Abstract
Most eukaryotic organisms display specialized cellular and behavioral oscillations with a period of approximately 24 hours, which are called circadian rhythms. The biological clock generates a rhythm that conveys temporal information over a day. Through this system, most eukaryotic organisms appropriately respond to daily or seasonal environmental changes by regulating their physiology and development in a time-dependent manner, conferring the organism with an adaptive advantage. In plants, the endogenous timing system also controls many important physiological processes including flower opening, hormone synthesis, metabolic pathways and gene expression so that these sessile species may survive efficiently in changing environments. Temperature compensation (TC) is one of the defining features of the clock mechanism. Under this mechanism, the pace of the clock, or period, remains stable over a broad range of physiologically relevant temperatures, which is unlikely to happen in other biochemical reactions. Thus, this mechanism allows organisms to sustain their ordinary life in various thermal environments by providing an accurate measure of the passage of time, regardless of the ambient temperature. Considering the current global climate changes our planet is undergoing, understanding the fundamental mechanism underlying TC cannot be overemphasized. In this review, we discuss the molecular organization of the plant circadian clock and the concept of TC, as well as the significance of plant TC in conferring fitness under the current increasing thermal environments.
-
Key words: Circadian clock, Temperature compensation, Global warming, Heat tolerance, Crop production
The plant circadian clock
The daily rotation of the earth on its axis and its annual revolution around the sun confer recurrent and rhythmic but predictable environmental changes (
McClung, 2011). To face such challenges actively, most organisms have evolved the capability to measure time. The circadian clock is the time measuring system that establishes an endogenous rhythm with a 24 hr period, called a circadian rhythm (
Jolma et al., 2010;
McClung, 2011). The rhythmic oscillation is persistent (self-sustaining), able to be synchronized to ambient environments (entrainable), and is stable in physiologically-ranged thermal environments (temperature compensated) (
de Montaigu et al., 2010;
McClung, 2011;
McWatters and Devlin, 2011).
In plants, about 10,000 transcripts are known to be regulated by the clock system (
de Montaigu et al., 2010). Thus, many biological processes, including leaf movement, flower opening, hormone synthesis, metabolic pathways and gene expression, are rhythmic, displaying peaks and troughs in their activity during particular times of the day. As such, this clock system, present in plants and many other eukaryotes, confers fitness benefits, efficiently coordinating many important biological functions by integrating and anticipating the daily and seasonal bases of environmental changes (
de Montaigu et al., 2010;
McWatters and Devlin, 2011).
The clock mechanism
The rhythmic oscillation by the circadian clock follows a common molecular signature in eukaryotes, including fungi, flies, mammals, and plants, wherein the mechanism is underpinned by transcription-translation feedback loops (TTFLs)(
Baker et al., 2011;
Jolma et al., 2010;
McClung, 2011;
McWatters and Devlin, 2011). Arabidopsis circadian oscillators, like those of many other eukaryotes, are based on multiple-interlocked TTFLs consisting of three main ones (
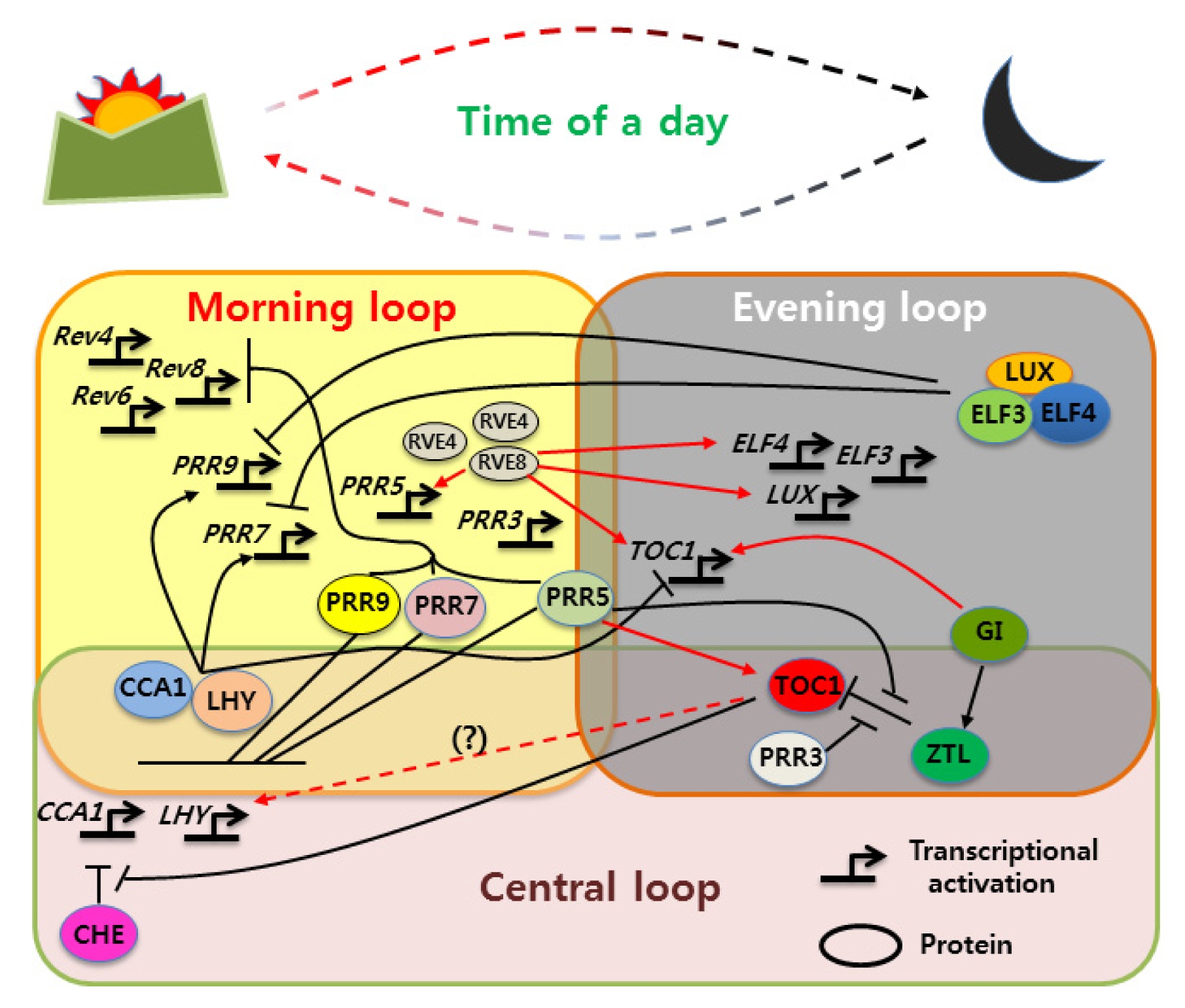
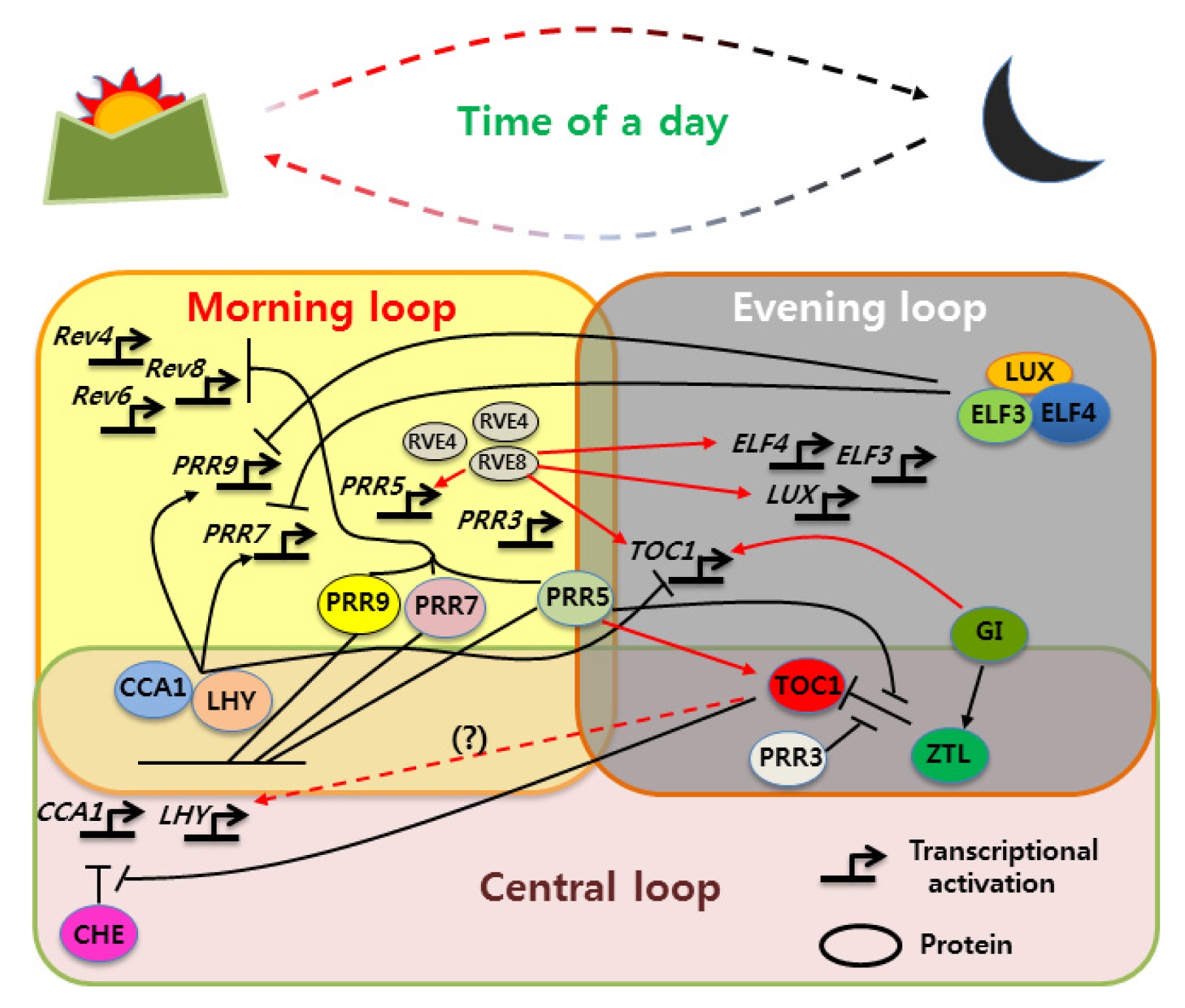
Figure 1) (
Harmer, 2009;
Locke et al., 2006;
McClung, 2011). These includes a central loop and two interlocked loops termed “morning” and “evening,” based on the times of day at which the loop components are maximally expressed (
Alabadi et al., 2001;
Locke et al., 2006). To date, more than 20 clock or clock-associated components have been identified in the Arabidopsis oscillator.
The central loop consists of a pseudo-response regulator (PRR), TIMING OF CAB EXPRESSION 1 (TOC1), and two single Myb domain transcription factors (TF): CIRCADIAN CLOCK ASSOCIATED1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) (
Alabadi et al., 2002;
Mizoguchi et al., 2002). CCA1 and LHY are negative regulators of
TOC1, inhibiting its expression until the evening (
Alabadi et al., 2001;
Alabadi et al., 2002) (
Figure 1). CCA1 and LHY physically interact with each other and bind to a motif in the TOC1 promoter, called the evening element (EE), to inhibit
TOC1 expression. It was recently reported that TOC1 is a DNAbinding transcriptional repressor of
CCA1 and
LHY (
Gendron et al., 2012;
Huang et al., 2012). CCA1 HIKING EXPEDITION (CHE), a TCP transcription factor, might coordinate the link between TOC1 and the regulation of
CCA1 in the central loop (
Pruneda-Paz et al., 2009) (
Figure 1). CHE, as another transcriptional repressor of
CCA1, recruits TOC1 to a transcriptional complex at the CCA1 promoter. However, the antagonistic interaction between those two components results in the indirect activation of
CCA1 (
Pruneda-Paz, 2009).
In the “morning” loop, CCA1 and LHY are positive regulators of two TOC1 relatives,
PRR7 and
PRR9 (
Harmer and Kay, 2005;
Mizuno and Nakamichi, 2005). Conversely, PRR5, PRR7 and PRR9 bind to the CCA1 and LHY promoters to repress their expression (
Nakamichi et al., 2010), which requires a co-repressor, encoded by members of the TOPLESS/TOPLESS-RELATED (TPL/ TPR) gene family (
Wang et al., 2013). The peak binding of the three proteins occurs sequentially from early morning (PRR9) through midday (PRR7) to late afternoon/early night (PRR5), resulting in the repression of
CCA1 and
LHY expression (
Nakamichi et al., 2010).
Although many clock genes of the evening phase, including
TOC1,
PRR3,
PRR5, ZEITLUPE (ZTL),
GIGANTEA (
GI) and others are assigned in the evening loop, the negative feedback mechanism by which they are regulated is still elusive (
Fujiwara et al., 2008;
Kim et al., 2007;
McClung, 2011). Instead, sophisticated post-translational mechanisms have been characterized in the evening loop (
Fujiwara et al., 2008;
Kim et al., 2007;
McClung, 2011). ZTL, an F-box-containing protein which is part of a Skp/Cullin/F-box (SCF) E3 ubiquitin ligase complex, directly targets both TOC1 and PRR5, leading to their degradation via the proteosome pathway (
Fujiwara et al., 2008;
Kiba et al., 2007;
Mas, 2003). Due to the light-sensing ability of ZTL, its involvement in the degradation of TOC1 and PRR5 is regulated by light (
Fujiwara et al., 2008;
Somers et al., 2000;
Somers and Fujiwara, 2009). The light sensing function of the LOV domain was unveiled by the discovery that GI is essential for the stability of the ZTL protein (
Kim et al., 2007;
Somers and Fujiwara, 2009). GI, with no recognizable functional domain, physically interacts with ZTL in a blue light-stimulated manner (
Kim et al., 2007;
Somers and Fujiwara, 2009).
GI transcription is clock-controlled, and GI protein levels closely follow the
GI mRNA rhythm. Thus, it is the cycling of GI that confers a circadian rhythm to the abundance of ZTL rather than
ZTL transcription, which is constitutive. As a result, the GI-mediated stabilization of ZTL affects the abundance of the ZTL substrates, TOC1 and PRR5, in a time-dependent manner, which is necessary for proper clock function (
Kim et al., 2007;
Somers and Fujiwara, 2009). Recently, the reciprocal stabilization of cytosolic GI by ZTL was reported, which further implicates the importance of the ZTL-GI relationship by demonstrating a role for ZTL in sequestering a portion of GI to the cytosol (
Kim et al., 2013).
Further control of the TOC1 levels comes from PRR5, whose phosphorylation promotes interaction with TOC1 and enhances the TOC1 accumulation in the nucleus, preventing it from undergoing the ZTL-dependent protein turn-over in the cytoplasm (
Wang, 2010). In addition, a TOC1/PRR3 phosphorylation-dependent interaction may protect TOC1 from ZTL-mediated degradation (
Fujiwara et al., 2008).
Other evening phased genes, including LUX, ELF3 and ELF4, interact to form the ‘evening complex’, which represses the expression of the day-phased clock gene
PRR9. Around late night, the TOC1 protein in the nucleus starts acting as a DNA binding transcriptional repressor of
CCA1 and
LHY (
Gendron et al., 2012;
Huang et al., 2012;
Pokhilko et al., 2010;
Wang et al., 2013). However, the expression of
CCA1 and
LHY is resumed just before dawn, which might result from collaborative action with another CCA1 transcriptional repressor, CHE, in an undefined manner (
Pruneda-Paz, 2009).
Recently, the Myb transcription factor REVEILLE8 (RVE8, also known as
LHY and
CCA1-like 5, LCL5) was assigned as an important clock component (
Hsu, 2013;
Hsu and Harmer, 2014;
Rawat, 2011).
RVE8 shows dawn-phased gene expression, providing an important signal to the plant early in the morning. Like CCA1 and LHY, REV8 protein, with an afternoon-phased peak, binds to the EE (
Rawat, 2011). However, it induces hundreds of evening genes that contain EE in the promoter, including
PRR5, TOC1, GI, LUX and
ELF4, which is entirely opposite from the repressor role of CCA1 and LHY (
Hsu, 2013)
. PRR9, as well as
PRR5, is also activated by RVE8 (
Hsu, 2013;
Rawat, 2011). PRR9, 7, and 5 repress
RVE8, suggesting a model whereby the PRRs and RVEs regulate each other to form a negative feedback loop (
Hsu, 2013). Two close homologs of REV8, RVE4 and RVE6, were reported to have partially redundant roles with RVE8, interconnecting the morning and evening loops
Overall
, the dedicated empirical and computational studies carried out in this field have developed the current models of the plant clock. However, the roles of significant portions of the identified clock genes in the clock network have yet to be identified. Furthermore, diverse post-transcriptional mechanisms, post-translational regulation and chromatin remodeling are emerging as important mechanisms in the clock network, adding a new layer of clock regulation (
Cibois, 2010;
Nagel and Kay, 2012). Thus, much remains to be understood to accommodate such mechanisms in the context of the current clock network.
The relevance of the Arabidopsis clock to other crop species
Although most of the knowledge of plant clocks has come from studies on Arabidopsis, homologs of these genes in other plant species have recently begun to be characterized. In addition, the agronomic importance of the circadian system in plants is becoming more apparent (
Hsu and Harmer, 2014;
McClung, 2013). In the major crop species including rice (
Filichkin et al., 2011;
Mockler et al., 2007;
Nagano et al., 2012), maize (
Hayes et al., 2010;
Khan et al., 2010), papya (
Zdepski et al., 2008) and tomato (
Facella et al., 2008), clock control of the transcriptome levels has been characterized for many essential functions (e.g., photosynthesis, carbohydrate metabolism, light sensing and signaling).
In addition, key clock orthologues of CCA1, LHY and the PRR family have been found in many crop species, including bean (
Phaseolus vulgaris) (
Kaldis et al., 2003), chestnut (
Castanea sativa) (
Ramos et al., 2005), pea (
Pisum sativum) (
Hecht et al., 2005;
Liew et al., 2009;
Ramos et al., 2005), soybean (
Glycine max) (
Hudson, 2010), Brassica rapa (
Kim et al., 2012;
Lou et al., 2012), tomato (
Solanum lycopersicum) (
Facella et al., 2008) and rice (Oryza sativa)(
Murakami et al., 2007a;
Murakami et al., 2007b). More recently, other clock genes such as GI and ELF3 were functionally characterized in rice (
Izawa et al., 2011;
Yang et al., 2013). In many cases, these clock genes have been shown to cycle with similar phases as their Arabidopsis counterparts. Thus, it can be speculated that there has been significant evolutionary conservation of clock function and organization in many cases (
Song et al., 2010). However, much remains to be understood about the clocks of non-model plant systems.
Temperature compensation (TC)
Along with the signal of light, temperature is one of the most important environmental cues in many important physiological and developmental responses, such as flowering and hormonal regulation (
McClung and Davis, 2010). TC, as one of the defining features of circadian rhythms, refers to the relative insensitivity of the period length to ambient temperature over a range of physiologically relevant temperatures. The rate of a typical biochemical reaction doubles with an increase in temperature of 10°C (expressed as Q
10=2), but the rate of the circadian clock is relatively constant (Q
10 ~ 1) (
Jolma et al., 2010;
McClung and Davis, 2010). The reactions induced by change in the ambient temperature are presumably constrained by the TC mechanism. Otherwise, the period of the clock would fluctuate greatly, depending on the ambient temperature (
Jolma et al., 2010;
McClung and Davis, 2010).
According to the research on one of most extensively studied organisms for TC,
Drosophila melanogaster, the factors regulating the accumulation, dimerization, and nuclear transport of the core clock components PERIOD (PER) and TIMELESS (TIM) are important for the maintenance of TC (
Hamblen et al., 1998;
Price, 1997). This suggests that TC acts directly from the intrinsic properties, wherein the dynamic interactions of core clock proteins function across a wide temperature range (
Hamblen et al., 1998;
Price, 1997). In contrast, Neurospora TC studies have suggested other mechanisms, such as the importance of altered levels or alternative isoforms of core clock proteins at certain temperatures (
Baker et al., 2011;
Jolma et al., 2010;
Liu et al., 1997;
Price, 1997). At present, the mechanistic bases of TC have been incompletely defined, although it is likely that multiple pathways and mechanisms are involved (
McClung and Davis, 2010).
In Arabidopsis, several studies have reported that both of the morning and evening loops play a role in TC. Within the evening loop, Hall and colleagues reported that loss of GI function reduces the robustness and period length at high (27°C) and low (12°C) temperatures, demonstrating that
GI plays a critical role in TC (
Gould et al., 2006). They also found that
LHY and
CCA1 contribute differentially to TC at high and low temperatures, suggesting that a dynamic balance between
LHY and
GI is key for TC at high temperatures, whereas
LHY appears to be replaced by
CCA1 at low temperatures (
Gould et al., 2006).
In the morning loop, PRR7 and PRR9, negative regulators of
CCA1 and
LHY, were suggested to be important components of the TC mechanism by modulating the expression levels of those genes (
Salome et al., 2010;
Salome and McClung, 2005). While simultaneous inactivation of both PRR7 and PRR9 has no effect on the periods at low temperature, it results in extreme period lengthening at high temperatures (overcompensation) (
Salome et al., 2010). Inactivation of
CCA1 and
LHY, in turn, fully suppresses the overcompensation effects of
prr7prr9 double mutations, indicating that the temperature-dependent period lengthening of
prr7prr9 is due to the hyperactivation of
CCA1 and
LHY (
Salome et al., 2010). Portolés and Más elaborated on the emphasized role of CCA1 at low temperature by showing the antagonizing posttranslational modulation of CCA1 activity by CASEIN KINASE2 (CK2)-mediated phosphorylation of CCA1 (
Portoles and Mas, 2010). At low temperatures, CK2-mediated phosphorylation is reduced, but a consequent enhancement in the CCA1 binding activity occurs, indicating that these antagonizing actions may sustain a constant period at various ambient temperatures (
Portoles and Mas, 2010).
Therefore, the TC mechanism in Arabidopsis may not be an intrinsic property of the circadian oscillator, but rather may be derived from the recruitment of other genes which regulate the oscillator components (
Edwards et al., 2005;
Portoles and Mas, 2010;
Salome et al., 2010). As with other areas, much remains to be characterized for this aspect of clock regulation in plants.
The significance of TC in fitness under extreme thermal environments
Plants, as sessile species, successfully grow and develop across a wide range of ambient temperatures. The species have adapted to persevere over a range of ~50°C, including extremes of growth near 0°C at the coastlines of Antarctica and at near 50°C in the deserts of the hottest places on Earth (
McClung and Davis, 2010). To prevail in such diverse thermal environments, the plant populations in contrasting thermal environments may have accumulated relevant genetic modifications/natural variations as a result of differential selective pressures (
Leinonen et al., 2009).
Considering the dominant role of the clock in the development and physiology of plants, a TC mechanism that maintains the proper oscillator speed under physiologically relevant temperatures could be one of the most critical targets of the evolutionary forces/natural selection (
Edwards et al., 2005;
Johnson, 2005). In an attempt to find genetic variations in TC, Millar and colleagues performed a QTL study on two natural accessions adapted in different thermal environments (
Edwards et al., 2005): one adapted to a hot climate, Cape Verde Island (Cvi), and the other originating from Northern Europe (Landsberg, L
er). In spite of the similar period phenotype between the parental lines, several temperature-specific QTLs for the clock period were identified, suggesting that natural genetic variations in TC mechanisms occur (
Edwards et al., 2005). Among the QTLs, the core clock components, GI and ZTL, were proposed as strong candidates, accounting for much of the variation (
Edwards et al., 2005). By sequence comparison, they postulated that the amino acid substitutions of GI
L718F and ZTL
P35T (L
er to Cvi) are the underlying causal factors for the period variation at the tested temperatures (22°C and 27°C respectively) (
Edwards et al., 2005). Since the effects of specific mutations in these genes on the period are in opposite directions at different temperatures, the Cvi’s clock system may have accumulated alleles with opposing effects epistatically acting at the different temperature regimes. This action would theoretically neutralize the effect on the period by balancing of the two across a range of relevant temperatures to attain TC. In reality, it was found that Cvi’s period and the physiological response were much more stable at the high temperature (30°C) compared to L
er (
Edwards et al., 2005;
Kusakina et al., 2014). Thus, it is likely that an antagonistic balance among the alleles observed in Cvi’s clock, including GI
L718F and ZTL
P35T, may be the result of an adaptation to different local thermal environments, presumably to the hot climate of Cape Verde Island (CVI) (
Edwards et al., 2005).
In the plant clock, as in other circadian systems, an oscillator whose period is consonant with the external environment significantly enhances the survival rates, which is the result of better physiological performances (
Dodd et al., 2005). Therefore, the recalibrated TC-related clock property through GI/ZTL natural variations that constrain the period to remain at a similar value even under the increasing thermal conditions, as in the CVI, may have significant fitness value.
Concluding remarks: the significance of TC study for future crop improvements
The process of crop domestication has led to the production of plants with many innovative features suitable for agriculture, but human selection has been trading off the natural diversity of gene/allelic pools, which may be critical for successful adaptation to various local environments (
Vigouroux et al., 2011). Thus, it is not just a coincidence that agriculture is affected by recent climate change, especially the global increase in temperature (
Lobell et al., 2011). For example, it is estimated that the mean temperature rise between 1981 and 2002 reduced the yields of major cereals by $5 billion per year (
Lobell et al., 2011). Therefore, developing crops that are better adapted to the current temperature situation will be important for food production around the world (
Varshney et al., 2011).
Plant thermo tolerance is a multigenic character; numerous biochemical and metabolic traits are orchestrated in the development and maintenance of heat tolerance, including the maintenance of membrane stability, scavenging of reactive oxygen species (ROS), production of antioxidants, accumulation and adjustment of compatible solute, and induction of chaperone signaling and transcriptional activation (
Bita and Gerats, 2013). Thus, breeding to provide greater stress tolerance is a great challenge due to the complexity. In this regard, implementing naturally modified diverse TC functions which exist in nature can be a reasonable approach in order to establish fundamental solutions for sustainable crop production under these changing thermal environments. Thus, it is important to understand and characterize the TC mechanisms as a parameter for temperature-related adaptive processes more precisely, so that such information can be applied more efficiently in related fields for various purposes.
ACKNOWLEDMENTS
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education, Science and Technology (2014-0188).
Fig. 1
Multiple-interlocked transcription-translation feedback loop model of Arabidopsis circadian clock.
In the “morning”, expressed CCA1 and LHY positively regulate PRR7 and PRR9; however, PRR7 and PRR9 proteins, along with the afternoon-expressed PRR5, bind to the CCA1 and LHY promoters to sequentially repress the CCA1/LHY transcriptional expression. In the evening, ZTL, an F-box-containing protein and part of a Skp/Cullin/F-box (SCF) E3 ubiquitin ligase complex, directly targets both TOC1 and PRR5 protein in the cytosol, leading to their degradation via the proteosome pathway. The evening-expressed gene GI, with no recognizable functional domain, physically interacts with ZTL in a blue light-stimulated manner, conferring ZTL with the post-transcriptional oscillation. This supports the amplitude of TOC1 protein to be time dependent, which is necessary for proper clock function. Phosphorylation of PRR5 promotes interaction with TOC1, enhancing its accumulation in the nucleus and preventing it from undergoing ZTL-dependent protein turn-over in the cytoplasm. In addition, a TOC1/PRR3 phosphorylation-dependent interaction may protect TOC1 from ZTL-mediated degradation. Meanwhile, evening phased genes, LUX, ELF3 and ELF4, interact to form the ‘evening complex’, which represses expression of the day-phased clock gene PRR9. TOC1 protein in the nucleus starts acting as a DNA binding transcriptional repressor of CCA1 and LHY; however, CCA1 and LHY expression is resumed just before dawn, which might be the result of a collaborative action with another CCA1 transcriptional repressor, CHE, in an undefined manner. The Myb transcription factor RVE8, a dawn-phased gene expression, provides an important signal to the plant early in the morning. REV8 protein, with the afternoon-phased peak, binds to the EE, which induces the expression of hundreds of evening genes, including PRR5, TOC1, GI, LUX and ELF4. PRR9, as well as PRR5, was also found to be activated by RVE8. Conversely, PRR9, 7 and 5 repress RVE8, suggesting a model in which PRRs and RVEs regulate each other to form a negative feedback loop. The two close homologs of REV8, RVE4 and RVE6, were reported to have partially redundant roles with RVE8, interconnecting the morning and evening loops. Red solid arrow: activation; red dashed arrow: indirect activation; black perpendicular bars: repression.
CCA1, CIRCADIAN CLOCK ASSOCIATED 1; LHY, LATE ELONGATED HYPOCOTYL; TOC1, TIMING OF CAB2 EXPRESSION 1; GI, GIGANTEA, ZTL, ZEITLUPE, PRR, PSEUDO-RESPONSE REGULATOR; LUX, LUX ARRHYTHMO; ELF, EARLY FLOWERING, CHE, CCA1 HIKING EXPEDITION REV, REVEILLE.
References
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA. 2001. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science. 293: 880-883.
- Alabadi D, Yanovsky MJ, Mas P, Harmer SL, Kay SA. 2002. Critical Role for CCA1 and LHY in Maintaining Circadian Rhythmicity in Arabidopsis. Curr Biol. 12: 757-761.
- Baker CL, Loros JJ, Dunlap JC. 2011. The circadian clock of Neurospora crassa. FEMS Microbiol Rev. 36: 95-110.
- Bita CE, Gerats T. 2013. Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front Plant Sci. 4: 273
- Cibois M, Gautier-Courteille C, Legagneux V, Paillard L. 2010. Post-transcriptional controls - adding a new layer of regulation to clock gene expression. Trends Cell Biol. 20: 533-41.
- Toth R, Coupland G. 2010. Plant development goes like clockwork. Trends Genet. 26: 296-306.
- Dodd AN, Salathia N, Hall A, Kévei E, Tóth R, Nagy F, Hibberd JM, Millar AJ, Webb AA. 2005. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science. 309: 630-633.
- Edwards KD, Lynn JR, Gyula P, Nagy F, Millard AJ. 2005. Natural allelic variation in the temperature-compensation mechanisms of the Arabidopsis thaliana circadian clock. Genetics. 170: 387-400.
- Facella P, Lopez L, Carbone F, Galbraith DW, Giuliano G, Perrotta G. 2008. Diurnal and circadian rhythms in the tomato transcriptome and their modulation by cryptochrome photoreceptors. PLoS One. 3: e2798
- Filichkin SA, Breton G, Priest HD, Dharmawardhana P, Jaiswal P, Fox SE, Michael TP, Chory J, Kay SA, Mockler TC. 2011. Global profiling of rice and poplar transcriptomes highlights key conserved circadian-controlled pathways and cis-regulatory modules. PLoS One. 6: e16907
- Fujiwara S, Wang L, Han L, Suh SS, Salomé PA, McClung CR, Somers DE. 2008. Post-translational regulation of the Arabidopsis circadian clock through selective proteolysis and phosphorylation of pseudo-response regulator proteins. J Biol Chem. 283: 23073-23083.
- Gendron JM, Pruneda-Paz JL, Doherty CJ, Gross AM, Kang SE, Kay SA. 2012. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc Natl Acad Sci U S A. 109: 3167-3172.
- Gould PD, Locke JC, Larue C, Southern MM, Davis SJ, Hanano S, Moyle R, Milich R, Putterill J, Millar AJ, Hall A. 2006. The molecular basis of temperature compensation in the Arabidopsis circadian clock. Plant Cell. 18: 1177-1187.
- Hanblen MJ, White NE, Emery PT, Kaiser K, Hall JC. 1998. Molecular and behavioral analysis of four period mutants in Drosophila melanogaster encompassing extreme short, novel long, and unorthodox arrhythmic types. Genetics. 149: 165-178.
- Harmer SL. 2009. The circadian system in higher plants. Annu Rev Plant Biol. 60: 357-77.
- Harmer SL, Kay SA. 2005. Positive and negative factors confer phase-specific circadian regulation of transcription in Arabidopsis. Plant Cell. 17: 1926-1940.
- Hayes KR, Beatty M, Meng X, Simmons CR, Habben JE, Danilevskaya ON. 2010. Maize global transcriptomics reveals pervasive leaf diurnal rhythms but rhythms in developing ears are largely limited to the core oscillator. PLoS One. 5: e12887
- Hecht V, Foucher F, Ferrándiz C, Macknight R, Navarro C, Morin J, Vardy ME, Ellis N, Beltrán JP, Rameau C, Weller JL. 2005. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 137: 1420-1434.
- Hsu PY, Devisetty UK, Harmer SL. 2013. Accurate timekeeping is controlled by a cycling activator in Arabidopsis. Elife. 2: e00473
- Hsu PY, Harmer SL. 2014. Wheels within wheels: the plant circadian system. Trends Plant Sci. 19: 240-249.
- Huang W, Perez-Garcia P, Pokhilko A, Millar AJ, Antoshechkin I, Riechmann JL, Mas P. 2012. Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science. 336: 75-79.
- Hudson KA. 2010. The Circadian Clock-controlled Transcriptome of Developing Soybean Seeds. Plant Genome. 3: 3-13.
- Izawa T, Mihara M, Suzuki Y, Gupta M, Itoh H, Nagano AJ, Motoyama R, Sawada Y, Yano M, Hirai MY, Makino A, Nagamura Y. 2011. Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell. 23: 1741-1755.
- Johnson CH. 2005. Testing the adaptive value of circadian systems. Methods Enzymol. 393: 818-837.
- Jolma IW, Laerum OD, Lillo C, Ruoff P. 2010. Circadian oscillators in eukaryotes. Wiley Interdiscip Rev Syst Biol Med. 2: 533-549.
- Kaldis AD, Kousidis P, Kesanopoulos K, Prombona A. 2003. Light and circadian regulation in the expression of LHY and Lhcb genes in Phaseolus vulgaris. Plant Mol Biol. 52: 981-997.
- Khan S, Rowe SC, Harmon FG. 2010. Coordination of the maize transcriptome by a conserved circadian clock. BMC Plant Biol. 10: 126
- Kiba T, Henriques R, Sakakibara H, Chua NH. 2007. Targeted degradation of PSEUDO-RESPONSE REGULATOR5 by an SCFZTL complex regulates clock function and photomorphogenesis in Arabidopsis thaliana. Plant Cell. 19: 2516-2530.
- Kim J, Geng R, Gallenstein RA, Somers DE. 2013. The F-box protein ZEITLUPE controls stability and nucleocytoplasmic partitioning of GIGANTEA. Development. 140: 4060-4069.
- Kim JA, Kim JS, Hong JK, Lee YH, Choi BS, Seol YJ, Jeon CH. 2012. Comparative mapping, genomic structure, and expression analysis of eight pseudo-response regulator genes in Brassica rapa. Mol Genet Genomics. 287: 373-388.
- Kim WY, Fujiwara S, Suh SS, Kim J, Kim Y, Han L, David K, Putterill J, Nam HG, Somers DE. 2007. ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature. 449: 356-360.
- Kusakina J, Gould PD, Hall A. 2014. A fast circadian clock at high temperatures is a conserved feature across Arabidopsis accessions and likely to be important for vegetative yield. Plant Cell Environ. 37: 327-340.
- Leinonen PH, Sandring S, Quilot B, Clauss MJ, Mitchell-Olds T, Agren J, Savolainen O. 2009. Local adaptation in European populations of Arabidopsis lyrata (Brassicaceae). Am J Bot. 96: 1129-1137.
- Liew LC, Hecht V, Laurie RE, Knowles CL, Vander Schoor JK, Macknight RC, Weller JL. 2009. DIE NEUTRALIS and LATE BLOOMER 1 contribute to regulation of the pea circadian clock. Plant Cell. 21: 3198-3211.
- Liu Y, Garceau NY, Loros JJ, Dunlap JC. 1997. Thermally regulated translational control of FRQ mediates aspects of temperature responses in the neurospora circadian clock. Cell. 89: 477-486.
- Lobell DB, Schlenker W, Costa-Roberts J. 2011. Climate trends and global crop production since 1980. Science. 333: 616-620.
- Locke JC, Kozma-Bognár L, Gould PD, Fehér B, Kevei E, Nagy F, Turner MS, Hall A, Millar AJ. 2006. Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol Syst Biol. 2: 59
- Lou P, Wu J, Cheng F, Cressman LG, Wang X, McClung CR. 2012. Preferential retention of circadian clock genes during diploidization following whole genome triplication in Brassica rapa. Plant Cell. 24: 2415-2426.
- Más P, Kim WY, Somers DE, Kay SA. 2003. Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana. Nature. 426: 567-570.
- Mcclung CR. 2011. The genetics of plant clocks. Adv Genet. 74: 105-139.
- Mcclung CR. 2013. Beyond Arabidopsis: the circadian clock in non-model plant species. Semin Cell Dev Biol. 24: 430-436.
- Mcclung CR, Davis SJ. 2010. Ambient thermometers in plants: from physiological outputs towards mechanisms of thermal sensing. Curr Biol. 20: R1086-R1092.
- McWatters HG, Devlin PF. 2011. Timing in plants-a rhythmic arrangement. FEBS Lett. 585: 1474-1484.
- Mizoguchi T, Wheatley K, Hanzawa Y, Wright L, Mizoguchi M, Song HR, Carré IA, Coupland G. 2002. LHY and CCA1 are partially redundant genes required to maintain circadian rhythms in Arabidopsis. Dev Cell. 2: 629-641.
- Mizuno T Nakamichi N . 2005. Pseudo-Response Regulators (PRRs) or True Oscillator Components (TOCs). Plant Cell Physiol. 46: 677-685.
- Mockler TC, Michael TP, Priest HD, Shen R, Sullivan CM, Givan SA, McEntee C, Kay SA, Chory J. 2007. The DIURNAL project: DIURNAL and circadian expression profiling, model-based pattern matching, and promoter analysis. Cold Spring Harb Symp Quant Biol. 72: 353-363.
- Murakami M, Tago Y, Yamashino T, Mizuno T. 2007a. Characterization of the rice circadian clock-associated pseudo-response regulators in Arabidopsis thaliana. Biosci Biotechnol Biochem. 71: 1107-1110.
- Murakami M, Tago Y, Yamashino T, Mizuno T. 2007b. Comparative overviews of clock-associated genes of Arabidopsis thaliana and Oryza sativa. Plant Cell Physiol. 48: 110-121.
- Nagano AJ, Sato Y, Mihara M, Antonio BA, Motoyama R, Itoh H, Nagamura Y, Izawa T. 2012. Deciphering and prediction of transcriptome dynamics under fluctuating field conditions. Cell. 151: 1358-1369.
- Nagel DH, Kay SA. 2012. Complexity in the wiring and regulation of plant circadian networks. Curr Biol. 22: R648-R657.
- Nakamichi N, Kiba T, Henriques R, Mizuno T, Chua NH, Sakakibara H. 2010. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell. 22: 594-605.
- Pokhilko A, Hodge SK, Stratford K, Knox K, Edwards KD, Thomson AW, Mizuno T, Millar AJ. 2010. Data assimilation constrains new connections and components in a complex, eukaryotic circadian clock model. Mol Syst Biol. 6: 416
- Portolés S, Más P. 2010. The functional interplay between protein kinase CK2 and CCA1 transcriptional activity is essential for clock temperature compensation in Arabidopsis. PLoS Genet. 6: e1001201
- Price JL. 1997. Insights into the molecular mechanisms of temperature compensation from the Drosophila period and timeless mutants. Chronobiol Int. 14: 455-468.
- Pruneda-Paz JL, Breton G, Para A, Kay SA. 2009. A functional genomics approach reveals CHE as a component of the Arabidopsis circadian clock. Science. 323: 1481-1485.
- Ramos A, Pérez-Solís E, Ibáñez C, Casado R, Collada C, Gómez L, Aragoncillo C, Allona I. 2005. Winter disruption of the circadian clock in chestnut. Proc Natl Acad Sci U S A. 102: 7037-7042.
- Rawat R, Takahashi N, Hsu PY, Jones MA, Schwartz J, Salemi MR, Phinney BS, Harmer SL. 2011. REVEILLE8 and PSEUDO-REPONSE REGULATOR5 Form a Negative Feedback Loop within the Arabidopsis Circadian Clock. PLoS Genet. 7: e1001350
- Salomé PA, McClung CR. 2005. PSEUDO-RESPONSE REGULATOR 7 and 9 Are Partially Redundant Genes Essential for the Temperature Responsiveness of the Arabidopsis Circadian Clock. Plant Cell. 17: 791-803.
- Salomé PA, Weigel D, McClung CR. 2010. The role of the Arabidopsis morning loop components CCA1, LHY, PRR7, and PRR9 in temperature compensation. Plant Cell. 22: 3650-3661.
- Somers DE, Fujiwara S. 2009. Thinking outside the F-box: novel ligands for novel receptors. Trends Plant Sci. 14: 206-213.
- Somers DE 1, Schultz TF, Milnamow M, Kay SA. 2000. ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis. Cell. 101: 319-329.
- Song YH, Ito S, Imaizumi T. 2010. Similarities in the circadian clock and photoperiodism in plants. Curr Opin Plant Biol. 13: 594-603.
- Varshney RK, Bansal KC, Aggarwal PK, Datta SK, Craufurd PQ. 2011. Agricultural biotechnology for crop improvement in a variable climate: hope or hype? Trends Plant Sci. 16: 363-371.
- Vigouroux Y, Barnaud A, Scarcelli N, Thuillet AC. 2011. Biodiversity, evolution and adaptation of cultivated crops. C R Biol. 334: 450-457.
- Wang L, Fujiwara S, Somers DE. 2010. PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock. EMBO J. 29: 1903-1915.
- Wang L, Kim J, Somers DE. 2013. Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription. Proc Natl Acad Sci U S A. 110: 761-766.
- Yang Y, Peng Q, Chen GX, Li XH, Wu CY. 2013. OsELF3 Is Involved in Circadian Clock Regulation for Promoting Flowering under Long-Day Conditions in Rice. Molecular Plant. 6: 202-215.
- Zdepski A, Wang W, Priest HD, Ali F, Alam M, Mockler TC, Michael TP. 2008. Conserved Daily Transcriptional Programs in Carica papaya. Trop Plant Biol. 1: 236-245.