Abstract
DNA methylation is a common and rapid evolving epigeneticmechanism among higher eukaryotic organisms including cytosine residues in CG and CNG sequences. NtROS2a gene isolated from tobacco plants (Nicotiana tabacum cv Xanthi nc) contains DNA glycosylase domain with significant similarities to Arabidopsis ROS1 and DME at the C-terminus. Purified NtROS2a protein gene is 5879 bp long and is designated as Nicotiana tabacum Repressor Of Silencing (NtROS: accession numbers; AB281588). RT-PCR analysis initially revealed the adaptive response of the lines overexpressing the gene in tobacco to various abiotic stresses. This was validated when phenotypic evaluation to cold, salt, simulated drought and osmotic stress tolerance was performed. Reduction in leaf wilting rate against high salinity treatment is remarkable in transgenic plants which showed at least 50% less wilting than the WT. Moreover, about 70% wilt rate difference was noted between the transgenic and WT in H2O2 treatment. This study showed that NtROS2a gene encoding DNA glycosylase which caused demethylation could regulate adaptive response of tobacco to various abiotic stress conditions.
-
Key words: DNA methylation, Demethylation, Repressor of silencing, Nicotiana tabacum, Transgenic plant
INTRODUCTION
Epigenetics is a change in gene expression through chromatin modifications without base sequence alteration (
Law and Jacobsen 2010), which facilitates recruitment of the RNA polymerase II transcriptional machinery to the promoter. This frequently occurs during somatic cell differentiation, in which the clonal expansion of a single cell leads to a diversity of cell types (
Zhang et al. 2006). Such a cellular inheritance is common during ontogeny but is usually erased before gametes are produced (
Reik and Walter 2001). In eukaryotes, DNA methylation has important roles in the establishment of developmental programs in fungi, plants, and animals. DNA methylation of mammalian and plant is restricted to symmetrical CpG sequences, but plants also have significant levels of cytosine methylation in the symmetric context CpNpG (where N is any nucleotide), CpNpN (N is A, T or C) and even in a symmetric contexts (
Bird 2002). DNA methylation is associated with numerous functions, depending on the model organism and the environment component. In the experiment involving mice, DNA methylation has been shown to be essential for proper embryonic development (
Li et al. 1992). Similarly, a loss of DNA methylation caused developmental defects in
Xenopus embryos (
Stancheva and Meehan 2000). In parallel with plants, reduced levels of DNA methylation were shown to be responsible for a large number of developmental abnormalities (
Finnegan et al. 1996;
Ronemus et al. 1996;
Cao and Jacobsen 2002;
Kankel et al. 2003). The triple mutant that lacks DRM1, DRM2, and CMT3, which has greatly reduced non-CG methylation (
Cao and Jacobsen 2002) and the mutant that lacks MET1, in which both CG and, to a lesser extent, non-CG methylation are decreased. Superficial similarities in some of the developmental defects have been interpreted to reflect a conserved function of DNA methylation. However, the molecular consequences of genomic DNA methylation have been found to be surprisingly diverse and broad (
Cao and Jacobsen 2002;
Kankel et al. 2003). However, unlike methylation, studies on de-methylation are still limited. In contrast to the well-studied genetics, biology and biochemistry of cytosine DNA methyltransferases, the enzymatic basis of active demethylation have remained elusive (
Morales-Ruiz et al. 2006). Direct removal of the methyl group from 5-meC residues has been reported, but this mechanism, which involves a thermodynamically unfavorable breakage of the carbon–carbon bond, has been questioned and could not be independently reproduced (
Agius et al. 2006). Recent work in plants has added genetic evidence for a role of DNA glycosylases in an active demethylation pathway. In Arabidopsis, recent genetic and biochemical studies demonstrated that two bifunctional DNA glycosylase/lyases, ROS1 and Demeter, function as DNA demethylase (
Gehring et al. 2006,
Agius et al. 2006,
Morales-Ruiz et al. 2006). ROS1 can specifically recognize methylated DNA substrate. Its glycosylase activity removes the 5-methylcytosine base, and its lyase activity nicks the DNA backbone at the abasic site by α, β elimination mechanism (
Agius et al. 2006,
Morales-Ruiz et al. 2006). Methylation status dynamically changes depending upon cell type, developmental stages and even environmental conditions (
Demeulemeester et al. 1999;
Steward et al. 2002;
Chakrabarty et al. 2003;
Wada et al. 2004). First, because DNA methylation in nucleosomes induces alteration of gene expression by changing chromatin structures; vast demethylation may serve as a common switch for many genes that are simultaneously controlled upon environmental cues. Second, because artificial demethylation induces heritable changes in plant phenotype (
Sano et al. 1990); altered DNA methylation may result in epigenetic inheritance, in which gene expression is modified without changing the nucleotide sequence. To investigate the function of
NtROS2a gene encoding DNA glycosylase in various abiotic stress conditions, we developed transgenic tobacco plants using overexpression and RNAi technique and subjected them to cold, salt and heat stress.
MATERIALS AND METHODS
Gene isolation and cloning
Total RNA was isolated from tobacco samples by acid guanidium thiocyanate-phenol -chloroform (AGPC) method (
Chomczynski and Sacchi 1987) with a slight modification, and used for cDNA library construction with the ZapII vector (Stratagene, La Jolla, CA, USA). In brief, cDNAs were ligated to the vector at the
EcoRI and
XhoI sites, and after transformation, the library was screened with 32P- labeled probes. Positive clones were rescued in the pBluescript SK-phagemid vector by
in vivo excision, and amplified in
Escherichia coli DH5α. The nucleotide sequences were determined by the dideoxynucleotide chain termination method (PRISM BigDye Terminator, ABI, Sunnyvale, CA, USA), and analyzed with appropriate computer software (GeneWorks, National Center for Biotechnology Information, and PSORT server). Genomic sequence was obtained by PCR using genomic DNA as the template and appropriate primer sets designed after the cDNA sequence. Resulting fragment was 10,243 bp, including a 1918 bp upstream region from the transcription initiation site (accession number AB267678).
To produce transgenic plants carrying NtROS2a gene, the NtROS2a cDNA was ligated into the pBI121 and pKANNIBAL vector and electroplated into Escherichia coli strain XL10-Gold (Stratagene) resulting in 1.5 × 106 transformants. The construct was confirmed by PCR amplification using the 35s promoter (F-5′ TGAGACTTTTCAACAAAGGGTA), and NtROS2a (R-3′ GGGAACTGATGAGCGGTTTG) primers. The construct was electroporated into Agrobacterium tumefaciens strain LBA4404 (Koncz and Schell 1986) and 2.6 × 106 cfu/ml inoculum was obtained 48 hours after shake-incubation of the bacteria in 25 ml LB medium at 240 rpm speed at 28ºC and stored in 50% glycerol at −80ºC until use.
Generation of NtROS2a transgenic tobacco
Seeds of wild type tobacco were surface sterilized by washing with 70% ethanol for 1 min, 3% sodium hypochlorite for 10 min, and rinsed four times with sterile distilled water. The seeds were then allowed to germinate in glass containers with 25 cm
3 of MS-based medium (
Murashige and Skoog 1962). For transformation experiments, tobacco was grown on MS medium in a Magenta culture box (Sigma, USA) in a culture room at 28ºC under an 18 h (day)/6 h (night) photoperiod. Sterile leaf disks were co-cultivated with
Agrobacterium and planted onto a medium without antibiotics. After incubation at 28ºC for two days, the leaf discs were transferred to MS medium supplemented with 6-benzylaminopurine (BA) 1 mg/L and 1-naphtalene acetic acid (NAA) 0.1 mg/L for callus and shoot induction. For root induction, MS medium containing hygromycin 50 mg/L was used. The media for callus and shoot, and root induction contained 8 g/L of agar, 50 mg/L hygromycin and the cultures were maintained at 28ºC under light condition. After root induction, putative transgenic plants were transferred immediately to MS medium without plant hormones.
Total DNA of transgenic tobacco was extracted from a ground freeze-dried leaf tissue (200 mg) using the cetyltrimethy1 ammonium bromide (CTAB) method. After adding the CTAB buffer, the samples were incubated in a 55ºC water bath for 15 min. After incubation, the extract mixture was spun at 1200 rpm for 5 min and the supernatant were transferred into clean tubes and added with 250 μl of chloroform: isoamy1 alcohol (25:1). The mixture was mixed gently by inverting for 5 min and was centrifuge at 13,000 rpm for 3 min at room temperature. The aqueous phase were collected into a clean tube added with 0.08 vol. of cold 7.5 M ammonium acetate and 0.8 vol. of cold isopropanol (−20ºC). The mixture was kept in the freezer for 30 min, and the DNA was collected by centrifugation at 13,000 rpm for 10 min. The pellet was washed with 500 μl 70% ethanol, air-dried and resuspended in 50–100 μl of ddH2O.
PCR analysis
The presence of transgene in putative transgenic plants was detected by PCR method with 35s promoter F-5′ (5′-TGAGACTTTTCAACAAAGGGTA-3′) and NtROS2a gene R-(5′-GGGAACTGATGAGCGGTTTG-3′) primer pair. The assay was performed in a total 50 μl volume containing 100 pg DNA, 10 pmol each of primers, 2.5mM dNTPs, 10X PCR buffer, and 0.5 U Taq-polymerase (Takara, Japan). The PCR reaction was carried out by denaturing the template at 94ºC for 5 min followed by 35 cycles of amplification (1 min at 94ºC, 1 min at 50ºC and 2 min at 72ºC) and by extension at 72ºC for 5 min.
Gene expression analysis under the abiotic stresses
Tobacco Seeds (
Nicotiana tabacum cv Xanthinc) were grown under controlled conditions (28ºC day/25ºC night temperatures, 14 h light/10 h darkness, 75% relative humidity) in MS medium in a Magenta culture box (Sigma, USA). After three weeks of germination, seedlings were exposed to high-salinity (250 mM), cold (4ºC), H
2O
2 (3%), and 100 μM ABA stresses. The plant tissues were harvested after 0, 30 min, 6h, and 8h of stress treatment, immediately frozen in liquid nitrogen, and stored at −80ºC. Total RNA was extracted using Trizol reagent (Invitrogen, USA) according to the manufacturer’s protocol. Real-time PCR was conducted using RT pre-mix (TOYOBO Co., Japan) in a Bio-RAD I Cycler IQ5 machine. The threshold cycle (Ct) values of the triplicate PCRs were averaged and the relative quantification of the expression levels was performed using the comparative Ct method (
Livak et al. 2001). The fold change in total RNA (target gene) relative to the reference gene (actin gene) was determined by the following formula: fold change = 2
−ΔΔCt, where ΔΔCt = (Ct target gene – Ct actin gene) transgenic plants (Ct target gene – Ct actin gene) wild-type plants. The following primers were used for the analysis,
NtROS2a: (5′-CACTT-TTGGAAGGGTTGGACA-3′) and R (5′-CGTATGCTGTTGCATGAGGAA-3′), and Actin F: (5′-CGCCCGTCATCGTGCGGTTC-3′) and R: (5′-TCCGCGGGGTCCATGG TCTG-3′). RT-PCR analysis of vegetative tissue (leaf, stem, and root) was conducted using 50 ng of total RNA treated with DNase I. The amplification program consisted of an initial step at 42ºC for 60 min, 94ºC for 5 min, 40 cycles (94ºC for 15 sec, 55ºC for 30 sec, 72ºC for 30 sec), and a final step at 72ºC for 10 min, and the amplified fragments were loaded on 1.2% agarose gel.
All seeds in the following assays were surface sterilized and kept at 4ºC for 2 d to break dormancy before subjecting them to any of the experimental treatments. For the germination assay, the tobacco seeds were plated in one-half MS medium (0.6% agar) added separately with PEG6000 or NaCl, and the number of germinated seeds was counted every day. For root elongation assay, the germinated seeds were first plated upright in MS plates until the roots were about 0.5 cm, then the seedlings were transferred into the same medium supplemented with PEG6000 or NaCl, in the growth chamber. The root lengths were recorded before and after the treatments.
Leaf discs assay and chlorophyll contents in transgenic plants
Tobacco seeds (
Nicotiana tabacum cv Xanthi nc) were grown under controlled conditions having 28ºC day/25ºC night temperatures, 14 h (light)/10 h (darkness) in MS medium in a Magenta culture box (Sigma, USA), and 75% relative humidity. Three weeks after germination, healthy and fully expanded leaves of wild-type and transgenic plants were detached. Leaf discs of about 1 cm in diameter were cut and floated in 1/2 MS medium containing separately either 250 mM NaCl or 3% H
2O
2 for 6 d. The treatment was performed in continuous white light at 25ºC. To examine the chlorophyll contents, the leaf discs obtained from transgenic and WT plants were floated in 1/2 MS liquid medium containing 250 mM NaCl, 3% H
2O
2 without sucrose (MSH) for 7 d under light at 25ºC. Measurement of chlorophyll fluorescence was performed according to
Aono et al. (1993).
RESULTS
Characterization of NtROS2a genes
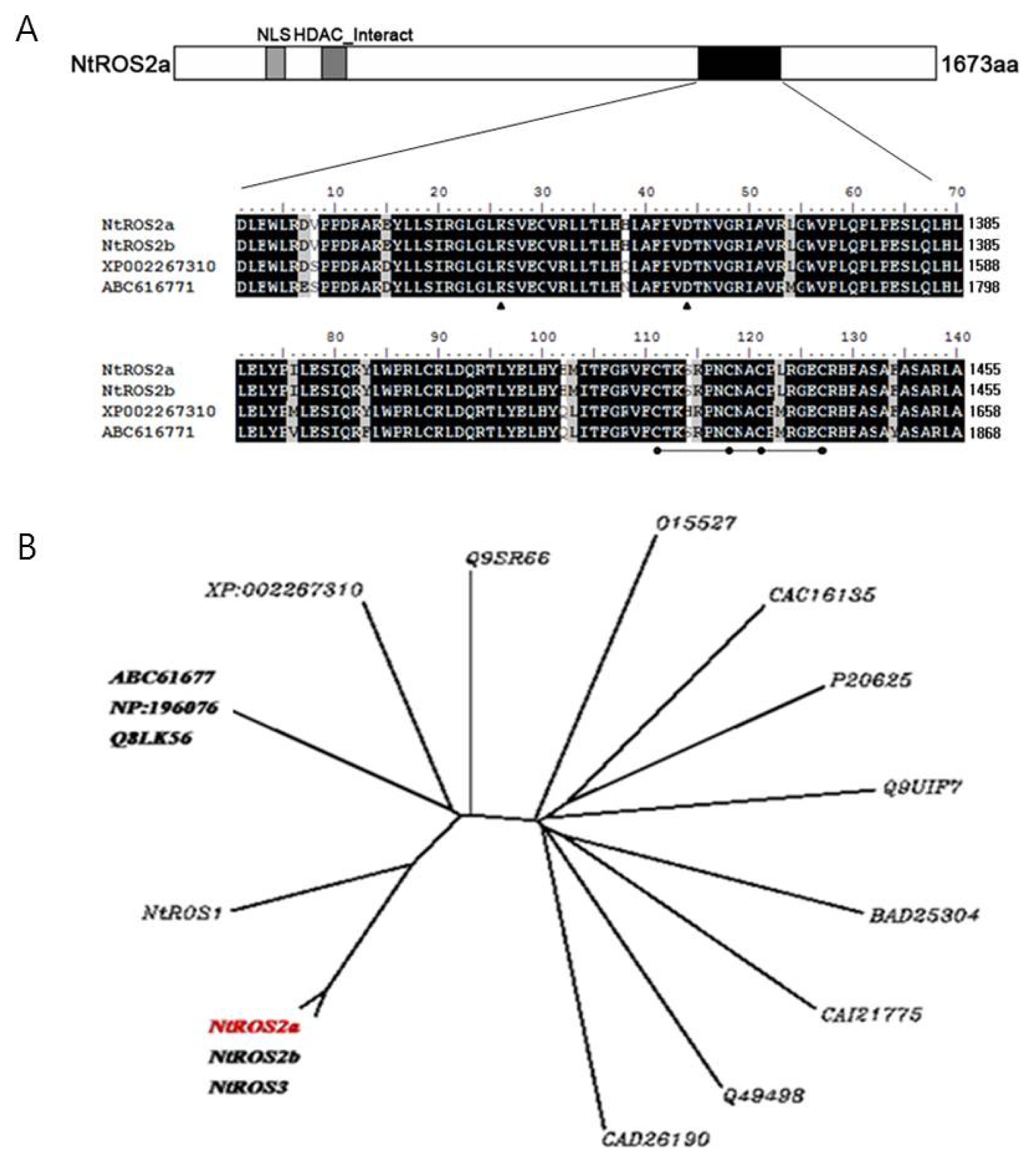
Two cDNA fragments were initially isolated from tobacco leaves RNA by reverse transcriptase-PCR using a set of degenerate primers which were designed based on the conserved DNA glycosylase domain being known as plant 5-methylcytosine DNA glycosylase proteins (
http://www.chromdb.org). One of the two fragments cloned, designated as
Nicotiana tabacum Repressor Of Silencing 2a (
NtROS2a), (
Fig. 1A) (accession numbers AB281588 for
NtROS2a) is 5879bp long and is highly associated to XP_002267310 (
Vitis vinifera) and ABC61677 (
A. thaliana). This clone encoded a protein of 1,673 amino acids with predicted molecular mass of 186 kDa. A pair-wise comparison of their deduced amino acid sequences with ABC61677 (
A. thaliana) showed the similarity of
NtROS2a at 32.2%. However, the similarity at the glycosylase domain with ABC61677 (
A. thaliana) was 94.3% for
NtROS2a (amino acid positions, 1,291–1,459). A pair-wise comparison of
NtROS2a indicated the similarity of
NtROS2b to be 99.6%. Notably, only five out of 1,673 total amino acids (0.4%) differed between
NtROS2a and
NtROS2b, suggesting them to be isoforms. Clustering starts in the root node with the training and evaluation of 17 DNA glycosylase classifiers (
Fig. 1B). The species representing the DNA glycosylase group are divided into two groups under the same parent roots.
NtROS2a gene is grouped to other roots compared with genes related to demethylation. This result indicates that
NtROS2a is a unique gene related to demethylation. GFP fluorescence in BY-2 cell suspensions was examined after transformation. About 70% of round- shaped calli that were obtained after the
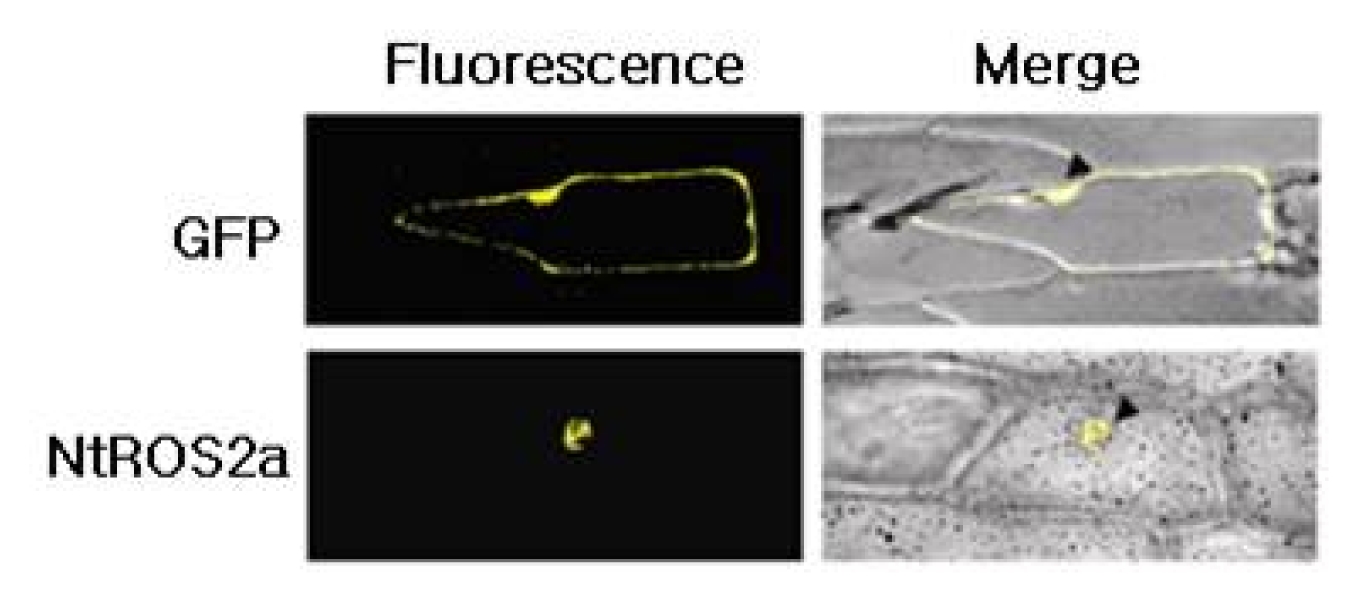
Agrobacterium- mediated transformation of BY-2 cells displayed GFP fluorescence intensity sufficient for reliable evaluation of its homogeneity (data not shown). Subcellular localization of
NtROS2a gene was analyzed by
Agrobacterium- mediated transient expression of a
NtROS2a–GFP fusion gene in onion epidermal cells (
Fig. 2). The cells transformed with
NtROS2a–GFP displayed fluorescence specifically in the nucleus, suggesting that
NtROS2a is a nucleus- localized protein.
In plants,
NtROS2a play a direct regulatory role in response to multiple signal stimulation. To clarify the potential function of
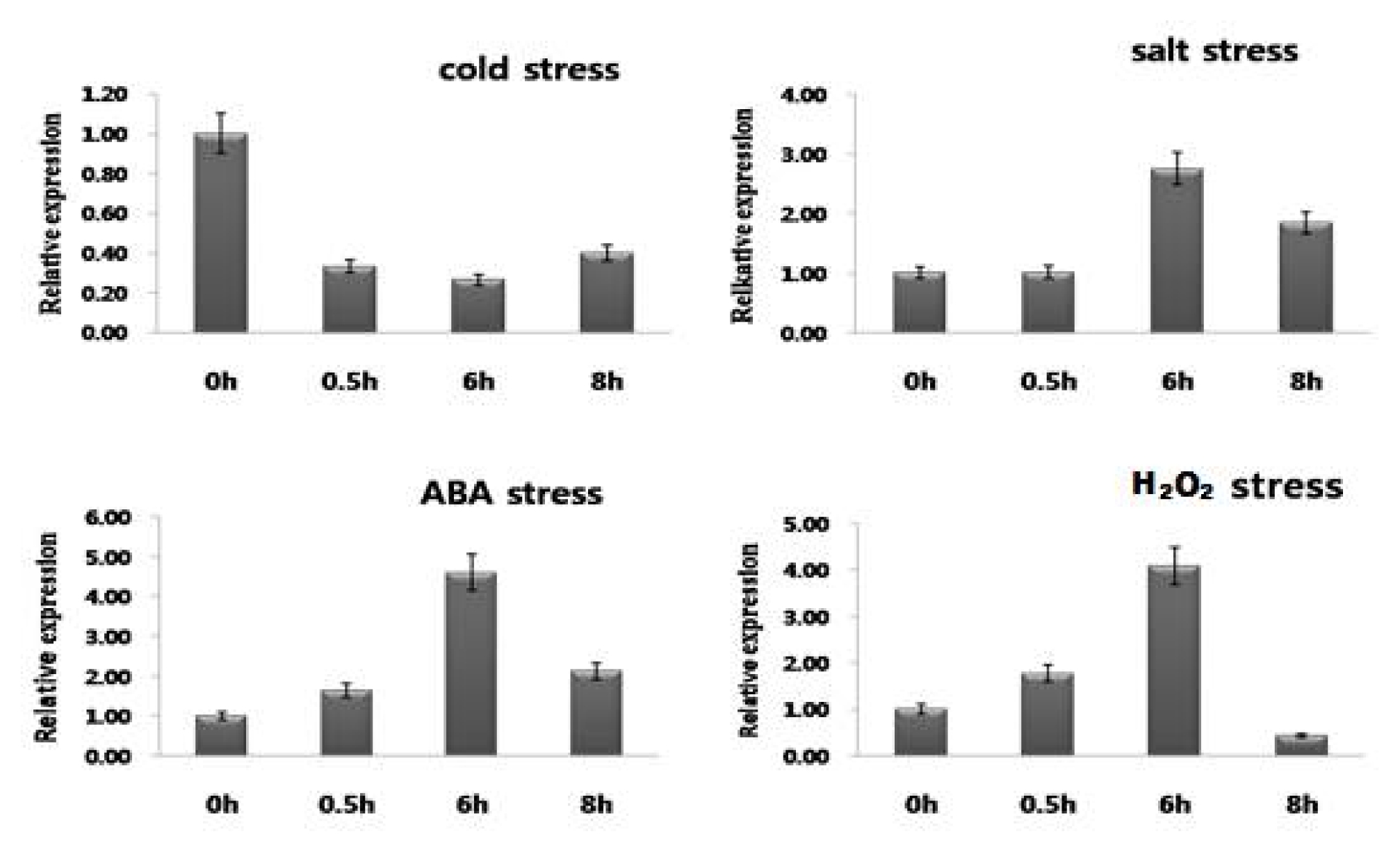
NtROS2a in response to different stimuli, Expression pattern of
NtROS2a gene was investigated under various abiotic stresses (
Fig. 3). The
NtROS2a gene was induced expression after ABA, cold, salt and H
2O
2 treatments.
NtROS2a mRNA was accumulated at 6 hr. after stresses treatment except for cold. At 8 hr. after stresses treatment, the expression level was declined back to the levels at 6 hr. treatment. However, there was no significant accumulation of
NtROS2a mRNA under cold stress. The lowest expression was observed in plants under normal conditions (data not shown).
To generate transgenic plants
, NtROS2a cDNA was cloned into the binary expression vector pBI121 under the control of the 35S promoter (
Fig. 4A). RNAi vector was designed for directional insertion of PCR products on either side of the PDK intron (
Fig. 4B). The
NtROS2a gene fragment was amplified through PCR, appended with the restriction sites and cloned sequentially on either side of the intron to become the two arms of the hairpin. The tobacco leaf discs infected with
Agrobacterium on MS medium with selectable kanamycin showed callus formation. The callus formation initiated from only the putative transformed leaf discs and the untransformed leaf discs began to yellow after 3–5 days. During the following 5–10 days, the leaf discs turned brown and did not initiate callus formation. The transformed leaflets developed into bright green calli and then the shoots emerged with 3 – 5 leaflets in 25–30 days (
Fig. 4C-a, b). The emerging shoots with 3–5 leaflets showed root formation in 20–25 days (
Fig. 4C-c). The roots continued to grow in the putative transgenic plants, but the root growth in untransformed plants did not develop (data not shown). All transgenic plants showed normal phenotype. The T
1 putative transgenic plants were transferred from the 1/2 MS medium to a selectable kanamycin supplemented MS medium. The plantlets remained green and developed elongated roots as they were resistant to kanamycin, while the untransformed plants became yellow while the roots ceased to elongate because of their sensitivity to the antibiotic (
Fig. 4C-d). PCR analysis of transgenic lines in T
2 generation was confirmed integration to tobacco genome both
NtROS2a and nptII gene.
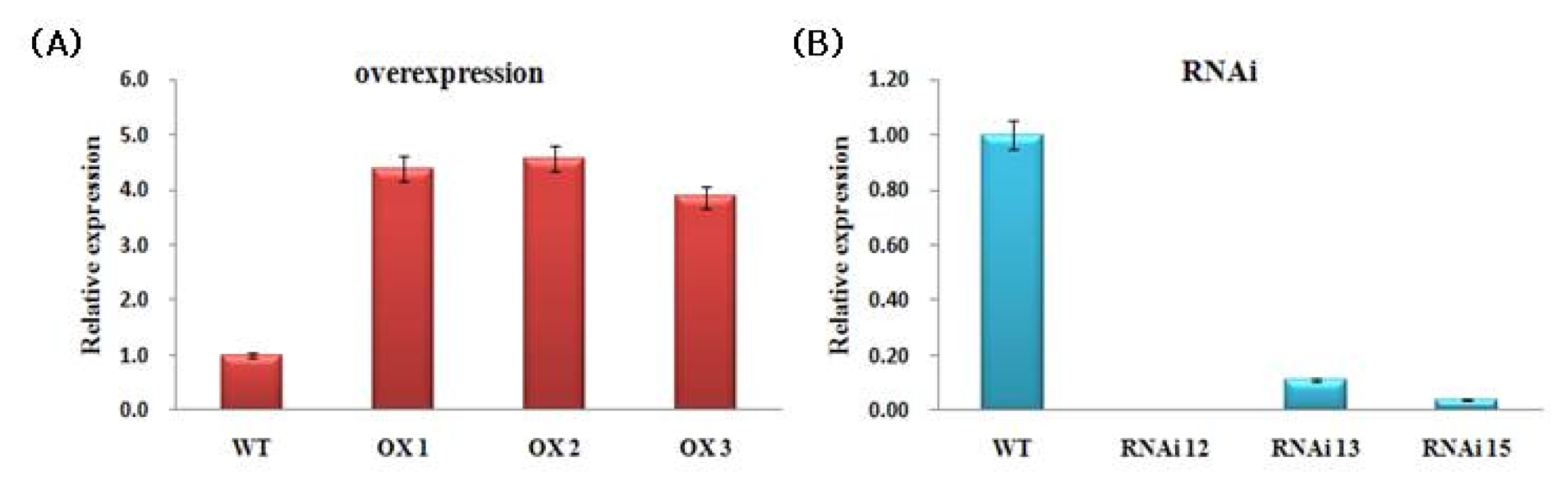
The transcript levels of
NtROS2a in the transgenic plants were determined by RT-PCR analysis using gene-specific primer sets (
Table 1). Total RNAs were extracted from leaf of 14-d-old seedlings grown under normal growth conditions.
NtROS2a expression level was higher in all pBI121::
NtROS2a transgenic lines than in the non-transgenic (WT) controls (
Fig. 5A). By contrast, the pKANNIBAL::
NtROS2a transgenic lines showed lower expression level of
NtROS2a gene than in non-transgenic (
Fig. 5B).
To examine the effect of abiotic stress in transgenic tobacco plants, gene expression patterns were investigated between the transgenic and non-transgenic plants under salt and H
2O
2 stress (
Fig. 6).
NtROS2a transcript was induced by salt- and H
2O
2 stress. The leaves of T
2 NtROS2a transgenic lines OX 1 and RNAi 13 and non-transgenic WT were incubated at 250 mM NaCl and 3% H
2O
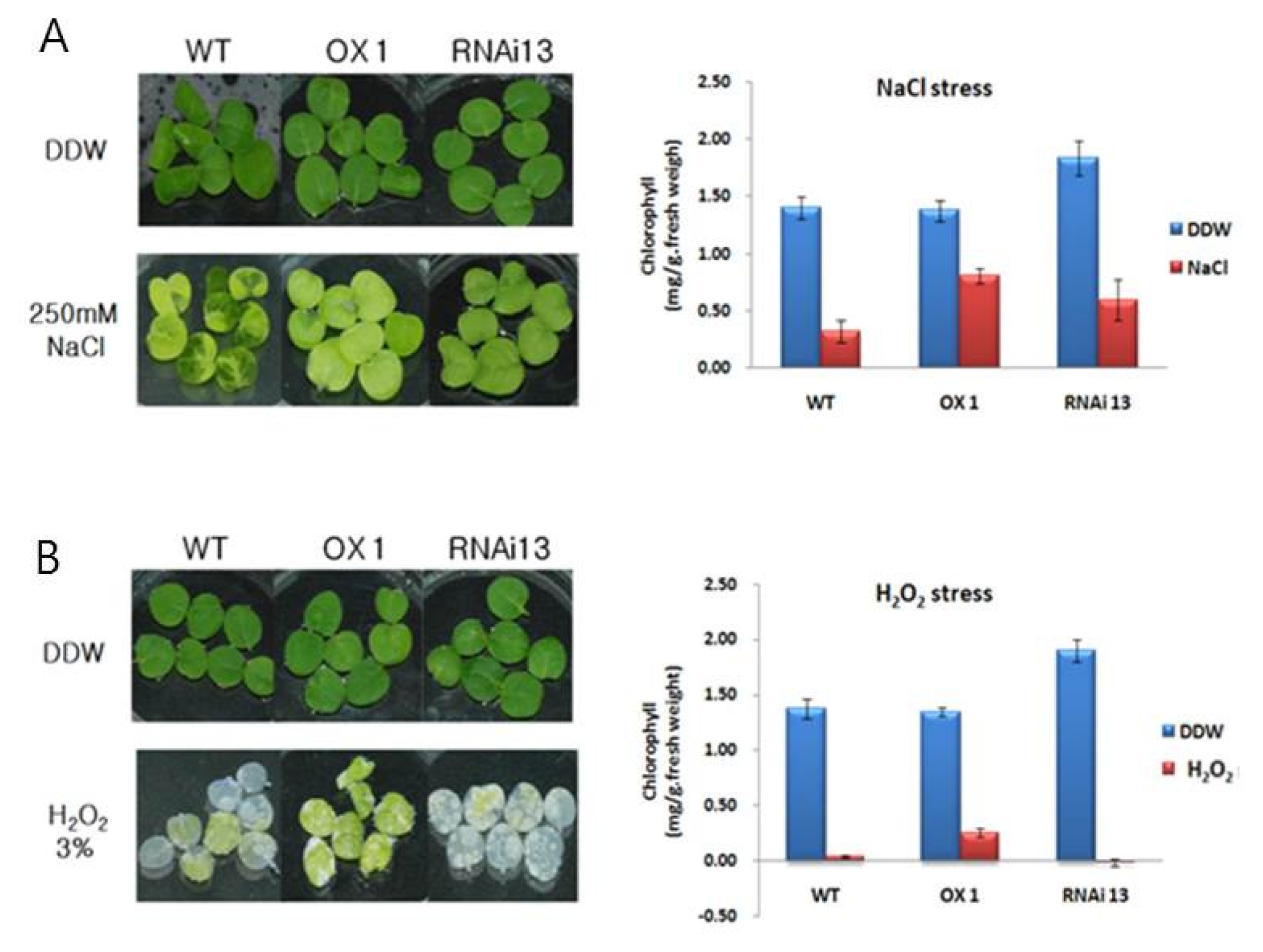
2 for 5 days. In NaCl, most of WT leaves (83%) wilted, whereas only 30.7% seedlings of transgenic line 1 and 24.4% of line 13 wilted (
Fig. 6A). In H
2O
2, most leaves of WT (90%) and transgenic line 13 (99%) were wilted, while only 20% seedlings of transgenic line 1 showed the symptom (
Fig. 6B).
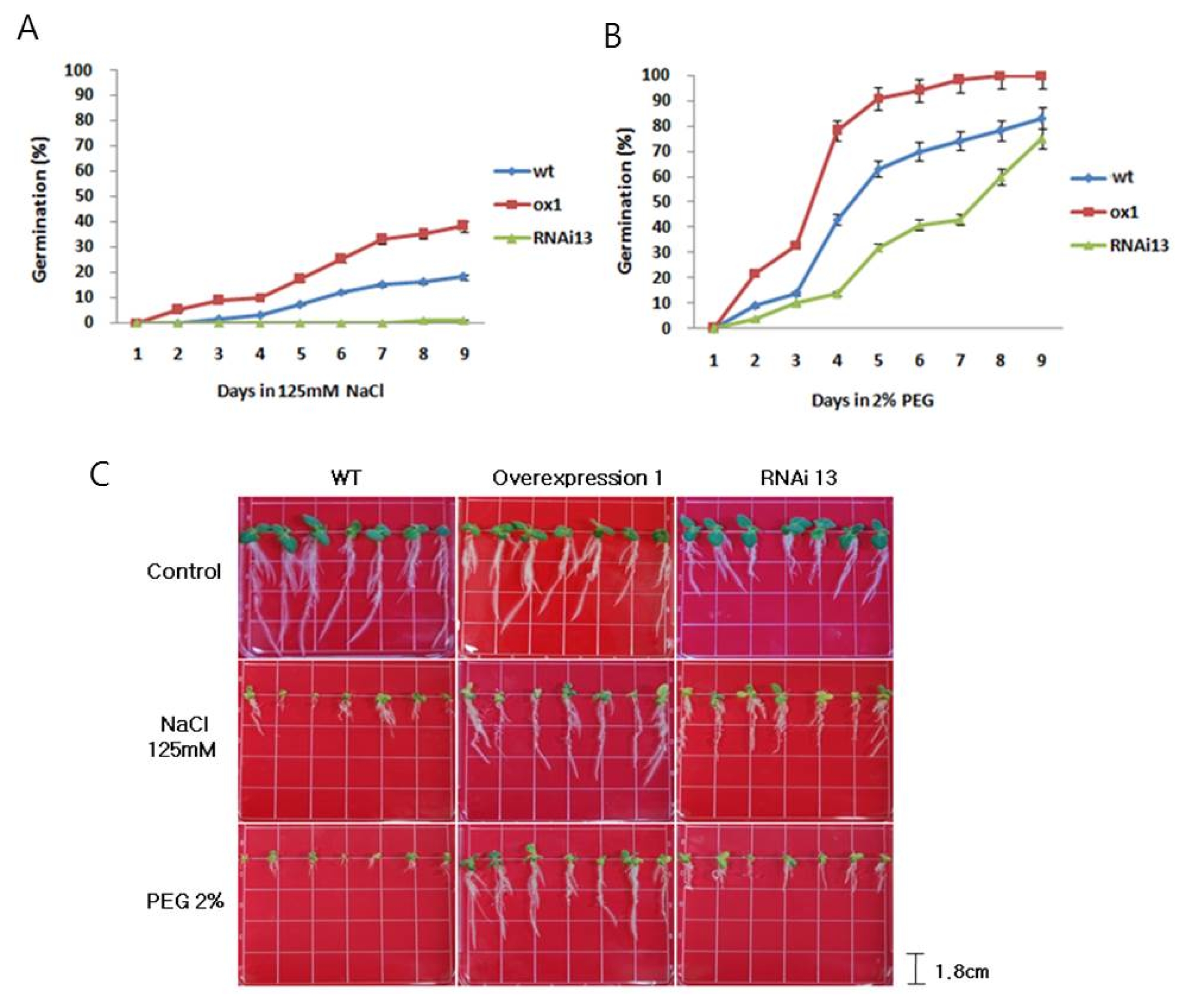
Transgenic lines (OX 1, RNAi 13) were subjected to simulated osmotic stress, and the effects on root elongation were assessed after germination. Germination in OX 1 lines was clearly higher than that in WT and RNAi 13 (
Fig. 7A and B). After 9 d of 125 mM NaCl treatment, seed germination in OX 1 line was 40% which is higher than the 15% germination in WT. Similarly, the germination percentage in OX 1 lines was twice higher than that in WT after 2% PEG6000 treatment for 9 d. After 14 d of the same treatment, the roots of OX 1 line plants were about twice longer than those in wild type and RNAi 13. Interestingly, when subjected to higher concentrations of PEG (2%) for the same duration, roots of OX 1 line were not only about 34% longer than those of wild types but also produced more lateral roots. These results indicate that
NtROS2a increases the adaptation to salt and osmotic stress during seed germination and root elongation in tobacco (
Fig. 7C). Also, the change of DNA methylation affects the gene expression and stability. Insertion of
NtROS2a is expected to cause changes in the genome of recipient plant. Total DNA of transgenic lines was digested with
MspI and
HpaII, which are unable to cleave the sites m
5CCGG and Cm
5CGG, respectively. DNA methylation affects was analyzed by RAPD analysis, but there were no different bands amplified (data not shown). This became part of the band overlap in demethylation, or a very specific band that a fragment is considered.
DISCUSSION
DNA glycosylases are subdivided into four groups according to their substrate specificity: excision of uracil, uracil-containing mismatches, alkylated bases and oxidized bases (
Begum et al. 2007). NtROS proteins are structurally related to
Arabidopsis ROS1 and DME, which belong to a large family of uracil-containing mismatch DNA glycosylases. However, apart from the m
5C DNA glycosylase domain and regions at the carboxyl-terminus, their similarity was low, suggesting specific function of NtROS proteins. Since NtROS1 and NtROS2a were shown to possess m
5C removal activity, and to be localized in nucleus, it is conceivable that they are closely associated with DNA and engaged in DNA demethylation. Conserved lysine and aspartic acid residues located in the helix-hairpin-helix domain in DNA glycosylases (
Krokan et al. 1997) and four cysteine residues adjacent to the DNA glycosylase domain were also shown in NtROS proteins. The latter cluster is considered to play a role in DNA binding. NtROS proteins also possess nuclear localization signal and histone deacetylase interacting domains (
http://smart.embl-heidelberg.de/) (
Fig. 1), suggesting them to localize in nucleus and to interact with DNA or chromatin (
Gehring et al. 2006). In our studies, subcellular localization of
NtROS2a gene was localized a nucleus protein (
Fig. 2). Their induction profile upon abiotic stresses is indicative, as DNA glycosylases are considered to deal with oxidative damages, which are commonly caused by reactive oxygen species (
Choi and Sano 2007;
Begum et al. 2007). The
NtROS2a gene was induced expression after ABA, cold, salt and H
2O
2 treatments.
NtROS2a mRNA was accumulated at 6 hr. after stresses treatment except for cold. At 8 hr. after stresses treatment, the expression level was declined back to the levels at 6 hr. treatment. However, there was no significant accumulation of
NtROS2a mRNA under cold stress (
Fig. 3). To cope with such damages, a set of genes must be expressed to repair and/or strengthen defense reactions. If these genes are locked by methylation under non-stressed condition, demethylation is absolutely necessary to globally switch them on. It is tempting to speculate that NtROS proteins participate in this task, thereby finely adjusting gene expression through methylation/demethylation pathways. This study showed a close correlation between demethylation and expression of a tobacco gene encoding a glycerophosphodiesterase-like protein upon exposure to abiotic stresses. This speculation is in accordance with the present finding, showing that
NtROS2a is distantly related to other members, and localized not only at cell surface (plasma membrane) but also in nucleus, suggesting a novel, so far unknown biological functions. In healthy leaves grown under non-stressed conditions,
NtROD2a was not expressed, and its coding region was found to be heavily methylated. Upon exposure to abiotic stress, its transcripts were induced and genomic locus was partly demethylated, while upon biotic stress, neither transcripts nor demethylation were induced. In this report, transgenic lines (OX 1, RNAi 13) were subjected to simulated osmotic stress, and the effects on root elongation were assessed after germination. Germination in OX 1 lines was clearly higher than that in WT and RNAi 13 (
Fig. 7). After 9 d of 125mM NaCl treatment, seed germination in OX 1 line was 40% which is higher than the 15% germination in WT. This strengthened the close correlation between gene expression and methylation, and shows clear demethylation activity towards m
5C in DNA and ubiquitously expressed in plant tissues (
Aguis et al. 2006). Its involvement in stress response has yet to be confirmed, but the present findings indicate that this possibility warrants attention.
ACKNOWLEDGMENTS
This work was supported by a research grant from Hankyong National University in the year of 2014
Fig. 1(A); Alignment of conserved m5C DNA glycosylase domain of NtROS2a, AB281588; NtROS2b, AB281589 (Nicotian tabaccum); XP_002267310 (Vitis vinifera); ABC61677 (A. thaliana) Common amino acids among for proteins are indicated by closed box, and grey boxes indicate conservative changes. Conserved lysine and aspartic acid residues in the glycosylase/lyase active site (InterPRO search, IPRO11257) are indicated by closed arrowheads, and four cysteine residues predicted to function in DNA binding are indicated by closed circles. (B); Unrooted phylogenetic tree. Amino acid sequences were aligned using the ClustalW program. Genebank accession numbers areNtROS1, AB281587; NtROS2a, AB281588; NtROS2b, AB281589; NtROS3, AB281590 (N. tabacum); XP_002267310 (Vitis vinifera); Nth1, CAC16135; AtDME, Q8LK56; AtDML2, Q9SR66; AtDML3, Q49498: DME, NP_196076; DEMETER, ABC61677 (A. thaliana); MutY, Q9UIF7 Ogg1, O15527; A2, CAI21775 (Homo sapiens); Nth, P20625 (Escherichia coli); A2, BAD25304 (Oryza sativa japonica Group); GB-M1,CAD26190 (Encephalitozoon cuniculi GB-M1 Eukaryota; Fungi; Microsporidia; Unikaryonidae; Encephalitozoon).
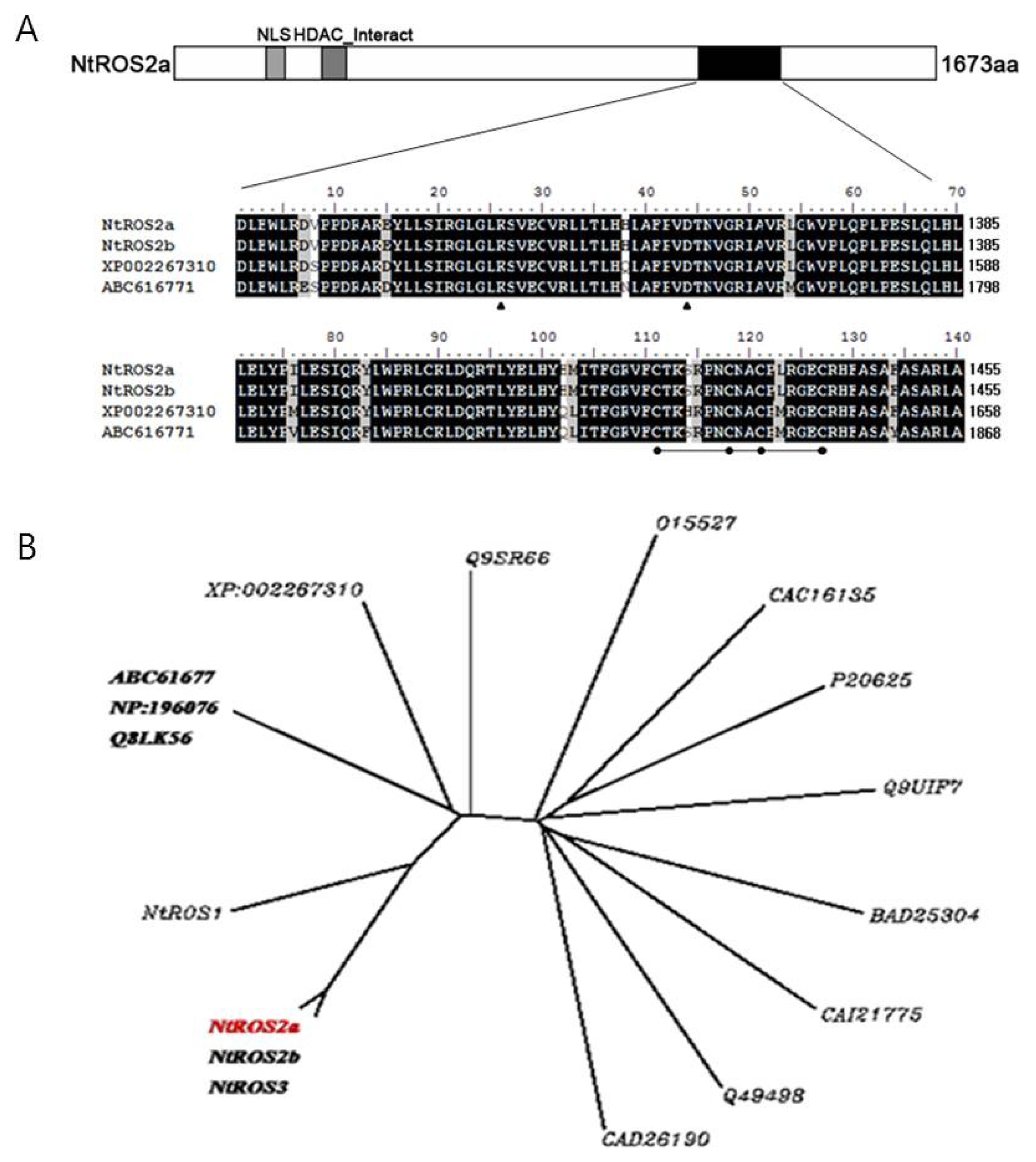
Fig. 2Cellular localization. Onion epidermal cell layers were bombarded with gold particles coated 35SGFP alone or 35SNtROS2a-GFP, and observed by epifluorescence for GFP. Bright field mages are merged to show positions of nucleus indicated by arrowheads.
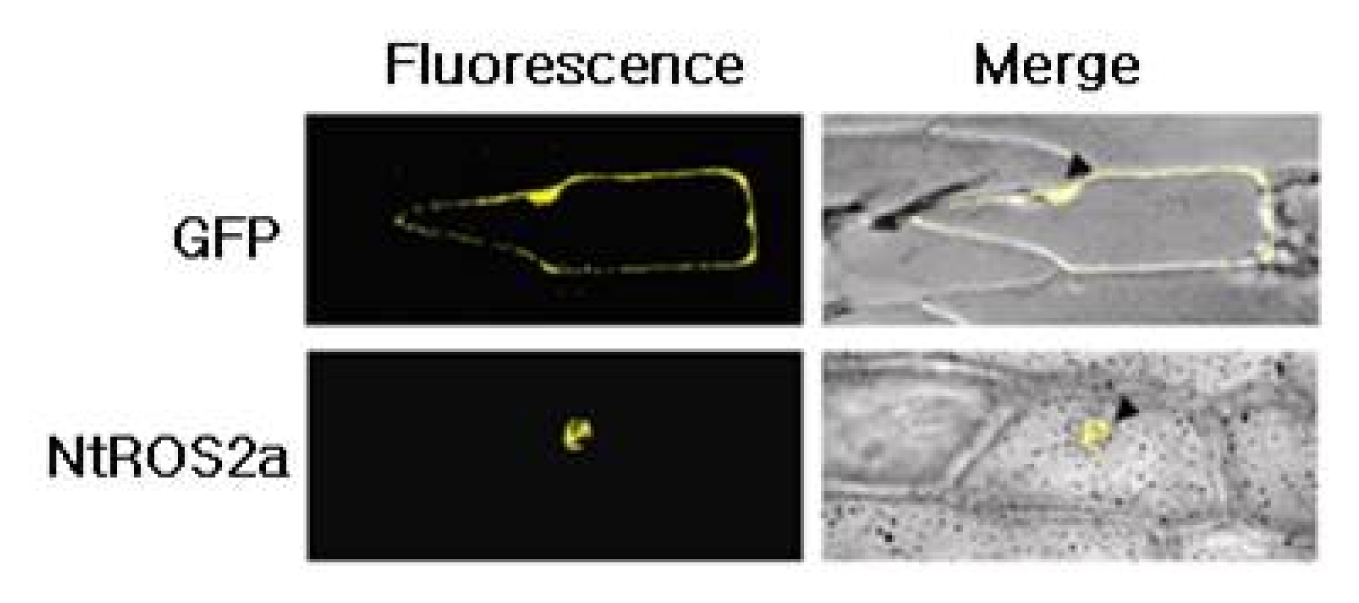
Fig. 3Expression of NtROS2a in reponse to abiotic stress in tobacco. The real-time qRT-PCR analysis of NtROS2a in tobacco seedlings subjected to abiotic stresses in cluding cold (4°C), salt (250 mM NaCl), and ABA (100 μM), H2O2(3%) stress. Total RNA was purified from the tobacco seedlings at Three-leaf stage after various treatments and used in real-time qRT-PCR assays.
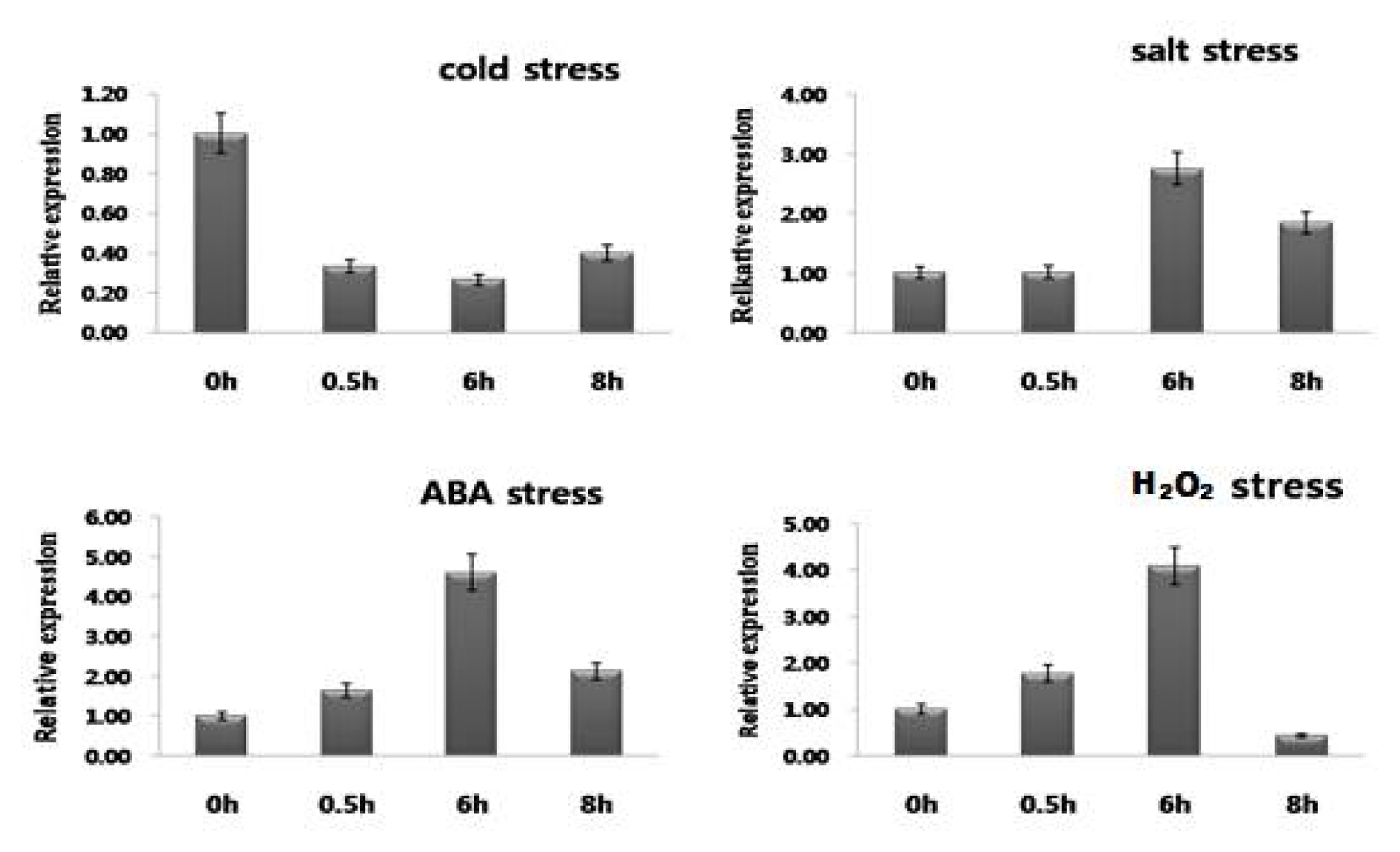
Fig. 4Ti-plasmid vectors for overexpression and silencing NtROS2a gene and transgenic plants. (A); Ti-plasmid vector for overexpression of NtROS2a gene. (B); Ti-plasmid vector for gene silencing (RNAi) of NtROS2a gene. (C); Transformation with Agrobacterium on MS medium with selectable kanamycin, (a); Survival response of explants in the selection medium after 4 weeks of co-cultivation (b); Appearance of shoot buds from the cut edges of the surviving green explants and development of small multiple shoots (c); Differentiation and elongation of green resistant shoots under selection after 8 weeks of culture (d); Acclimation of transgenic plants.

Fig. 5Quantitative RT-PCR analysis. WT; Wild type tobacco (Nicotiana tabacum cv Xanthi nc), (A); pBI121::NtROS2a (OX lines), (B); pKANNIBAL::NtROS2a (RNAi lines). CT values were calculated using actin expression level as a control. Error bars show the standard error of the mean for three replicate measurements.
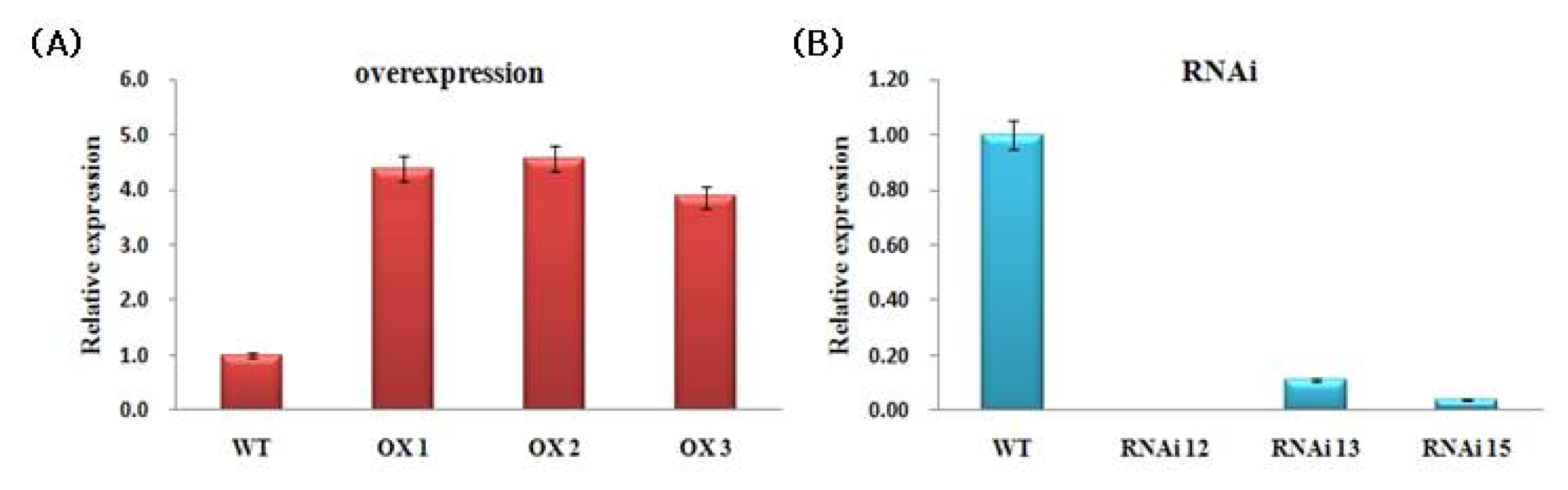
Fig. 6NaCl and H2O2 sensitivity in transgenic plants. (A) and (B) showed the phenotype of leaf discs from the WT plants, OX lines and RNAi lines after incubation in 250 mM NaCl and 3% H2O2 solution for 5 d, and corresponding measurement of chlorophyll contents respectively. Leaf discs floating in water served as the control. The standard deviation is represented by the vertical bar in graph (n=3).
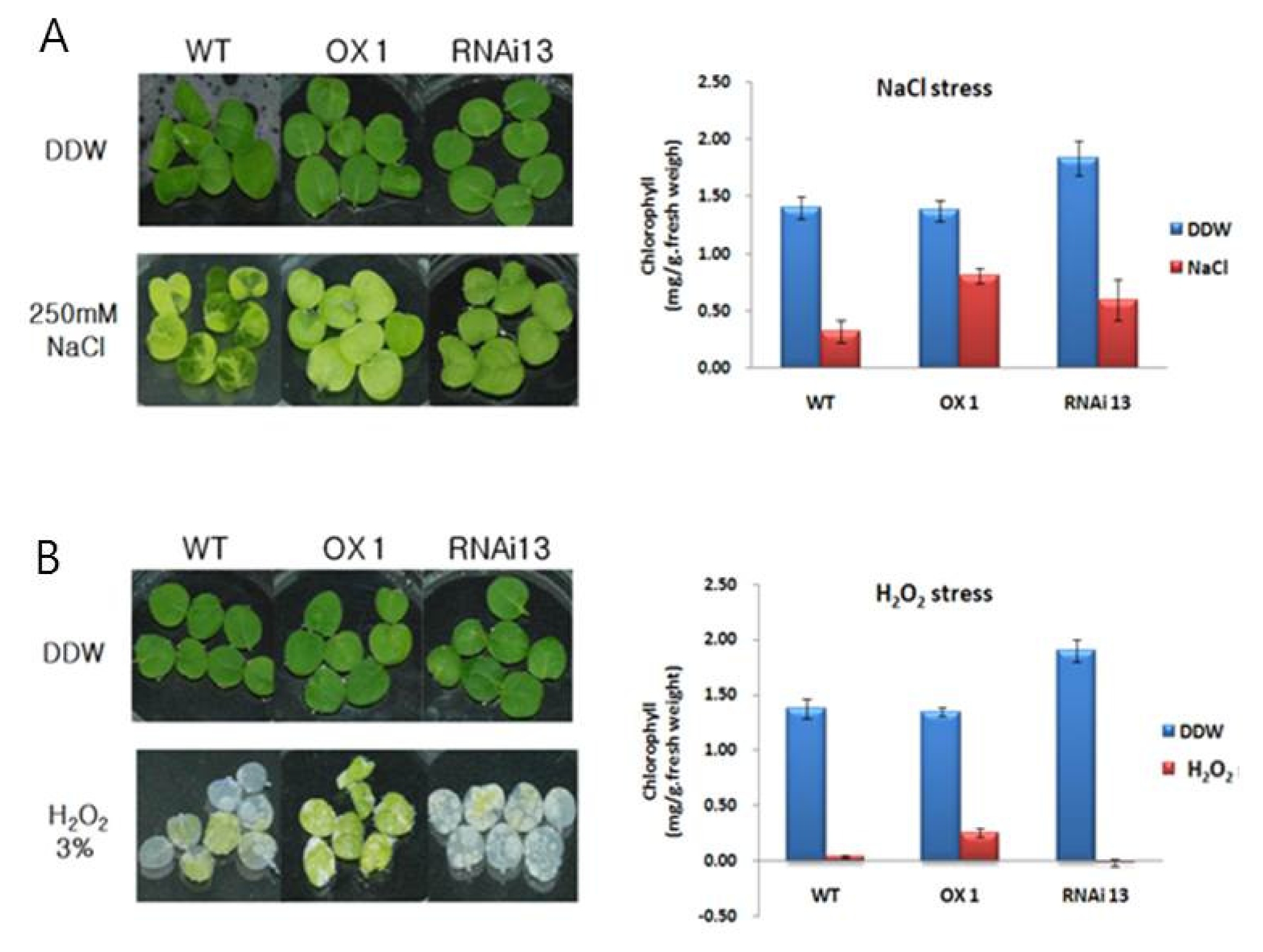
Fig. 7Germination assay under salt and osmotic stresses in transgenic lines (A); 125 mM NaCl stresses, (B); 2% PEG stresses. (C); Root elongation under salt and osmotic stresses in transgenic lines. Root growth under 125 mM NaCl and 2% PEG for 2 weeks after germination.
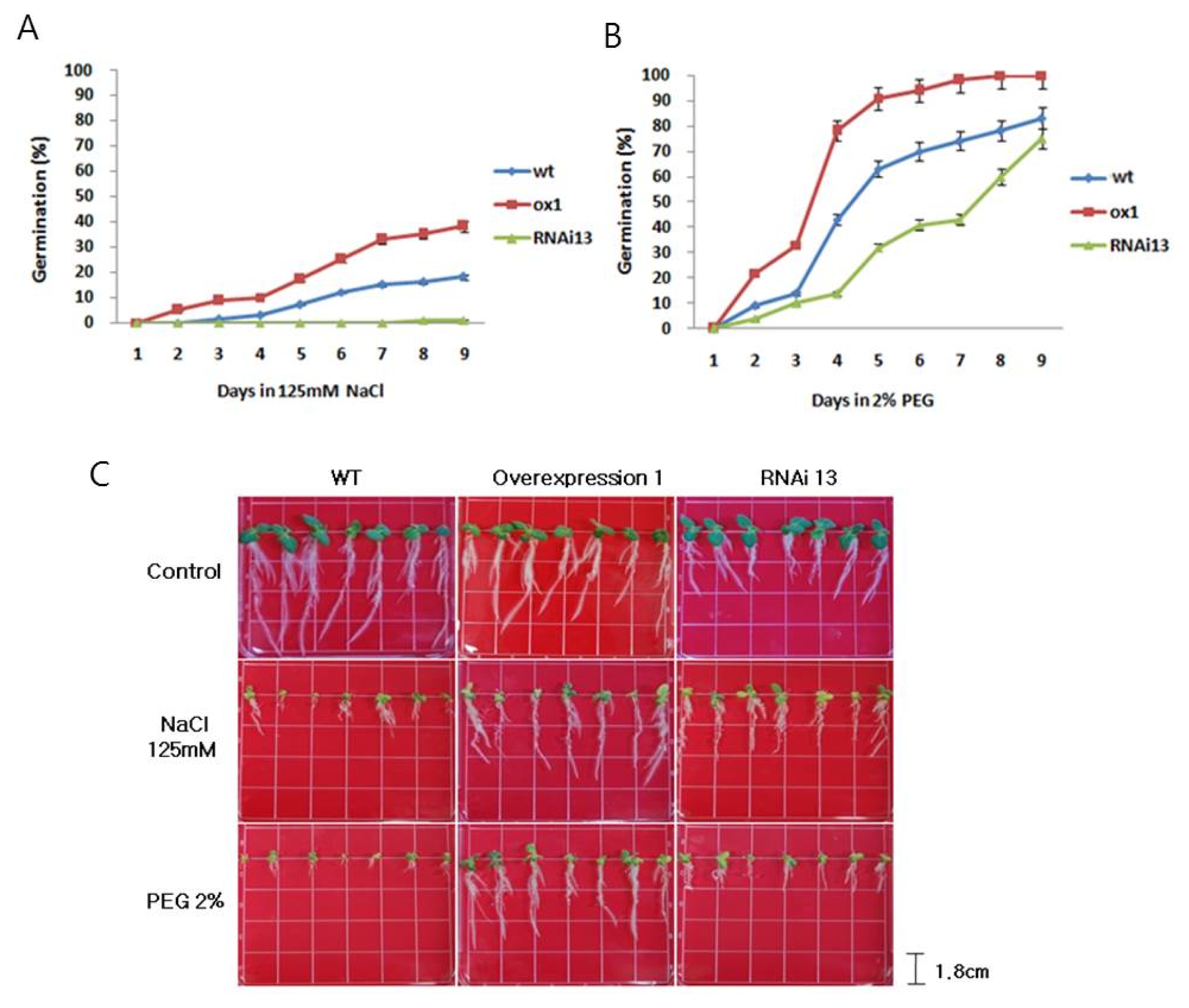
Table 1Primers sequence used for NtROS2a analysis.
Table 1
|
Primers |
Sequence(5′-3′) |
|
NtROS2a-RT F |
TGTAAGTCTGAGCGAAGAGC |
|
NtROS2a-RT R |
GGAAAGCTAGGTGGTGAAGT |
|
actin-RT F |
TACATGTTCACCACCACTGC |
|
actin-RT R |
AAGCTCCTGCTCGTAGTCAA |
|
35s-F |
TGAGACTTTTCAACAAAGGGTA |
|
NtROS2a-R |
ACAATGGGCTCTGGTGTTGC |
|
Kana F |
ATGATTGAACAAGATGGATTGCAC |
|
Kana R |
TCAGAAGAACTCGTCAAGAAGGC |
|
URP250 |
CCAGTTGCTATAAGAGGCAT |
|
URP251 |
ATCCAAGGTCCGAGACAACC |
|
URP253 |
CCCAGCAACTGATCGCACAC |
|
URP255 |
ATGTGTGCGATCAGTTGCTG |
References
- Aguis F, Kapoor A, Zhu JK. 2006. Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation. Proc Natl Acad Sci USA. 103: 11796-11801.
- Aono M, Kubo A, Saji H, Tanaka K, Kondo N. 1993. Enhanced tolerance to photooxidative stress of transgenic Nicotiana tabacum with high chloroplastic glutathione reductase activity. Plant Cell Physiology. 34: 129-135.
- Begum NA, Izumi N, Nishikori M, Nagaoka H, Shinkura R, Honjo T. 2007. Requirement of non-canonical activity of uracil DNA glycosylase for class switch recombination. J Biol Chem. 282: 731-742.
- Bird AP. 2002. DNA methylation patterns and epigenetic memory. Genes Dev. 16: 6-21.
- Cao X, Jacobsen SE. 2002. Locus-specific control of asymmetric and CpNpG methylation by the DRM and CMT3 methyltransferase genes. Proc Natl Acad Sci USA. 16491-16498.
- Chakrabarty D, Yu KW, Paek KY. 2003. Detection of DNA methylation changes during somatic embryogenesis of Siberian ginseng (Eleuterococcus senticosus). Plant Sci. 165: 61-68.
- Chomczynski P, Sacchi N. 1987. Single Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol- Chloroform Extraction. Anal Biochem. 162: 156-159.
- Choi CS, Sano Hiroshi. 2007. Abiotic-stress induces demethylation and transcriptional activation of a gene encoding a glycerophosphodiesterase-like protein in tobacco plants. Mol Genet Genomics. 277: 589-600.
- Demeulemeester MAC, Van Stallen N, De Proft MP. 1999. Degree of DNA methylation in chicory (Cichorium intybus L.). influence of plant age and vernalisation. Plant Sci. 142: 101-108.
- Finnegan EJ, Peacock WJ, Dennis ES. 1996. Reduced DNA methylation in Arabidopsis thaliana results in abnormal plant development. Proc Natl Acad Sci USA. 93: 8449-8454.
- Gehring M, Huh JH, Hsieh TF, Penterman J, Choi Y, Harada JJ, Goldberg R, Fischer RL. 2006. DEMETER DNA glycosylase establishes MEDEA polycomb gene self- imprinting by allele-specific demethylation. Cell. 124: 495-506.
- Kankel MW, Ramsey DE, Stokes TL, Flowers SK, Haag JR, Jeddeloh JA, Riddle NC, Verbsky ML, Richards EJ. 2003. Arabidopsis MET1 cytosine methyltransferase mutants. Genetics. 163: 1109-1122.
- Krokan HE, Standal R, Slupphaug G. 1997. DNA glycosylases in base excision repair of DNA. Biochem J. 325: 1-16.
- Law JA, Jacobsen SE. 2010. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat Rev Genet. 11: 204-220.
- Li E, Bestor TH, Jaenisch R. 1992. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell. 69: 915-926.
- Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods. 25: 402-408.
- Morales-Ruiz T, Ortega-Galisteo AP, Ponferrada-Marin MI, Martinez-Macias MI, Ariza RR, Roldan-Arjona T. 2006. DEMETER and REPRESSOR OF SILENCING 1 encode 5-methylcytosine DNA glycosylases. Proc Natl Acad Sci USA. 103: 6853-6858.
- Murashige T, Skoog F. 1962. A revesed medium for rapid growth and bioassays with tobacco cultures. Physiol Plant. 15: 473-497.
- Reik W, Dean W, Walter J. 2001. Epigenetic reprogramming in mammalian development. Science. 293: 1089-1093.
- Ronemus MJ, Galbiati M, Ticknor C, Chen J, Dellaporta SL. 1996. Demethylation-induced developmental pleiotropy in Arabidopsis. Science. 273: 654-657.
- Stancheva I, Meehan RR. 2000. Transient depletion of xDnmt1 leads to premature gene activation in Xenopus embryos. Genes Dev. 14: 313-327.
- Sano H, Kamada I, Youssefian S, Katsumi M, Wabiko H. 1990. Asingle treatment of rice seedlings with 5-azacytidine induces heritable dwarfism and under methylation of genomic DNA. Molecular Genetics and Genomics. 220: 441-447.
- Steward N, Ito M, Yamakuchi Y, Koizumi N, Sano H. 2002. Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress. J Biol Chem. 277: 37741-37746.
- Wada Y, Miyamoto K, Kusano T, Sano H. 2004. Association between up-regulation of stress-responsive genes and hypomethylation of genomic DNA in to tobacco plants. Mol Genet Genomics. 271: 658-666.
- Zhang X, Yazaki J, Sundaresan A, Cokus S, Chan SW, Chen H, Henderson IR, Shinn P, Pellegrini M, Jacobsen SE, et al. 2006. Genome-Wide high-resolution mapping and functional analysis of DNA methylation in Arabidopsis. Cell. 126: 1189-1201.