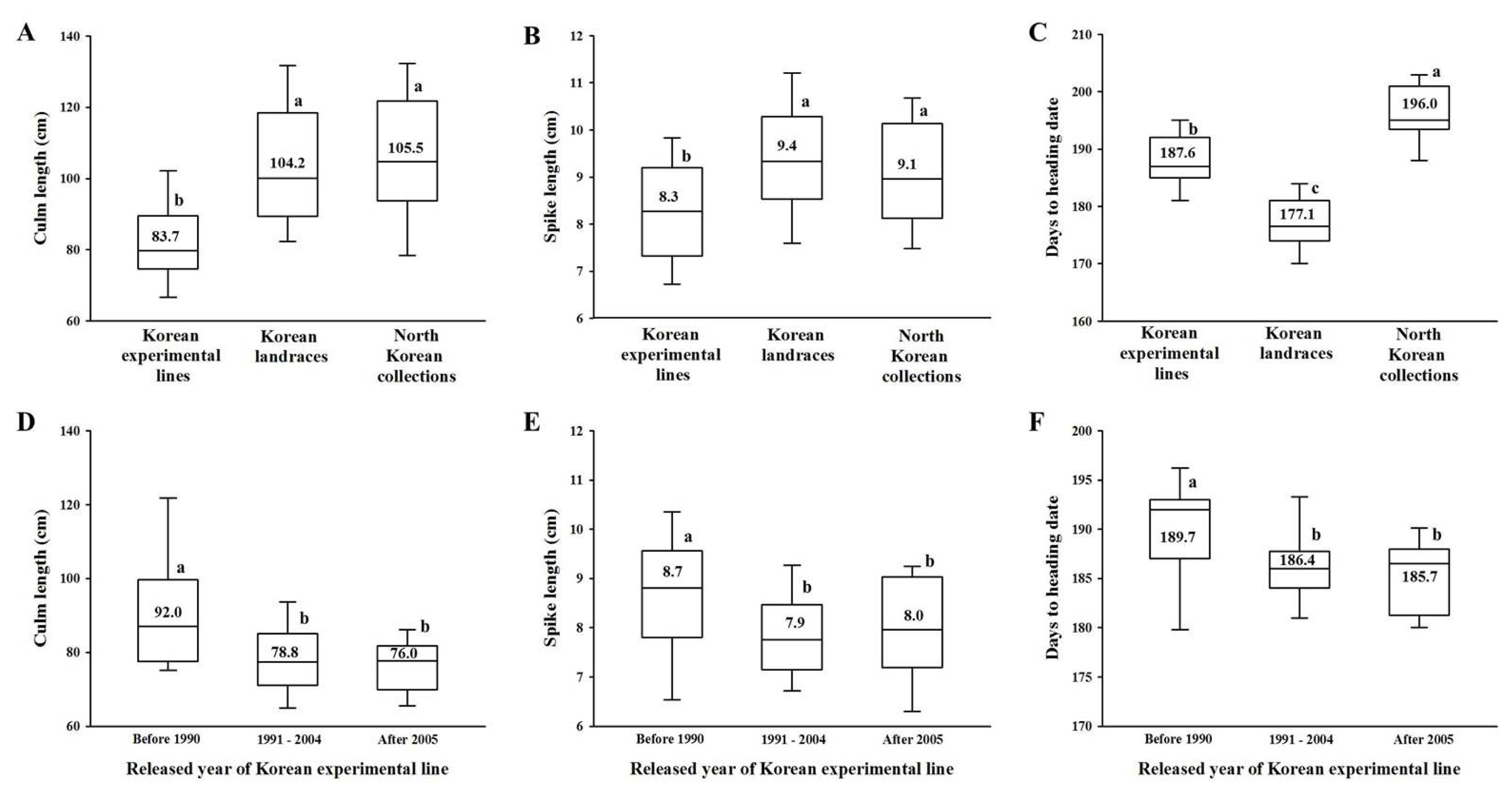
Table 1Differences in culm length, spike length and days to heading date in Korean wheats according to different alleles at the Rht-1, Vrn-1 and Ppd-1 loci.
Table 1
|
No. line |
Culm length (Cm) |
Spike length (Cm) |
Days to heading date |
|
Rht-B1a
|
318 |
101.4az)
|
9.2a |
182.1a |
|
Rht-B1b
|
92 |
90.0b |
8.7b |
185.1a |
|
Rht-D1a
|
322 |
102.3a |
9.1a |
183.3a |
|
Rht-D1b
|
88 |
86.3b |
9.0a |
180.7a |
|
Rht-B1a Rht-D1a
|
233 |
107.3a |
9.2ab |
182.7b |
|
Rht-B1a Rht-D1b
|
85 |
87.5ab |
8.9ab |
180.4b |
|
Rht-B1b Rht-D1a
|
90 |
90.2ab |
8.7b |
184.9b |
|
Rht-B1b Rht-D1b
|
2 |
82.0b |
10.5a |
194.5a |
|
vrn-B1
|
385 |
99.4a |
9.1a |
182.7b |
|
Vrn-B1a
|
23 |
87.4a |
8.3a |
183.1b |
|
Vrn-B1b
|
2 |
89.0a |
9.0a |
193.5a |
|
|
vrn-D1
|
271 |
100.8a |
9.3a |
183.9a |
|
Vrn-D1a
|
139 |
95.0b |
8.7b |
180.5b |
|
vrn-B1 vrn-D1
|
250 |
102.0a |
9.3a |
184.0a |
|
vrn-B1 Vrn-D1a
|
135 |
95.1a |
8.7a |
180.2a |
|
Vrn-B1a vrn-D1
|
20 |
87.9a |
8.3a |
182.2a |
|
Vrn-B1a Vrn-D1a
|
3 |
84.7a |
8.7a |
189.7a |
|
Vrn-B1b vrn-D1
|
1 |
74.0a |
8.0a |
194.0a |
|
Vrn-B1b Vrn-D1a
|
1 |
104.2a |
10.0a |
193.0a |
|
|
Ppd-B1a
|
25 |
81.2b |
8.2b |
183.6a |
|
Ppd-B1b
|
385 |
100.0a |
9.1a |
182.7a |
|
Ppd-D1a
|
308 |
93.5b |
8.8b |
182.6a |
|
Ppd-D1b
|
102 |
114.8a |
9.7a |
183.5a |
|
Ppd-B1a Ppd-D1a
|
25 |
81.2c |
8.3c |
183.6a |
|
Ppd-B1b Ppd-D1a
|
283 |
94.6b |
8.9b |
182.5a |
|
Ppd-B1b Ppd-D1b
|
102 |
114.8a |
9.7a |
183.5a |
References
- Beales J, Turner A, Griffiths S, Snape JW, Laurie DA. 2007. A pseudo-response regulator is mis-expressed in photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum). Theor Appl Genet. 115: 721-733.
- Bentley AR, Turner AS, Gosman N, Leigh FJ, Maccaferri M, Dreisigacker S, Greenland A, Laurie DA. 2011. Frequency of photoperiod-insensitive Ppd-A1a alleles in tetraploid, hexaploid and synthetic hexaploid wheat germplasm. Plant Breed. 130: 10-15.
- Börner A, Plaschke J, Korzun V, Worland AJ. 1996. The relationships between the dwarfing genes of wheat and rye. Euphytica. 89: 69-75.
- Börner A, Worland AJ, Plaschke J, Schumann E, Law CN. 1993. Pleiotrophic effects of genes for reduced (Rht) and day-length insensitivity (Ppd) on yield and its components for wheat growing middle Europe. Plant Breed. 111: 204-216.
- Chen F, Gao M, Zhang J, Zuo A, Shang X, Cui D. 2013. Molecular characterization of vernalization response genes in bread wheat from the Yellow and Huai Valley of China. BMC Plant Biol. 13: 199
- Cho EJ, Kang CS, Yoon YM, Park CS. 2015a. The effects of Rht semi-dwarfing alleles on agronomic traits in Korean wheat cultivars. J Agric Sci Tech. Submitted.
- Cho EJ, Kang CS, Yoon YM, Park CS. 2015b. The relationship between allelic variations of Vrn-1 and Ppd-1 and agronomic traits in Korean wheat cultivars. Indian J Genet Plant Breed. Accepted.
- Dyck JA, Matus-Cádiz MA, Hucl P, Talbert L, Hunt T, Dubuc JP, Nass H, Clayton G, Dobb J, Quick J. 2004. Agronomic performance of hard red spring wheat isolines sensitive and insensitive to photoperiod. Crop Sci. 44: 1976-1981.
- Ellis MH, Spielmeyer W, Gale KR, Rebetzke GJ, Richards RA. 2002. “Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theor Appl Genet. 105: 1038-1042.
- Flintham E, Borner A, Worland A, Gale M. 1997. Optimizing wheat grain yield effects of Rht (gibberellin-insensitive) dwarfing genes. J Agric Sci. 128: 11-25.
- Foulkes MJ, Sylvester-Bradley R, Worland AJ, Snape JW. 2004. Effect of a photoperiod response gene Ppd-D1 on yield potential and drought resistance in UK winter wheat. Euphytica. 135: 63-73.
- Fu D, Szűcs P, Yan L, Helguera M, Skinner JS, von Zitzewitz J, Hays PM, Dubcovsky J. 2005. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol Gen Genom. 273: 54-65.
- Goncharov NP. 2004. Response to vernalization in wheat: its quantitative or qualitative nature. Cereal ResComm. 32: 323-330.
- Iqbal M, Navabi A, Yang R-C, Salmon DF, Spaner D. 2007. Molecular characterization of vernalization response genes in Canadian spring wheat. Genome. 50: 511-516.
- Kamran A, Iqbal M, Spaner D. 2014. Flowering time in wheat (Triticum aestivum L.): a key factor for global adaptability. Euphytica. 197: 1-26.
- Knopf C, Becker H, Ebmeyer E, Korzun V. 2008. Occurrence of three dwarfing Rht genes in German winter wheat varieties. Cereal ResComm. 36: 553-560.
- Korzun V, Röder MS, Ganal MW, Worland AZ, Law CN. 1998. Genetic analysis of the dwarfing gene (Rht8) in wheat. Part I. molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.). Theor Appl Genet. 96: 1104-1109.
- MAF.2014. Food, agriculture, forestry and fisheries statistical yearbook.
- Santra DK, Santra M, Allan RE, Campbell KG, Kidwell KK. 2009. Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1, and Vrn-D1 in spring wheat germplasm from the Pacific Northwest Region of the U.S.A. Plant Breeding. 128: 576-584.
- Shcherban AB, Emtseva MV, Efremova TT. 2012. Molecular genetical characterization of vernalization genes Vrn-A1, Vrn-B1 and Vrn-D1 in spring wheat germplasm from Russia and adjacent regions. Cereal ResComm. 40: 351-361.
- Singh SK, Singh AM, Jain N, Singh GP, Ahlawat AK, Ravi I. 2013. Molecular characterization of vernalization and photoperiod genes in wheat varieties from different agroclimatic zones of India. Cereal ResComm. 41: 376-387.
- Snape JW, Butterworth K, Whitechurch E, Worland AJ. 2001. Waiting for fine times: genetics of flowering time in wheat. Euphytica. 119: 185-190.
- Stelmakh AF. 1998. Genetic systems regulating flowering response in wheat. Euphytica. 100: 359-369.
- Tosovic-Maric B, Kobiljski B, Obrehti D, Vapa L. 2008. Evaluation of wheat Rht genes using molecukar markers. Genetika. 40: 31-38.
- Worland AJ, Law CN. 1986. Genetic-analysis of chromosome 2D of wheat .1. the location of genes affecting height, day-length insensitivity, hybrid dwarfism and yellow-rust resistance. J Plant Breed. 96: 331-345.
- Worland AJ, Appendino ML, Sayers EJ. 1994. The distribution, in European winter wheats, of genes that influence ecoclimatic adaptability whist determining photoperiodic insensitivity and plant height. Euphytica. 80: 219-228.
- Worland AJ, Börner A, Korzun V, Li WM, Petrovíc S, Sayers EJ. 1998. The influence of photoperiod genes on the adaptability of European winter wheats. Euphytica. 100: 385-394.
- Worland AJ. 1996. The influence of flowering time genes on environmental adaptability in European wheats. Euphytica. 89: 49-57.
- Yamada T. 1990. Classification of GA response, Rht genes and culm length in Japanese varieties and landraces of wheat. Euphytica. 50: 221-239.
- Yan L, Helguera M, Kato K, Fukuyama S, Sherman J, Dubcovsky J. 2004. Allelic variation at the VRN-1 promoter region in polyploidy wheat. Theor Appl Genet. 109: 1677-1686.
- Yang FP, Zhang XK, Xia XC, Laurie DA, Yang WX, He ZH. 2009. Distribution of photoperiod insensitive Ppd-D1a allele in Chinese wheat cultivars. Euphytica. 165: 445-452.
- Zhang XK, Xiao YG, Zhang Y, Xia XC, Dubcovsky J, He ZH. 2008. Allelic variation at the vernalization genes Vrn-A1, Vrn-B1, Vrn-D1, and Vrn-B3 in Chinese wheat cultivars and their association with growth habit. Crop Sci. 48: 458-470.
- Zhang J, Wang Y, Wu S, Yang J, Liu H, Zhou Y. 2012. A single nucleotide polymorphism at the Vrn-D1 promoter region in common wheat is associated with vernalization response. Theor Appl Genet. 125: 1697-1704.
- Zhang X, Yang S, Zhou Y, He Z, Xia X. 2006. Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers. Euphytica. 152: 109-116.