Abstract
Plants have adapted the ability to respond to various abiotic stresses such as high salinity, osmotic stress, high and low temperatures, and drought in order to survive. Small heat shock proteins (sHsps) play important and extensive roles in plant defenses against abiotic stresses. Herein, we cloned an sHsp gene from the rice, which we named Oshsp16.9 based on the molecular weight of the protein. Real-time PCR analysis showed that expression of the Oshsp16.9 gene was rapidly and strongly induced by stresses including high-salinity (250 mM NaCl), osmotic stress (300 mM mannitol), 100 μM ABA, cold (4°C) and heat (45°C). Subcellular localization assay indicated that Oshsp16.9 was localized specifically in the cytoplasm. In addition, overexpression of Oshsp16.9 in rice conferred tolerance of transgenic plants to salt and drought stress. Taken together, these results suggest that the Oshsp16.9 gene is an important determinant of stress response in plants.
-
Key words: Abiotic stress, Heat shock protein, Heat stress, Salt stress, Drought stress, ABA treatment
INTRODUCTION
Plants have adapted the ability to respond to various abiotic stresses such as high salinity, osmotic stress, high and low temperatures, and drought in order to survive. In response to high temperatures and other abiotic stresses, all organisms universally respond by accumulating heat shock proteins (HSPs) (
Vierling, 1991). These HSPs are mainly involved in peptide-folding, protein assembly and transport, protection against irreversible protein denaturation, maintaining proteins in the normal folding state, and promoting the degradation of mis-folded proteins under various stresses, which can be summed as acting through “chaperone-like” activities
(Krishna et al. 1995;
Keeler et al. 2000;
Sato and Yokoya 2008). Heat-shock proteins are classified on the basis of their approximate molecular weight into the following groups: Heat-shock proteins 100 kDa, i.e. Hsp100 (CIp), Hsp90, Hsp70 (DnaK), Hsp60 (GroEL), and small HSP (sHsps), where the molecular weight ranges from 15 to 42 kDa (
Trent 1996). These sHsps usually make a complex of small subunits wherein the molecular weight ranges from 200 to 800 kDa (
Kim et al. 1998). The sHsps, the most abundant stress-induced proteins, have highly conserved C-terminals, where an α-crystallin domain (ACD) is found (
Vierling 1991;
Parsell and Lindquist 1993). Plants contain a wide array of sHsps that are divided into six classes based on their sequence alignments and immunological cross-reactivity. Three of these classes (CI, CII, and CIII) are located in the cytoplasm or nucleus (
Scharf et al. 2001), while the others exist in the endoplasmic reticulum (
Helm et al. 1993), mitochondria and plastids (
Osteryoung and Vierling 1994). Plants vary greatly in the amount as well as type of Hsps expressed (
Hamilton et al. 1996).
Arabidopsis thaliana is the most highly studied species of plant, wherein the response to heat-shock treatment occurs through the participation of a number of different Hsps (
Swindell et al. 2007). Higher plants are characterized by the presence of at least 20 types of sHsps, but one species could contain as many as 40 (
Vierling 1991). These sHsps are usually undetectable in plant cells under physiological conditions and are induced upon stress and plant tolerance to stress, including drought, salinity, oxidized species, and low temperatures (
Löw et al. 2000;
Hamilton and Heckathorn 2001;
Scharf et al. 2001;
Zhang et al. 2008. It is believed that this diversification and abundance of the sHsps in plants is a reflection of the adaptation of the plant to heat stress (
Waters et al. 1996). Earlier studies demonstrated that OsHsps can act as a protein chaperone, protecting pollen mother cells and tapetal cells against extreme heat (
Murakami et al. 2004;
Zou et al. 2009). To examine further its functioning and molecular mechanism under abiotic stresses, we transformed
Oshsp16.9 into rice as our model system. Our goal was to investigate how the overexpression of
Oshsp16.9 influences plant tolerance to stress, and to improve our overall understanding of the roles played by OsHsps during the stress response.
MATERIALS AND METHODS
Isolation of full-length Oshsp16.9 cDNA
The Oshsp16.9 fragment was identified in the dbEST division of GenBank by the BLAST program using the α-crystallin domain of the molecular chaperone as the probe sequence. The full-length cDNA was isolated by RACE technology with the SMART RACE-PCR kit (Clontech, Palo Alto, CA, USA).
Subcellular localization analysis
The full open reading frame (ORF) of Oshsp16.9 was amplified using the cDNA clone mentioned above as a template. The PCR product was ligated into the pGWB5-GFP vector, resulting in an in-frame fusion protein of the GFP (green fluorescent protein) gene and the Oshsp16.9 ORF. The construct (p35S:GFP-Oshsp16.9) was transformed into onion epidermal cells by particle bombardment using a Biolistic PDS-1000/He gene gun system (BIO-RAD). After incubation of the transformed onion epidermal cells for 24h, the GFP signal was detected by confocal fluorescence microscopy.
Gene expression analysis under abiotic stress
Seeds of the rice (
Oryza sativa) var. Dongjin were grown under controlled conditions, with 28°C day/25°C night temperatures, 14 h light/10 h darkness in Yosida solution (Yosida
et al. 1976) at 75% relative humidity. After 20 days of germination, seedlings were exposed to high-salinity (250 mM), osmotic (15% polyethylene glycol (PEG) 6000, 15% PEG6000 + 250 mM NaCl), dehydration (300mM mannitol), and 100 μM ABA stresses. The plants were harvested after 0 h, 0.25 h, 0.5 h, 1 h, 5 h, 10 h, and 24 h of exposure to stress, frozen in liquid nitrogen, and stored at −80°C until further analysis (
Rabbani et al. 2003). Total RNA was extracted using Trizol reagent (Invitrogen, USA), according to the manufacturer’s protocol. Real-time PCR was conducted using a Bio-RAD I Cycler IQ5 machine. The threshold cycle (Ct) values of triplicate PCR reactions were averaged, and relative quantification of the expression levels was performed using the comparative Ct method (
Livak et al. 2001). The fold change in total RNA (target gene) relative to the reference gene (actin) was determined using the following formula: fold change = 2−ΔΔ
Ct, where ΔΔCt = (Ct
target gene − Ct
actin gene) transgenic plants − (Ct
target gene − Ct
actin gene) wild-type plants. The following primers were designed through the website
http://primer3.ut.ee/ and employed for analysis:
OshspF1 5′-GCGTCAAGAAGGAGGAGGT-3′ and
OshspR1 5′-GTGCCACTTGTCGTTCTTGT-3′;
ActinF 5′-GCAGT-CCAAGAGAGGTATCC-3′ and
ActinR 5′-GTAGAA-GGTGTGATGCCAGA-3′. RT-PCR analysis of various tissues (root, stem, leaf, spike and seed) was conducted using 50ng of total RNA after treatment with
DNase I. The amplification program consisted of an initial step at 42°C for 60 min, followed by 94°C for 5 min, 30 cycles of amplification (94°C for 1 min, 58°C for 1 min, 72°C for 1 min), and a final step at 72°C for 10 min. The amplified fragments were run on 1.2% agarose gels.
The
Oshsp16.9 gene was introduced into the pB2GW7.0 gateway vector for
Agrobacterium-mediated plant transformation. The construct including the
Oshsp16.9 gene was transformed into rice as described by
Toki (2006). Transgenic rice plants were selected in MS medium containing 3 mg/L phosphinotricine (PPT) and analyzed (T
2 generation). For extraction of genomic DNA, rice seedlings were harvested, immediately frozen in liquid N
2, and stored at −80°C. Genomic DNA was extracted from leaves using Nucleon PhytoPure (GE Healthcare, Piscataway, NJ, USA) according to the manufacturer’s protocol. For
35S::Oshsp16.9 transgenic analysis, the PCR primer sets 35SF (5′-CGCACAATCCCACTATCCTT-3′),
OshspR1 (5′-GTGCCACTTGTCGTTCTTGT-3′), and barF (5′-GTCATCAGATTTCGGTGACG-3′), barR (5′-TTATT-GCGCGTTCAAAAGTC-3′) were designed and synthesized. The amplification program consisted of an initial denaturation step at 94°C for 2 min, 30 cycles (94°C for 15 s, 53°C 30 s, 72°C for 1 min), and a final extension step at 72°C 10 min. Total RNA from 14-day-old seedlings was isolated for RT-PCR analysis using the RBC plant RNA extraction kit. For southern blot, DNA aliquots (40 μg) were digested overnight at 37°C with
EcoRI, a site that was not predicted in the
Oshsp16.9 cDNA. The transfer and hybridization were performed similar to that of the Southern blot analysis, according to the procedure of
Yang et al. (2005).
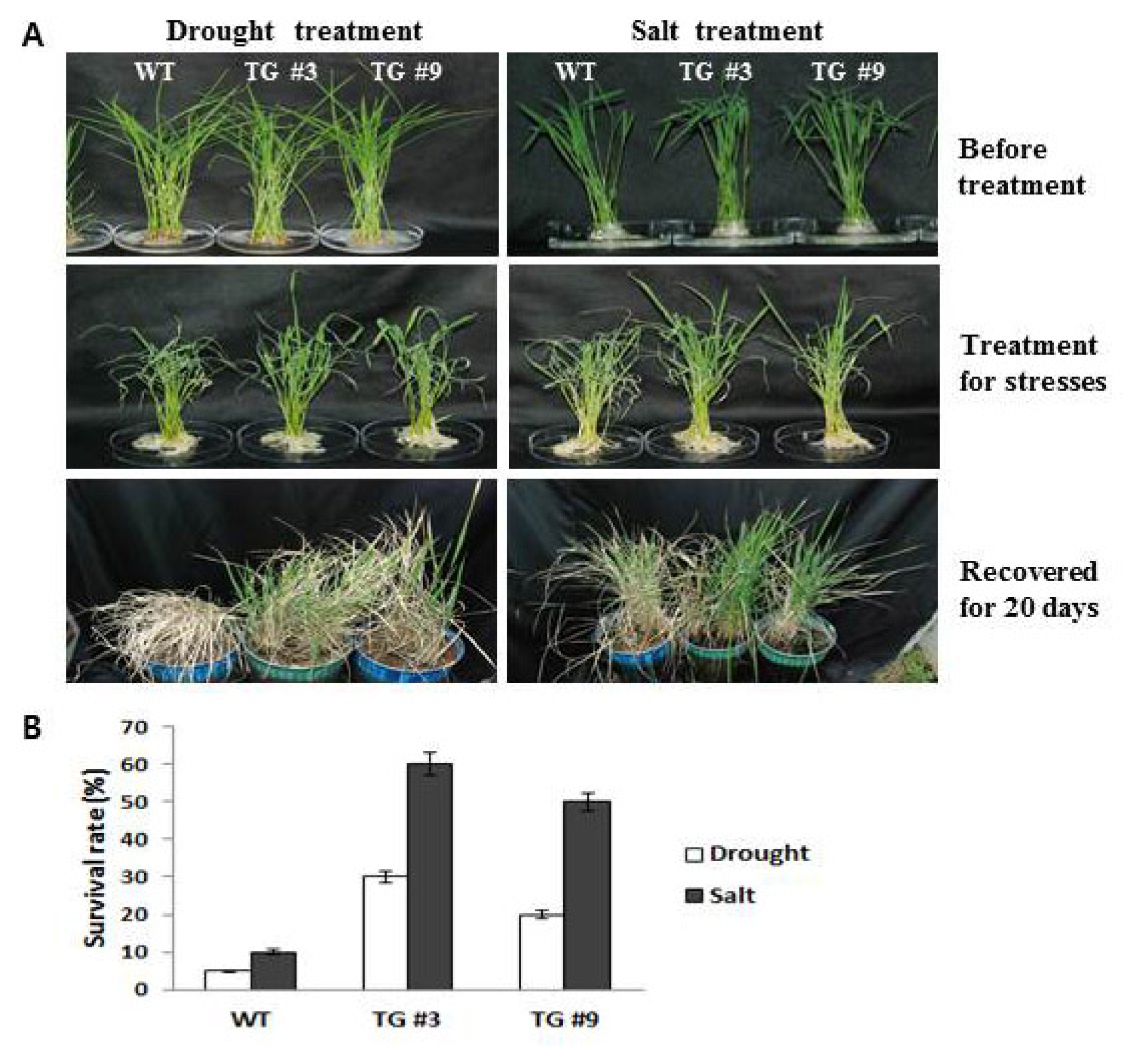
Two homozygous T2 generation transgenic rice plants lines were used in all subsequent abiotic stress assays, with application of 20–30 samples in 2 independent biological experimental replicates, unless otherwise noted. WT and transgenic line seeds were germinated and grown on 1/2 x MS medium in a culture room under a 16:8 h light:dark photoperiod at 24°C. For the drought and salt stress treatments, each of the transgenic seedlings were grown as stated above for 14d. The plants for salt treatment were then transferred to 1/2 x MS media containing 250 mM NaCl, while another treatment plot was exposed to drought conditions. After 1 day of exposure to the stress conditions, plants were photographed and the survival rates were measured after recovery for 20 days. Statistical analyses were performed using the Student’s t-test in Microsoft Excel. Significance was determined for P-values below 0.05.
RESULTS
Cloning and sequence analysis of Oshsp16.9
A partial cDNA of
Oshsp16.9 was identified by a database search of the dbEST division of GenBank using the heat shock protein 16.9 motif as the probe sequence. The full-length cDNA of
Oshsp16.9 was isolated using RACE-PCR, which revealed the gene to be 754 bp in length, consisting of a 117 bp 5′-untranslated region (UTR), a complete ORF of 453 bp encoding a polypeptide of 150 amino acids, and a 3′-UTR of 184 bp (
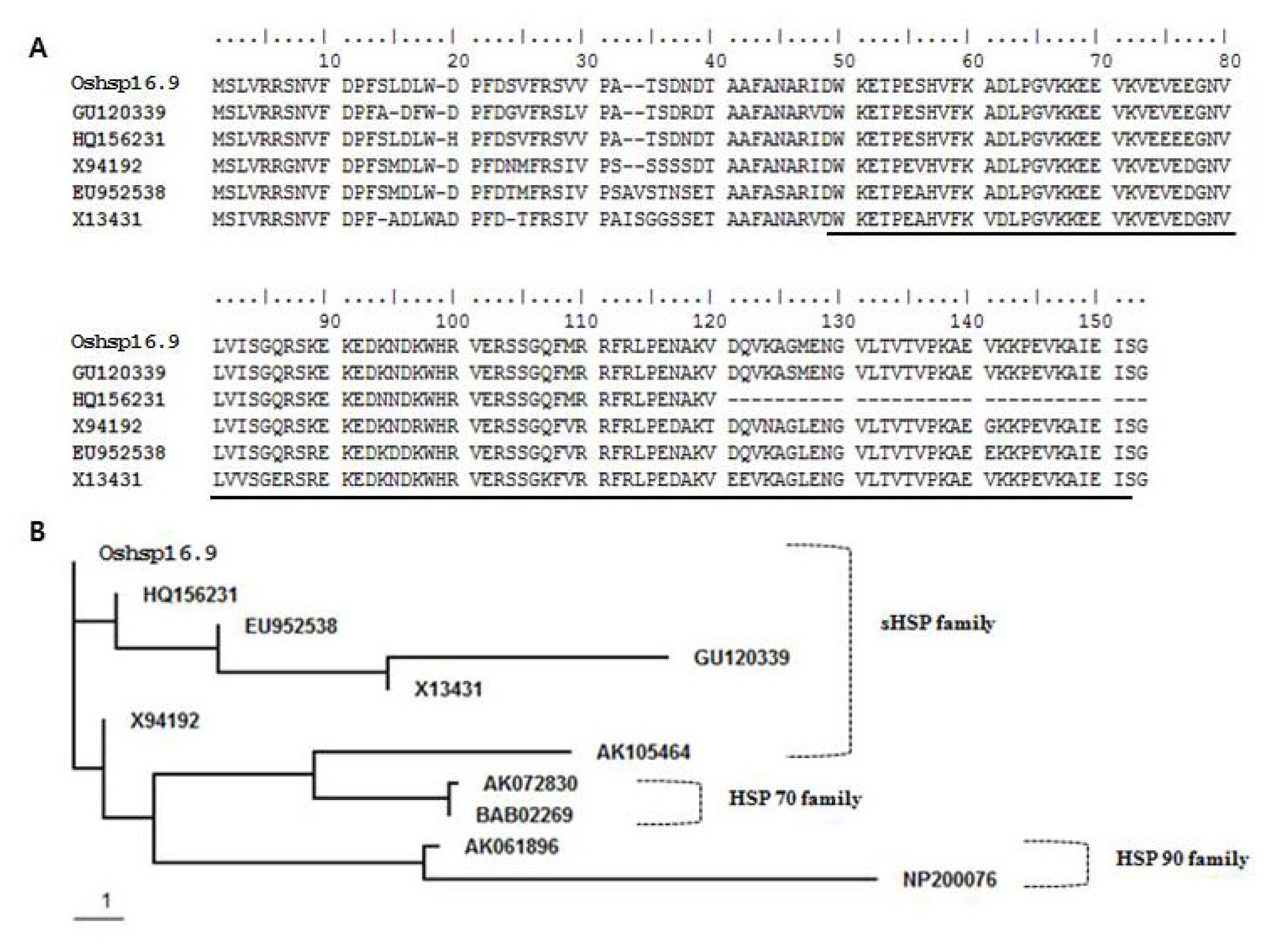
Fig. 1). BLAST protein sequence analysis showed that
Oshsp16.9 exhibited high identity with GU120339 (
Oryza sativa Japonica group) (98%) over the entire protein length, and a similarly high sequence identity, though just over the α-crystallin domain (ACD), with HQ156231 (
Oryza sativa Indica group) (98%), X94192 (
Cenchrus americanus) (98%), EU952538 (
Zea mays) (96%) and X13431 (Triticum aestivum) (94%) (
Fig. 1A). That the ACD shared more than 94% identity suggested origin from a single ancestral gene that most likely underwent recent events of tandem duplication. The conserved α-crystallin domain of sHSPs was used as a query to search the genome of
O. sativa, revealing the existence of 23 genes encoding sHSPs. The phylogenetic relationship of the deduced amino acid sequence of
Oshsp16.9 to sHSP proteins from other organisms was determined using ClustalW analysis from the BioEdit program (
Fig. 1B). In the phylogenetic tree, the 23 sHSPs were divided into three main groups, which showed close relationships with the hsp 70 sequences of other plant species.
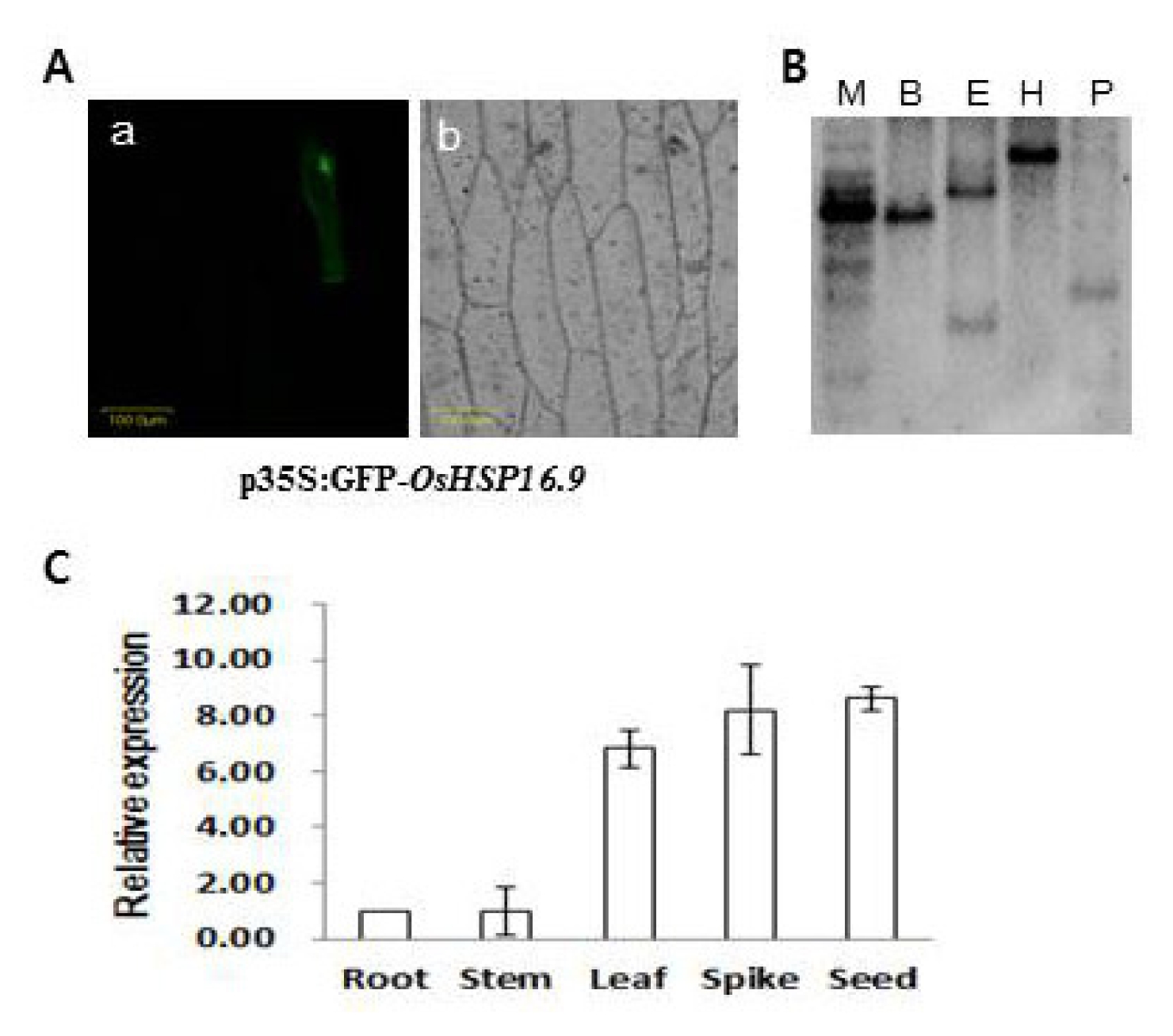
The subcellular localization of
Oshsp16.9 was also analyzed by particle bombardment for transient expression analysis of the
Oshsp16.9–GFP fusion gene in onion epidermal cells. As shown in
Fig. 2A, cells transformed with
Oshsp16.9–GFP displayed fluorescence specifically in the cytoplasm, suggesting that
Oshsp16.9 is a cytoplasmically- localized protein. Southern blot analysis was also carried out using probes specific to the C-terminal region. The probes revealed a single band, suggesting that each member of the
Oshsp16.9 genes existed as a single copy in the rice genome (
Fig. 2B). Expression analysis of the
Oshsp16.9 gene transcript was carried out on various tissues (root, stem, leaf, spike and seed). Semi-quantitative RT-PCR analysis showed broad expression of the
Oshsp16.9 transcript across tissues, with the highest expression in the seed and spike, among reproductive tissues, and in the leaves, among the vegetative tissues (
Fig. 2C).
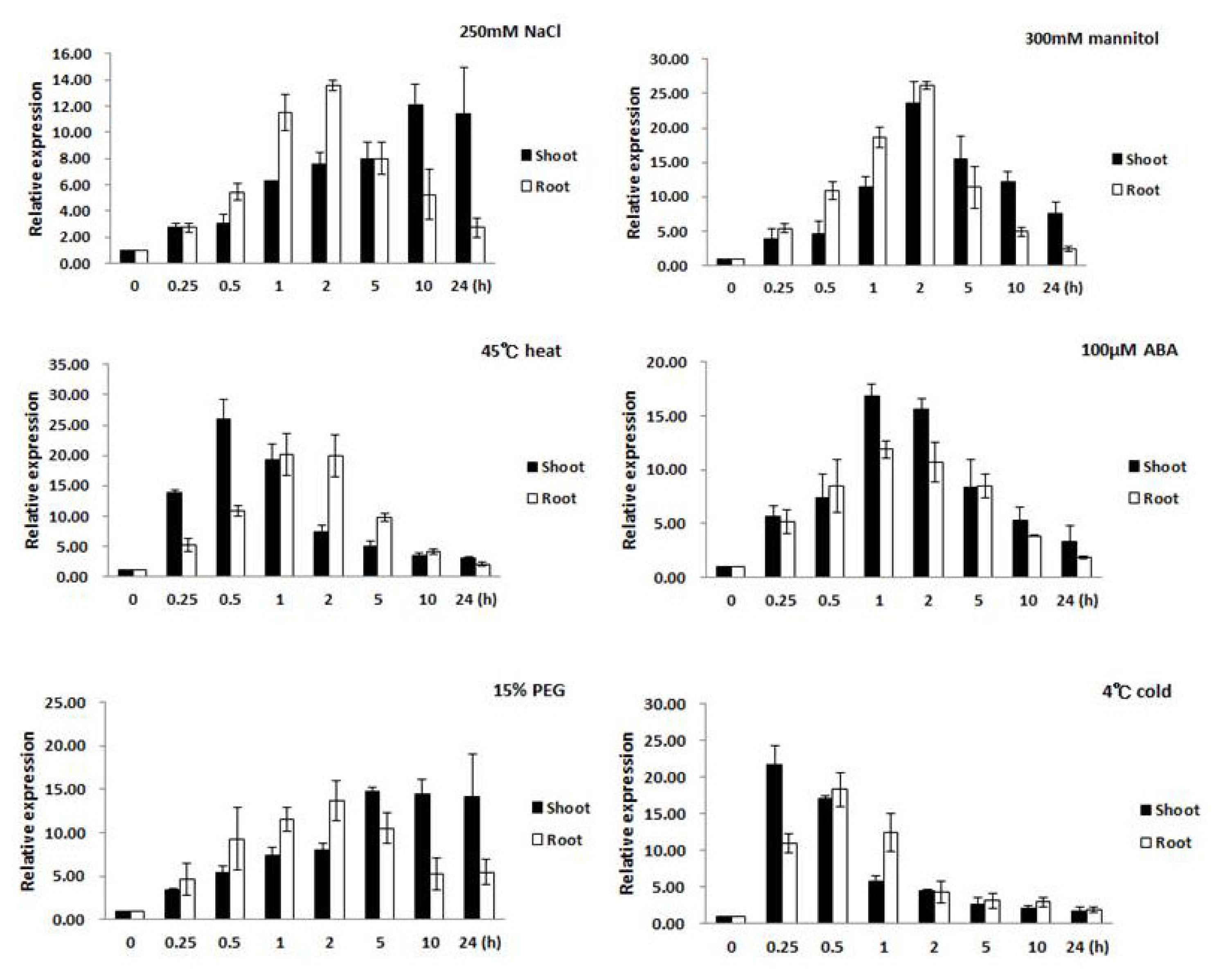
Changes of expression in the root and leaf tissues were examined separately to gain an understanding of the role or contribution each made to the stress response. The results from real-time PCR analysis showed that expression of
Oshsp16.9 was induced under various abiotic stresses in seedlings. In the presence of salt stress (250 mM NaCl), the transcript levels were rapidly induced by several fold after 0.5 hour in the root, reaching a peak after 2 hours, then returning to pre-treatment levels (
Fig. 3). The
Oshsp16.9 transcript also showed rapid induction in leaf tissue, peaking at 14 fold and 22 fold after 0.25 hour of exposure to heat (45°C) and cold (4°C) shock, followed by a gradual decrease in expression to the pre-treatment levels during 24 hours of exposure.
Oshsp16.9 was also examined after treatment with ABA to determine if this stress hormone might be involved in regulating the transcript levels. Exogenous ABA treatment (100 μM) resulted in strong
Oshsp16.9 induction within 0.5 hours, which gradually decreased to the pre-treatment levels over 24 hours. Finally, the roles of dehydration and osmotic stress in the induction of
Oshsp16.9 transcript were also examined. In the presence of dehydration stress (300 mM mannitol), transcript levels were rapidly induced by 5–12 fold after 0.25~0.5 hours in the root, reaching peak induction after 2 hours, then returning to pre-treatment levels. In the case of osmotic stress (15% PEG), increase in the transcript levels in the root by 8 fold after 0.5 hours was observed, which gradually increasing during treatment until 5 hours, then continued to increase, reaching a peak of induction after 24 hours (
Fig. 3).
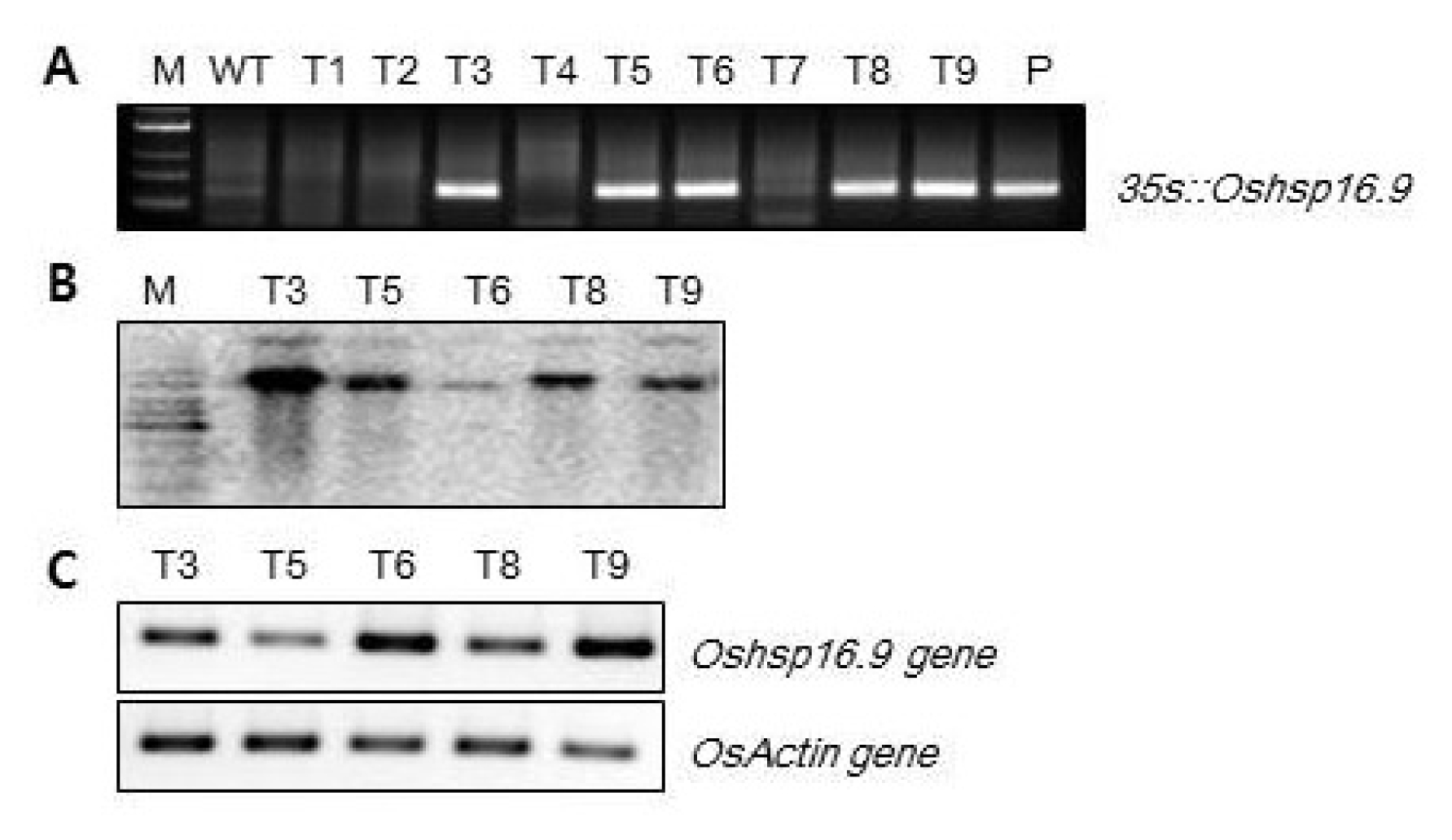
To study the physiological functions of
Oshsp16.9, several
Oshsp16.9-overexpressing transgenic rice plant lines were generating by fusing the full-length cDNA sequence to the CaMV 35S promoter in the pB2GW7.0 gateway vector. Five positive transgenic plant lines, noted as TG #3, #5, #6, #8 and #9, were confirmed by genomic PCR, Southern blot, and RT-PCR to contain a single and highly expressed copy of
Oshsp16.9, not normally found in the rice cultivar (
Fig. 4A–C). No significant gross morphological defects were observed between the transgenic and non-transgenic plants under standard growth conditions. Transgenic lines also showed similar patterns of growth to the WT under non-stressed conditions, although slight overall increases in fresh weight (FW) could be observed for some of the lines (
Fig. 5). The transgenic lines were then examined for responses to various abiotic stresses and compared with the WT, as detailed below. For examination of the survival rate, the seedlings were subjected to stress through exposure to salt (250 mM NaCl) and drought (
Fig. 5A and B). After salt and drought stresses, all seedlings (WT, TG #3, and TG #9) showed significant reduction in the measured survival rate; however, the transgenic lines (TG #3 and TG #9) showed greater tolerance to salt and drought stresses than WT, with significant increases in response to the stress conditions for 1 day. Plants were also examined after given a recovery period of 20 days. After recovery for 20 days, the survival rates of the
Oshsp16.9 transgenic plants (about 20–60%) were much higher than those of the control plants (only 5–10%) (
Fig. 5B). These results suggest that the overexpression of
Oshsp16.9 in transgenic rice enhanced drought tolerance (
P-values of 0.05, respectively, by Student’s
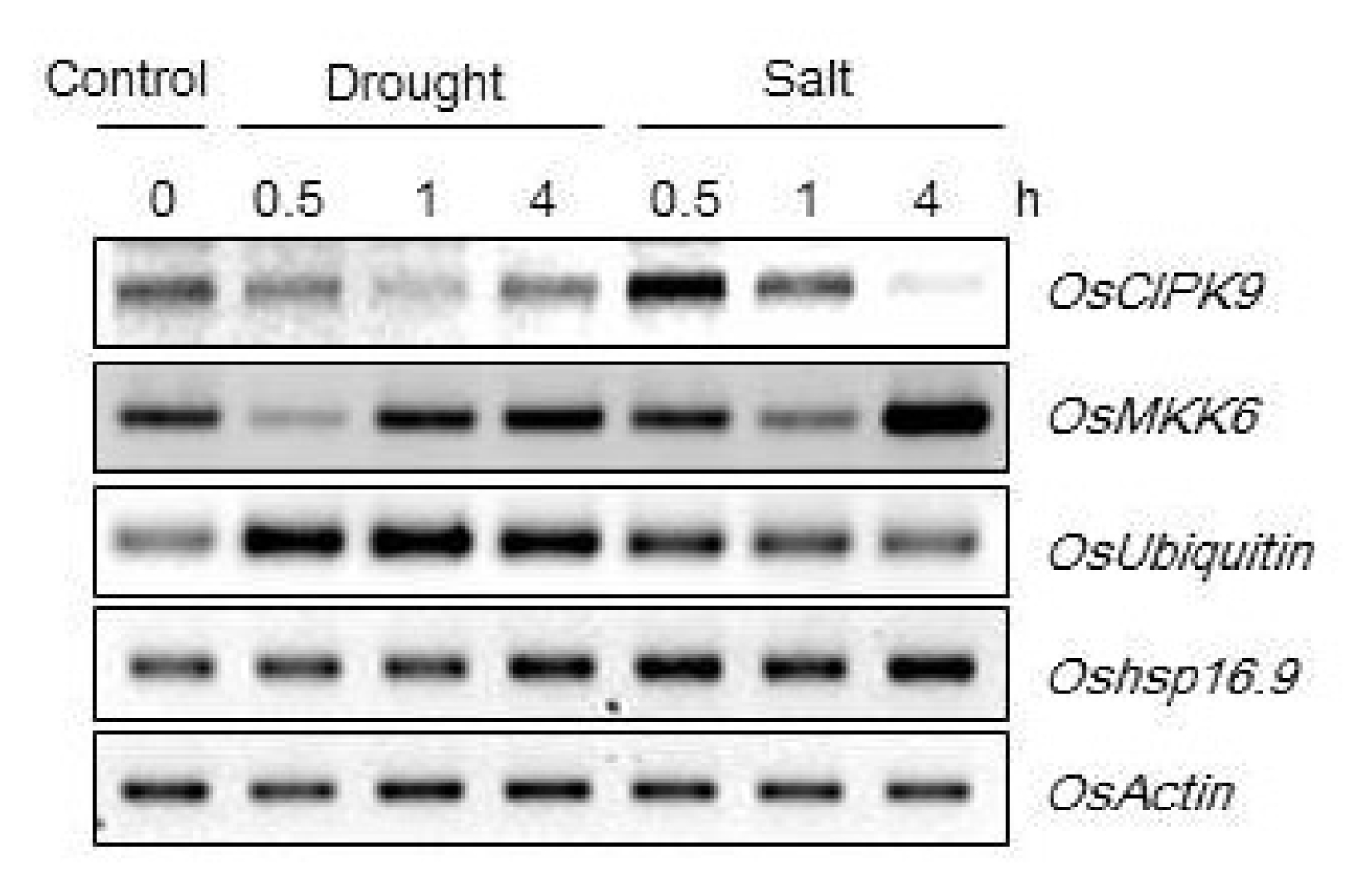
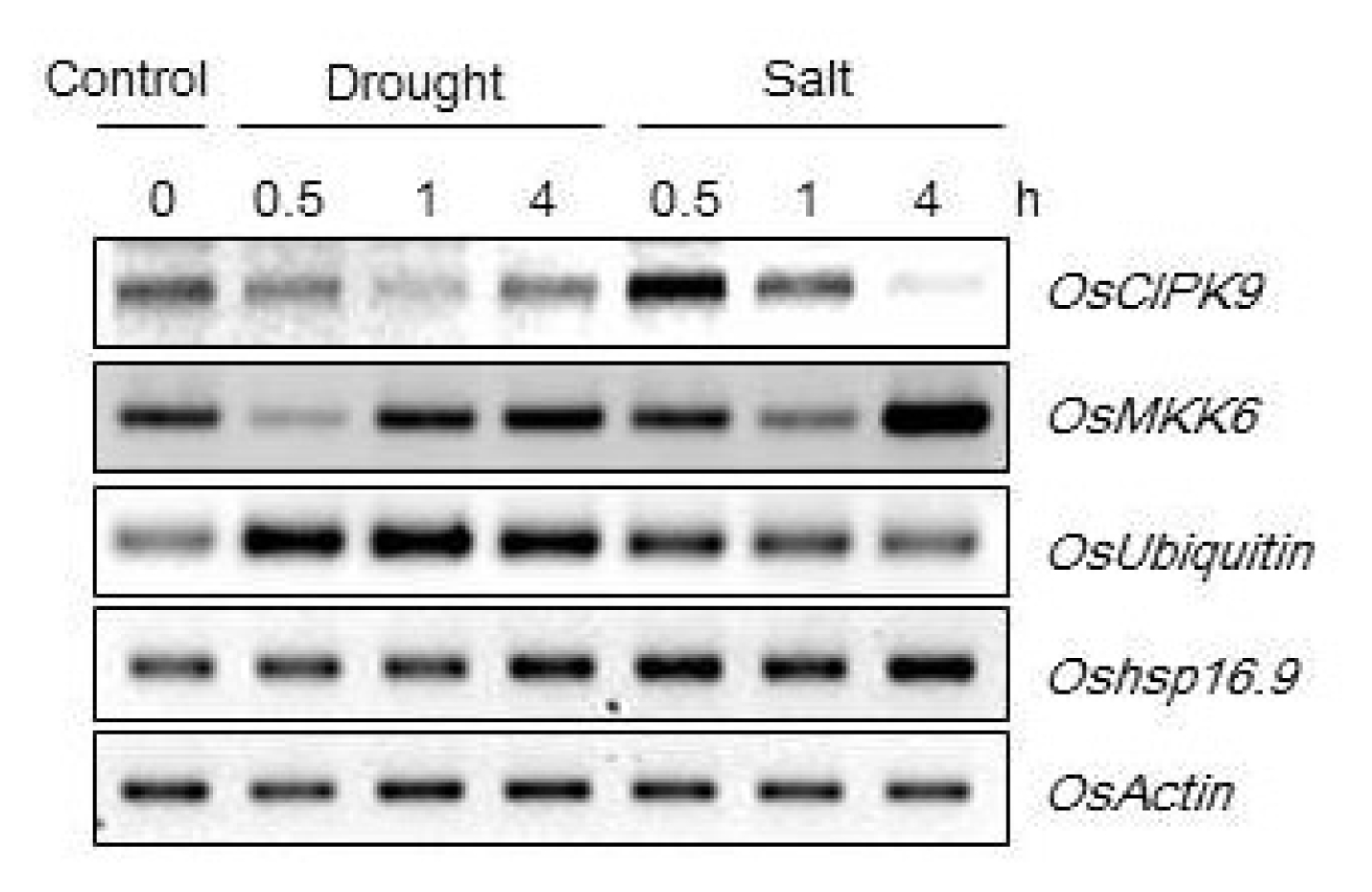
t-test). In addition, the transgenic lines showed high relative expression levels of candidate genes (CBL9, protein kinase, Ubiquitin) under drought and salt stress (250 mM NaCl) (
Fig. 6). In conclusion, this study showed that transgenic rice plants overexpressing
Oshsp16.9 had significant improvement of the tolerance to drought and salinity. Together, these results suggest that
Oshsp16.9 plays a strong role in several stress-related processes, and that its overexpression may lead to the creation of transgenic plants which are tolerant to salt, cold, heat and dehydration stresses. These results suggest that this gene may be useful for the genetic improvement of stress tolerance in rice.
DISCUSSION
Heat shock proteins (HSPs) are conserved in prokaryotes and eukaryotes, and are especially abundant in plants (
Trent, 1996). They are one of the main products expressed in cells in response to stress. HSPs can be divided into six families, including HSP100, HSP90, HSP70, HSP60, small heat shock proteins (sHSPs) and ubiquitin (8.5 kDa) (
Scharf et al. 2001;
Hu et al. 2009). The group of sHSPs, with molecular masses ranging from 15 to 42 kDa, are conserved intra-cellular chaperones for other proteins (
Trent, 1996). They are associated with the nuclei, cytoskeleton, membranes, and both localized and protein-protein interactions, such as folding in organelles. Based on the sequence of the flanking region, we obtained the 453 bp full-length cDNA of a putative sHSP gene in rice by RACE, tentatively designated as
Oshsp16.9 (
Fig. 1). This gene was annotated by the RGAD Rice Genome Annotation Database (
http://rgp.dna.affrc.go.jp/RiceHMM/). Despite the significant amount of data concerning the structure, gene regulation and function of the genes in the plant sHsp family, attributing specific roles to individual proteins in stress tolerance has been difficult (
Mu et al. 2011). Southern blot analysis revealed that the
Oshsp16.9 gene yielded a single band, suggesting that each member of the
Oshsp16.9 genes existed as a single copy in the rice genome (
Fig. 2). Previous studies on
Oryza sativa heat shock protein (OsHSP) genes showed that they exhibited diverse expression in different organs and enhanced the tolerance of transgenic plants to various stresses, including abiotic stress (
Sun et al. 2002;
Zou et al. 2009). In addition, sHSPs are known to function in pollen development and seed germination, as well as resistance to UV damage, enhancement of cell survival against both thermal extremes (50°C and 4°C) and seed and root development (
Sung et al. 2001;
Murakami et al. 2004;
Zou et al. 2009). Expression of the
Oshsp16.9 gene transcript was observed in different parts of the plant, including the root, stem, leaf, spike and seed. Higher expression in the seeds, leaves and spikes was detected than in other tissues of the plant (
Fig. 2). Some sHsps are also activated in response to osmotic and salt stress. The expression of
OsHSP23.7 in rice was reported to increase during salt stress treatment, and the
OsHSP24.1 gene was enhanced following treatment with 10 % PEG (
Zou et al. 2009).
At-HSP17.6A and
At-HSP17.6-II were induced by 0.2 M NaCl and 20 % PEG with similar kinetics in Arabidopsis (
Sun et al. 2002).
OsHsfC1b was induced in
O. sativa roots after 30 min of salt stress, while expression was down regulated in the roots after mannitol treatment (
Ruibal et al. 2013). In the present study, after the seedlings were faced with various stresses, the expression of the
Oshsp16.9 gene was increased by many fold until the maximum time limit, whereat the induction capability reached a peak (
Fig. 3). Many molecular chaperones are stress proteins, and many were originally identified as heat-shock proteins. Molecular chaperones are key components contributing to cellular homeostasis in both optimal and adverse growth conditions. Exogenous ABA treatment demonstrated that the levels of
Oshsp16.9 transcript were higher at the initial stage of induction, but eventually decreased to the pretreatment levels. Exogenous ABA, heavy metals and dehydration stress induces heat tolerance in maize seedlings (
Bonham-Smith et al. 1987). Numerous studies have shown that the plant hormone abscisic acid (ABA) plays a crucial role in controlling the downstream responses which are essential for adaptation to abiotic stress. As part of the osmotic stress response, regulation of gene expression occurs in both an ABA-dependent and ABA-independent manner (
Shinozaki Yamaguchi-Shinozaki, 2000;
Ruibal et al. 2013). Interestingly, the gene
Oshsp16.9 was activated in response to both osmotic and dehydration stresses (
Fig. 3). Individual members of each class of HSPs/chaperones have particular functions, but the co-operation between different HSP/chaperone networks appears to be a central principle of the integrated HSP/chaperone machinery. The transgenic plants, which were transformed to overexpress
Oshsp16.9, showed similar performance in most of the growth parameters under non-stress conditions (
Fig. 4). However, after exposure to salt and drought stresses, the transgenic rice plants showed significant growth differences compared to the WT, confirming that overexpression of
Oshsp16.9 in transgenic rice enhanced tolerance to drought (
Fig. 5). The response of plants under drought and heat stress is composed of suppression of photosynthesis, enhancement of respiration, induction of a large number of defense genes, including genes related to pathogen defense, and changes in genes related to sugar metabolism (
Rizhsky et al. 2002,
Guo et al. 2007). This study showed that transgenic rice overexpressing
Oshsp16.9 had significant improvement of the tolerance to drought and salinity. Together, these results suggest that
Oshsp16.9 plays a strong role in several stress-related processes, and that its overexpression may lead to the generation of transgenic plants with tolerance to salt, cold, heat and dehydration stresses. Our results suggest that this gene is an important determinant of stress response in plants.
Fig. 1A, Sequence alignment of molecular chaperones, including the α-crystallin domains (ACDs) from different proteins. The α-crystallin domains (ACD) were marked by underline. B, Sequence alignment of the known members of the HSP family. Accession numbers are as follows: OsHSP16.9, AK105464, AK072830 and AK061896 (Oryza sativa Japonica group); HQ156231 (Oryza sativa Indica group); EU952538 (Zea mays); GU120339 (Oryza sativa Japonica group); X13431 (Triticum aestivum); X94192 (Cenchrus americanus); BAB02269 and NP200076 (Arabidopsis thaliana).
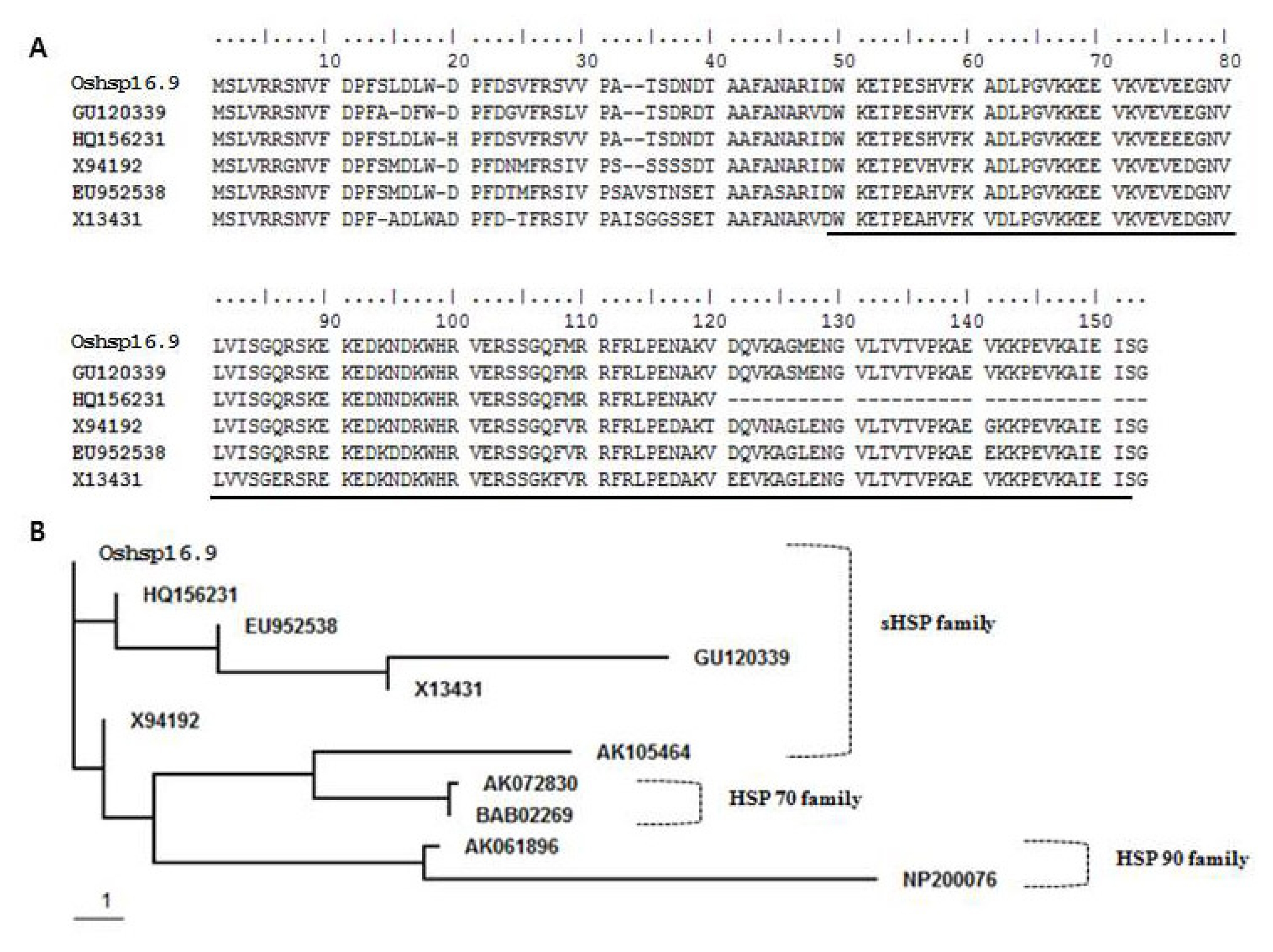
Fig. 2A, Cytoplasmic localization of the Oshsp16.9 gene. p35S:GFP-Oshsp16.9 was transiently expressed in onion epidermal cells. The photograph was taken in the dark field for green fluorescence (a), and under bright light for the morphology of the cell (b). B, Southern blot analysis. The specific region of Oshsp16.9 was used as a probe. Restriction enzymes used were as follows; M: molecular marker, B: BamHI, E: EcoRI, H: HindIII, P: Pst I. C, Relative expression of Oshsp16.9 in organs collected from rice. Data presented are means and standard deviations of 3 independent replicated experiments.
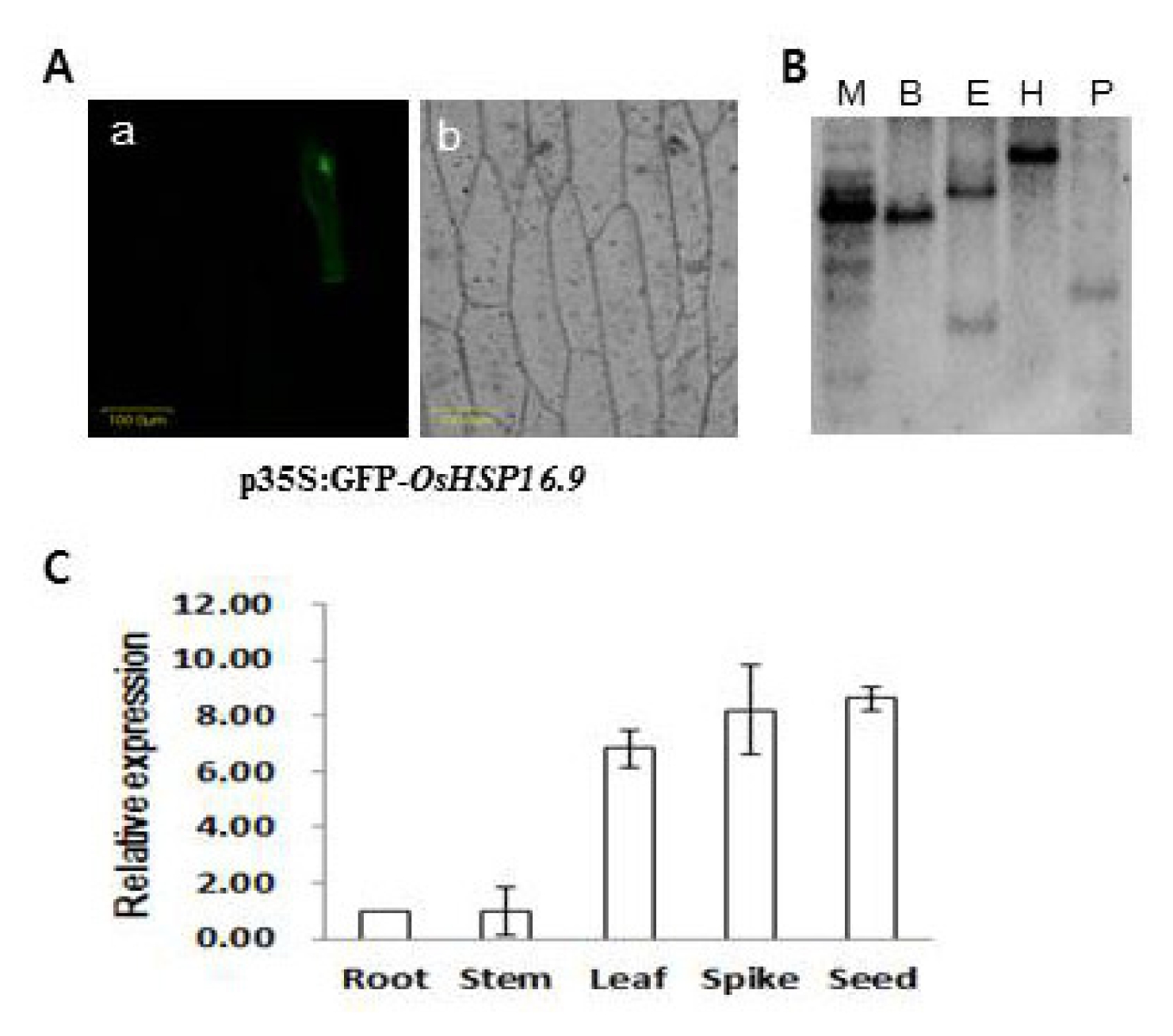
Fig. 3Expression profiles of Oshsp16.9 in shoots and roots of rice after abiotic stress treatments. Expression levels were examined via real-time PCR analysis at various times after treatment with salt (250 mM NaCl), heat (45°C), cold (4°C), 100 μM abscisic acid (ABA), 300mM mannitol and osmotic stress (15% PEG6000) in tissue from 20-day-old plants. Actin was used as a positive control for relative expression for each treatment. Data presented are means ± SD of three independent replicated experiments.
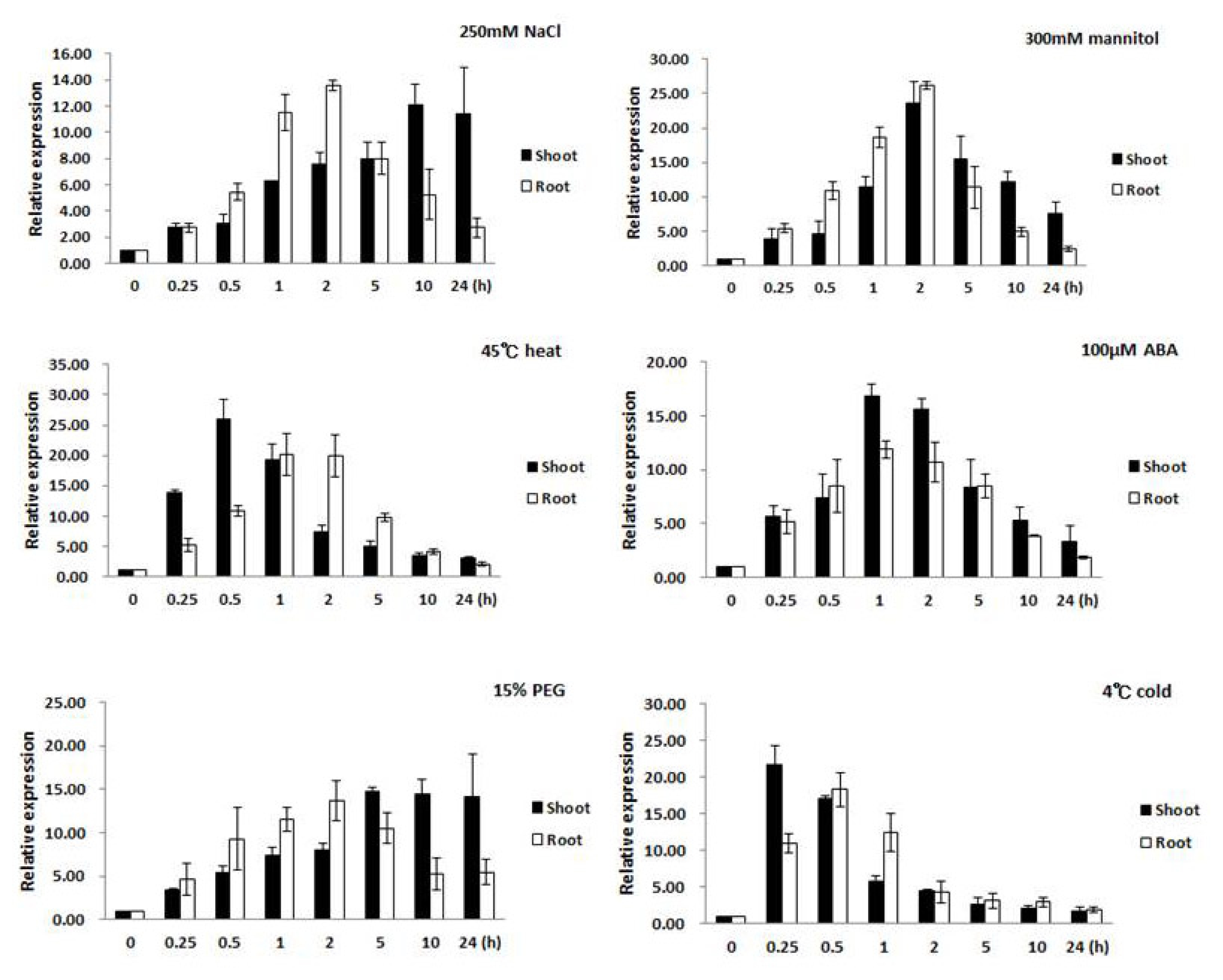
Fig. 4Expression levels of Oshsp16.9-overexpressing lines. A: PCR amplification of transferred Oshsp16.9 genes in transgenic rice lines. The amplification products were separated using a 1% agarose gel. Lane M: 1-kb DNA ladder; Lane WT: wild-type plant; Lanes T1–T9: PCR products generated from the DNA template of independent transgenic lines; and Lane P: PCR product of the vector plasmid that contained Oshsp16.9. B: Southern blot analysis for detection of the Oshsp16.9 gene in transgenic plants. Genomic DNA of five transgenic lines were digested with EcoRI, separated with 0.8% agarose gel electrophoresis and hybridized with a Oshsp16.9 fragment probe. C: Analysis of Oshsp16.9 gene expression in the transgenic rice lines by RT-PCR. As a loading control, the samples were also amplified with specific primers for the rice actin gene.
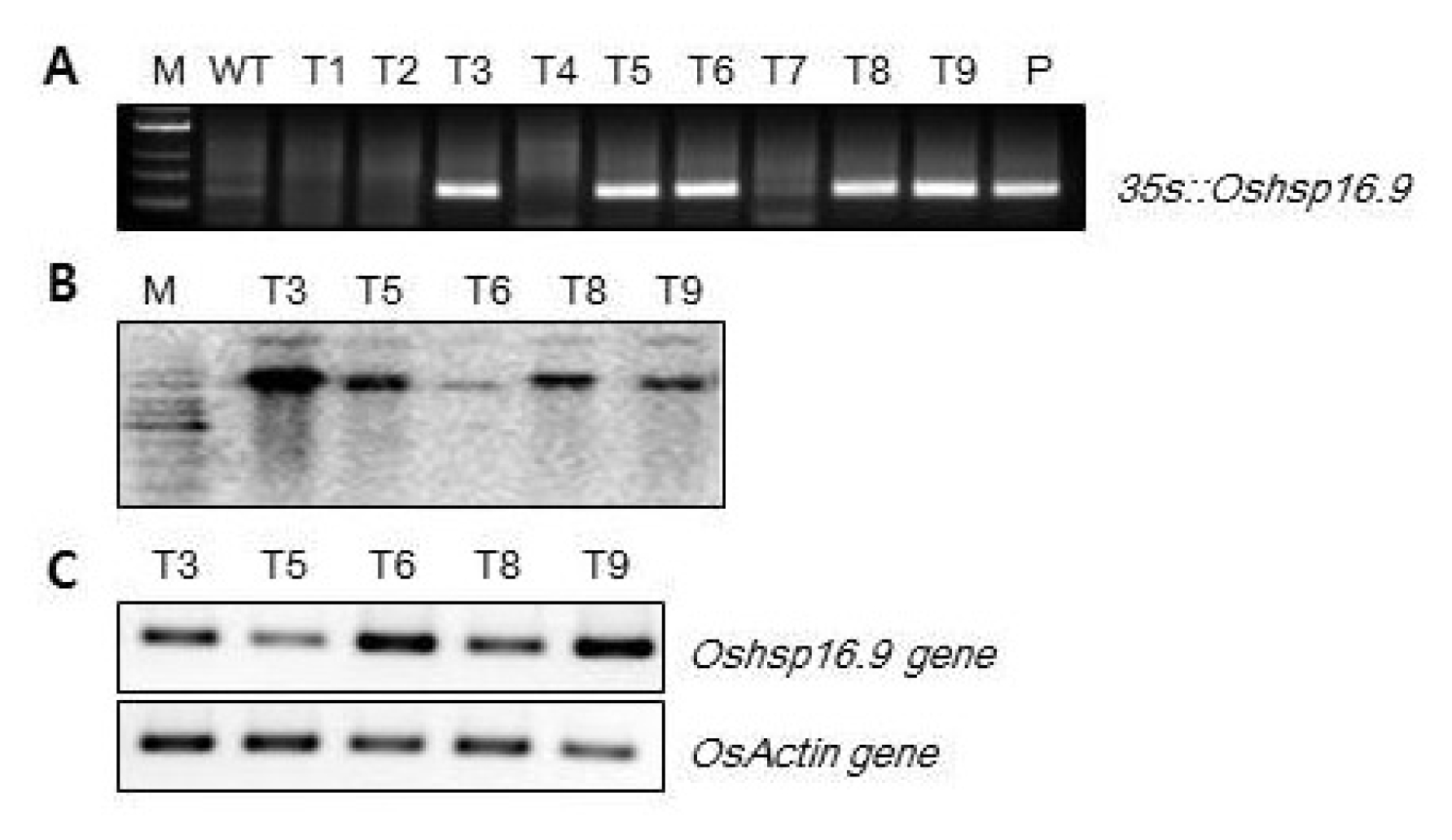
Fig. 5A, Drought and salt tolerance assays of Oshsp16.9 transgenic rice. Three-week-old seedlings were exposed to air conditions for 10 h, and then transferred to normal hydroponic conditions and allowed to recover for 20 days. In addition, three-week-old seedlings were watered with 250 mM NaCl solution for 24 h, and then allowed to recover for 20 days. B, Survival rates of the WT and transgenic rice lines. The data presented are the means ±SE of three independent experiments. The values with significant differences according to t-tests are at the level of P < 0.05.
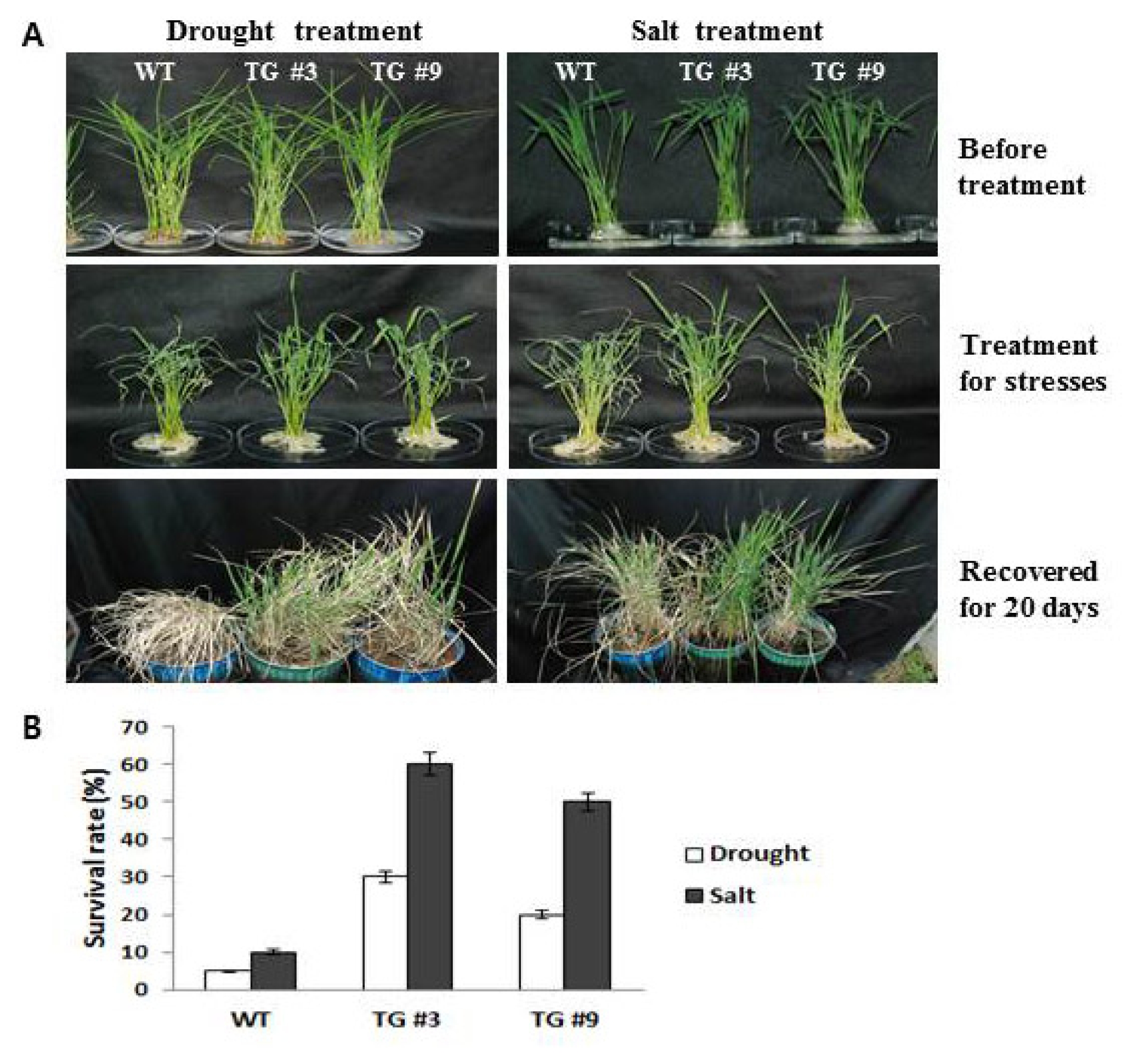
Fig. 6Relative expression levels of candidate genes in Oshsp16.9-overexpressing lines under drought (exposed to air) and salt stresses (250 mM NaCl). Total RNA was extracted from three-week-old rice seedlings.
References
- Bonham-Smith PC, Kapoor M, Bewley JD. 1987. Establishment of thermo tolerance in maize by exposure to stresses other than a heat shock does not require heat shock protein synthesis. Plant Physiol. 85: 575-580.
- Guo SJ, Zhou HY, Zhang XS, Li XG, Meng QW. 2007. Overexpression of CaHSP26 in transgenic tobacco alleviates photo inhibition of PSII and PSI during chilling stress under low irradiance. J Plant Physiol. 164: 126-136.
- Hamilton EW, Heckathorn SA, Downs CA, Schwarz TE, Coleman JS, Hallberg RL. 1996. Heat shock proteins are produced by field-grown naturally occurring plants in the summer in the temperate northeast. US Bulletin of the Ecologic Soc Am. 77: Suppl Part 2. 180. (Abstr.).
- Hamilton EW, Heckathorn SA. 2001. Mitochondrial adaptations to NaCl. Complex I is protected by anti-oxidants and small heat shock proteins, whereas complex II is protected by proline and betaine. Plant Physiol. 126: 1266-1274.
- Helm KW, Lafayete PR, Nago RT, Key JL, Vierling E. 1993. Localization of small heat shock proteins to the higher plant endo membrane system. Mol Cell Biol. 13: 238-247.
- Hu W, Hu G, Han B. 2009. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Sci. 176: 583-589.
- Keeler SJ, Boettger CM, Haynes JG, Kuches KA, Johnson MM, et al. 2000. Acquired thermo tolerance and expression of the HSP100/ClpB genes of Lima Bean. Plant Physiol. 123: 1121-1132.
- Kim KK, Yakota H, Santoso S, Lerner D, Kim R, Kim SH. 1998. Purification, crystallization and preliminary X-ray crystallographic data analysis of a small heat shock protein homolog from Methanococcus jannaschii, a hyperthermophile. J Struct Biol. 121: 76-80.
- Krishna P, Sacco M, Cherutti JF, Hill S. 1995. Cold-induced accumulation of hsp90 transcripts in Brassica napus. Plant Physiol. 107: 915-923.
- Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the DDCt method. Methods. 25: 402-408.
- Low D, Brandle K, Nover L, Forreiter C. 2000. Cytosolic heat-stress proteins Hsp17.7 class I and Hsp17.3 class II of tomato act as molecular chaperones in vivo. Planta. 211: 575-582.
- Mu C, Wang S, Zhang S, Pan J, Chen N, Li X. 2011. Small heat shock protein LimHSP16.45 protects pollen mother cells and tapetal cells against extreme temperatures during late zygotene to pachytene stages of meiotic prophase I in David Lily. Plant Cell Rep. 30: 1981-1989.
- Murakami T, Matsuba S, Funatsuki H, Kawaguchi K, Saruyama H, Tanida M, et al. 2004. Over-expression of a small heat shock protein, sHSP17.7, confers both heat tolerance and UV-B resistance to rice plants. Mol Breeding. 13: 165-175.
- Osteryoung KW, Vierling E. 1994. Dynamics of small heat shock protein distribution within the chloroplasts of higher plants. J Biol Chem. 269: 28676-28682.
- Parsell DA, Lindquist S. 1993. The function of heat-shock proteins in stress tolerance: degradation and reactivation of proteins. Annu Rev Genet. 27: 437-96.
- Rabbani MA, Maruyama K, Abe H, Khan MA, Katsura K, Ito Y, Yoshiwara K, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. 2003. Monitoring expression profiles of rice (Oryza sativa L.) genes under cold, drought and high-salinity stresses, and ABA application using both cDNA microarray and RNA gel blot analyses. Plant Physiology. 133: 1755-1767.
- Rizhsky L, Liang H, Mittler R. 2002. The Combined Effect of Drought Stress and Heat Shock on Gene Expression in Tobacco. J Plant Physiology. 1143-1151.
- Ruibal C, Alexandra C, Valentina C, László S, Sabina V. 2013. Recovery from heat, salt and osmotic stress in Physcomitrella patens requires a functional small heat shock protein PpHsp16.4. BMC Plant Biology. 174: 2-18.
- Sato Y, Yokoya S. 2008. Enhanced tolerance to drought stress in transgenic rice plants overexpressing a small heat-shock protein smHSPs17.7. Plant Cell Reports. 27: 329-334.
- Scharf KD, Siddique M, Vierling E. 2001. The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing alpha-crystallin domains (Acd proteins). Cell Stress Chaperones. 6: 225-237.
- Shinozaki K, Yamaguchi-Shinozaki K. 2000. Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr Opin Plant Biol. 3: 217-223.
- Sun W, Motangu MV, Verbruggen N. 2002. Small heat shock proteins and stress tolerance in plants. Biochim Biophys Acta. 1577: 1-9.
- Sung DY, Kaplan F, Guy CL. 2001. Plant Hsp70 molecular chaperones: protein structure, gene family, expression and function. Physiol Plantarum. 113: 443-451.
- Sung DY, Vierling E, Guy CL. 2001. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 126: 789-800.
- Swindell M Huebner, Weber AP. 2007. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genomics. 8: 125
- Toki S, Hara N, Ono K, Onodera H, Tagiri A, Oka S, Tanaka H. 2006. Early infection of scutellum tissue with Agrobacterium allows high-speed transformation of rice. Plant J. 47: 969-976.
- Trent JD. 1996. A review of acquired thermo tolerance, heat-shock proteins and molecular chaperones in Archaea Fems Microbiol. Rev. 18: 249-258.
- Vierling E. 1991. The roles of heat shock proteins in plants. Annu Rev Plant Physiol Plant Mol Biol. 42: 579-620.
- Waters ER, Lee GJ, Vierling E. 1996. Evolution, structure and function of the small heat shock proteins in plants. J Exp Bot. 47: 325-338.
- Yang L, Ding J, Zhang C, Jia J, Weng H, Liu W, Zhang D. 2005. Estimating the copy number of transgenes in transformed rice by real-time quantitative PCR. Plant Cell Rep. 23: 759-763.
- Zhang JH, Wang LJ, Pan QH, Wang YZ, Zhan JC, Huang WD. 2008. Accumulation and subcellular localization of heat shock proteins in young grape leaves during cross- adaptation to temperature stresses. Scientia Horticulturae. 117: 231-240.
- Zou J, Liu A, Chen X, Zhou X, Gao G, Wang W, et al. 2009. Expression analysis of nine rice heat shock protein genes under abiotic stresses and ABA treatment. J Plant Physiol. 166: 851-856.