Abstract
Panicle blast is a serious disease in rice production in Korea. To develop rice cultivar resistant to panicle blast, the Pb1 gene, originating in the indica cultivar ‘Modan’, was incorporated into an elite japonica cultivar ‘Ilmi’, which is highly susceptible to panicle blast, by marker assisted backcross breeding. ‘Hwayeong’ was selected as the donor parent for the Pb1 gene based on 10 years of panicle blast data. The inheritance of panicle blast resistance in ‘Hwayeong’ is controlled by a single dominant gene. This panicle blast resistance gene, Pb1, from ‘Hwayeong’ was incorporated into the susceptible variety, ‘Ilmi’, by marker assisted backcross approach using DNA marker RM206. As a result, ‘Saeilmi’ containing the Pb1 gene showed resistance to panicle blast. The agronomical and grain quality traits of ‘Saeilmi’ were similar to those of the recurrent parent ‘Ilmi’. Molecular background analysis revealed that ‘Saeilmi’ had the least donor segment introgression in the locus of the Pb1 gene on chromosome 11.
-
Key words: Rice, Panicle blast, Marker assisted selection, Saeilmi
INTRODUCTION
Disease and insect pests seriously constrain rice yields. Among these, rice blast, which is caused by the fungus
Magnaporthe grisea, is the most widespread disease in most commercial rice-growing areas of the world. Annual losses caused by the rice blast fungus vary from 10% to 30% of the harvest, with even 10% loss being enough to feed 6o million people for one year (
Skamnioti et al. 2009). To minimize the loss from rice blast, the use of resistant varieties is one of the most effective ways to control the disease. However, the useful life span of resistant varieties was only about three years due to the breakdown of resistance induced by the pathogenic variability of
M.
grisea rice (
Khush and Brar 1991). In the 1970s, Tongil-type cultivars accounted for more than 75% of the total rice production in Korea. However, the proliferation of diverse cultivars triggered genetic mutations of rice blast leading to differentiation of the blast fungus,
M. grisea. This development eventually led to the breakdown and loss of blast resistance in Tongil-type rice cultivars (
Lee and Park 1979;
Ryu et al. 1987). The same virulent
M. grisea isolates also caused great damage to leaf and panicle blast in
japonica rice. This is attributed to the narrow genetic diversity of
japonica cultivars to panicle blast resistance and the rapid expansion of cultivation area (
Han et al. 2001;
Kwon et al. 2002). Thus, the development of a blast-resistant cultivar has still been considered to be the most effective and economical breeding strategy to control the spread of the disease (
Bonman 1992). Up to date, approximate 50 resistance genes to blast disease have been identified. Some major genes, such as
Pi-z,
Pi-zt,
Piz-5 (Pi2), Pi5,
Pi9, and
Pi40, were reported to have resistance to a broad spectrum of
M. grisea isolates (
Hayashi et al. 2004;
Jeon et al. 2003;
Liu et al. 2002;
Suh et al. 2009). However, those blast resistance genes were identified for a leaf blast disease not to panicle blast. The genetic and QTLs of the resistance to leaf blast and panicle blast might be independently controlled by different genes (
Sirithunya et al. 2002). The damage of panicle blast is more destructive than that of leaf blast in practical rice cultivation. Therefore, development of high resistance to panicle blast variety is considered in breeding programs.
A broad-spectrum panicle blast resistance gene
Pb1, a single dominant locus, was found in the
japonica rice cultivar ‘Tsukinohikari’ derived from the
indica cultivar ‘Modan’ (
Fujii et al. 1999;
Fujii et al. 2000). The
Pb1 gene was identified on chromosome 11 encoding coiled–coil (CC)-NBS-LRR and contains two putative CC domains in the N terminus (
Hayashi et al. 2010). The
Pb1 gene has been classified as a field resistance gene (or adult resistance) because of its durability and quantitative nature of resistance (
Hayashi et al. 2010). Rice cultivars containing
Pb1 gene were rather weakly resistant to leaf blast, but remarkable resistant to panicle blast (
Fujii et al. 2000;
Hayashi et al. 2010).
The use of molecular markers increases the speed of the selection process in breeding programs. The marker-assisted backcrossing strategy is to transfer a specific allele at the target locus from a donor line to a recipient line while selecting against donor introgressions across the rest of the genome (
Jairin et al. 2009;
Suh et al. 2011;
Wongsaproma et al. 2010). The PCR based marker RM206 for panicle blast resistant gene was identified by
Hayashi et al. (2004). However, there has been no scientific report in the development of a rice cultivar resistant to the panicle blast. Here, we report a new variety ‘Saeilmi’ carrying
Pb1 gene using the marker-assisted backcrossing approach.
MATERIALS AND METHODS
Plant materials and development of near isogenic lines (NILs)
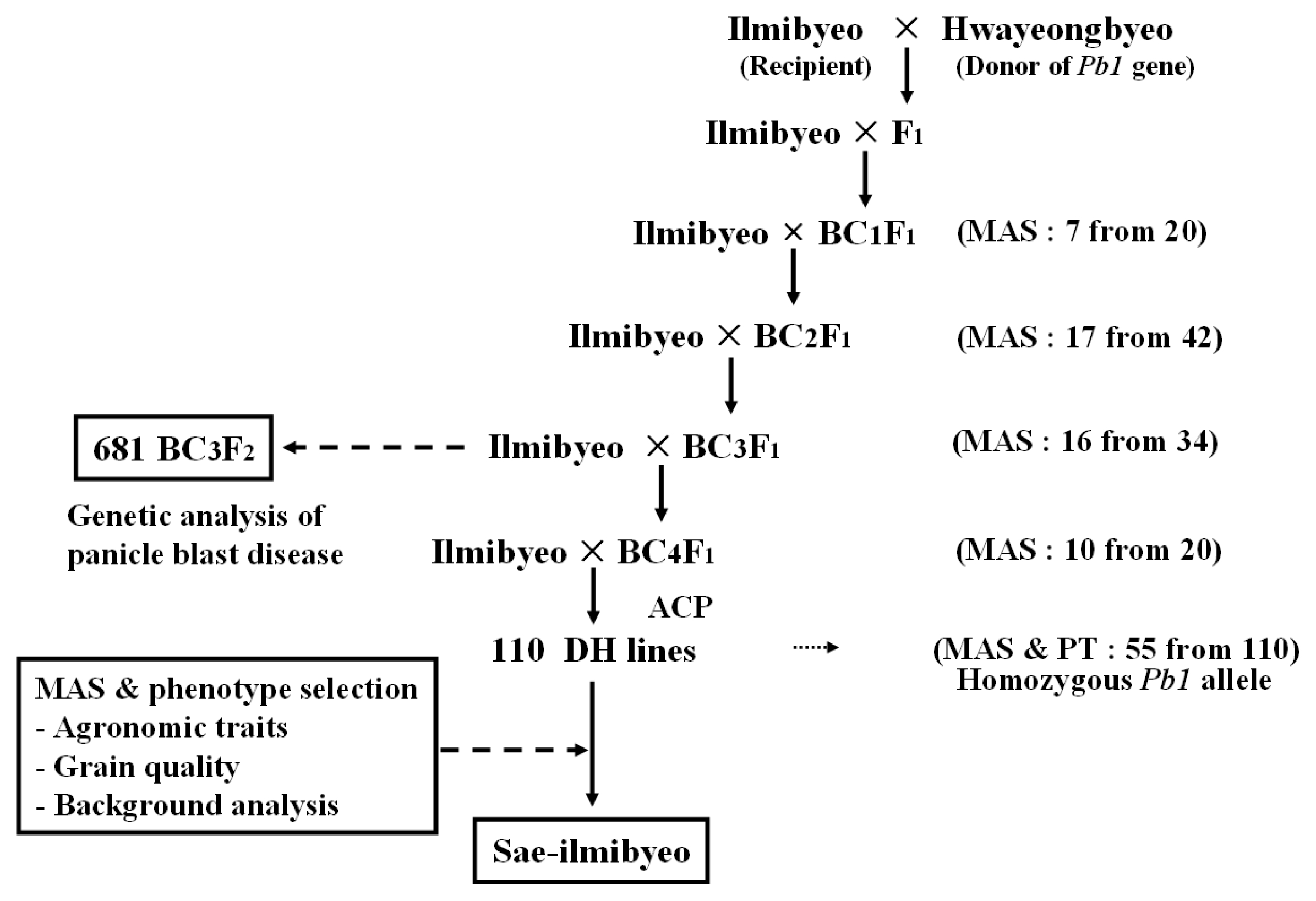
The rice cultivars ‘Ilmi’ and ‘Hwayeong’ were used as the parents in the marker assisted backcross program. ‘Ilmi’ was used as the recipient variety and ‘Hwayeong’, one of the ‘Modan’-derived panicle blast resistant cultivars, was used as the donor of
Pb1 gene. Near isogenic lines (NILs) were developed through the marker assisted backcross strategy
. In each backcross generation (BC
nF
1), individual BC
nF
1 were selected based on the heterozygous allele of SSR marker RM206 linked to the
Pb1 gene based on a report of
Hayashi et al. (2004). A total of 687 individuals of BC
3F
2 were obtained and used to identify the
Pb1 gene. One hundred and ten NILs were developed by anther culture method in the BC
4F
1 generation and used to evaluate the agronomic traits, their grain quality, and association of SSR marker RM206 analysis.
We carried out field tests of panicle blast using the standard evaluation method (
RDA 2003). One hundred and ten NILs and 687 individuals of BC
3F
2 were grown in an experimental plot at the Department of the Functional Crop, Miryang, Korea. Seeds were sown on 30 June and transplanted on 20 July in 2007 and in 2008. The planting density was 22.22 m
−2 (15 cm × 30 cm) and N-P
2O
5-K
2O fertilizers were applied at the level of 90 - 45 - 57 kg/ha. The incidence of panicle blast was evaluated based on the percentage of infected panicles per plant at 40–50 days after heading. The number of panicles showing symptoms of panicle blast disease was expressed as percent infection.
The parents and eight NILs derived from marker assisted backcross were planted in a eight-row plot with 200 plants per row by 30 × 15 cm spacing in a randomized complete block design with three replications. The were evaluated for agronomic traits in the rice experimental plot of the Department of the Functional Crop, Miryang, Korea, using the standard evaluation method of rice (
RDA 2003). For each line, ten plants in the middle rows were used to determine heading date, culm length, panicle length, panicle number, number of grains per panicle, grain filling ratio, and grain yield. Grain yield per plot was evaluated based on a grain harvest of 100 plants in the central row of each plot.
Grain quality was estimated for chalkiness (0: non-chalkiness, 3: high chalkiness) of brown rice, amylose content and protein content of milled rice. Amylose content was determined by the relative absorbency of starch-iodine color by
Juliano’s (1971) modified method. Protein content was calculated by multiplying grain nitrogen content by 5.95, the nitrogen–protein conversion factor. Grain nitrogen content was measured by the Micro-Kjeldahl method (2300 Kjeltec Analyzer, Foss). Palatability characteristics of the rice samples were measured using a TOYO meter (MB-90A and MA-90B, Toyo Co.) to determine the glossiness of the cooked rice. The head rice ratio of milled rice was measured by a single grain rice inspector (RN600, Kett Co.)
The SSR marker RM206, which is the most tightly linked to panicle blast resistance gene Pb1, was used to confirm the presence of Pb1 gene in marker assisted backcross lines. Polymerase chain reaction (PCR) was performed in an ABI 9700 Thermocycler (Germany) with a total volume of 20 μl containing 20 ng of template DNA, 5 pmoles of each primer, 1.5 mM of MgCl2, 0.2 mM of dNTP and 0.5 U of Taq polymerase. The PCR amplification condition was one cycle of denaturation at 95°C for 5 min, followed by 40 cycles at 95°C for 30 s, 55°C for 30 s, and 72°C for 1 min, with a final extension at 72°C for 10 min. After determining the amplification success, the PCR product was detected using 3% agarose gel electrophoresis with ethidium bromide staining.
Molecular analysis of genetic background
A total of 192 SSR markers distributed evenly on the 12 chromosomes with an average interval of 1.0–2.0 Mb were used in a genome-wide survey to identify the chromosome segment substitution locations in ‘Saeilmi’. The polymorphism analysis was conducted following the procedure of
McCouch et al. (2002). The PCR products were detected using 4% denaturing polyacrylamide gel electrophoresis.
The least significant difference (LSD) was used for multiple mean comparisons using the SAS statistical software package (version 8.2; SAS Institute, Cary, NC).
RESULTS
Donor parent selection and genetic background of panicle blast resistance gene Pb1
In order to select the donor parent of
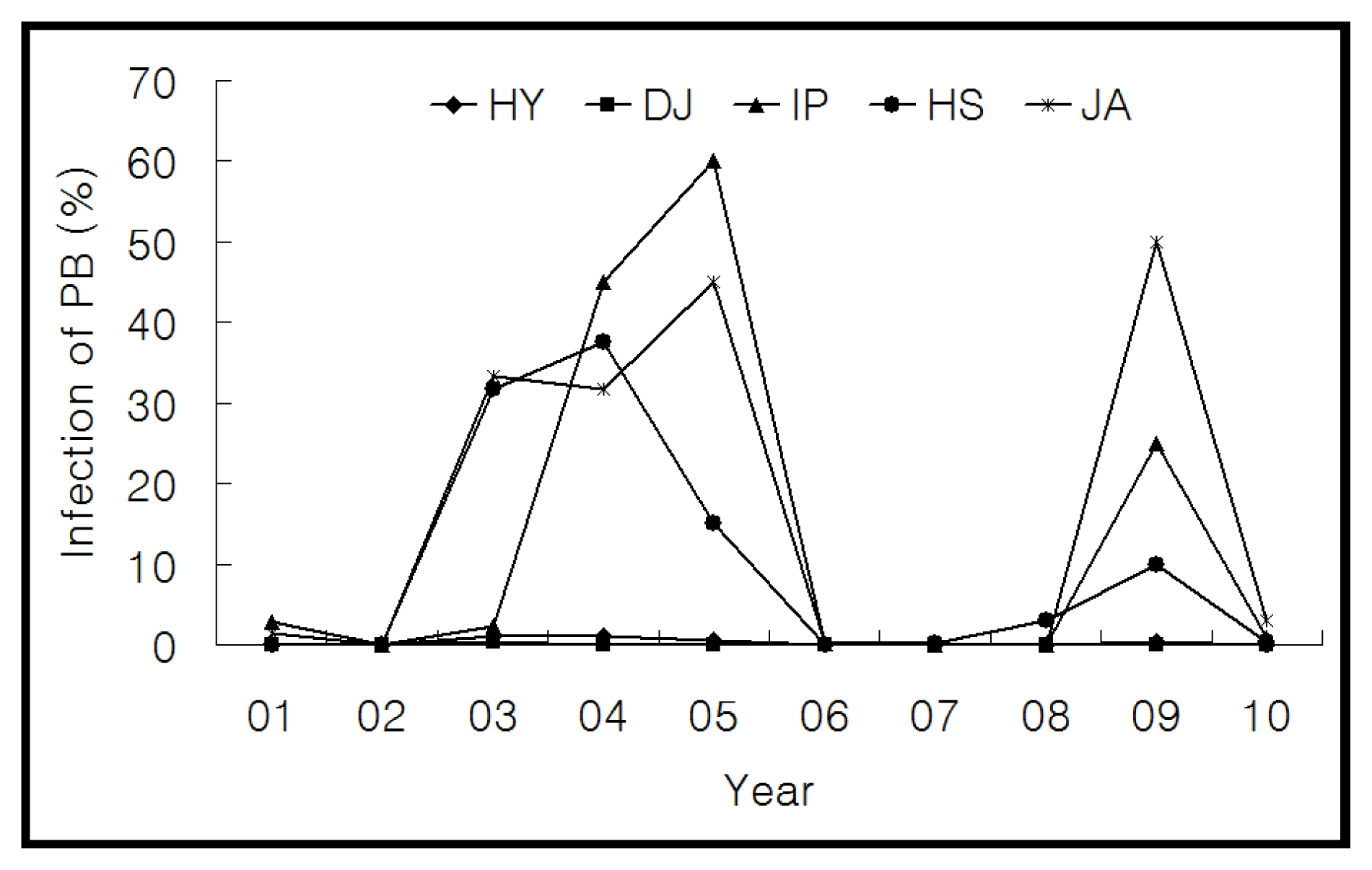
Pb1 gene among Korean
japonica rice, varietal reactions data to panicle blast was surveyed from 2001 to 2010 in Jecheon province, a known area prone to panicle blast in Korea. During these 10 years, the incidence of panicle blast in ‘Hwayeong’ and ‘Dongjin’ ranged from 0.0 to 1.0% and 0.0 to 0.4%, respectively. However, the incidence of panicle blast in ‘Ilpum’, ‘Hwaseong’, and ‘Juan’ ranged from 0.0 to 45.0%, 0.0 to 37.5%, and 0.0 to 50.0%, respectively (
Fig. 1). The incidence of panicle blast of ‘Hwayeong’ was significantly lower than that of susceptible cultivars ‘Ilpum’, ‘Hwaseong’, and ‘Juan’. As a result, we selected the ‘Hwayeong’ as the donor parent for
Pb1 gene based on the 10 years of field tests. In pedigree analysis, the panicle blast resistance gene,
Pb1, derived from ‘Hwayeong’ was introduced from
indica cultivar ‘Modan’ at an early stage of the
japonica rice breeding program in Korea.
F
1 progenies were produced from a cross between the panicle blast susceptible cultivar ‘Ilmi’ and the
Pb1 donor ‘Hwayeong’. The BC
1 progenies were obtained by backcrossing the F
1 plants with ‘Ilmi’ as recurrent parent. In the BC
nF
1 generation, individual plants showing heterozygous at the
Pb1 gene locus were identified and used for further backcrossing with the recurrent parent using a marker-assisted backcrossing scheme (
Fig. 2). The advanced backcross progenies of BC
2, BC
3, and BC
4 were obtained from the crosses of selected resistant BC
1F
1 (7 plants from 20 plants), BC
2F
1 (17 plants from 42 plants), BC
3F
1 (16 plants from 34 plants), and BC
4F
1 (10 plants from 20 plants) plant based from marker allele of RM206.
The panicle blast reaction was carried out using the modified method (
Roh et al. 2007). ‘Hwayeong’ showed significantly lower infection ratio to panicle blast than ‘Ilmi’ at 40–50 days after heading. The average percentage of panicle blast infection was 3.0% for ‘Hwayeong’ and 67.3% for ‘Ilmi’, respectively (
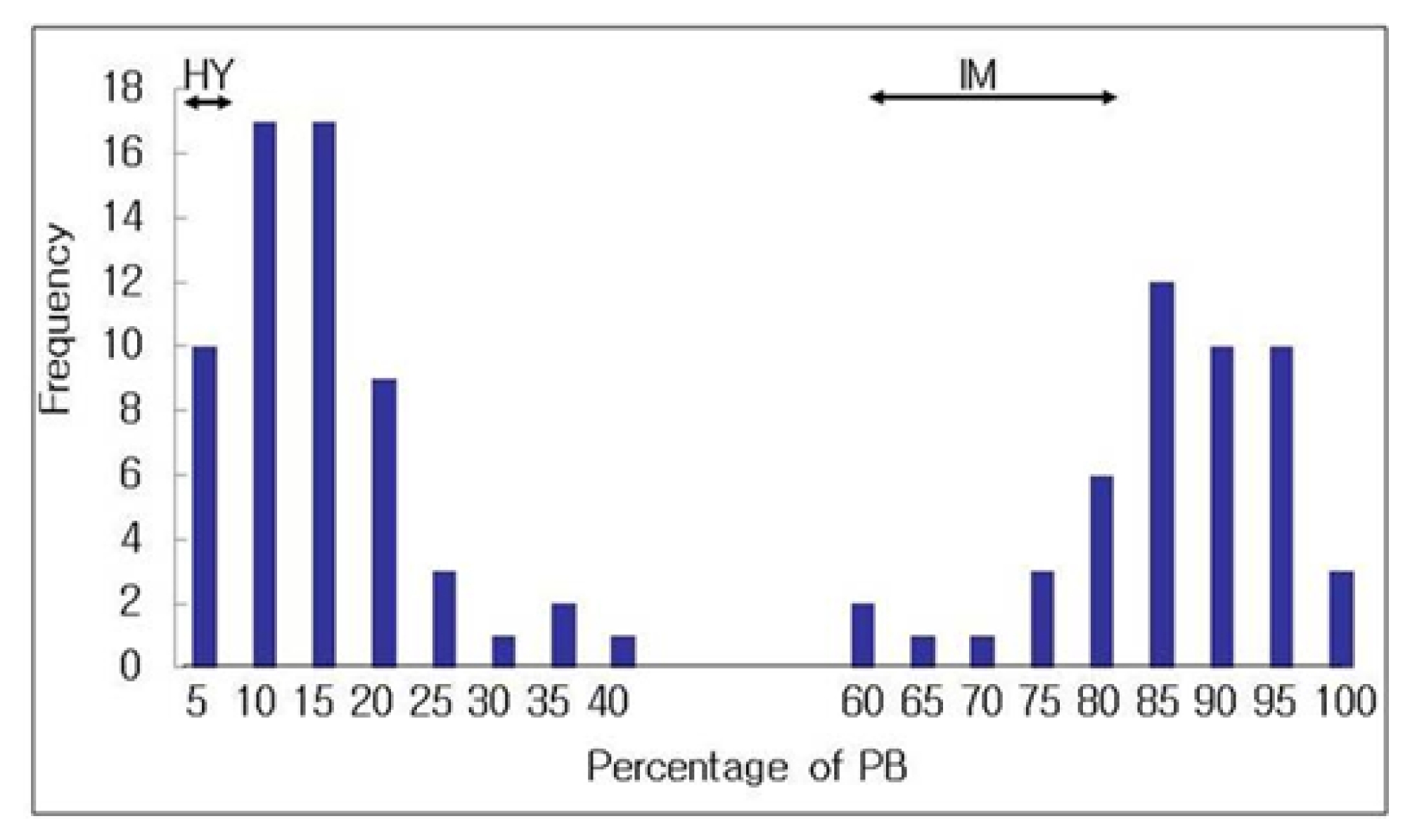
Fig. 3). The frequency distribution of panicle blast in the BC
4DH population (N=110) was bimodal distribution, resistance (0 to 40%) and susceptible (60 to 100%), suggesting that the inheritance of panicle blast resistance in ‘Hwayeong’ is controlled by a single gene (
Table 1).
The mode of panicle blast inheritance in donor parent, ‘Hwayeong’, was investigated in the BC
3F
2 population developed by the serial backcross with the panicle blast susceptible recurrent parent ‘Ilmi’. A total of 679 individuals of BC
3F
2 were classified as either resistant or susceptible, based on the infected percentage of panicle blast with 40% as the boundary. Segregation ratio of 519 resistant, and 160 susceptible plants was fit to a 3:1 ratio and (χ
2 = 0.747, df = 1,
p = 0.388) (
Table 1). This result confirmed that the inheritance of panicle blast resistance in ‘Hwayeong’ was controlled by a single dominant gene.
The field performances of the selected NILs were evaluated in 2007. The days to heading of elite NILs were almost the same as those of ‘Ilmi’. The culm length of NILs varied from 73.6 to 78.5 cm with an average of 76.2 cm. In major traits of yield components, the average number of grains per panicle in NILs was 125.5, which was much higher compared to that of ‘Ilmi’. The grain filling ratio and brown rice per rough rice ratio were also almost the same. Compared to that of ‘Ilmi’, NILs showed a little bit higher rice yield, with an average of 7.06 MT/ha. Consequently, except the number of grains per panicle, the main agronomic traits of the NILs were similar to those of ‘Ilmi’ because of the backcross progenies possessing undesirable agronomic traits were eliminated in early generations. As a results, one of the elite NIL of YR26754Acp13 possessing the
Pb1 gene with similar grain quality was selected and designated as ‘Saeilmi’ through a local adaptability test over 3 year (2009 to 2011). And most of the grain features in NILs, such as amylose and protein contents, were the same as those of ‘Ilmi’ (
Table 3).
‘Saeilmi’, developed from the backcross of recurrent parent ‘Ilmi’ and panicle blast resistant donor ‘Hwayeong’, was investigated for background degree of the whole genome using SSR markers. A total of 188 SSR markers were used to identify introgression of chromosome segments of donor parent. The marker polymorphism between ‘Hwayeong’ and ‘Ilmi’ was 24.6%. In ‘Saeilmi’, the introgression chromosome segments of ‘Hwayeong’ were distributed around the region of the
Pb1 gene located on chromosome 11 only. The percentage of donor parent chromosome substitution on chromosome 11 was 31.3% (
Table 4).
Field response of
Pb1 gene was conducted for panicle blast disease prone area such as Icheon, Jaecheon, Yaesan, and Iksan during the 3 years from 2009 to 2011. The average infected panicle blast percentage of the ‘Saeilmi’ ranged between 0.6% and 3.3%. The panicle blast resistance varieties ‘Dongjin’ and ‘Hwayeong’ also showed 0.6% and 1.3% of infected panicle blast, respectively. However, the average percentage of infected panicle blast of the susceptible varieties, ‘Hwaseong’ and ‘Ilpum’ ranged between 4.7% and 18.7% and between 5.7% and 25.0%, respectively (
Table 5).
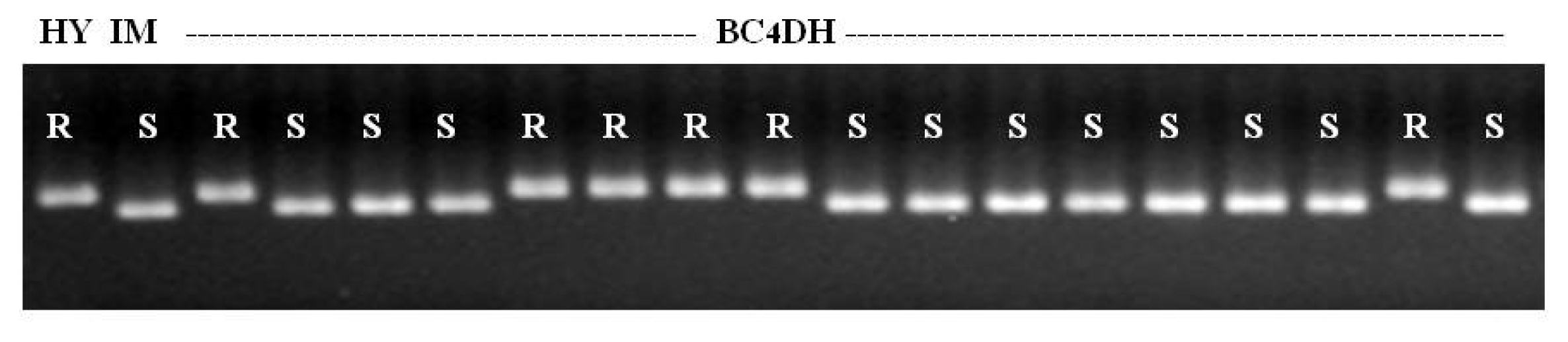
To confirm the presence of the
Pb1 gene, RM206, which is tightly linked to panicle blast gene
Pb1, was screened in BC
4 DH lines. Consequently, the RM206 DNA marker was well associated with the field resistance of panicle blast in each individual line (
Fig. 4). And as shown in
Fig. 5, ‘Saeilmi’ showed clear resistance, while ‘Ilmi’ was susceptible to panicle blast.
DISCUSSIONS
The main objective of this study was to combine panicle blast resistance gene
Pb1 into an elite cultivar using marker assisted backcross. Conventional breeding programs for disease resistance are laborious, time consuming, and highly dependent on environmental conditions compared to marker assisted selection, which is simple, efficient, and precise (
Babujee and Gnanamanickam 2000). Application of a marker assisted selection program has been successfuly utilized in developing new varieties for resistance to blast diseases (
Jairin et al. 2009;
Suh et al. 2011).
At present, most of the Korean
japonica cultivars to carry the blast resistance genes
Pia, Pib, Pii, Pik,
Pita, and
Piz based on pedigree analysis, molecular marker analysis, and reaction to leaf blast disease (
Cho et al. 2007;
Kwon et al. 2008). However, because of difficulty in infection of panicle blast under fluctuating environmental conditions in which each plant’s tendency to escape infection during the heading and maturing stage, which is the genetic and molecular basis of panicle blast resistance, has been poorly understood in Korea.
Fujii et al. (1999) reported that the
Pb1 gene is located on the long arm of chromosome 11 and linked to the RSV resistance gene originating from
indica cultivar ‘Modan’. The
Pb1 gene has been classified as a field resistance gene because of its durability and quantitative reaction. Interestingly, the
Pb1 gene was blast susceptible during young vegetative stages, however the resistance level increased as the plant grew, and persisted even after heading (
Fujii and Hayano-Saito 2007).
The best way to understand the durability of the panicle blast resistance mechanism is probably to examine a cultivar or genotype that has remained resistant to a highly variable pathogen for a long period of cultivation. ‘Hwayeong’ was selected as the donor parent of the
Pb1 gene for marker-assisted backcross in this study because ‘Hwayeong’ has shown the durable resistance to panicle blast from 2001 to 2010 in Jecheon. In analysis of breeding pedigree, ‘Hwayeong’ revealed that the
Pb1 gene originated from Japanese cultivar ST.No.1, which is derived from the
indica cultivar ‘Modan’ (
Fujii et al. 1999).
In rice genetic improvement such as disease resistance and abiotic stress tolerance, the amount of linkage drag would increase as more major resistance genes combine into a variety (
Jairin et al. 2009;
Lee et al. 2005). So it is important to consider how to optimize the combination of a broader spectrum resistance. When the target gene is linked with an unfavorable allele, background selection using molecular marker can be performed to break down the linkage drag (
Jena and MacKill 2008). Although marker-assisted selection technology is well known as a powerful tool for dissecting linkage drag, the most efficient way in a breeding program is the utilization of donor parent with no undesirable alleles in the target gene locus.
The agronomic characteristics and grain quality traits of NILs developed from marker assisted backcross were almost the same as ‘Ilmi’, indicating that any linkage drags with unfavorable alleles of ‘Hwayeong’ did not occurs. In addition, NILs developed from marker assisted backcross showed resistance to K
3 race of bacterial blight disease. In this respect, it is clear that the bacterial blight resistance gene
Xa3 of
‘Hwayeong’ located on the end of the long arm of chromosome 11 (
Kang et al. 2003) is closely linked to
Pb1 gene.
Agronomic traits related to yield stability and sustainability are major concerns of rice breeding efforts. No significant difference was statistically observed in agronomic characteristics and grain quality between ‘Ilmi’ and NILs developed from marker assisted backcross. However, the grain yield of the NILs was slightly increased than greater of ‘Ilmi’, apparently because of increased number of grains per plant. Finally, as a result, we selected the best line among of the NILs and developed a new variety, ‘Saeilmi’. The results obtained from the NILs clearly showed that ‘Saeilmi’ had similar agronomic charateristics to the recurrent parent ‘Ilmi’.
The effectiveness of marker-assisted backcrossing in enhancing the introgression of major resistance QTL alleles has been well documented (
Jairin et al. 2009;
Suh et al. 2011). The present study demonstrated that marker assisted backcross was sufficient to recover the majority of the important characteristics of the original plant without any linkage drag of unfavorable background. In many cases, molecular markers linked to disease and insect resistance are available to facilitate direct selection in the breeding program. And those techniques apparently enhance stable resistance to elite varieties that are renowned for their wide adaptation and production stability across environments.
The majority of resistance breeding has depended on field screening methods in Korea. It was difficult to develop a durable resistance to panicle blast using conventional breeding methods due to the failures to infect satisfactorily individual plants by fluctuating environmental conditions. In this report, we demonstrated that marker assisted backcross effectively enhanced the introgression of panicle resistance gene Pb1. We have successfully developed a near isogenic variety ‘Saeilmi’ which is carrying the Pb1 gene and is identical to japonica elite cultivar ‘Ilmi’ by using marker-assisted backcrossing. Thus, marker assisted backcross was shown to be a great tool in breeding programs.
ACKNOWLEDGMENTS
This work was supported by the Rural Development Administration (Project No. PJ0102682014), Republic of Korea.
Fig. 1Fluctuation of the percentage of blast-diseased panicle in Jecheon located in the northern part of south Korea for 10 years (2001 to 2010). HY: Hwayeong, DJ: Dongjin, IP: Ilpum, HS: Hwaseong, JA: Juan.
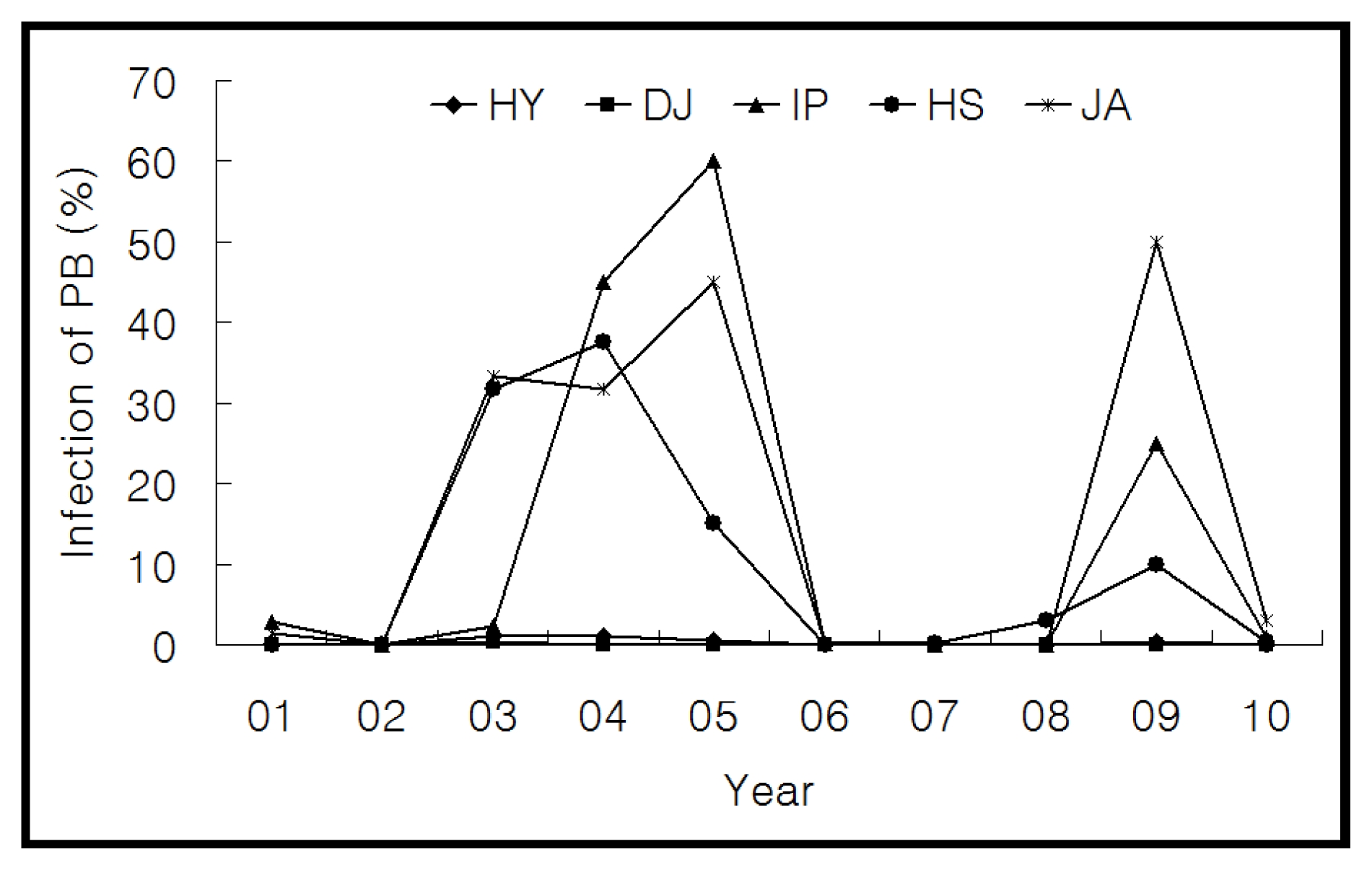
Fig. 2Flow diagram showing steps involved in marker-assisted backcross breeding program.
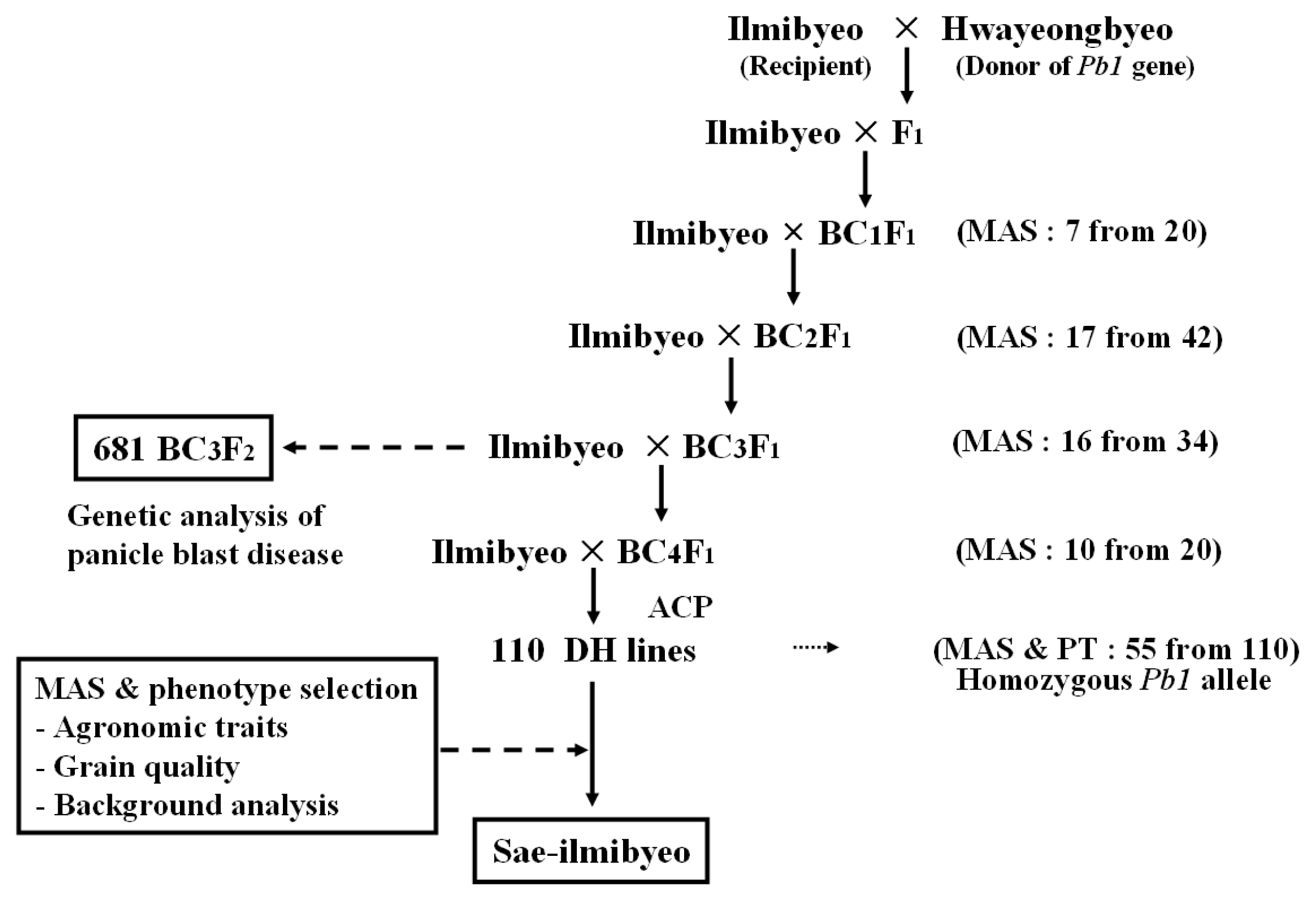
Fig. 3Frequency distribution of the percentage of panicle blast in 110 backcrossed double haploid lines (BC4DH) derived from a donor parent Hwayeong and a recurrent parent Ilmi.
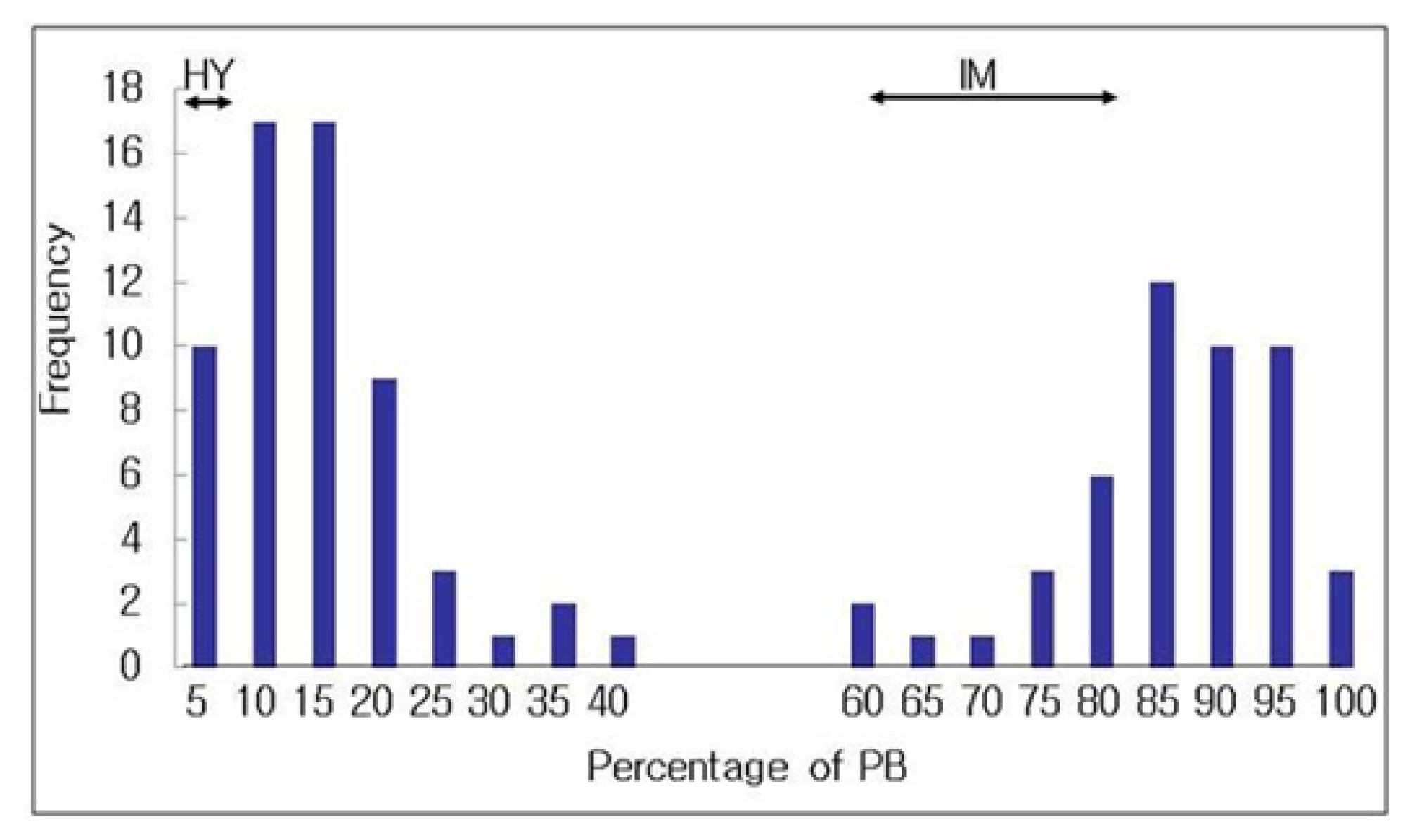
Fig. 4Association between reaction to panicle blast disease and marker allele of RM206 tightly linked to Pb1 gene in NILs. HY: Hwayeong (Donor parent), IM: Ilmi (recurrent parent), R: resistance, S: susceptible.
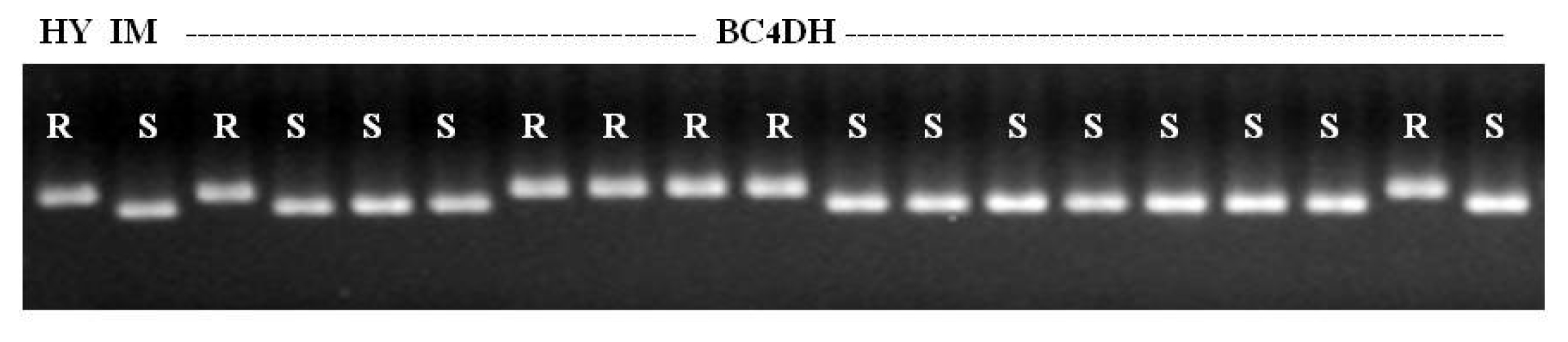
Fig. 5Symptoms of rice panicle blast in cultivar Ilmi (Pb1−) and Saeilmi (Pb1+).

Table 1Genetic analysis of resistance to panicle blast disease BC4 DH and BC3F2 population derived from a backcross Ilmi (recurrent) and Hwayeong (donor).
Table 1
|
Designation |
Generation |
No. of tested plants |
Reaction to PBz)
|
Expected ratio |
χ2
|
P-value |
|
|
Resistance |
Susceptible |
|
YR26754 |
BC4-DH |
110 |
59 |
51 |
1:1 |
0.582 |
0.446 |
|
YR26175 |
BC3F2
|
679 |
519 |
160 |
3:1 |
0.747 |
0.388 |
Table 2Agronomic traits of eight BC4 DH lines showing resistance to panicle blast.
Table 2
|
Designation |
z) DH1) (days) |
CL2) (cm) |
PL3) (cm) |
NP4)
|
NGP5)
|
GFR6) (%) |
HR7) (%) |
GY8) (MT/ha) |
|
Ilmi |
85y)
|
75.0 |
21.1 |
16.1 |
111.4 |
89.5 |
82 |
6.82 |
|
Hwayeong |
78 |
75.4 |
22.4 |
15.8 |
110.8 |
90.6 |
83 |
6.19 |
|
YR26754Acp13 |
83 |
76.9 |
20.9 |
15.7 |
124.5 |
89.9 |
82 |
7.27 |
|
YR26754Acp15 |
83 |
78.5 |
20.6 |
15.1 |
141.0 |
90.3 |
82 |
7.30 |
|
YR26754Acp29 |
84 |
77.1 |
20.8 |
15.4 |
124.1 |
91.9 |
82 |
7.10 |
|
YR26754Acp40 |
85 |
73.6 |
19.2 |
15.5 |
119.7 |
90.7 |
80 |
6.43 |
|
YR26754Acp58 |
84 |
76.9 |
20.5 |
14.8 |
133.3 |
89.0 |
82 |
7.18 |
|
YR26754Acp61 |
83 |
74.8 |
21.3 |
15.6 |
121.9 |
84.5 |
82 |
7.16 |
|
YR26754Acp79 |
84 |
75.9 |
20.7 |
15.8 |
120.5 |
89.3 |
82 |
6.96 |
|
YR26754Acp91 |
84 |
75.8 |
20.0 |
14.6 |
118.7 |
86.7 |
82 |
7.12 |
|
Mean |
84 |
76.2 |
20.5 |
15.3 |
125.5 |
89.0 |
82 |
7.06 |
Table 3Grain qualities of eight BC4 DH lines resistant to panicle blast.
Table 3
|
Designation |
z) AC1) (%) |
PC2) (%) |
HRR3) (%) |
WC/WB4) (0–9) |
PA5)
|
|
Ilmi |
19.1y)
|
6.2 |
93.1 |
0/0 |
68.9 |
|
Hwayeong |
18.6 |
6.5 |
92.1 |
0/0 |
67.4 |
|
YR26754Acp13 |
19.0 |
6.3 |
94.7 |
0/0 |
67.6 |
|
YR26754Acp15 |
18.9 |
6.2 |
94.6 |
0/0 |
67.8 |
|
YR26754Acp29 |
19.2 |
6.3 |
96.0 |
0/0 |
67.8 |
|
YR26754Acp40 |
19.3 |
6.2 |
95.0 |
0/0 |
67.9 |
|
YR26754Acp58 |
18.5 |
6.3 |
95.3 |
0/0 |
67.0 |
|
YR26754Acp61 |
19.6 |
6.2 |
93.4 |
0/0 |
68.0 |
|
YR26754Acp79 |
19.4 |
6.3 |
93.6 |
0/0 |
68.6 |
|
YR26754Acp91 |
19.1 |
6.4 |
92.9 |
0/0 |
69.0 |
|
Mean |
19.1 |
6.2 |
94.4 |
0/0 |
68.0 |
Table 4Simple sequence repeat markers with polymorphism between recurrent parent and donor parent and substituted chromosome segments from donor parent in Saeilmi.
Table 4
|
Chromosome No. |
No. of tested SSR markers |
Chromosome length (Mb) |
Marker interval (Mb) |
Polymorphism (%) |
substituted chromosome segments (%) |
|
1 |
16 |
43.2 |
2.7 |
12.5 |
0 |
|
2 |
16 |
35.9 |
2.2 |
6.3 |
0 |
|
3 |
16 |
36.2 |
2.3 |
6.3 |
0 |
|
4 |
16 |
35.5 |
2.4 |
33.3 |
0 |
|
5 |
16 |
29.6 |
2.0 |
6.7 |
0 |
|
6 |
16 |
30.7 |
1.9 |
6.3 |
0 |
|
7 |
16 |
29.6 |
1.9 |
20.0 |
0 |
|
8 |
15 |
28.4 |
1.9 |
40.0 |
0 |
|
9 |
15 |
22.6 |
1.5 |
26.7 |
0 |
|
10 |
16 |
22.7 |
1.4 |
25.0 |
0 |
|
11 |
16 |
28.4 |
1.8 |
43.8 |
31.3 |
|
12 |
16 |
27.6 |
1.7 |
68.8 |
0 |
|
Total |
188 |
30.9 |
2.0 |
24.6 |
2.6 |
Table 5Field test of panicle blast in disease prone experimental plots for 3 years from 2009 to 2011.
Table 5
|
Varieties |
Icheon |
Jaecheon |
Yaesan |
Iksan |
Average |
Rangez)
|
|
Hwaseong |
3.9 |
4.5 |
3.8 |
6.6 |
4.7 |
0.0 – 18.7 |
|
Ilpum |
7.2 |
9.2 |
0.8 |
5.5 |
5.7 |
0.0 – 25.0 |
|
Dongjin |
0.9 |
0.0 |
1.4 |
0.0 |
0.6 |
0.0 – 4.3 |
|
Hwayeong |
1.3 |
0.1 |
3.6 |
0.1 |
1.3 |
0.0 – 10.8 |
|
Saeilmi |
1.9 |
0.5 |
0.0 |
0.1 |
0.6 |
0.0 – 3.3 |
References
- Babujee L, Gnanamanickam SS. 2000. Molecular tools for characterization of the rice blast pathogen (Magnaporthe grisea) population and molecular marker-assisted breeding for disease resistance. Curr Sci. 78: 248-257.
- Bonman JM. 1992. Durable resistance to rice blast disease - environmental influences. Euphytica. 63: 115-123.
- Cho YC, Kwon SJ, Choi IS, Lee SK, Jeon JS, Oh MK, Roh JH, Hwang HG, Yang SJ, Kim YG. 2007. Identification of major blast resistance genes in Korean rice varieties (Oryza sativa L.) using molecular markers. J Crop Sci Biotechnol. 10: 265-276.
- Fujii K, Hayano-Saito Y, Sugiura N, Hayashi N, Saka N, Tooyama T, Izawa T, Shumiya A. 1999. Gene analysis of panicle blast resistance in rice cultivars with rice stripe resistance. Breed Res. 1: 203-210.
- Fujii K, Hayano-Saito Y, Saito K, Sugiura N, Hayashi N, Tsuji T, Izawa T, Iwasaki M. 2000. Identification of a RFLP marker tightly linked to the panicle blast resistance gene, Pb1, in rice. Breed Sci. 50: 183-188.
- Fujii K, Hayano-Saito Y. 2007. Genetics of durable resistance to rice panicle blast derived from an indica rice variety Modan. Jpn J Plant Sci. 1: 69-76.
- Han SS, Ryu JD, Shim HS, Lee SW, Hong YK, Cha KH. 2001. Breakdown of resistance of rice cultivars by new race KI1117a and race distribution of rice blast fungus during 1999–2000 in Korea. Plant Dis. 7: 86-92.
- Hayashi K, Kato T, Hayano-Saito Y. 2004. Delimitation of chromosomal location of rice panicle blast gene, Pb1, by the analysis using with DNA markers. Jpn J Phytopathol. 70: 193.
- Hayashi N, Inoue H, Kato T, Funao T, Shirota M, Shimizu T, Kanamori H, Yamane H, Hayano-Saito Y, Matsumoto T, Yano M, Takatsuji H. 2010. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J. 64: 498-510.
- Jairin J, Teangdeerith S, Leelagud P, Kothcharerk J, Sansen K, Yi M, Vanavichit A, Toojinda T. 2009. Development of rice introgression lines with brown planthopper resistance and KDML105 grain quality characteristics through marker-assisted selection. Field Crops Res. 110: 263-271.
- Jena KK, Mackill DJ. 2008. Molecular markers and their use in marker-assisted selection in rice. Crop Sci. 48: 1266-1276.
- Jeon JS, Chen D, Yi GH, Wang GL, Ronald PC. 2003. Genetic and physical mapping of Pi 5(t), a locus associated with broad spectrum resistance to rice blast. Theor Appl Genet. 269: 280-289.
- Juliano BO. 1971. A simple assay for milled rice amylose. Cereal Sci Today. 16: 334-336.
- Kang HJ, Kim HS, Nam JK, Lee YT, Lee SY, Kim SD. 2003. Mapping of RFLP Markers Linked to Bacterial Blight Resistant Genes (Xa-1, Xa-3) in Rice. Korean J of Crop Sci. 48: 419-423.
- Khush GS, Brar DS. 1991. Genetics of resistance to insects in crop plants. Adv Agron. 45: 223-274.
- Kwon SJ, Ahn SN, Hong HC, Cho YC, Suh JP, Kim YG, Kang KH, Han SS, Choi HC, Moon HP, Hwang HG. 2002. Identification of DNA markers linked to resistance genes to rice blast (Pyricularia grisea Sacc.). Korean J Breed. 34: 105-110.
- Kwon SW, Cho YC, Kim YG, Suh JP, Jeung JU, Roh JH, Lee SK, Jeon JS, Yang SJ, Lee YT. 2008. Development of near-isogenic japonica rice lines with enhanced resistance to Magnaporthe grisea. Mol Cells. 25: 407-416.
- Lee EW, Park SZ. 1979. Interpretation on the epidemic outbreak of rice blast disease in Korea, 1978. Korean J Crop Sci. 24: 1-10.
- Lee JH, Yeo US, Kwak DY, Park DS, Oh BG, Ku YC, Kim HY, Sohn JK. 2005. QTL analysis for ripening traits of BPH resistant backcross inbred lines in rice. Korean J Breed Sci. 37: 172-177.
- Liu G, Lu G, Zeng L, Wang GL. 2002. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol Genet Genomics. 267: 472-480.
- McCouch SR, Teytelman L, Xu YB, Lobos KB, Clare K, Walton M, Fu BY, Maghirang R, Li ZK, Xing YZ, Zhang QF, Kono I, Yano M, Fjellstom R, Declerck G, Scheider D, Cartinhour S, Ware D, Stein L. 2002. Development and mapping of 2240 new SSR markers of rice (Oryza sativa L.). DNA Res. 9: 199-207.
- RDA(Rural Development Administration).2003. Manual for standard evaluation method in agricultural experiment and research. Suwon (Korea): RDA. pp. 838.
- Roh JH, Cho YC, Kim YG, Oh IS, Lee JY, Han SS. 2007. Blast Resistance of High Quality Rice Cultivars. Korean J Bred Sci. 39: 288-295.
- Ryu JD, Yeh WH, Han SS, Lee YH, Lee EJ. 1987. Regional and annual fluctuation of races of Pyricularia oryzae during 1977–1985 in Korea. Korean J Plant Pathol. 3: 174-179.
- Skamnioti P, Gurr SJ. 2009. Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol. 27: 141-150.
- Sirithunya P, Tragoonrung S, Vanavichit A, Pa-In N, Vongsaprom C, Toojinda T. 2002. Quantitative trait loci associated with leaf and neck blast resistance in recombinant inbred Line population of rice (Oryza Sativa). DNA Research. 9: 79-88.
- Suh JP, Roh JH, Cho YC, Han SS, Kim YG, Jena KK. 2009. The Pi40 gene for durable resistance to rice blast and molecular analysis of Pi40-advanced backcross breeding lines. Phytopathology. 99: 243-250.
- Suh JP, Yang SJ, Jeung JU, Pamplona A, Kim JJ, Lee JH, Hong HC, Yang CI, Kim YG, Jena KK. 2011. Development of elite lines conferring Bph18 gene-dereived resistance to brown planthopper (BPH) by marker-assisted selection and genome-wide background analysis in japonica rice (Oryza sativa L.). Field Crops Res. 120: 215-222.
- Wongsaproma C, Sirithunyab P, Vanavichitc A, Pantuwand G, Jongdeee B, Sidhiwongf N, Lanceras-Siangliwa J, Toojindaa T. 2010. Two introgressed quantitative trait loci confer a broad-spectrum resistance to blast disease in the genetic background of the cultivar RD6 a Thai glutinous jasmine rice. Fields Crop Res. 119: 245-251.