Abstract
Common wheat (Triticum aestivum L.) and durum wheat (T. turgidum L. subsp. Durum) are major staple food crops in the world. However, their production are limited by environmental stress such as drought. In order to evaluate wheat’s response to drought, a total of 77 common wheat and durum wheat consisted of 19 Korean common wheat, 30 Tunisian common wheat and 28 Tunisian durum wheat were used in this study. Drought stress was applied for 21 days by suspending water application starting at the third leaf-expansion stage, followed by watering for the recovery of wheat until harvesting. Phenotypic parameters such as plant height, leaf length, tiller number, chlorophyll content, days to flowering and dry weight were scored during and after the treatment. Drought tolerance trait index (DTTI) and drought tolerance index (DTI) were calculated and used as criteria for selection of drought tolerance. At the end of treatment, most of the parameters except tiller numbers significantly decreased. Even after re-watering, plant height, leaf length, and dry weight continuously decreased. However, leaf chlorophyll content, and days to flowering of both stressed and non-stressed plants showed no significant differences. A total of 17 drought tolerance related simple sequence repeats (SSR) markers were used to identify genetic distance between Korean and Tunisian cultivars and elucidate possible use of marker systems for drought resistance. The common wheat and durum wheat cultivars formed different clusters for drought tolerance (resistance, moderate resistance, susceptible) using the SSR data. The results obtained in this study could help to increase genetic resources and breeding program for drought tolerance.
-
Key words: Common wheat, Drought tolerance, Durum wheat, SSR marker
INTRODUCTION
There is a current concern for future global food production using crops amidst an increasing world population (
Brown & Funk 2008). Environmental problems such as drought, floods, and salinity have been reported to be more severe in some areas. In semi-arid areas, it is predicted that the grain yield of main cereal crops such as wheat and rice would decrease in the following 20 years (
Lobell et al. 2008). To overcome these problems, crop improvements not only to increase yield but also to enhance tolerance to environmental stress are needed to improve global food security (
Takeda & Matsuoka 2008).
Wheat (
Triticum spp.) is a major staple food crop around the world (
Munns et al. 2006). Common wheat (
T. aestivum L.) and durum wheat (
T. turgidum L. subsp.
Durum) are major species of cultivated wheat. Drought stress is one of the global problems that limit crop production (
Pan et al. 2002). About 32% of the wheat cultivating regions in developing countries experience drought stress during the growing season (
Rajaram 2001).
Microsatellites or simple sequence repeats (SSR) have been widely utilized in plants as molecular markers for specific phenotypic traits (
Gupta & Varshney 2000). Marker-assisted selection is useful for improving drought tolerance in wheat (
Quarrie et al. 2003). SSR markers can show the genetic diversity for drought stress in wheat (
El Siddig et al. 2013).
Recent studies involving
in vitro selection using polyethylene glycol (PEG) or mannitol applied to generate water stress in plants were performed to develop drought- tolerant plants (
Rai et al. 2011). However, some results of
in vivo and
in vitro studies are not necessarily associated and had to be considered individually (
Farshadfar et al. 2012). Further, plant responses to PEG treatment could not indicate the drought tolerance for the entire plant growth period. Drought stress is a limiting factor to plant growth and dry matter (
Jaleel et al. 2009); therefore, morphological traits can be used as a criteria for drought tolerance.
Korea is located in East Asia. The winter climate is cold and dry, whereas it is hot and humid during summer. In Korea, wheat is cultivated as a winter crop. Accordingly, wheat is subjected to drought stress during its early stages of growth in Korea.
Tunisia is located in North Africa; it is bordered by the Mediterranean Sea on the north and east, whereas the Sahara desert is positioned to its south. The climate in the north is warm with dry summer climate (Mediterranean climate), whereas the climate in the mid-south is semi-arid, where soil moisture barely meets wheat seedling growth and rainfall is low. Thus, in Tunisia, wheat experiences drought stress during its entire growing period.
The main objective of this study was to screen Korean common wheat, Tunisian common wheat, and Tunisian durum wheat cultivars for drought tolerance by using morphological traits and SSR markers to utilize them for future breeding programs.
MATERIALS AND METHODS
Plant materials and experimental conditions
A total of 77 cultivars (49 common wheat lines and 28 durum wheat lines) were used in this study. Of these, 19 lines were Korean common wheat, and 58 lines were Tunisian wheat. Seeds of Korean wheat cultivars were provided by the National Institute of Crop Science (NICS), Rural Development Administration (RDA) of Korea. The Tunisian seeds were obtained from the National Agrobiodiversity Center (NAC), RDA and the National Plant Germplasm System (NPGS), United States Department of Agriculture (USDA). Information about the Tunisian common wheat and durum wheat accessions was obtained from the RDA-Genebank Information Center (
http://www.genebank.go.kr/) and the Agricultural Research Service-Germplasm Resources Information Network (ARS-GRIN) (
http://www.ars-grin.gov/).
The experiment was conducted during the 2013 growing season at the greenhouse of the Research Farm of Korea University (Namyangju-si, Gyeonggi-do, Korea). Two seeds were planted per pot (5′ 5′ 16 cm) filled with soil (Sunshine mix #1). Immediately after emergence, seedlings were thinned out to let each plant grow in each pot. Five plants per line from each control and treatment groups were subjected to the test.
All plants were grown with sufficient watering until the drought treatment was initiated. Drought stress was imposed at the fully expanded third leaf stage by suspending water application, whereas plants in the non-stressed condition received sufficient water every two days. The treatment was applied for three weeks. After the treatment, all plants were provided with sufficient water and grown until spike harvest.
Morphological traits measurement
Phenotypic parameters such as plant height, average leaf length, number of tillers per plant, and leaf chlorophyll content were measured at three time points (the initiation of treatment, the end of treatment, and 28 days after the end of treatment). The first measurement was used for normalization, and the others were used in calculating the degree of recovery after drought stress. Days to flowering and shoot dry weight were also scored.
Leaf chlorophyll content was recorded by using a portable chlorophyll meter (SPAD-502, Minolta, Japan). SPAD-502 can measure leaf chlorophyll content in a fast and nondestructive way (Yıldırım
et al. 2001). Drought tolerance trait index (DTTI) and drought tolerance index (DTI) were calculated using the formula of
Shahzad et al. (2012) and
Ali et al. (2007). If the mean DTTI for a trait was >100, it indicates that the trait would increase during drought stress. Thus, the reciprocal of DTTI was used to calculate DTI.
DTI was calculated as the mean of DTTIs for morphological traits showing the tendency to drought stress and recovery. Based on DTI, drought tolerance of 77 common wheat and durum wheat cultivars was determined as resistant lines, moderate lines, and susceptible lines.
SSR marker analysis
A total of 17 SSR primers were used in the analysis (
Table 2). Information on the markers was obtained from the Agricultural Research Service-GrainGenes 2.0 (
http://wheat.pw.usda.gov). Polymerase chain reactions (PCRs) were performed as described by
Ciucă & Petcu (2009), with minor modification. PCR products were separated on agarose gel, and visualized and recorded using a gel documentation system.
The analysis was performed using the NTSYS-pc version 2.1 software using qualitative data for the similarity analysis, Sequential Agglomerative Hierarchical Nesting (SAHN) based on Unweighted Pair-Group Method with Arithmetic average (UPGMA) for the cluster analysis, and tree plot for the graphic visualization of dendrograms.
RESULTS
Morphological traits measurement
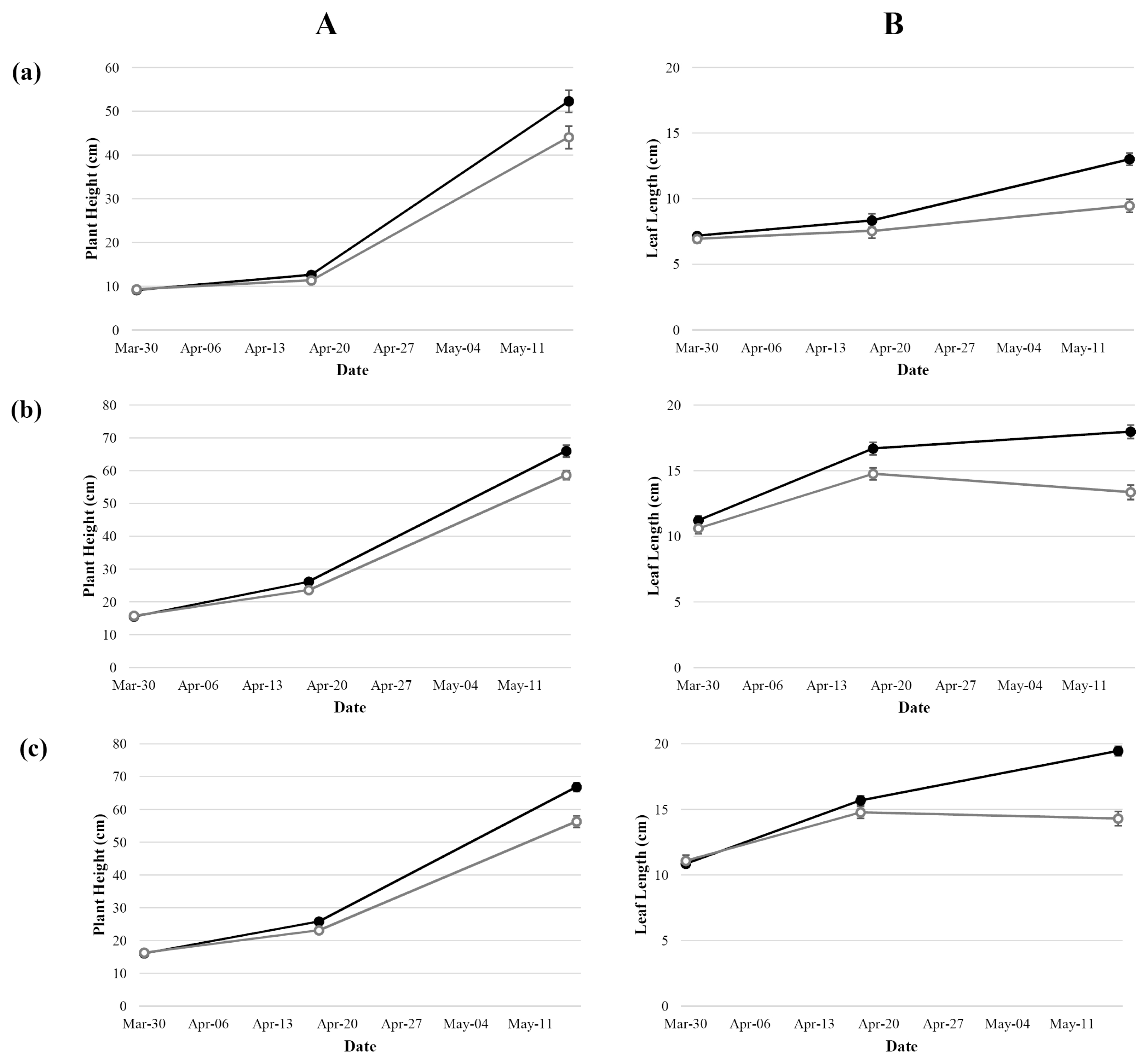
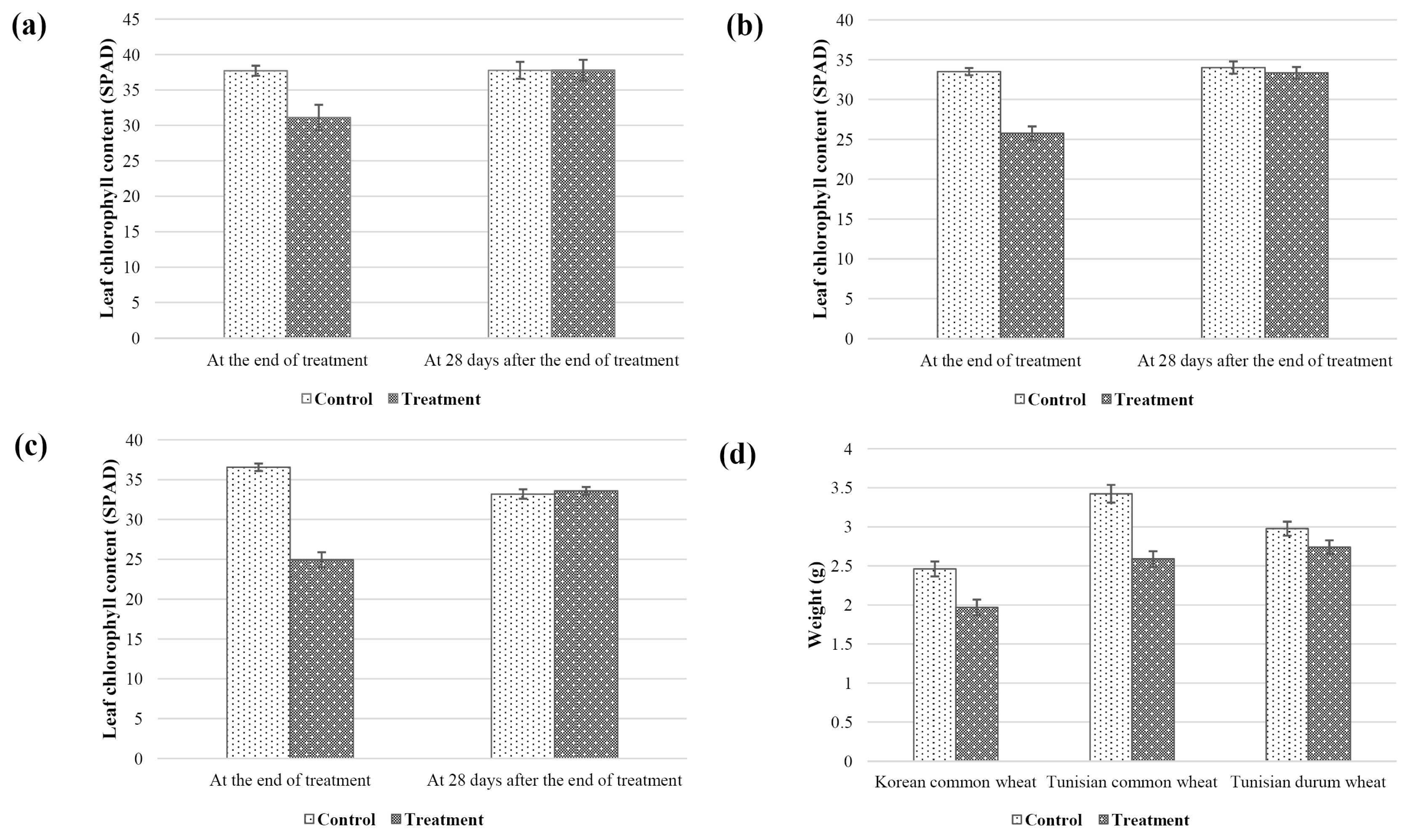
Agronomic traits of both control (non-stressed) and stressed plants were measured at three time points: at the initiation of the treatment, end of treatment (21 days without watering), and 28 days after the end of treatment. Compared to the non-stressed plant, plant height, leaf length, leaf chlorophyll content, and shoot dry weight of the treated plants decreased (
Fig. 1A, B, and
2). Despite re-watering, the gap of those parameters between control and treatment increased.
The magnitude of DTTI at the end of the treatment indicates the response of the plants to drought stress. At the end of the treatment, the mean DTTI for plant height was 88.3 ± 1.57. Tunisian durum wheat cultivars showed the highest plant height (88.7 ± 2.18%), followed by the Korean common wheat cultivars and Tunisian common wheat cultivars (
Table 3). The mean DTTI for average leaf length was 93.2 ± 1.88%. Korean common wheat cultivars showed the highest leaf length (94.7 ± 5.49%) followed by the Tunisian common wheat cultivars and Tunisian durum wheat cultivars (
Table 3). The mean DTTI for the number of tillers was 98.5 ± 3.27%. Korean common wheat cultivars showed the highest number of tillers (109.1 ± 4.33%) followed by the Tunisian common wheat cultivars and Tunisian durum wheat cultivars (
Table 3). The mean DTTI for leaf chlorophyll content was 75.3 ± 1.82%. Korean common wheat cultivars showed the highest leaf chlorophyll content (82.7 ± 4.57%), followed by the Tunisian common wheat cultivars and the Tunisian durum wheat cultivars (
Table 3). Overall, DTTIs in most genotypes were <100%, except for the number of tillers in Korean common wheat cultivars.
The magnitude of DTTI at 30 days after the end of the treatment indicates the ability of plants to recover from the drought stress. At 28 days after the end of the treatment, the mean DTTI for plant height was 84.3 ± 2.04%. Tunisian common wheat cultivars showed the greatest plant height (86.9 ± 3.59%), followed by the Tunisian durum wheat cultivars and the Korean common wheat cultivars (
Table 4). The mean DTTI for average leaf length was 75.5.2 ± 2.60%. Tunisian common wheat cultivars showed the highest leaf length (79.3 ± 4.03%), followed by the Korean common wheat and the Tunisian durum wheat cultivars (
Table 4). The mean DTTI for the number of tillers was 132.5 ± 6.72%. Korean common wheat cultivars showed the highest number of tillers (154 ± 15.02%), followed by the Tunisian common wheat and the Tunisian durum wheat cultivars (
Table 4). The mean DTTI for leaf chlorophyll content was 100.2 ± 1.24%. Tunisian durum wheat cultivars showed the highest leaf chlorophyll content (101.9 ± 2.14), followed by the Korean common wheat and Tunisian common wheat cultivars (
Table 4). Compared to measurement at the end of the treatment, the mean DTTIs for plant height and average leaf length decreased, whereas those of the number of tillers per plant and leaf chlorophyll content increased.
For shoot dry weight, the mean DTTI was 84.2 ± 2.22%; the Tunisian durum wheat cultivars showed the highest shoot dry weights, followed by the Korean common wheat cultivars and Tunisian common wheat cultivars (
Table 5). For days to flowering, the mean DTTI was 101.0 ± 0.35% and the Tunisian durum wheat cultivars showed the highest values, followed by the Tunisian common wheat cultivars and Korean common wheat cultivars (
Table 5). Most of the shoot dry weight in the treatment was lower than in the control, whereas the days to flowering in both the treatment and the control were almost same.
The magnitude of DTI indicates a combined plant response using four parameters (plant height, average leaf length, leaf chlorophyll content, and dry shoot weight) and two stages (21 days after treatment and 28 days after the end of the treatment). The average DTI of 77 wheat cultivars was 85.5%. Based on the magnitude of DTI, drought tolerance of the 77 wheat cultivars was determined as resistant lines (above 90%), moderate lines (81–90%), and susceptible lines (below 80%). From the 77 cultivars, 23 lines were resistant, 33 lines were moderate, and 21 lines were susceptible (
Table 6). Four cultivars that showed >100% DTI were selected as the most resistant lines (
Table 7).
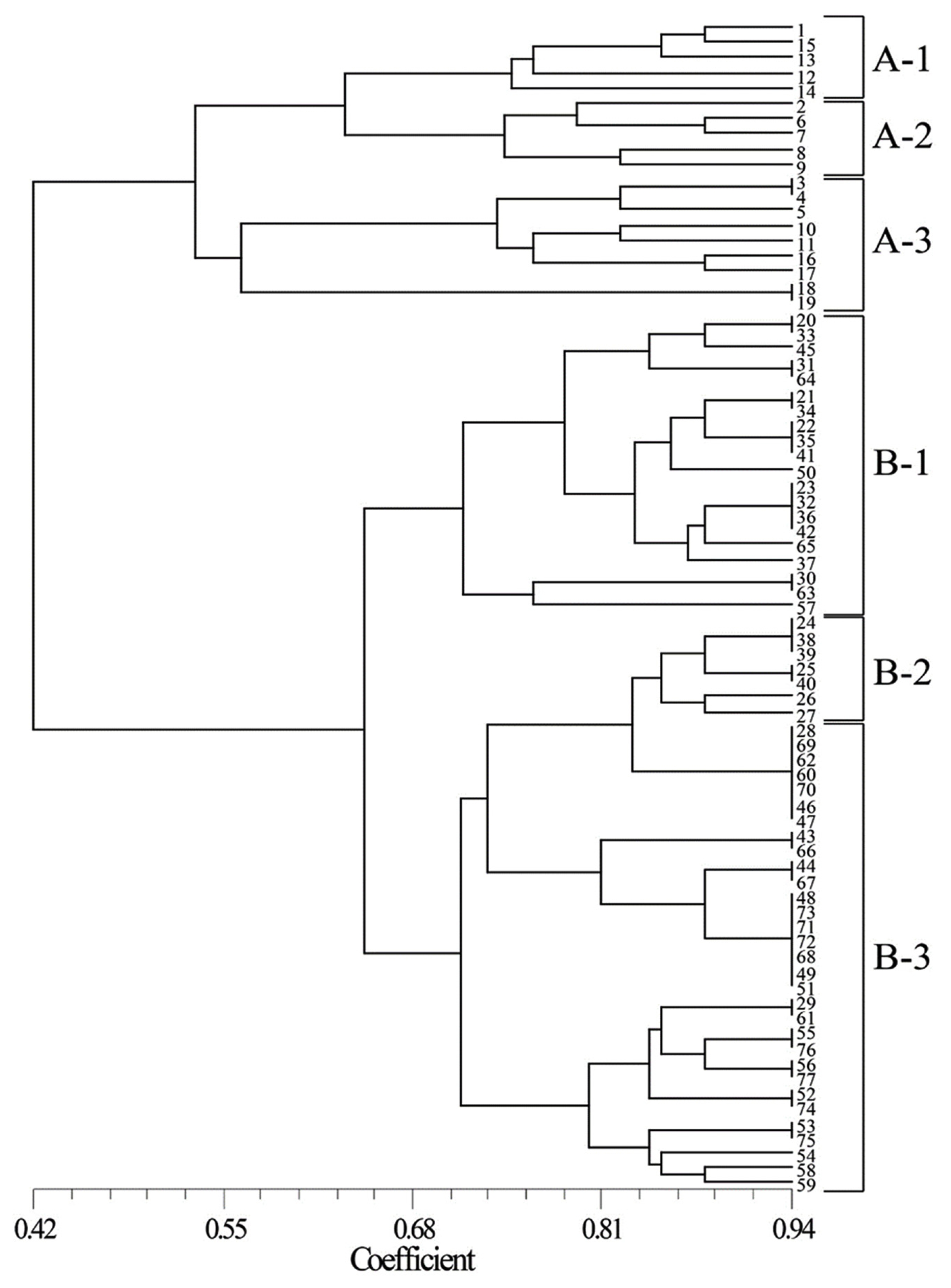
SSR analysis was performed to identify markers for drought tolerance. A dendrogram was constructed from the cluster analysis of similarity coefficients of SSR markers (
Fig. 3). A total of 77 cultivars were classified into two groups: group A (comprised of Korean cultivars) and group B (composed of Tunisian cultivars). Groups A and B were further categorized into three clusters each. Cluster A-1 is comprised of moderate resistant and resistant lines and had average DTI of 90.9 ± 4.45%. The average DTI of cluster A-2 was 72.4 ± 5.52%, which is comprised of mostly susceptible lines and a few moderate lines. The average DTI of cluster A-3 was 92.5 ± 2.88%, which is comprised of mostly resistant lines and a few moderate lines. The average DTI of cluster B-1 was 92.5 ± 1.96%, composed mostly of resistant lines and a few of moderate lines. The average DTI of cluster B-2 was 89.7 ± 1.38%, consisted of resistant lines and moderate lines. The average DTI of cluster B-3 was 80.1 ± 1.15%, consisted of of susceptible lines and moderate lines.
DISCUSSION
This study demonstrates that wheat cultivars are sensitive to drought treatment. Plant height, average leaf length, and leaf chlorophyll content decreased, whereas the number of tillers did not change. After re-watering, plant height and average leaf length continued to decrease, whereas leaf chlorophyll content showed similar values to that of the control and the number of tillers increased. Shoot dry weight also decreased, whereas days to flowering did not change. SSR analysis showed the genetic differences between the Korean and Tunisian cultivars based on drought tolerance.
Based on morphological trait measurement, the Korean common wheat and Tunisian common and durum wheat cultivars were affected by drought stress. Plant height and average leaf length did not recover fully, decreasing at the end of treatment and at 28 days after the end of treatment. Further, drought stress resulted in a reduced shoot dry weight. Thus, drought stress may result in the inhibition of growth. According to
Zhu (2001), water stress inhibits plant growth by inhibiting cell division. However, the leaf chlorophyll content of the drought-stressed plants decreased at the end of treatment, whereas it increased to levels similar to that of the control at 28 days after the end of the treatment. This indicates that the chlorophyll content of the newly produced leaves after re-watering in the treatment condition was similar to that in the control condition.
In terms of the number of tillers, Korean common wheat cultivars increased at both the end of treatment and 28 days after the end of treatment, whereas that of the Tunisian common wheat and durum wheat cultivars decreased at the end of treatment. According to
Veesar et al. (2007), Indus-66, which is a more tolerant wheat genotype to water stress than other genotypes, formed a lower number of tillers. In addition,
Castillo et al. (2007) reported that the number of non-productive tillers, which developed after restoration, increased with osmotic stress in rice. There might be a negative relationship between the number of non-productive tillers and drought tolerance.
Further, there was no clear difference between the control and treatment in terms of days to flowering, indicating that the number of tillers and days to flowering were not affected by drought stress. According to
Foulkes et al. (2002), flowering date was typically neutral to drought stress.
Anjum et al. (2011) reported that plant height, stem diameter, and leaf area decreased under drought stress condition. These factors are related to average leaf length, and shoot dry weight. Further, there was a positive correlation between chlorophyll content (SPAD) and high transpiration efficiency, wherein SPAD could be utilized as an indicator of drought tolerance (
Fotovat et al. 2007). Therefore, the calculation of DTI with morphological traits such as plant height, average leaf length, dry weight, and leaf chlorophyll weight can reflect drought tolerance.
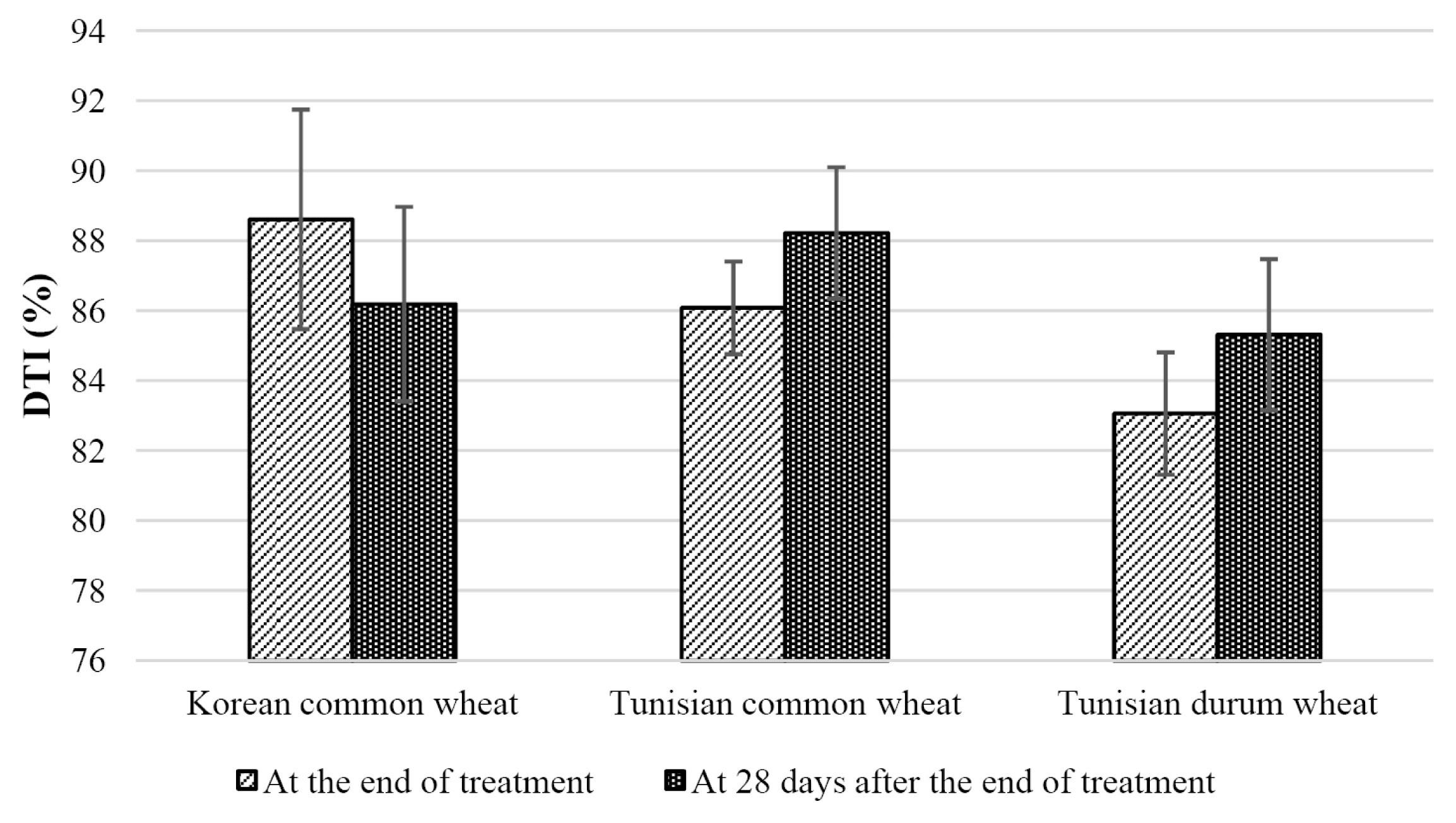
Korean wheat and Tunisian wheat showed different DTIs at the end of treatment and 28 days after the end of treatment (
Fig. 4). The DTI of Korean wheat cultivars was higher at the end of treatment than at 28 days after the end of treatment. However, the DTI of Tunisian common wheat and durum wheat cultivars were lower at the end of treatment than at 28 days after the end of treatment. This indicates that the tolerance to drought stress and the ability to restore must be considered separately.
Costa et al. (1997) reported that the stress index between continuous stress and transient stress was different. Therefore, separate strategies should be used for these cultivars to improve drought tolerance.
The dendrogram showed distant relationship between the Korean and Tunisian cultivars (
Fig. 3). Environmental conditions could affect genetic diversity (
Li et al. 2012). Consequently, the genetic distance of Korean common wheat and Tunisian common wheat and durum wheat reflects the environmental differences between Korea and Tunisia.
In Tunisian cultivars, common wheat and durum wheat were completely mixed within the clusters, indicating that the SSR markers used in this study can be applied to both common wheat and durum wheat. In addition, both clusters A and B were categorized into some groups with drought tolerance. This suggests that the cluster analysis using SSR markers can be useful in evaluating drought tolerance in common wheat and durum wheat.
In conclusion, morphological traits and SSR markers could be used in evaluating drought tolerance in Korean and Tunisian wheat cultivars. It is necessary to establish an accurate standard to effectively evaluate tolerance to drought stress for wheat improvement. The results obtained in this study could help increase genetic resources and improve breeding programs for drought tolerance in wheat.
ACKNOWLEDGMENT
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MEST) (No. 2012K1A3A1A09028123) and carried out with the support of “Cooperative Research Program for Agriculture Science & Technology Development (Project title: Development of high yielding wheat with stress tolerance via molecular breeding strategies, Project No. PJ008031)”, Rural Development Administration, Republic of Korea.
Fig. 1Average plant height (A) and average leaf length (B) of Korean common wheat cultivars (a), Tunisian common wheat cultivars (b), and Tunisian durum wheat cultivars (c). Control (closed symbols) and treatment (open symbols).
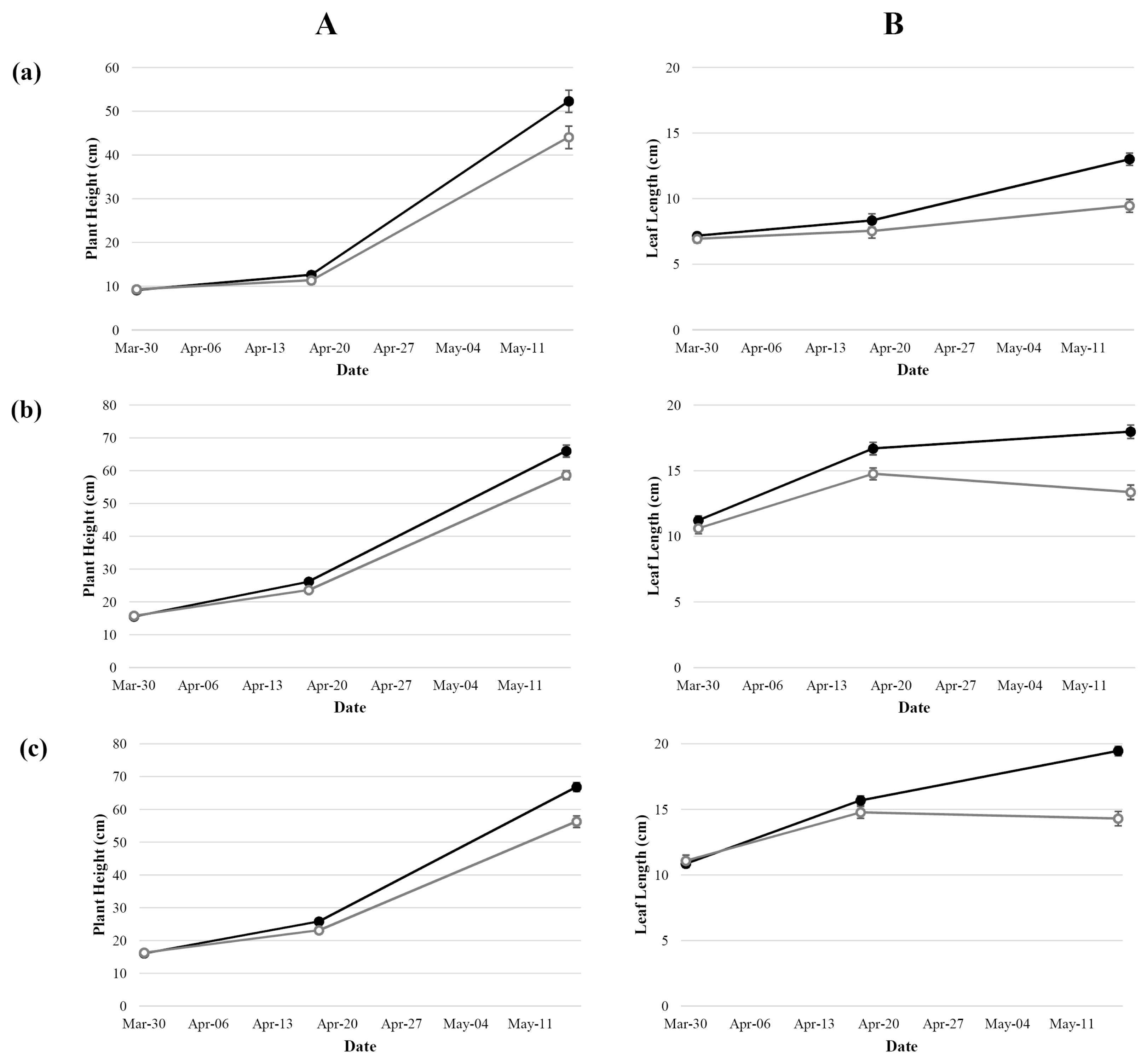
Fig. 2Average leaf chlorophyll content of Korean common wheat cultivars (a), Tunisian common wheat cultivars (b), and Tunisian durum wheat cultivars (c), and shoot dry weight of each cultivar (d).
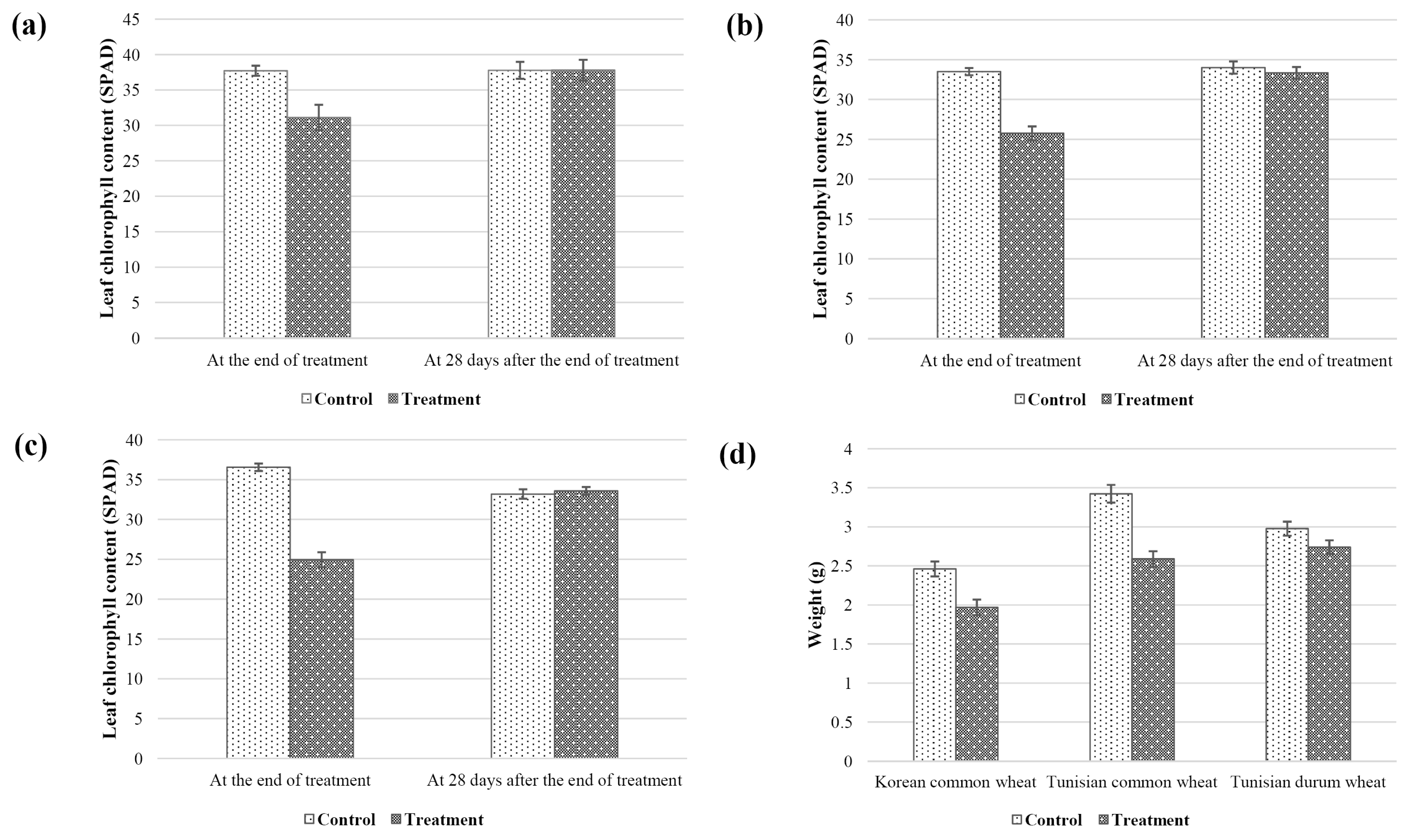
Fig. 3SSR-based genetic relationship among 77 wheat cultivars.
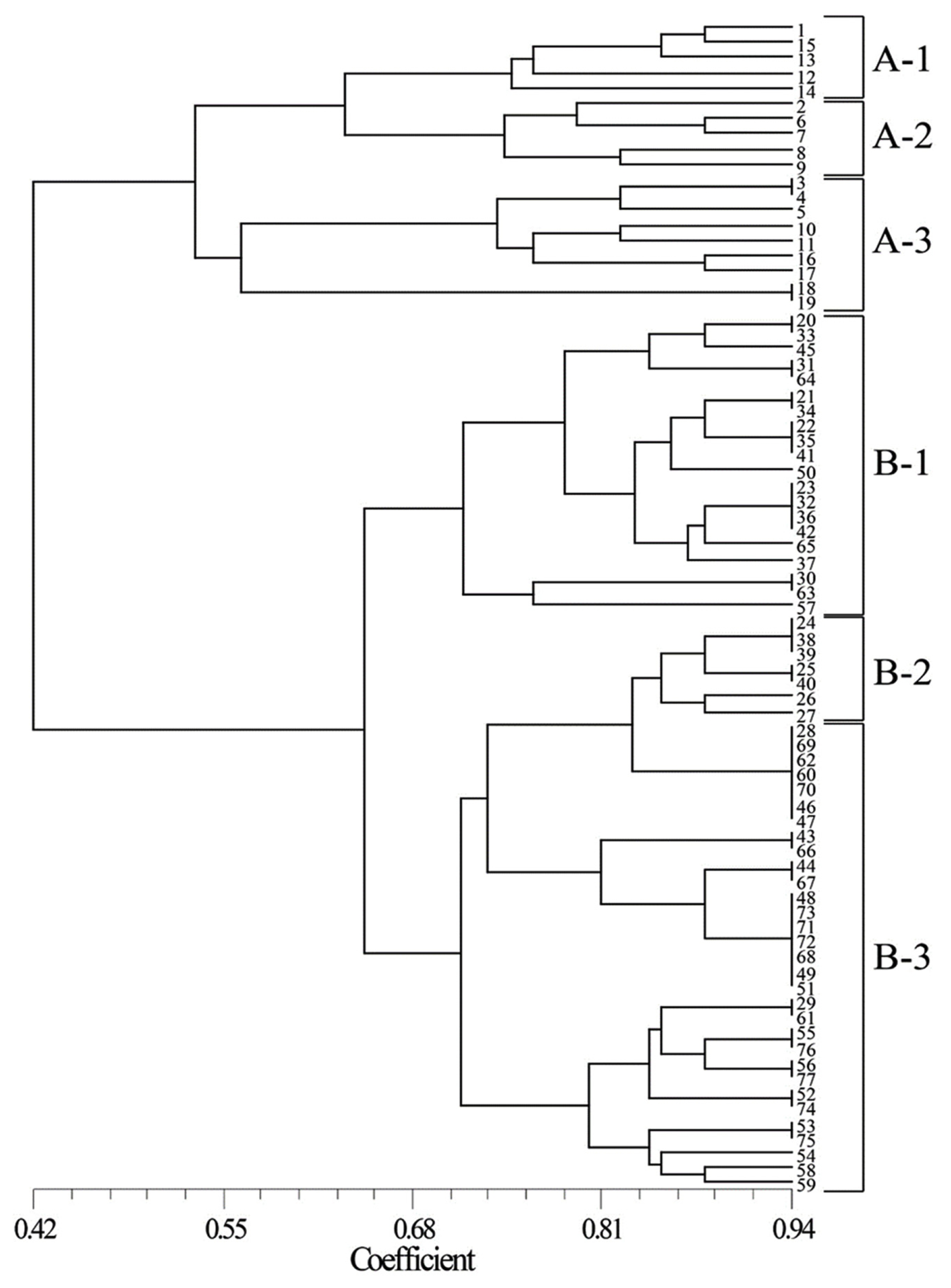
Fig. 4Drought tolerance index (DTI) at the end of treatment and at 28 days after the end of treatment.
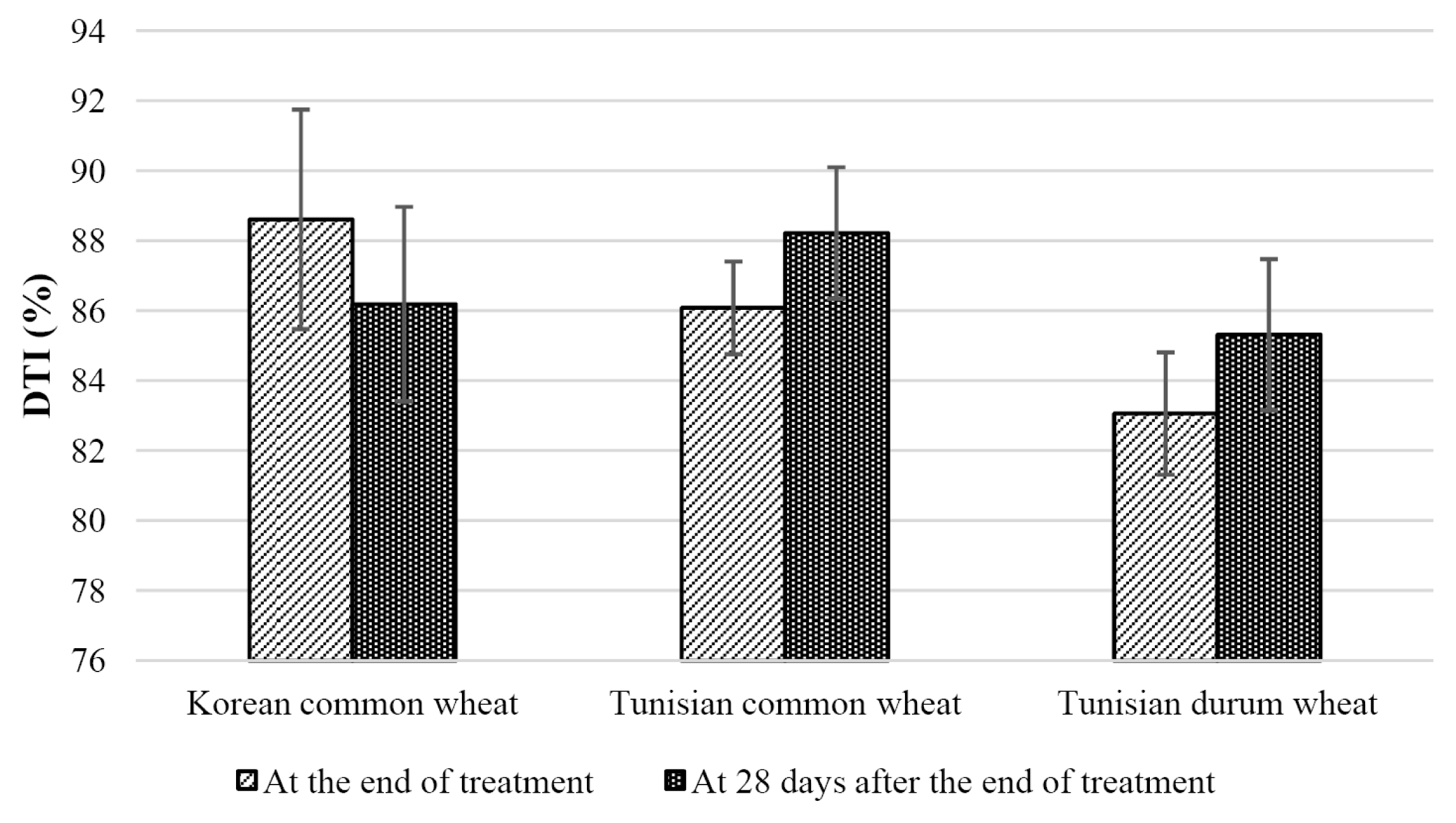
Table 1The list of wheat cultivars used in this study.
Table 1
|
Entry No. |
Name |
Species |
Location |
Supplier |
Accession Number |
|
1 |
Chunggyemil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-4z)
|
|
2 |
Olgeurumil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-9z)
|
|
3 |
Alchanmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-10z)
|
|
4 |
Gobunmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-11z)
|
|
5 |
Keumgangmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-12z)
|
|
6 |
Seodunmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-14z)
|
|
7 |
Saealmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-13z)
|
|
8 |
Jinpummil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-15z)
|
|
9 |
Joeunmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-17z)
|
|
10 |
Anbaekmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-18z)
|
|
11 |
Sinmichalmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-20z)
|
|
12 |
Jonongmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-22z)
|
|
13 |
Jokyoungmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-23z)
|
|
14 |
Younbaekmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-24z)
|
|
15 |
Sukangmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-29z)
|
|
16 |
Hanbaekmil |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-30z)
|
|
17 |
Sooan |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-33z)
|
|
18 |
Dajung |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-36z)
|
|
19 |
Goso |
T. aestivum L. |
Korea |
NICS, RDA |
01-0006-35z)
|
|
20 |
ZAAFRANE |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205894y)
|
|
21 |
U14014 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205910y)
|
|
22 |
U14018 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205912y)
|
|
23 |
U14021 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205915y)
|
|
24 |
U14038 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205922y)
|
|
25 |
U14040 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205923y)
|
|
26 |
U14045 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205926y)
|
|
27 |
U14046 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205927y)
|
|
28 |
U14054 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205933y)
|
|
29 |
U14057 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205935y)
|
|
30 |
U14076 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 205938y)
|
|
31 |
FRIGUI 2 |
T. aestivum L. |
Tunisia |
NAC, RDA |
IT 229475y)
|
|
32 |
U14010 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205908y)
|
|
33 |
U14031 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205917y)
|
|
34 |
U14050 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205929y)
|
|
35 |
U14053 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205932y)
|
|
36 |
U14055 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205934y)
|
|
37 |
U14078 |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205940y)
|
|
38 |
SINLIKAT |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205941y)
|
|
39 |
AGIM SINLIKA |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205943y)
|
|
40 |
MELANGE |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205944y)
|
|
41 |
BEDI |
T. turgidum L. subsp. Durum
|
Tunisia |
NAC, RDA |
IT 205947y)
|
|
42 |
Allorca |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 41032x)
|
|
43 |
Florence Aurore |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 150608x)
|
|
44 |
Florence Aurore |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 150609x)
|
|
45 |
Koudiat A2 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 174665x)
|
|
46 |
Florence 135 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 185357x)
|
|
47 |
Mahon 73 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 185372x)
|
|
48 |
Mahon 124 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 185373x)
|
|
49 |
Roussla |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 189775x)
|
|
50 |
Cailloux |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 191446x)
|
|
51 |
Mahon 73 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 191734x)
|
|
52 |
Irakie 231 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 191738x)
|
|
53 |
Mahon 124 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 191929x)
|
|
54 |
Mahon 73 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 192024x)
|
|
55 |
Cailloux |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 304915x)
|
|
56 |
Baroota 52 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 410889x)
|
|
57 |
Soltane |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 428416x)
|
|
58 |
Ariana 66 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 519955x)
|
|
59 |
BT 2288 |
T. aestivum L. |
Tunisia |
NPGS, USDA |
PI 559656x)
|
|
60 |
Huguenot Bariole 360 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 174646x)
|
|
61 |
Jenah Retifah 24 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 184537x)
|
|
62 |
Mekki 13 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 185194x)
|
|
63 |
Sbei 7 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 185195x)
|
|
64 |
Ble Dur 116 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 189772x)
|
|
65 |
Ble Dur 870 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 189773x)
|
|
66 |
Jenah Retifah 1 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 192516x)
|
|
67 |
Jenah Retifah 24 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 192517x)
|
|
68 |
Sbei 272 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 278383x)
|
|
69 |
Mahmoudi |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 306573x)
|
|
70 |
Inrat 69 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 324939x)
|
|
71 |
Maghrebi |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 422313x)
|
|
72 |
Roussia |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 428434x)
|
|
73 |
Maghrebi 72 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 428462x)
|
|
74 |
Amal 72 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 433749x)
|
|
75 |
Badri |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 433751x)
|
|
76 |
Maghrebi 72 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 433758x)
|
|
77 |
Maghrebi 72 |
T. turgidum L. subsp. Durum
|
Tunisia |
NPGS, USDA |
PI 520062x)
|
Table 2The list of SSR markers used in this study.
Table 2
|
Name |
F Primer |
R Primer |
Annealing Temperature (°C) |
|
Xbarc108
|
GCGGGTCGTTTCCTGGAAATTCATCTAA |
GCGAAATGATTGGCGTTACACCTGTTG |
50 |
|
Xbarc121
|
ACTGATCAGCAATGTCAACTGAA |
CCGGTGTCTTTCCTAACGCTATG |
50 |
|
Xgwm573
|
AAGAGATAACATGCAAGAAA |
TTCAAATATGTGGGAACTAC |
50 |
|
Xgwm130
|
AGCTCTGCTTCACGAGGAAG |
CTCCTCTTTATATCGCGTCCC |
60 |
|
Xgwm332
|
AGCCAGCAAGTCACCAAAAC |
AGTGCTGGAAAGAGTAGTGAAGC |
60 |
|
Xgwm635
|
TTCCTCACTGTAAGGGCGTT |
CAGCCTTAGCCTTGGCG |
60 |
|
Xwmc233
|
GACGTCAAGAATCTTCGTCGGA |
ATCTGCTGAGCAGATCGTGGTT |
61 |
|
Xwmc9
|
AACTAGTCAAATAGTCGTGTCCG |
GTCAAGTCATCTGACTTAACCCG |
61 |
|
Xwmc596
|
TCAGCAACAAACATGCTCGG |
CCCGTGTAGGCGGTAGCTCTT |
61 |
|
Xwmc603
|
ACAAACGGTGACAATGCAAGGA |
CGCCTCTCTCGTAAGCCTCAAC |
61 |
|
Xwmc695
|
GAGGGCACCTCGTAAGTTGG |
GGCAGGAGCCCCTACAAGAT |
61 |
|
Xwmc65
|
TGGATGGGAAGGAGAATAAGTG |
ATCCAACCGGAACTACCGTCAG |
61 |
|
Xgwm260
|
GCCCCCTTGCACAAATC |
CGCAGCTACAGGAGGCC |
55 |
|
Xgwm350
|
ACCTCATCCACATGTTCTACG |
GCATGGATAGGACGCCC |
55 |
|
Xgwm276
|
ATTTGCCTGAAGAAAATATT |
AATTTCACTGCATACACAAG |
55 |
|
Xwmc17
|
ACCTGCAAGAAATTAGGAACTC |
CTAGTGTTTCAAATATGTCGGA |
51 |
|
Xwmc182
|
GTATCTCACGAGCATAACACAA |
GAAAGTGTATGGATCATTAGGC |
51 |
Table 3Drought tolerance trait indices (DTTIs) at 21 days after drought treatment.
Table 3
|
Group |
Plant Height (%) |
Average Leaf Length (%) |
No. of Tillers (%) |
Leaf Chlorophyll Content (%) |
|
Korean common wheat |
88.5 ± 3.70 |
94.7 ± 5.49 |
109.1 ± 4.33 |
82.7 ± 4.57 |
|
Tunisian common wheat |
87.9 ± 2.67 |
93.6 ± 2.50 |
98.3 ± 6.63 |
76.8 ± 2.17 |
|
Tunisian durum wheat |
88.7 ± 2.18 |
91.9 ± 2.56 |
91.6 ± 4.39 |
68.6 ± 2.74 |
|
|
Total |
88.3 ± 1.57 |
93.2 ± 1.88 |
98.5 ± 3.27 |
75.3 ± 1.82 |
Table 4Drought tolerance trait indices (DTTIs) at 28 days after re-watering.
Table 4
|
Group |
Plant Height (%) |
Average Leaf Length (%) |
No. of Tillers (%) |
Leaf Chlorophyll Content (%) |
|
Korean common wheat |
82.1 ± 4.21 |
76.1 ± 5.8 |
154 ± 15.02 |
100.3 ± 2.83 |
|
Tunisian common wheat |
86.9 ± 3.59 |
79.3 ± 4.03 |
105.6 ± 8.24 |
98.5 ± 1.74 |
|
Tunisian durum wheat |
83.1 ± 2.99 |
70.9 ± 4.11 |
146.8 ± 11.03 |
101.9 ± 2.14 |
|
|
Total |
84.3 ± 2.04 |
75.5 ± 2.6 |
132.5 ± 6.72 |
100.2 ± 1.24 |
Table 5Drought tolerance trait indices (DTTIs) for shoot dry weight and days to flowering.
Table 5
|
Group |
Shoot Dry Weight (%) |
Days to Flowering (%) |
|
Korean common wheat |
83.2 ± 6.09 |
100.5 ± 1.00 |
|
Tunisian common wheat |
76.8 ± 3.04 |
100.9 ± 0.41 |
|
Tunisian durum wheat |
92.7 ± 2.39 |
101.5 ± 0.51 |
|
|
Total |
84.2 ± 2.22 |
101.0 ± 0.35 |
Table 6Drought tolerance grouping of wheat cultivars based on drought tolerance index (DTI).
Table 6
|
Drought Tolerance Category |
Range of DTI |
No. of Cultivars |
Entry No. (Name) |
|
Resistant |
>90% |
23 |
1 (Chunggyemil), 4 (Gobunmil), 5 (Keumgangmil), 14 (Younbaekmil), 16 (Hanbaekmil), 17 (Sooan), 18 (Dajung), 19 (Goso), 22 (U14018), 23 (U14021), 26 (U14045), 27 (U14046), 31 (FRIGUI 2), 32 (U14010), 34 (U14050), 35 (U14053), 36 (U14055), 37 (U14078), 39 (AGIM SINLIKA), 40 (MELANGE), 41 (BEDI), 42 (Allorca), 45 (Koudiat A2) |
|
Moderate |
80–90% |
33 |
3 (Alchanmil), 8 (Jinpummil), 10 (Anbaekmil), 11 (Sinmichalmil), 12 (Jonongmil), 13 (Jokyoungmil), 15 (Sukangmil), 20 (ZAAFRANE), 21 (U14014), 24 (U14038), 25 (U14040), 30 (U14076), 33 (U14031), 38 (SINLIKAT), 47 (Mahon 73), 48 (Mahon 124), 49 (Roussla), 50 (Cailloux), 54 (Mahon 73), 55 (Cailloux), 56 (Baroota 52), 57 (Soltane), 60 (Huguenot Bariole 360), 61 (Jenah Retifah 24), 62 (Mekki 13), 63 (Sbei 7), 64 (Ble Dur 116), 65 (Ble Dur 870), 66 (Jenah Retifah 1), 68 (Sbei 272), 71 (Maghrebi), 75 (Badri), 76 (Maghrebi 72) |
|
Susceptible |
<80% |
21 |
2 (Olgeurumil), 6 (Seodunmil), 7 (Saealmil), 9 (Joeunmil), 28 (U14054), 29 (U14057), 43 (Florence Aurore), 44 (Florence Aurore), 46 (Florence 135), 51 (Mahon 73), 52 (Irakie 231), 53 (Mahon 124), 58 (Ariana 66), 59 (BT 2288), 67 (Jenah Retifah 24), 69 (Mahmoudi), 70 (Inrat 69), 72 (Roussia), 73 (Maghrebi 72), 74 (Amal 72), 77 (Maghrebi 72) |
Table 7The most resistant cultivars to drought stress.
Table 7
|
Entry No. |
Name |
Group |
DTI (%) |
|
14 |
Younbaekmil |
Korean common wheat |
104.7 |
|
34 |
U14050 |
Tunisian durum wheat |
102.6 |
|
42 |
Allorca |
Tunisian common wheat |
113.6 |
|
45 |
Koudiat A2 |
Tunisian common wheat |
106.8 |
References
- Ali Z, Salam A, Azhar FM, Khan IA. 2007. Genotypic variation in salinity tolerance among spring and winter wheat (Triticum aestivum L.) accessions. South Afr J Bot. 73: 70-75.
- Anjum SA, Xie XY, Wang LC, Saleem MF, Man C, Lei W. 2011. Morphological, physiological and biochemical responses of plants to drought stress. Afr J Agric Res. 6: 2026-2032.
- Brown ME, Funk CC. 2008. Food security under climate change. Science. 319: 580-581.
- Castillo EG, Tuong TP, Ismail AM, Inubushi K. 2007. Response to salinity in rice: Comparative effects of osmotic and ionic stresses. Plant Prod Sci. 10: 159-170.
- Ciucă M, Petcu E. 2009. SSR markers associated with membrane stability in wheat (Triticum aestivum L.). Romanian Agric Res. 26: 21-24.
- Costa LD, Vedove GD, Gianquinto G, Giovanardi R, Peressotti A. 1997. Yield, water use efficiency and nitrogen uptake in potato: influence of drought stress. Potato Res. 40: 19-34.
- El Siddig MA, Baenziger S, Dweikat I, El Hussein AA. 2013. Preliminary screening for water stress tolerance and genetic diversity in wheat (Triticum aestivum L.) cultivars from Sudan. J Genet Eng Biotechnol. 11: 87-94.
- Farshadfar E, Jamshidi B, Cheghamirza K, da Silva JAT. 2012. Evaluation of drought tolerance in bread wheat (Triticum aestivum L.) using in vivo and in vitro techniques. Ann Biol Res. 3: 465-476.
- Fotovat R, Valizadeh M, Toorchi M. 2007. Association between water-use efficiency components and total chlorophyll content (SPAD) in wheat (Triticum aestivum L.) under well-watered and drought stress conditions. J Food Agric Environ. 5: 225-227.
- Foulkes MJ, Scott RK, Sylvester-Bradley R. 2002. The ability of wheat cultivars to withstand drought in UK conditions: formation of grain yield. J Agric Sci. 138: 153-169.
- Gupta PK, Varshney RK. 2000. The development and use of microsatellite markers for genetic analysis and plant breeding with emphasis on bread wheat. Euphytica. 113: 163-185.
- Jaleel CA, Manivannan P, Wahid A, Farooq M, Al-Juburi HJ, Somasundaram R, Panneerselvam R. 2009. Drought stress in plants: a review on morphological characteristics and pigments composition. Int J Agric Biol. 11: 100-105.
- Korea Seed & Variety Service. 2014. http://www.seed.go.kr/
- Li XJ, Xu X, Yang XM, Li XQ, Liu WH, Gao AN, Li LH. 2012. Genetic diversity of the wheat landrace Youzimai from different geographic regions investigated with morphological traits, seedling resistance to powdery mildew, gliadin and microsatellite markers. Cereal Res Commun. 40: 95-106.
- Lobell DB, Burke MB, Tebaldi C, Mastrandrea MD, Falcon WP, Naylor RL. 2008. Prioritizing climate change adaptation needs for food security in 2030. Science. 319: 607-610.
- Munns R, James RA, Läuchli A. 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J Exp Bot. 57: 1025-1043.
- Pan XY, Wang YF, Wang GX, Cao QD, Wang J. 2002. Relationship between growth redundancy and size inequality in spring wheat population mulched with clear plastic film. Acta Phytoecol Sinica. 26: 177-184.
- Quarrie SA, Dodig D, Pekiç S, Kirby J, Kobiljski B. 2003. Prospects for marker-assisted selection of improved drought responses in wheat. Bulg J Plant Physiol. 2003. 83-95.
- Rai MK, Kalia RK, Singh R, Gangola MP, Dhawan AK. 2011. Developing stress tolerant plants through in vitro selection—An overview of the recent progress. Environ Exp Bot. 71: 89-98.
- Rajaram S. 2001. Prospects and promise of wheat breeding in 21st century. Euphytica. 119: 3-15.
- RDA. 2014. RDA-Genebank Information Center. http://www.genebank.go.kr/
- Shahzad A, Ahmad M, Iqbal M, Ahmad I, Ali GM. 2012. Evaluation of wheat landrace genotypes for salinity tolerance at vegetative stage by using morphological and molecular markers. Genet Mol Res. 11: 679-692.
- Takeda S, Matsuoka M. 2008. Genetic approaches to crop improvement: responding to environmental and population changes. Nat Rev Genet. 9: 444-457.
- USDA. 2014. The Agricultural Research Service - Germplasm Resources Information Network (ARS-GRIN). http://www.ars-grin.gov/
- USDA. 2014. The Agricultural Research Service - The GrainGenes 2.0. http://wheat.pw.usda.gov/
- Veesar NF, Channa AN, Rind MJ, Larik AS. 2007. Influence of water stress imposed at different stages on growth and yield attributes in bread wheat genotypes Triticum aestivum L. Wheat Inform Ser. 104: 15-19.
- Yıldırıma M, Kılıçb H, Kendalb E, Karahan T. 2011. Applicability of chlorophyll meter readings as yield predictor in durum wheat. JPlant Nutri. 34: 151-164.
- Zhu JK. 2001. Plant salt tolerance. Trends Plant Sci. 6: 66-71.