Abstract
The functions of DUF640/ALOG (Arabidopsis LSH1 and Oryza G1) domain proteins, which are found in most land plants, have not been well characterized, but some of these proteins regulate inflorescence architecture in rice and specify organ boundaries in Arabidopsis. Arabidopsis DUF640-domain genes are initially identified as LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes. Chinese cabbage leaves have large, white midribs and photosynthetic leaf blades (or lamina). A DUF640 domain gene of Brassica rapa, BrLSH2, is specifically expressed in the midrib of Chinese cabbage. Arabidopsis and rice possess ten LSH family genes, but B. rapa has 24 LSH genes, which can be categorized into two or four groups based on sequence identity. Here, we examined the expression patterns of the LSHs in various Brassica species and analyzed the promoter sequence of the BrLHS2 gene. The transcript levels of most LSH genes were very high in the midrib but low in the leaf blade. These genes were evenly expressed throughout the petiole region of Korean cabbage and highly expressed in the leaf base region near the stem and in the border area in B. oleracea. In addition, BrLSHs were expressed in both bundle and mesophyll cells of the midrib. These expression patterns suggest the possible use of these genes to generate leafy vegetables with altered leaf morphology. The BrLSH2 promoter, which contains auxin- and cytokinin-responsive elements as well as leaf development-related elements, may confer midrib-specific expression, suggesting that this promoter may be useful for the production of midrib-targeted transgenic Chinese cabbage.
-
Key words: ALOG, DUF640, Leaf boundary, Leaf morphology, Midrib
INTRODUCTION
DUF640 (domain of unknown function 640) domain genes, or ALOG (
Arabidopsis LSH1 and
Oryza G1) family genes, are found in land plants. DUF640 (InterPro: IPR006936) is also found in plant proteins including resistance protein-like protein. Ten ALOG family genes are present in rice and
Arabidopsis (
Yoshida et al. 2009;
Cho et al. 2011). All members of this family have a highly conserved region and a nuclear localization signal (NLS), KKRK, flanking the C-terminal region of this domain. Although the precise function of the DUF640 motif (or ALOG) is still unclear, the roles of some of these genes have been studied in rice,
Arabidopsis and tomato.
ALOG genes regulate inflorescence architecture in rice and tomato (
Yoshida et al. 2009,
2013;
MacAlister et al. 2012;
Yan et al. 2013), while they determine organ boundaries and influence light sensitivity in
Arabidopsis (
Zhao et al. 2004;
Cho and Zambryski 2011;
Takeda et al. 2011).
Rice
LONG STERILE LEMMA (
G1), the first identified
ALOG gene from monocots, specifies the identity of the sterile lemma by repressing lemma identity
via the regulation of downstream target genes (
Yoshida et al. 2009). Another rice DUF640 domain gene,
TRIANGULAR HULL1/BEAK- SHAPED GRAIN 1 (
TH1/BSG1), determines grain shape and size by regulating cell division and extension of the lemma and palea (
Li et al. 2012;
Yan et al. 2013).
TAWAWA1 (
TAW1) encodes an ALOG family protein in rice and regulates inflorescence architecture, partly through promoting the expression of
SHORT VEGETATIVE PHASE (
SVP)-like genes (
Yoshida et al. 2013). Tomato
TERMINATING FLOWER (
™F), like rice
TAW1, affects inflorescence organization (
MacAlister et al. 2012), possibly by preventing the early expression of orthologous genes of
Arabidopsis UNUSUAL FLORAL ORGANS (
UFO),
LEAFY (
LFY),
APETALLA 1 (
AP1) and
SEPALLATA (
SEP), which contribute to promoting floral fate (
Teo et al. 2014).
Arabidopsis contains ten
ALOG or
LSH (LIGHT-SENSITIVE HYPOCOTYLS) genes.
LSH1 was first identified as
Light-dependent
Short
Hypocotyl 1 (
Zhao et al. 2004) and later renamed “Light-Sensitive Hypocotyls”.
LSH1 is expressed in hypocotyls, shoot apices and lateral root primordia in
Arabidopsis, and its overexpression leads to the hypersensitive response to various light wavelengths, with plants exhibiting short hypocotyls and enlarged cotyledons (
Zhao et al. 2004).
LSH3 and
LSH4, known as
ORGAN BOUNDARY 1 (
OBO1) and
OBO4, are
Arabidopsis ALOG family genes that are expressed at the boundary of the shoot apical meristem (SAM) and lateral organs (
Cho et al. 2011;
Takeda et al. 2011).
LSH3 and
LSH4 are activated by
CUP-SHAPED COTYLDON 1 (
CUC1) (
Takeda et al. 2011) and might interact with UFO; constitutive expression of
LSH3 and
LSH4 partially mimics the phenotype exhibited by mutations in
UFO [similar to the interaction between
™F and
ANANTHA (
AN), an ortholog of
Arabidopsis UFO] (
Teo et al. 2014). Taken together, these observations suggest that ALOG family proteins are important regulators that affect inflorescence architecture through mediating the transition status of SAM differentiation due to their roles in maintaining meristem indeterminacy and suppressing floral identity, together with the unique expression patterns of
LSH and
™F genes (
Teo et al. 2014).
Chinese cabbage (
Brassica rapa ssp.
pekinensis) is one of the most important leafy vegetables in Asian countries including Korea. Compared to other plants, the leaf morphology of Chinese cabbage is quite peculiar, comprising a large midrib with a leaf blade but lacking a petiole when mature (
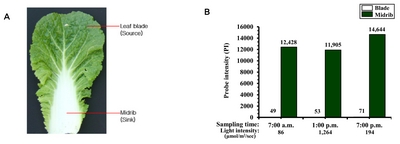
Fig. 1A). The outer leaf of mature Chinese cabbage, i.e., the leaf blade and midrib, can be divided into source and sink tissues, respectively, with respect to photosynthesis. The
B. rapa LSH2 gene (Brapa_ESTC024477 for Br300K array = Bra040188 for
http://brassicadb.org/brad/) is specifically expressed in midribs throughout the day (
Fig. 1B). In the current study, we examined the expression patterns of
LSH family genes in various
Brassica species. In addition, we discuss the possible application of these genes to biotechnology.
MATERIALS AND METHODS
Plant materials
Seeds of the Brassica rapa inbred lines ‘Chiifu’ and ‘Kenshin’ were obtained from Korea Brassica rapa Genome Resource Bank (KBGRB), and other seeds were purchased from a local market. The plants were grown in a greenhouse at Chungnam National University. Leaf tissues were sampled, frozen in liquid nitrogen and stored at −70°C until use.
RNA isolation and microarray analysis
Total RNA was isolated from midrib and leaf blade tissues using an Easy-BLUE™ Total RNA Extraction Kit (Invitrogen, U.S.A.) and purified using an RNeasy MinElute™ Cleanup Kit (Qiagen, Germany). Microarray experiments were carried out as described by
Dong et al. (2013); the full results of the analysis have not yet been published. To assess the reproducibility of the microarray analysis, the experiment was repeated twice with total RNAs from independent cultures and treatments. Normality of Cy3 intensities was tested with the qqline function in R Statistical Software. Data were subsequently normalized with cubic spline normalization using quantiles to adjust signal variations among chips from the robust multi-chip analysis and a median polish algorithm implemented in NimbleScan. Finally, the average value from the perfect match (PM) values of six probes was used to select responsive genes.
The total RNA from each plant sample amounting to 5 μg was combined with random hexamer primers in a Super Script first-strand cDNA synthesis system according to the manufacturer’s instructions (Invitrogen, U.S.A.). Complementary DNA was diluted 10-fold, and 1 μl of the diluted cDNA was used in each 20 μl PCR mixture. RT-PCR primers were designed using sequence information from
Arabidopsis thaliana,
B. rapa and
B. oleracea (
Table 1). Primer sequences used in this experiment are listed in
Table 2, and the primer set used for the control was designed based on
B. rapa actin1 (
BrACT1) (Forward sequence = 5′-ACACC-ATGATGTCTTGGCCTACCA and reverse sequence = 5′-AATGGTACCGGAATGGTCAAGGCT). Standard PCR was performed, with 5 min denaturation at 94°C followed by 25 cycles of 94°C for 30 s, 54°C for 30 s and 72°C for 60 s. The PCR products were analyzed following electrophoresis on a 1.5% agarose gel.
Multiple sequence alignments were conducted using ClustalX2.0 (
Larkin et al. 2007). The phylogenetic tree, based on the
LSH coding sequences (CDs), was constructed using the neighbor-joining (NJ) method. NJ trees were constructed using MEGA6 (
http://www.megasoftware.net/) with the ‘pairwise deletion’ option and ‘Kimura 2-Parameter’ model, with a bootstrap test of 1,000 replicates (
Tamura et al. 2013).
To analyze
cis-acting elements of
BrLSH2, a ca. 3,200 bp sequence upstream of the
LSH2 ATG start codon, which was obtained from BRAD (
http://brassicadb.org/), was analyzed with PlantPAN (
Chang et al. 2008) and PLACE (
Higo et al. 1999). Several important motifs are indicated in the figure.
RESULTS
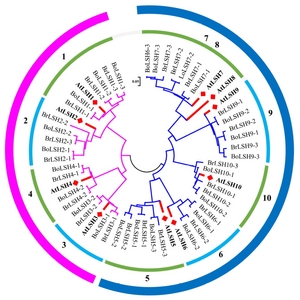
LSH genes from B. rapa and B. oleracea
A total of 24
B. rapa LSH (
BrLSH) genes corresponding to ten
Arabidopsis genes were found in the BRAD database (
http://brassicadb.org/brad/), with no ortholog of
AtLSH8 detected (
Table 1). A similar number of
LSH genes may be present in another
Brassica species,
B. oleracea. To examine the relationship among the members of the
LSH multigene family, we constructed an NJ tree using ClustalX2.0 and MEGA6 software (
Larkin et al. 2007;
Tamura et al. 2013) (
Fig. 1). As shown in
Figure 1, all
LSH family genes could be divided into two large groups; one group includes
LSH1, 2, 3 and
4, while the other includes
LSH5, 6, 7, 8, 9 and
10. The genes were further divided into four groups:
LSH1 and
2;
LSH3 and
4;
LSH5 and
6; and
LSH7, 8, 9 and
10. This classification reflects both the functional redundancy and divergence of the
LSH gene family.
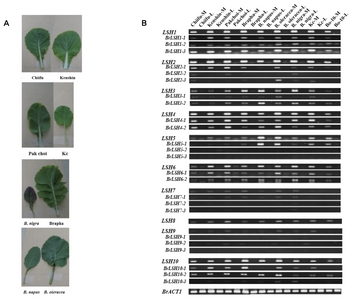
To examine the expression characteristics of
LSH genes from diverse
Brassica species, we performed RT-PCR analysis with a common primer set and an allele-specific primer set for each
LSH gene of
B. rapa (
Table 2 and
Fig. 3). The common primer set could be applied to all
Brassica species because it was designed from a highly conserved region among
Brassica genes, including
Arabidopsis sequences. As shown in
Figure 3,
LSH genes were specifically induced in the midribs of all
Brassica species examined. However,
LSH5-2, LSH5-3, LSH7s and
LSH9s were expressed at very low levels. Unexpectedly, although
LSH8 is not present in the
B. rapa genome, we detected
LSH8 transcripts in this species, implying possible contamination of other
LSH gene products, such as
LSH7, which is most closely related to
LSH8 (
Fig. 2).
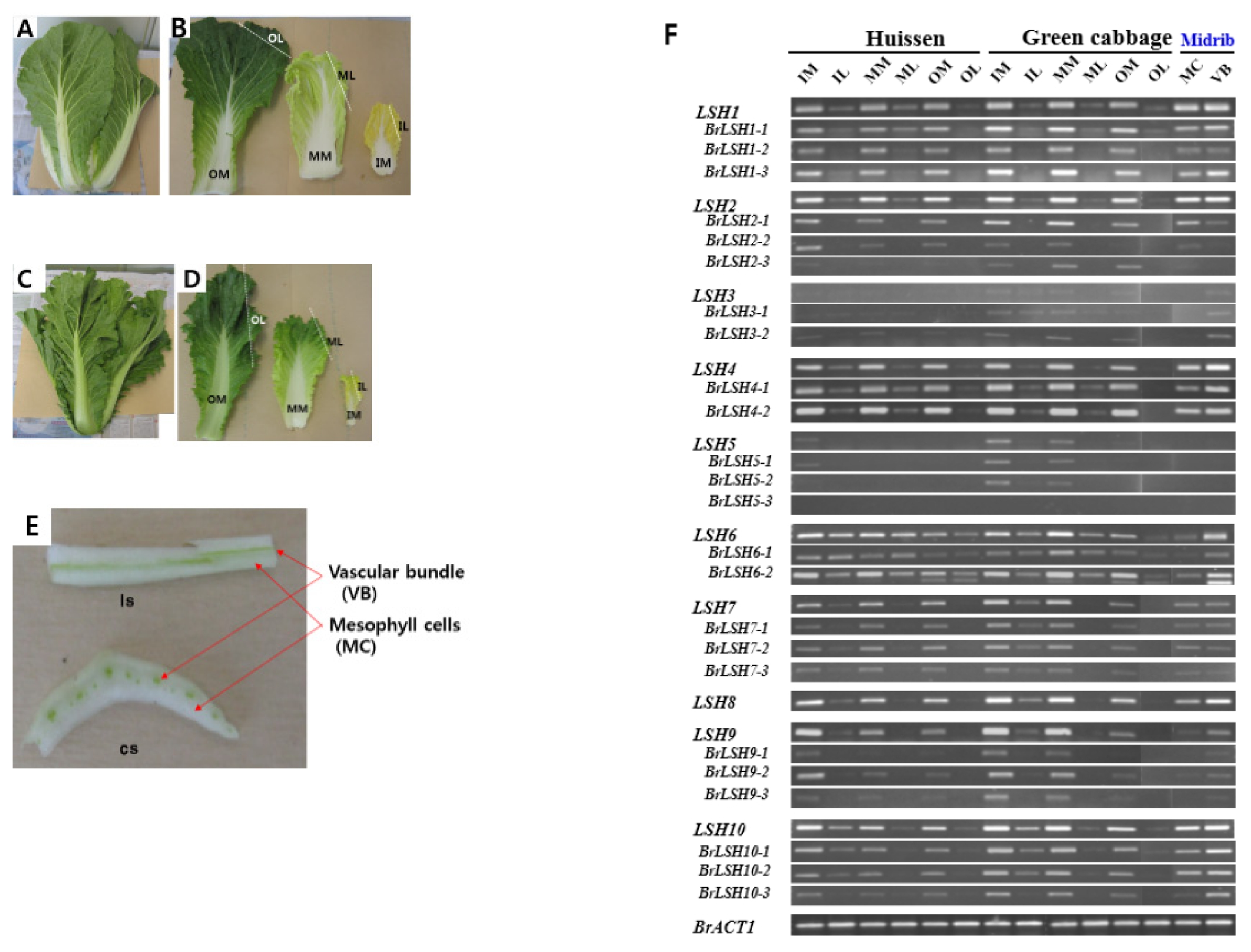
To investigate whether the expression of
LSH genes occurs in photosynthetic or sink tissues, various tissues of
B. rapa were subjected to RT-PCR analysis (
Fig. 4). Outer leaves, middle leaves and inner non-photosynthetic leaves from ‘Heissen’ (F
1 Korean cultivar) and green cabbage (F
1 Chinese cultivar) were sampled when they formed heads. In addition, the midribs of ‘Heissen’ leaves were further dissected into vascular bundles and mesophyll cells.
LSH genes were predominantly expressed in midribs from all parts of the plant. There was no difference in
LSH gene expression between mesophyll cells and vascular bundles, while
LSH6 was more highly expressed in vascular bundles than in mesophyll cells. Detection of
LSH transcripts in blades may have been due to the presence of veins in this tissue or PCR cycles. These results indicate that the expression of
LSH is not related to the photosynthetic status of the tissue but is, instead, midrib-specific.
To examine
LSH gene expression in different parts of the petiole, we performed RT-PCR analysis of Korean cabbage leaves, which have relatively long petiole-like midribs (
Fig. 5). As shown in
Figure 5,
LSH genes were evenly expressed throughout all areas of the petiole region but not in the blade. The levels of
LSH3 and
LSH9 expression were relatively low, indicating that different
LSH genes are expressed at different levels in various
Brassica species.
Changes in leaf morphology in
B. oleracea are quite dramatic throughout development, ranging from long petioles in young seedlings to blade-like petioles to no petiole-like structures during head formation (left panel of
Fig. 6). The expression levels of
LSHs were high in the midrib and petiole and relatively low in the leaf blade. The most interesting finding was that we could detect expression of
LSHs in both the midrib and leaf blade in samples harvested from the bottom halves of leaves. In particular,
LSH1 and
LSH2 were highly expressed in the 6
th to 13
th leaf blade portions (
Bo-2L,
Bo-3L and
Bo-4L) in the bottom halves of leaves. These results indicate that the
LSH genes are highly expressed in the leaf base region near the stem or border area in
B. oleracea.
To further dissect the gene expression patterns in
B. oleracea, we separated a cabbage leaf into 14 sections and subjected the sections to RT-PCR analysis (
Fig. 7). All
LSH genes were highly expressed in the midrib, and the expression of most of the genes was relatively high in the basal parts of the leaf blade. Only low or very low transcript levels were detected in parts 9 and 10. Compared to
B. rapa, cabbage leaves contain relatively large veins derived from the midrib, which may affect the expression of
LSH. These results, along with those shown in
Figure 6, indicate that
LSH genes are highly expressed in the basal parts of cabbage leaves.
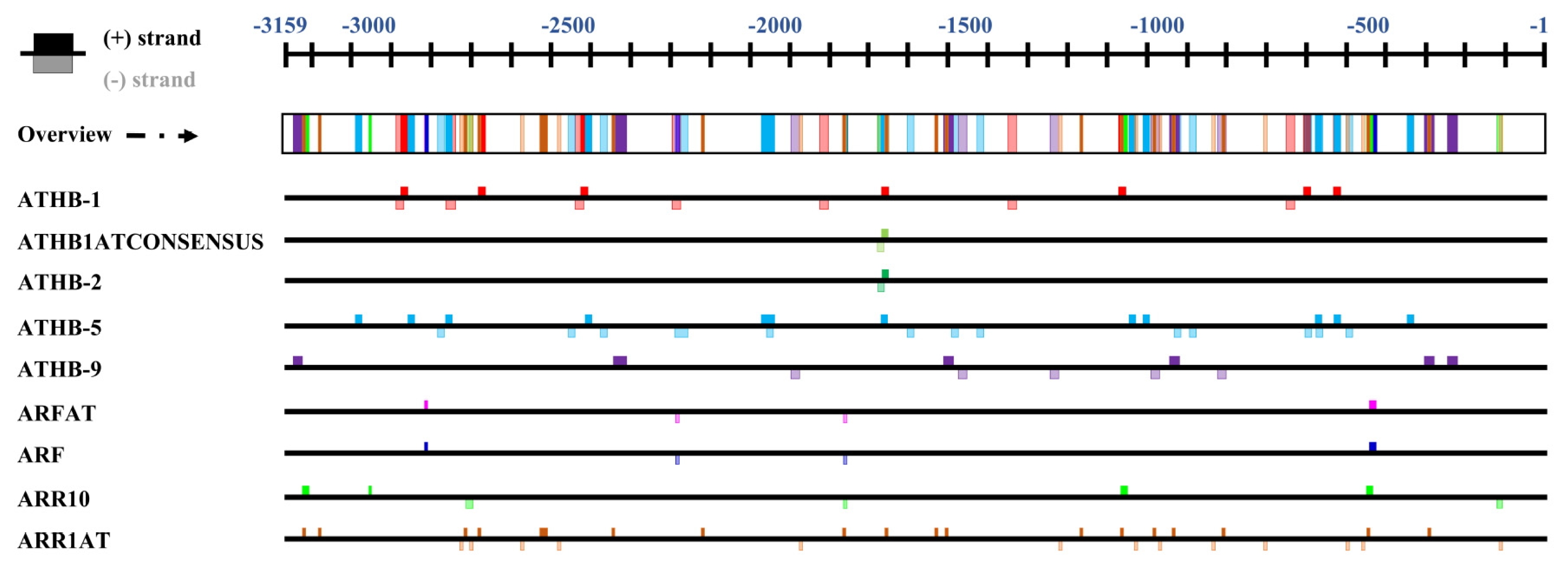
To understand why
LSH genes were specifically expressed in the midrib, we analyzed
cis-acting elements in the
B. rapa LSH2 gene (
BrLSH2) promoter, which shows the most prominent expression in the midrib compared to the leaf blade. The region 3,159 nucleotides upstream of
BrLSH2 contains 527 motifs corresponding to 67 transcription factors (data not shown). Among these, we identified the binding sites of eight transcription factors (auxin responsive:
ARF and
ARFAT, cytokinin response:
ARR10 and
ARR1AT, leaf development:
ATHB1,
ATHB1ATconsensus,
ATHB2 and
ATHB9), whose positions are shown in
Figure 8. These elements appear to be associated with leaf development or differentiation. Except for
ATGB-2 and
ATHB1ATconsensus, most elements are present at multiple sites. All
cis-elements are present within 1,700 nt of the ATG start codon, indicating that this region may confer midrib-specific expression of the
BrLSH2 gene.
DISCUSSION
Functions of DUF640 domain proteins
Although the precise functions of DUF640 domain (or ALOG) proteins have not yet been defined, there are four possible roles for these proteins based on published information, as follows: regulation of inflorescence architecture, determination of organ identity and differentiation, transport of RNA, and sensing of invading DNA.
ALOG genes control inflorescence architecture in rice and tomato (
Yoshida et al. 2009,
2013;
Li et al. 2012;
MacAlister et al. 2012;
Yan et al. 2013;
Teo et al. 2014).
Arabidopsis genes are expressed at the boundary region of the SAM and lateral organs, indicating their function in organ identity (
Cho et al. 2011;
Takeda et al. 2011). Moreover, sequence analysis of ALOG domains suggests that they help establish organ identity and differentiation by binding to specific DNA sequences and acting as transcription factors or recruiters of repressive chromatin (
Iyer and Aravind 2012). The current expression data also support the notion that all
LSH genes play a role in leaf formation and in determining the boundary between the leaf and the stem.
Solanum tuberosum LSH10 (
StLSH10) encodes an RNA-binding protein (B5RBP3) that binds to the 3′UTR of
StBEL5, a mobile RNA that controls tuber formation (
Cho et al. 2012;
Lin et al. 2013).
StBEL5 RNA is highly abundant in petioles, where it moves long distances through the phloem (
Cho et al. 2012). RBPs are commonly detected in companion cells and sieve elements of leaf veins where they serve as chaperones of mobile RNAs. The RNA levels of
B5RBP3 (
StLSH10) are remarkably low in leaves but extremely high in petioles, stolons and young tubers, suggesting that this protein plays a role in RNA transport as well as tuber development (
Cho et al. 2012). The expression patterns of
LSH genes detected in the current study, i.e., high levels of
LSH transcripts in petioles and low levels in leaf blades, suggest that
Brassica LSHs function in long-distance RNA transport. However, the levels of
BrLSHs transcripts were high in both bundle sheath cells and mesophyll cells of the midrib (
Fig. 4), suggesting that
BrLSHs play different roles from those of potato LSH10.
The ALOG domain is present in certain plant defense proteins, and domain analysis can help predict whether proteins might function as DNA sensors to detect invading DNA (
Iyer and Aranid 2012). Based on the expression data, we cannot yet determine whether
Brassica LSHs also participate in this defense response.
Midrib, the largest vein in a leaf, runs through the middle of the leaf and sometimes becomes the petiole. The midrib helps keep the leaf in an upright position and conducts foods and water
via its vascular bundles. Several recent reviews have focused on the development of leaf morphology (
Byrne 2012;
Fambrini and Pugliesi 2013), but no study on midrib development in dicots has been reported. Several such studies have been performed in monocotyledons, such as rice and sorghum. In rice, the well-studied
DROOPING LEAF (
DL) gene regulates midrib development (
Yamaguchi et al. 2004;
Ohmori et al. 2011). Moreover, the morphologies of the leaves of several
Arabidopsis mutants are similar to that of mature Chinese cabbage leaves, including plants harboring mutations in
LEAFY PETIOLE (
LEP) (
van der Graaff et al. 2000),
JAGGED (
JAG) (
Ohno et al. 2004),
BLADE-ON-PETION1 (
BOP1) and
BOP2 (
Ha et al. 2003,
2007;
Hepworth et al. 2005). In the current study, the expression pattern of
BOP1 resembled that of
LSH (data not shown), suggesting a possible interaction between these genes. Perhaps, LSHs interact with known gene products that regulate leaf shape and development.
Recent work has revealed that
CUC1 activates the expression of
LSH3 and
LSH4, which are specifically expressed in the boundary cells of various shoot organs, such as cotyledons, leaves and floral organs (
Takeda et al. 2011), indicating that
CUC1 is an upstream transcription factor of
LSH3 and
LSH4. Therefore, further studies should focus on identifying other upstream genes, as well as downstream genes, of
LSHs.
As shown in
Figure 8, major transcription factor-binding sites are related to the auxin response, the cytokinin response and leaf development. The plant hormone auxin affects plant growth and differentiation. The meristem-active regulatory genes
WUSCHEL (
WUS) and
SHOOTERMRISTEMLESS (
S™), which contain cytokinin-response regulator binding motifs (ARR1AT) and auxin-responsive elements, are highly expressed in leaf veins (
Bao et al. 2009), indicating that these elements may be related to midrib specificity or midrib differentiation. ATHB1 and ATHB2 are involved in the regulation of leaf development (
Aoyama et al. 1995) and the adaxial identity of subsequently formed organs (
Turchi et al. 2013), respectively. These observations suggest that the region 1,700 nt upstream of the ATG start codon of
BrLSH2 may contain a
cis-element required for leaf boundary formation or identity that can be used as a midrib-specific promoter.
Bacterial soft rot is the most severe and destructive disease of various crops including
Brassica family members. Chinese cabbage is highly susceptible to soft rot disease caused by the Gram-negative bacterium
Pectobacterium carotovorum subsp.
carotovorum (
Pcc) (
Ren et al. 2001;
Zhang et al. 2007). Infection tests have largely been carried out by midrib inoculation of
Brassica (
Vanjildorj et al. 2009;
Park et al. 2012), because infection starts at the stem or petiole. The analysis of the
BrLSH2 promoter performed in the current study suggests that this promoter can be used for the generation of transgenic plants exhibiting soft rot resistance. We are currently analyzing the
BrLSH2 promoter and the up- and down-stream of genes of
LSHs.
ACKNOWLEDGMENT
This work was supported by a grant from the Next- Generation BioGreen 21 Program (the Next-Generation Genomics Center No. PJ008118), RDA and by a grant National Research Foundation of Korea (NRF; 2012R1A-1A4A01012074).
Fig. 1Leaf morphology of Chinese cabbage (A) and levels of
LIGHT-SENSITIVE HYPOCOTYLS (
LSH)
2 transcripts measured throughout the day (B). Transcript levels were obtained from
B. rapa inbred line ‘Chiifu’ using a Br300K microarray (
Dong et al. 2013). Probe intensity (hybridization) values are shown at the top of the graph.
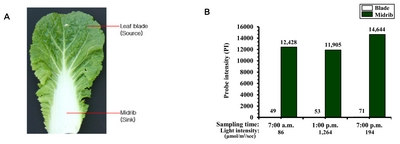
Fig. 2Phylogenetic relationships of LIGHT-SENSITIVE HYPOCOTYLS (LSH) gene families from several species. Trees were constructed using the NJ (neighbor-joining) method in MEGA (version 6.0). Red squares indicate genes from Arabidopsis. Br and Bo stand for B. rapa and B. oleracea, respectively. Numbers 1 to 10 indicate that the LSH gene families can be classified into ten groups.
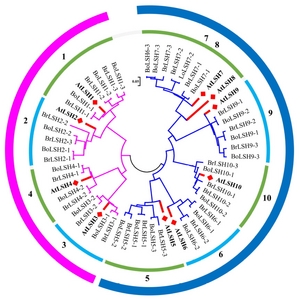
Fig. 3Expression levels of LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes in midrib (M) and leaf blade (L) tissues of various Brassica species. Abbreviations are as follows: Chiifu (Brassica rapa ssp. pekinensis, DH line Chiifu), Kenshin (Brassica rapa ssp. pekinensis, DH line Kenshin), Pakchoi (Brassica rapa ssp. chinensis), Brapha (Brassicoraphanus), Kc (Korean cabbage), Bo-10 (B. oleracea 10 days old), M (midrib) and L (leaf blade). Gene names without Br indicate transcripts amplified by common primer sets.
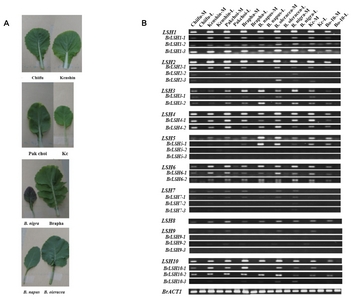
Fig. 4Expression profiles of LIGHT-SENSITIVE HYPOCOTYLS (LHS) genes in various tissues of Brassica rapa cultivars ‘Heissen’ (A, B, E) and Green cabbage (C, D). Blade (L) and midrib (M) tissues were sampled from outer (O), middle (M) and inner (I) leaves. E shows longitudinal (ls) and cross (cs) sections of ‘Heissen’ outer leaf midrib.
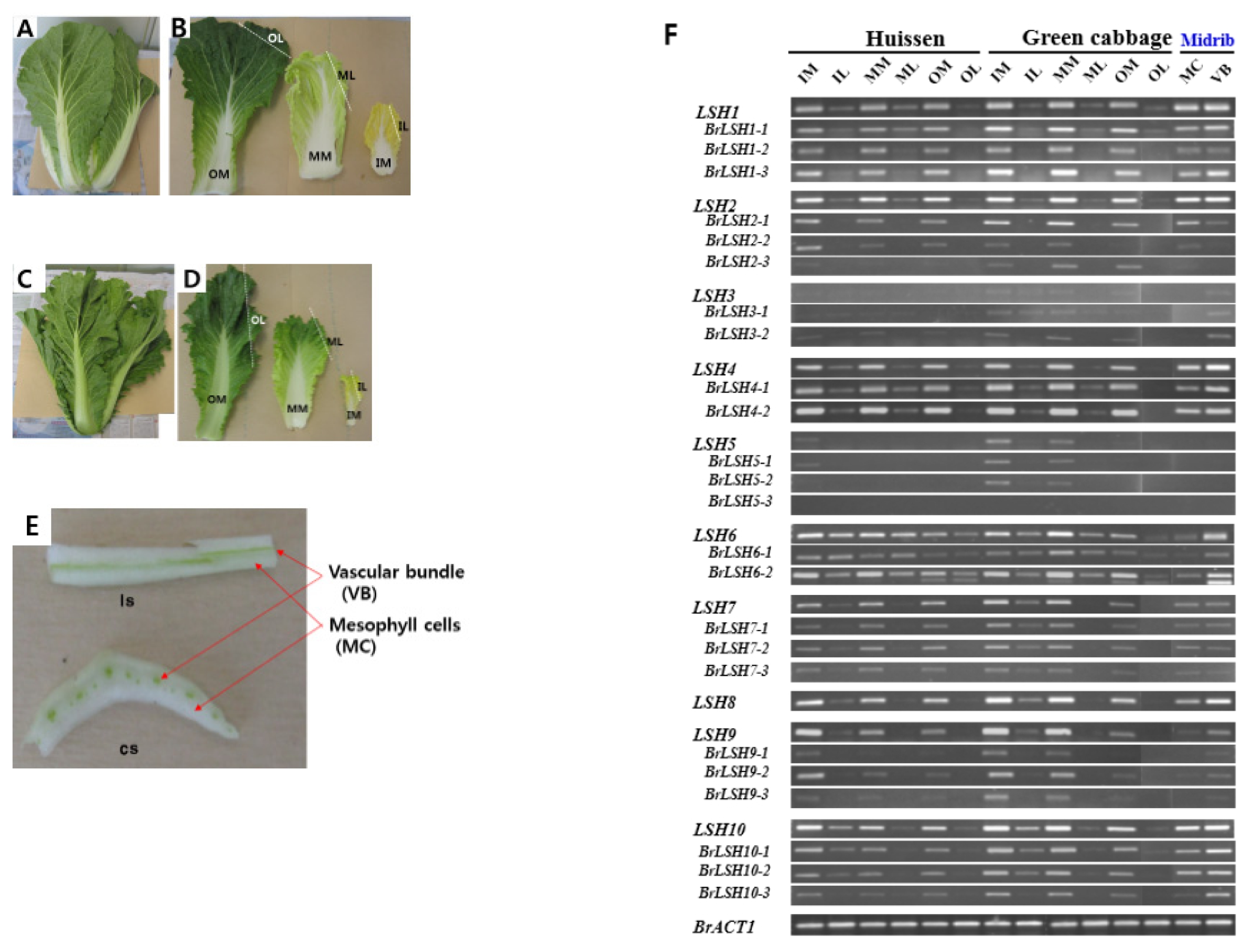
Fig. 5Expression of LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes in various parts of Korean cabbage leaves. Korean cabbage (Brassica lee ssp. namai), the progeny of a cross between Brassica rapa and B. oleracea, exhibits extremely long petioles (or extended midribs).

Fig. 6Expression of LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes in various B. oleracea tissues, beginning at the heading stage. Midrib (M) and leaf blade (L) tissues were sampled from 20-day-old B. oleracea plants (Bo-M, L, no photograph). Bo1, Bo2, Bo3 and Bo4 indicate 3rd, 6th, 10th and 13th leaves of 80-day-old plants (top middle), respectively. The sampled parts are indicated in the photographs.

Fig. 7Expression of LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes in different parts of a cabbage leaf. Samples were selected from 14 parts of a leaf, as indicated in the photograph.

Fig. 8Organization of
cis-elements in the promoter region of
BrLSH2 (Bra040188).
Cis-elements were identified using PlantPAN (
Chang et al. 2008) and PLACE (
Higo et al. 1999).
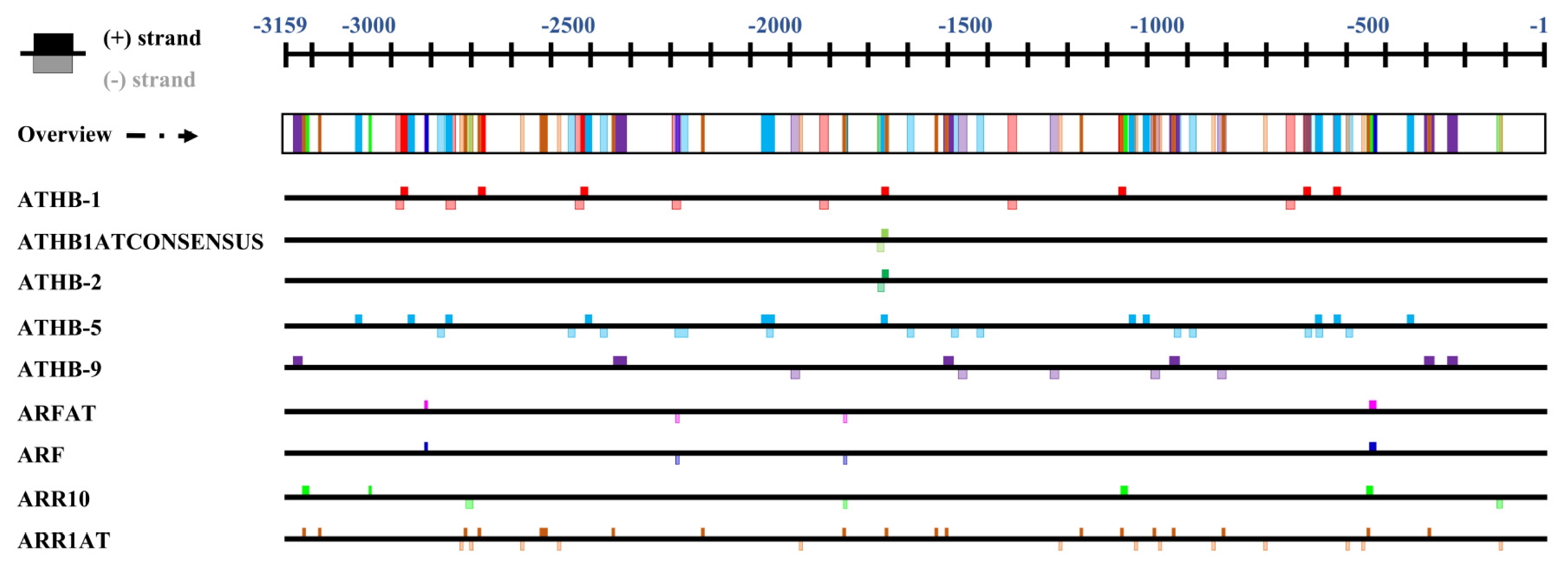
Table 1List of DUF640 superfamily (
LSH:
LIGHT-SENSITIVE HYPOCOTYLS) genes from
Arabidopsis, B. rapa and
B. oleracea. Numbers 1, 2 and 3 after the gene name indicate the highest to lowest identity to
Arabidopsis sequences.
Br_Locus and
Bo_Locus (Bol) were obtained by BlastN at
http://brassicadb.org/brad/ against the
Arabidopsis sequence. RNA_Seq Id of
B. oleracea are unpublished data.
Table 1
|
Arabidopsis thaliana
|
Brassica rapa
|
|
Brassica oleracea
|
|
|
Gene name |
At_Locus |
Gene name |
Br_Locus |
Gene name |
Bo_Locus |
RNA_ Seq Id |
|
AtLSH1
|
AT5G28490 |
BrLSH1-1
|
Bra020610 |
BoLSH1-1
|
- |
Locus_22975 |
|
BrLSH1-2
|
Bra009997 |
BoLSH1-2
|
- |
- |
|
BrLSH1-3
|
Bra036134 |
BoLSH1-3
|
Bol032364 |
Locus_36030 |
|
|
AtLSH2
|
AT3G04510 |
BrLSH2-1
|
Bra040188 |
BoLSH2-1
|
- |
Locus_48663 |
|
BrLSH2-2
|
Bra040137 |
BoLSH2-2
|
- |
Locus_27811/Locus_52336 |
|
BrLSH2-3
|
Bra001109 |
- |
- |
- |
|
|
AtLSH3
|
AT2G31160 |
BrLSH3-1
|
Bra018263 |
BoLSH3-1
|
Bol008257 |
Locus_49179 |
|
BrLSH3-2
|
Bra021707 |
BoLSH3-2
|
- |
Locus_10104 |
|
|
AtLSH4
|
AT3G23290 |
BrLSH4-1
|
Bra014935 |
BoLSH4-1
|
Bol037267 |
Locus_9116 |
|
BrLSH4-2
|
Bra023738 |
BoLSH4-2
|
Bol028747 |
Locus_19519 |
|
|
AtLSH5
|
AT5G58500 |
BrLSH5-1
|
Bra020365 |
BoLSH5-1
|
Bol015532 |
Locus_16670 |
|
BrLSH5-2
|
Bra002642 |
BoLSH5-2
|
Bol016895 |
- |
|
BrLSH5-3
|
Bra002644 |
BoLSH5-3
|
Bol016897 |
- |
|
|
AtLSH6
|
AT1G07090 |
BrLSH6-1
|
Bra015551 |
BoLSH6-1
|
Bol041101 |
Locus_13192 |
|
BrLSH6-2
|
Bra031552 |
BoLSH6-2
|
- |
Locus_21750 |
|
|
AtLSH7
|
AT1G78815 |
BrLSH7-1
|
Bra035066 |
BoLSH7-1
|
Bol038483 |
Locus_51379 |
|
BrLSH7-2
|
Bra008382 |
BoLSH7-2
|
Bol021549 |
Locus_15825 |
|
BrLSH7-3
|
Bra003637 |
BoLSH7-3
|
- |
- |
|
|
AtLSH8
|
AT1G16910 |
- |
- |
- |
- |
- |
|
|
AtLSH9
|
AT4G18610 |
BrLSH9-1
|
Bra021000 |
BoLSH8-1
|
Bol009398 |
Locus_39945 |
|
BrLSH9-2
|
Bra013330 |
BoLSH8-2
|
Bol029905 |
Locus_9841 |
|
BrLSH9-3
|
Bra012584 |
BoLSH8-3
|
Bol024421 |
Locus_14109 |
|
|
AtLSH10
|
AT2G42610 |
BrLSH10-1
|
Bra016865 |
BoLSH10-1
|
- |
Locus_52096/Locus_40164/Locus_55288 |
|
BrLSH10-2
|
Bra000269 |
BoLSH10-2
|
- |
Locus_25548/Locus_27130 |
|
BrLSH10-3
|
Bra004711 |
- |
- |
- |
Table 2Primer sequences used in RT-PCR experiments. Three to four reverse primer sequences for LIGHT-SENSITIVE HYPOCOTYLS (LSH) genes (except LSH8) were used to distinguish alleles, but one common forward primer was used. Among reverse primers, LSH-R primers were used to amplify all LSH genes from any Brassica species. LSH-R primers were designed with identical sequences among Arabidopsis, B. rapa and B. oleracea. However, BrLSH primers were specific for each allele of each LSH gene (to distinguish each allele). LSH8 primers were designed based on Arabidopsis and Eutrema salsugineum genes.
Table 2
|
Gene name |
Forward primer |
Reverse primer |
|
|
Name |
Sequence (5′→3′) |
Name |
Sequence (5′→3′) |
|
LSH1
|
LSH1-F |
CCGCCTACGAGGAGAACGGT |
LSH1-R |
TTGCACCCGAGTAATTAGCCATC |
|
BrLSH1-1 |
CGGCTGCGTCTGCGATAG |
|
BrLSH1-2 |
GCTGCTGTTCTTGTTGTTGCTGC |
|
BrLSH1-3 |
GTGGCTGCGTCTGCGGTAA |
|
|
LSH2
|
LSH2-F |
GAYTGGAACACTTTCTGCCAATACCT |
LSH2-R |
TCCACCGTTCTCCTCGTAGGC |
|
BrLSH2-1 |
GGCCTTGCTGTTGCTGCTCTA |
|
BrLSH2-2 |
CTTCTCTCTCATGCTGATGCTGTTGT |
|
BrLSH2-3 |
TGTAGAGGCGGCTGCAAACAC |
|
|
LSH3
|
LSH3-F |
TCGGTAAGACAAAAGTCCACACGA |
LSH3-R |
CTAGCTACGGCGGAGGAAGAAGA |
|
BrLSH3-1 |
TCACTTCTCAAGGTTTGATATTTGGG |
|
BrLSH3-2 |
GAGGCCGACAACTGAGGAACC |
|
|
LSH4
|
LSH4-F |
RTTYCTCAGGTACCTCGACCAATT |
LSH4-R |
CACGYGCTTTAGCCTGCGA |
|
BrLSH4-1 |
TTAGGGCTACTGGAAATCACCGAG |
|
BrLSH4-2 |
CTATTCGAAATCCCCGGCTGA |
|
|
LSH5
|
LSH5-F |
AGTACCTCGACCAGTTTGGTAAGACC |
LSH5-R |
AAGGGTTTGACTCNGGAAGACCAC |
|
BrLSH5-1 |
TCCATCATCGACGTTGGGAGTT |
|
BrLSH5-2 |
CTCCATCATCGGCGTTTAGAGTC |
|
BrLSH5-3 |
ATCCATCATGGGGATTTTGAGCT |
|
|
LSH6
|
LSH6-F |
CGTTCTTGCAGTACCTCAAGAACC |
LSH6-R |
GACTYTCTCTGACTTCYCTCAAGTAAATCC |
|
BrLSH6-1 |
TCACGGTGGTAACAGTTGTCGGT |
|
BrLSH6-2 |
CGGTGGTGACAGTTGGTGCC |
|
|
LSH7
|
LSH7-F |
ATGGCTAGTCMTAGCAACAAAGGC |
LSH7-R |
CTTSGCCTGTGAATCTCTCACTTC |
|
BrLSH7-1 |
CAAGTTGGAGCTACTAGTCCCTGTAGTAC |
|
BrLSH7-2 |
GTCCCTGCAGTACCATCTTCAACAT |
|
BrLSH7-3 |
GCTACTAGTCCCCGTAGTATCATCTTCAA |
|
|
LSH8
|
LSH8-F |
GCCGMGACTGGAACACGTTT |
LSH8-R |
AGCCGTCCGATCAAAGCATC |
|
|
LSH9
|
LSH9-F |
CAAGCTTGTGTCTTCTTCGGACAA |
LSH9-R |
CACGAGCCTTGGCTTGAGATT |
|
BrLSH9-1 |
CCTGTTCTTTTTCCTGTACGGAATC |
|
BrLSH9-2 |
TCTCCTCTTCTTCTTCCTGTACGAAATT |
|
BrLSH9-3 |
CCTCCTCTTCTTCTTCCTGTAAGGTATC |
|
|
LSH10
|
LSH10-F |
ATCATCAGGATCAGAGCCACCGGT |
LSH10-R |
GCCTGACACTCCCTAACCTCCC |
|
BrLSH10-1 |
CCACCACTCATCTCCGTCTTTG |
|
BrLSH10-2 |
CAACCATCTCCGTAGTTGGCTG |
|
BrLSH10-3 |
ACCCATCTCCGTCGTCGTTG |
References
- Aoyama CH, Dong CH, Wu Y, Carabelli M, Sessa G, Ruberti I, Morelli G, Chua NH. 1995. Ectopic expression of the Arabidopsis transcriptional activator Athb-1 alters leaf cell fate in tobacco. Plant Cell. 7: 1773-1785.
- Bao Y, Dharmawardhana P, Arias R, Allen MB, Ma C, Strauss SH. 2009. WUS and S™-based reporter genes for studying meristem development in poplar. Plant Cell Rep. 28: 947-962.
- Byrne ME. 2012. Make leaves. Curr Opin Plant Biol. 15: 24-30.
- Cary AJ, Che P, Howell SH. 2002. Developmental events and shoot apical meristem gene expression patterns during shoot development in Arabidopsis thaliana. Plant J. 32: 867-877.
- Chang WC, Lee TY, Huang HD, Huang HY, Pan RL. 2008. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene group. BMC Genomics. 9: 561
- Cho E, Zambryski PC. 2011. Organ boundary1 defines a gene expressed at the junction between the shoot apical meristem and lateral organs. Proc Natl Acad Sci USA. 108: 2154-2159.
- Cho SK, Kang IH, Carr T, Hannapel DJ. 2012. Using the yeast three-hybrid system to identify proteins that interact with a phloem-mobile mRNA. Front Plant Sci. 3: 189
- Dong X, Feng H, Xu M, Lee J, Kim YK, Lim YP, Piao ZY, Park YD, Ma H, Hur Y. 2013. Comprehensive analysis of genic male sterility-related genes in Brassica rapa using a newly developed Br300K oligomeric chip. PLoS One. 8: e72178
- Fambrini M, Fugliesi C. 2013. Usual and unusual development of the dicot leaf: involvement of transcription factors and hormones. Plant Cell Rep. 32: 899-922.
- Ha CM, Kim GT, Kim BC, Jun JH, Soh MS, Ueno Y, Machida Y, Tsukaya T, Nam HG. 2003. The BLADE- ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development. 130: 161-172.
- Ha CM, Jun JH, Nam HG, Fletcher JC. 2007. BLADE- ON-PETIOL1 and 2 control Arabidopsis lateral organ fate through regulation of LOB domain and adaxial- abaxial polarity genes. Plant Cell. 19: 1809-1825.
- Hepworth SR, Zhang Y, McKim S, Li X, Haughn GW. 2005. BLADE-ON-PETIOLE-dependent signaling controls leaf and floral patterning in Arabidopsis. Plant Cell. 17: 1434-1448.
- Higo K, Ugawa Y, Iwamoto M, Korenaga T. 1999. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 27: 297-300.
- Iyer LM, Arvind L. 2012. ALOG domains: provenance of plant homeotic and developmental regulators from the DNA-binding domain of a novel class of DIRS1-type retroposons. Biol Direct. 7: 39
- Larkin MA, Blackshields G, Brwon NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. 2007. Clustal W and Clustal X version 2.0. Bioinformatics. 23: 2947-2948.
- Lee SS, Lee SA, Yang J, Kim J. 2011. Developing stable progenies of xBrassicoraphanus, an intergeneric allopolyploid between Brassica rapa and Raphanus sativus, through induced mutation using microspore culture. Theor Appl Genet. 122: 885-891.
- Li X, Sun L, Tan L, Liu F, Zhu Z, Fu Y, Sun X, Sun X, Xie D, Sun C. 2012. TH1, a DUF640 domain-like gene controls lemma and palea development in rice. Plant Mol Biol. 78: 351-359.
- Lin T, Sharma P, Gonzalez DH, Viola IL, Hannapel DJ. 2013. The impact of the long-distance transport of a BEL1-like messenger RNA on development. Plant Physiol. 161: 760-772.
- MacAlister CA, Park SJ, Jinag K, Marcel F, Bendahmane A, Izkovich Y, Eshed Y, Lippman ZB. 2012. Synchonization of the flowering transition by the tomato TERMINATING FLOWER gene. Nat Genett. 44: 1393-1398.
- McConnell JR, Emery J, Eshed Y, Bao N, Bowman J, Barton MK. 2001. Role of PHABULOSA and PHAVOLUTA in determining radial patterning in shoots. Nature. 411: 709-713.
- Ohmori Y, toriba T, Nakamura H, Ichikawa H, Hirano HY. 2011. Temporal and spatial regulation of DROOPING LEAF gene expression that promotes midrib formation in rice. Plant J. 65: 77-86.
- Ohno CK, Reddy GV, Heisler MG, Meyerowitz EM. 2004. The Arabidopsis JAGGED gene encodes a zinc finger protein that promotes leaf tissue development. Development. 131: 1111-1122.
- Park YH, Choi C, Park EM, Kim HS, Park HJ, Bae SC, Ahn I, Kim MG, Park SR, Hwang DJ. 2012. Over- expression of rice leucine-rich repeat protein results in activation of defense response, thereby enhancing resistance to bacterial soft rot in Chinese cabbage. Plant Cell Rep. 31: 1845-1850.
- Ren JP, Petzoldt R, Dickson MH. 2001. Screening and identification of resistance to bacterial soft rot in Brassica rapa. Euphytica. 118: 271-280.
- Risseeuw E, Venglat P, Xiang D, Komendant K, Daskalchuk T, Babic V, Crosby W, Datla R. 2013. An activated form of UFO alters leaf development and produces ectopic floral and inflorescence meristem. PLoS One. 8: e83807
- Samach A, Klenz JE, Kohalmi Se, Risseeuw E, Haughn GW, Crosby WL. 1999. The UNUSUAL FLORAL GRGNAS gene of Arabidopsis thaliana is an F-box protein required for normal patterning and growth in the floral meristem. Plant J. 20: 433-445.
- Takeda S, Hanano K, Kariya A, Shimizu S, Zhao L, Matsui M, Tasaka M, Aida M. 2011. CUP-SHAPED COTYLEDON1 transcription factor activates the expression of LSH4 and LSH3, two members of the ALOG gene family, in shoot organ boundary cells. Plant J. 66: 1066-1077.
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 30: 2725-2729.
- Teo ZW, Song S, Wang YQ, Liu J, Yu H. 2014. New insights into the regulation of inflorescence architecture. Trends Plant Sci. 19: 158-165.
- Turchi L, Carabelli M, Ruzza V, Possenti M, Sassi M, Peñalosa A, Sessa G, Salvi S, Forte V, Morelli G, Ruberti I. 2013. Arabidopsis HD-Zip II transcription factors control apical embryo development and meristem function. Development. 140: 2118-2129.
- van der Graff E, Dulk-Ras AD, Hooykaas PJ, Keller B. 2000. Activation tagging of the LEAF PETIOLE gene affects leaf petiole development in Arabidopsis thaliana. Development. 127: 4971-4980.
- Vanjildorj E, Song SY, Yang ZH, Choi JE, Noh YS, Park S, Lim WJ, Cho KM, Yun HD, Lim YP. 2009. Enhancement of tolerance to soft rot disease in the transgenic Chinese cabbage (Brassica rapa L. ssp. pekinensis) inbred line Kenshin. Plant Cell Rep. 28: 1581-1591.
- Yamaguchi T, Nagasawa N, Kawasaki S, Matsumoka M, Nagato Y, Hirano HY. 2004. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell. 16: 500-509.
- Yan DW, Zhou Y, Ye SH, Zeng LJ, Zhang XM, He ZH. 2013. BEAK-SHAPED GRAIN 1/TRIANGULAR HULL 1, a DUF640 gene, is associated with grain shape, size and weight in rice. Sci China Life Sci. 56: 275-283.
- Yoshida A, Suzaki T, Tanaka W, Hirano HY. 2009. The homeotic gene long sterile lemma (G1) specifies sterile lemma identity in the rice spikelet. Proc Natl Acad Sci USA. 106: 20103-20108.
- Yoshida A, Sasao M, Yasuno N, Takagi K, Daimon Y, Chen R, Yamazaki R, Tokunaga H, Kitaguchi Y, Sato Y, Nagamura Y, Ushijima T, Kumamaru T, Iida S, Maekawa M, Kyozuka J. 2013. TAWAWA1, a regulator of rice inflorescence architecture, functions through the suppression of meristem phase transition. Proc Natl Acad Sci USA. 110: 767-772.
- Zhang SH, Yang Q, Ma RC. 2007. Erwinia carotovora ssp. carotovora infection induced “defense lignin” accumulation and lignin biosynthetic gene expression in Chinese cabbage (Brassica rapa L. ssp. pekinensis). J Integr Plant Biol. 49: 993-1002.
- Zhao L, Nakazawa M, Takase T, Manabe K, Kobayash M, Seki M, Shinozaki K, Matsui M. 2004. Overexpression of LSH1, a member of an uncharacterised gene family, causes enhanced light regulation of seedling development. Plant J. 37: 694-706.