Abstract
Milk thistle (Silybum marianum (L.) Gaertn.), a valuable medicinal plant with economic importance, serves as a source of both therapeutic compounds and vegetable oil for human use. Genome editing techniques have been widely applied to improve key agronomic traits in various species. Therefore, this study aims to develop an efficient method for seed sterilization and protoplast isolation in milk thistle. For seed sterilization, seed treated with hydrogen peroxide for either 4 hours or 7 hours with ethanol and detergent showed more sufficient to sterilize seed of milk thistle for in vitro growth than ethanol, sodium hypochlorite, and chlorine gas. For protoplast isolation in milk thistle, high-yielding, viable protoplasts were successfully isolated from the leaves of 21-day-old plants using a 2 hours enzymatic treatment containing 2% Viscozyme® L, 1% Celluclast® 1.5 L, and 1% Pectinex® Ultra SP-L. Additionally, transient expression of green fluorescent protein was observed following polyethylene glycol-mediated transfection. Therefore, the seed sterilization and protoplast isolation methods developed in this study can facilitate in vitro culture, gene function analysis, and genome editing aimed at improving agronomic traits in milk thistle.
-
Key words: Milk thistle, Silybum marianum, Protoplast isolation, Seed sterilization, Transient expression
Introduction
Milk thistle (Silybum marianum (L.) Gaertn.), a diploid species (2n = 34) belonging to the Asteraceae family, originates from the Mediterranean region and is now widespread globally. This annual or biennial plant is characterized by large, glossy, spiny leaves and purple flower heads. Currently, milk thistle is cultivated both as a medicinal plant and as a crop, primarily for the extraction of silymarin(
Andrzejewska et al. 2015;
Hetz et al. 1995a;
Zhang et al. 2024).
Furthermore, milk thistle seeds contain a relatively high oil content (20–30%), rich in essential fatty acids, including linoleic acid and oleic acid, along with antioxidants and bioactive compounds such as carotenoids, tocopherols, and sterols (
Liu et al. 2023;
Malekzadeh et al. 2011).
Milk thistle is commercially available in the form of tablets, capsules, and intravenous solutions. The global milk thistle market is valued at USD 115.10 million in 2025 and is projected to reach approximately USD 228.96 million by 2034 (
https://www.precedenceresearch.com/milk-thistle-market; (
Tolangi et al. 2024)). Despite growing interest in its cultivation and economic value in the herbal market, genetic research on this species remains limited. Several agronomic challenges hinder the development of high-yield cultivars, such as asynchronous flowering, variability in seed yield and silymarin content among genotypes, spiny foliage, and fruit shattering at maturity (
Marceddu et al. 2022;
Martinelli 2019;
Zhang et al. 2024).
Plant protoplasts are totipotent cells capable of regenerating into entire plants or specific organs. Consequently, they serve as valuable systems for investigating gene expression, protein–protein interactions and for facilitating plant breeding through techniques such as electroporation and polyethylene glycol (PEG)-mediated transfection. Recent studies report that Clustered Regularly Interspaced Short Palindromic Repeats and CRISPR-associated protein 9 (CRISPR/Cas9) technology is a potent tool for plant genome editing and generation of transgenic plants using
Agrobacterium-mediated transformation and protoplast transfection (
Bhuyan et al. 2023;
Chen et al. 2024a;
Chen et al. 2019;
Yao et al. 2023)
However,
Agrobacterium-mediated transformation and plant protoplast isolation/culture techniques for milk thistle remain underdeveloped, with few studies reported (
Hetz et al. 1995b;
Si-qing et al. 1990). Therefore, this study aims to investigate the protoplast isolation and transfection methods for effective genome editing in milk thistle.
Materials and Methods
Plant materials
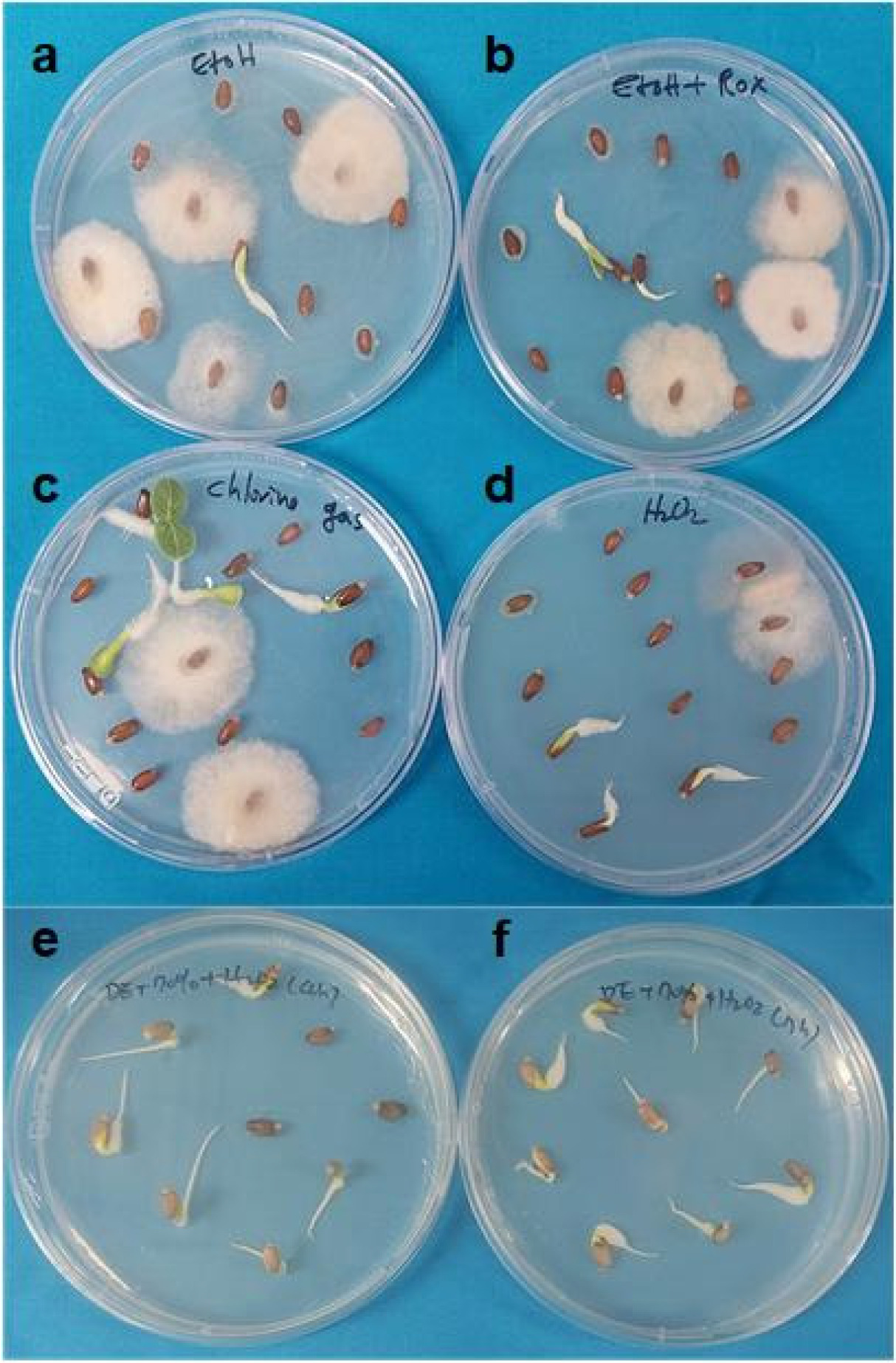
Milk thistle seeds were surface sterilized using three different treatments: commercial bleach (5.25% NaOCl), chlorine gas, and 3% (v/v) hydrogen peroxide (H2O2). For bleach treatment, seeds were first rinsed with 70% ethanol for 5 minutes, followed by immersion in 50% (v/v) commercial bleach for 30 minutes. For chlorine gas treatment, seeds were exposed to chlorine gas for 17 hours (h). Chlorine gas was generated by mixing 95 mL of 12% bleach with 5 mL of 37% hydrochloric acid in a desiccator placed under a fume hood. For H2O2 treatment, two approaches were used. In the first, milk thistle seeds were soaked in 3% H2O2 for 4 h. In the second, prior to 7 h H2O2 treatment, seeds were first washed with a detergent solution (two drops of Tween 20 in 100 mL water) for 30 minutes and immersed in 70% (v/v) ethanol for 5 minutes. After H₂O₂ treatment, the sterilized seeds were rinsed five times with sterile distilled water and germinated on half-strength Murashige and Skoog (1/2 MS) medium (Murashige et al. 1962) supplemented with 3% sucrose and 0.8% plant agar. Cultures were maintained under long-day conditions (16 h light/8 h dark) at 25℃ for 3 weeks. Subsequently, healthy leaves were used for protoplast isolation.
Protoplast isolation and purification protocol
Protoplast isolation and purification were performed following the protocol described by
Jeong et al. (2021), with slight modifications. Briefly, the two youngest leaves from 21-day-old plants were sliced into approximately 0.5 mm strips and incubated in 20 mL of enzyme solution containing 0.5 M mannitol, 10 mM 2-(N-morpholino)ethanesulfonic acid (MES; pH 5.7), 10 mM CaCl
2, 0.1% (w/v) bovine serum albumin, 2% Viscozyme
® L (Novozymes, Bagsværd, Denmark), 1% Celluclast
® 1.5 L, (Novozymes, Bagsværd, Denmark), and 1% Pectinex Ultra
® SP-L (Novozymes, Bagsværd, Denmark). The enzyme solutions were sterilized using a 0.2 µm syringe filter (Sartorius, Göttingen, Germany). Healthy leaves were digested using various enzyme combinations and incubation times (1-4 h) to evaluate protoplast yield and viability. Following enzymatic cell digestion of the cell walls, an equal volume of W5 solution (154 mM NaCl, 125 mM CaCl
2, 5 mM KCl, and 2 mM MES; pH 5.7) was added to terminate the reaction. Undigested tissues were removed using 40 µm cell strainers (SPL Life Sciences, Pocheon, South Korea). The filtrate containing protoplasts was centrifuged at 100 × g for 7 minutes at room temperature. The resulting protoplast pellet was resuspended in 2 mL of MMG solution (4 mM MES, pH 5.7, 0.5 M mannitol, 15 mM MgCl
2).
For purification, 2 mL of protoplast suspension was transferred into 6 mL of 0.6 M sucrose solution using a wide-bore pipette tip. The mixture was centrifuged at 500 rpm for 20 minutes. Subsequently, approximately 2 mL of viable protoplasts were transferred to 8 mL of W5 solution and centrifugation again at 500 rpm for 10 minutes. The collected protoplasts were resuspended in 2 mL of W5 solution and placed on ice for 30 minutes. Following a final centrifugation step at 500 rpm for 2 minutes, the protoplast density was adjusted to 2 × 106 protoplasts/mL in MMG solution using a hemocytometer under light microscopy (Leica Microsystems, Wetzlar, Germany) by using a hemocytometer.
Protoplast viability was evaluated using fluorescein diacetate (FDA; Sigma-Aldrich, St. Louis, MO, USA). A 2% (w/v) FDA stock solution was prepared by dissolving the FDA in 1 mL of acetone. Subsequently, 100 μL of protoplast suspension was mixed with 1 μL of the FDA stock solution and incubated in the dark at room temperature for 10 minutes. The stained protoplasts were observed under a fluorescence microscope (Olympus BX61, Olympus Corporation, Tokyo, Japan). Viability was calculated as follows: Viability = (the total number of protoplasts with green fluorescence through FDA staining/ the total number of observed protoplasts) x 100.
Green fluorescent protein reporter gene transfection assay
PEG-mediated transfection was performed following a previously described protocol (
Yoo et al. 2007) with slight modifications. Briefly, 200 μL of protoplast suspension was mixed with 20 μg of plasmid DNA carrying the GFP under the control of the pCsVMV-eGFP in a 2 mL microcentrifuge tube. Subsequently, 220 μL of 40% PEG-Ca
2+ solution containing 0.5 M mannitol, 100 mM CaCl
2, and 40% PEG 4000 (Sigma-Aldrich, St. Louis, MO, USA) was added. The mixture was incubated at room temperature for 15 minutes. The transfected protoplasts were washed with W5 solution (154 mM NaCl, 125 mM CaCl
2, 5 mM KCl, and 2 mM MES, pH 5.7) and incubated in the dark for 8 h at 25℃ in incubation buffer (0.5 M mannitol, 4 mM MES, and 20 mM KCl, pH 5.7). GFP expression was visualized and imaged using fluorescence microscopy.
Statistical Analysis was performed with Microsoft Excel Student’s t-test. All experiments were replicated at least three times, and all data are presented as means ± SD. Significance levels are marked as: * p < 0.05.
Results
Effect of different sterilization methods
The sterilization step in
in vitro culture is crucial for eliminating fungal and microbial contamination. Common surface sterilizing agents include ethanol, sodium hypochlorite (NaOCl), mercuric chloride (HgCl₂), and chlorine gas, and are widely used in various plant species, including milk thistle (
Elwekeel et al. 2012;
Hasanloo et al. 2008;
Rady et al. 2018). In this study, seeds were sterilized using four different agents.
Figs. 1a–1b. illustrates that treatment with 70% ethanol alone or 70% ethanol combined with 2% NaOCl was insufficient to achieve effective sterilization for
in vitro growth.
Chlorine gas is used to sterilize seeds of Arabidopsis, carnation, and soybean (
Clough et al. 1998;
Li et al. 2017;
Peluola et al. 2020). However, a 17 h chlorine gas treatment followed by inoculation on media was insufficient for effective sterilization to support
in vitro growth (
Fig. 1c).
Hydrogen peroxide (H
2O
2) at 3% concentration is used for sterilizing seeds of cotton,
Ribes cereum, and
Cannabis sativa (
Barampuram et al. 2014;
Rosner et al. 2003;
Sorokin et al. 2020). In this study, milk thistle seeds were treated with 3% H
2O
2 for 1 h (
Fig. 1d) and in separate experiments for 4 h or 7 h (
Figs. 1e–1f). For the 4 h and 7 h treatments, seeds were washed with a detergent solution (two drops of Tween-20 in 100 mL of water) for 30 minutes, followed by a 5 minutes wash with 70% (v/v) ethanol. Subsequently, the seeds were treated with 3% H
2O
2 for 4 h or 7 h with shaking at 200 rpm and washed five times with sterile distilled water.
Figs. 1e–1f. shows that the 4 h (
Fig. 1e) and 7 h (
Fig. 1f) H₂O₂ treatments achieved sterilization efficiencies of approximately 90% and 95%, respectively. These findings indicate that 3% H
2O
2 is effective for surface sterilization of milk thistle seeds under the conditions used in this study.
The plant cell wall is primarily composed of cellulose, hemicellulose, pectin, and lignin, with cellulose serving as the major structural component and being highly resistant to hydrolysis. Therefore, effective cellulose digestion is essential for successful protoplast isolation. Several factors influence protoplast isolation efficiency, such as enzyme type, digestion time, and incubation temperature. Among the enzymes used, Cellulase RS and Macerozyme R-10 (YAKULT Co. Ltd., Tokyo, Japan) are widely used in various plant species, including milk thistle (
Cabral et al. 1984;
Hetz et al. 1995b;
Si-qing et al. 1990). In this study, we used commercial enzyme preparations—Viscozyme
® L, Celluclast
® 1.5 L, and Pectinex Ultra
® SP-L (Novozymes, Bagsværd, Denmark)—each characterized by distinct enzymatic activities. Viscozyme
® L is a lignocellulose-specialized multi-enzyme complex containing various carbohydrases. Celluclast
® 1.5 L is a cellulase preparation enriched with β-glucosidase activity. Pectinex Ultra
® SP-L is a pectinase complex containing pectin transeliminase, polygalacturonase, and pectinesterase.
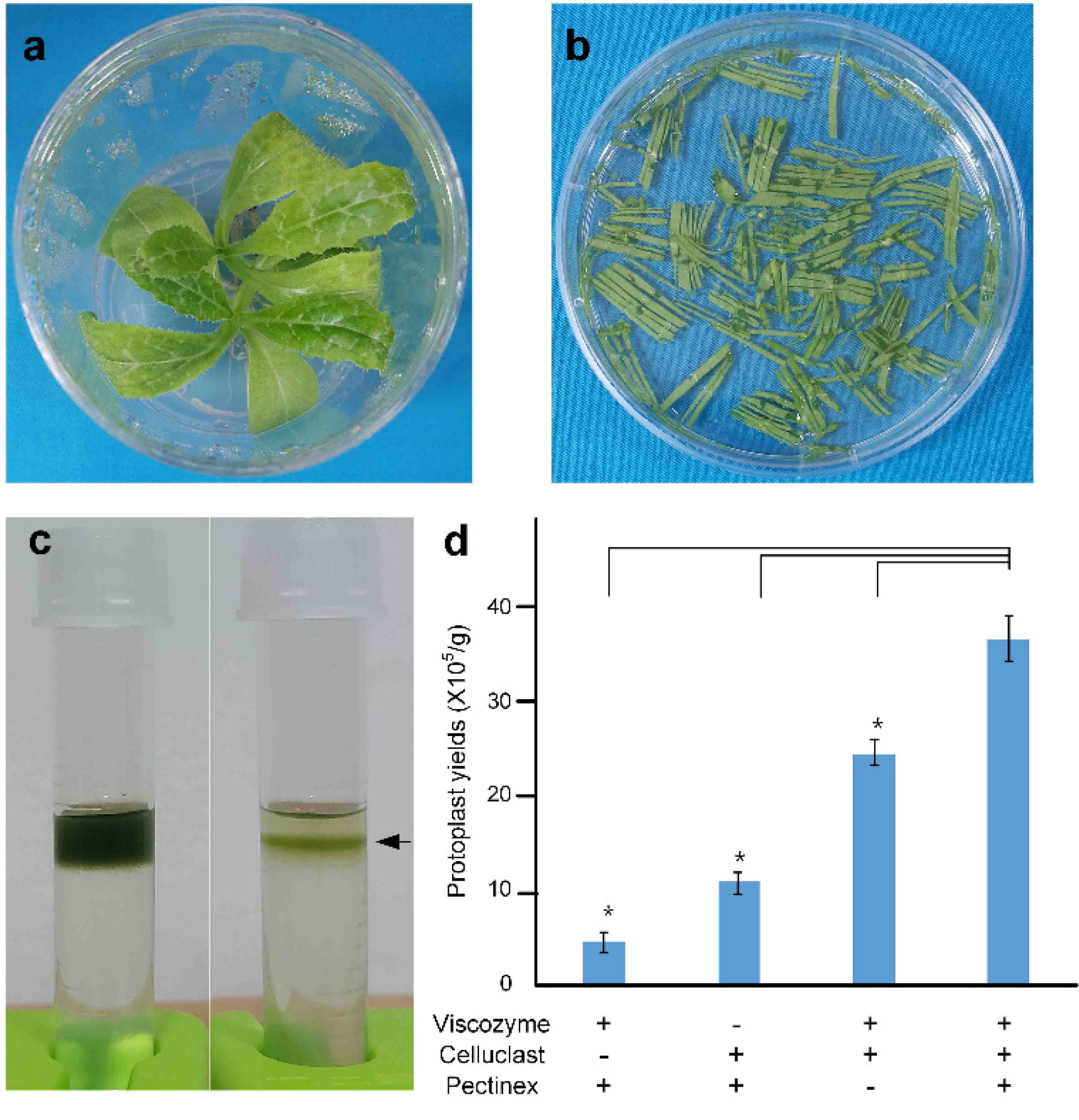
To evaluate the contribution of each enzyme in the isolation process, four enzyme mixtures were tested: one containing all components, one lacking Viscozyme
® L, one lacking Celluclast
® 1.5 L, and one lacking Pectinex Ultra
® SP-L. Protoplasts were isolated using the two youngest leaves of 21-day-old plants following a 2 h enzymatic digestion under each condition (
Figs. 2a–2b). After a 2 h digestion and sucrose gradient separation (
Fig. 2c), the enzyme combinations produced significant differences in protoplast yields. In the absence of Celluclast
® 1.5 L (–Celluclast) and Viscozyme
® L, the protoplast yield was very low, while the omission of Pectinex Ultra
® SP-L (–Pectinex) resulted in approximately 50% reduction in yield compared to the full enzyme mixture (
Fig. 2d). This result suggests that Viscozyme
® L and Celluclast
® 1.5 L are the most effective enzymes for isolating protoplasts from milk thistle leaves.
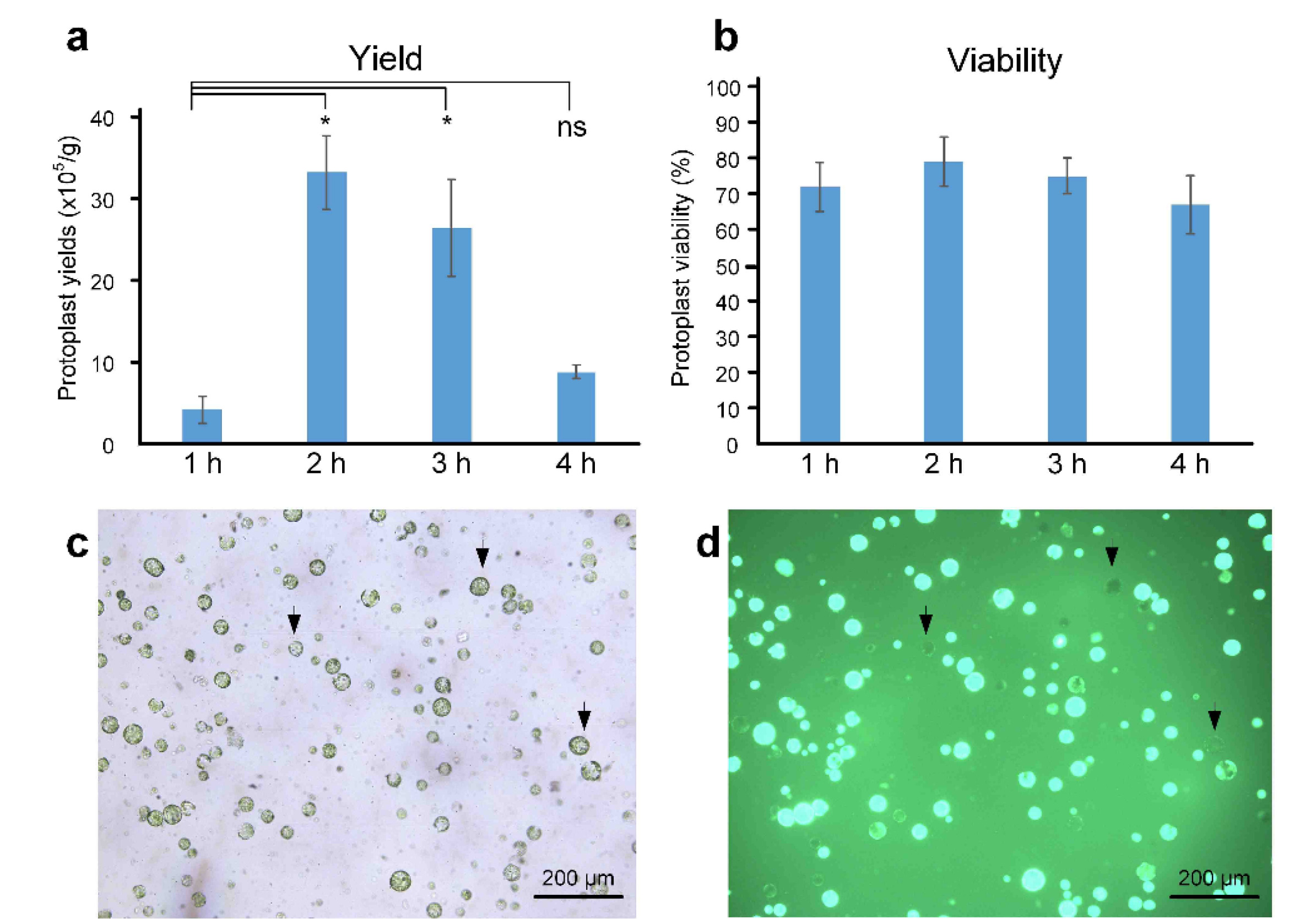
Beyond the enzyme composition in the digestion solution, enzyme digestion time is a critical factor influencing protoplast isolation from various plant materials. The optimal digestion time varies depending on the type of explants, such as leaves or calluses. For leaf tissues, complete degradation was achieved after a digestion time of 3–12 h (
Bart et al. 2006;
Jeong et al. 2021). In preliminary experiments with longer digestion times (2 h, 4 h, 6 h, and 8 h) in this study, protoplast yields were consistently low (data not shown). To determine the optimal digestion time for maximizing protoplast yield and viability, enzymatic digestion was performed for 1 h, 2 h, 3 h, and 4 h, and cell viability was assessed using an FDA staining assay. Digestion time significantly influenced both protoplast yield and viability. A 1 h treatment resulted in very low protoplast recovery (
Fig. 3). Yield increased significantly at 2 h but decreased after 3 h and 4 h of digestion. The highest yield and viability were observed at 2 h, followed by reduced values at 3 h and 4 h. These findings indicate that a 2 h digestion time is optimal for protoplast isolation.
Hetz et al. (1995b) reported that 16–18 h of immersion in an enzyme solution on a roller (3 rpm) is effective for protoplast isolation from milk thistle leaves and suspension cells. In contrast,
Si-qing et al. (1990) achieved callus-derived protoplast isolation using a 4.5–5 h digestion period. However, direct comparison of these protocols is challenging due to differences in enzyme composition, shaking speed, and plant genotype used in each study.
Direct delivery of plasmid DNA or ribonucleoproteins (RNPs) into plant cells is achieved via PEG-mediated or electroporation-based transfection. These methods facilitate the generation of transgene-free mutants and the analysis of gene expression. PEG-mediated transfection is widely used to deliver plasmids into plant protoplasts from species such as
Arabidopsis, maize, cabbage, tomato, and soybean (
Coy et al. 2022;
Kang et al. 2025;
Patil et al. 2022;
Stajič et al. 2023;
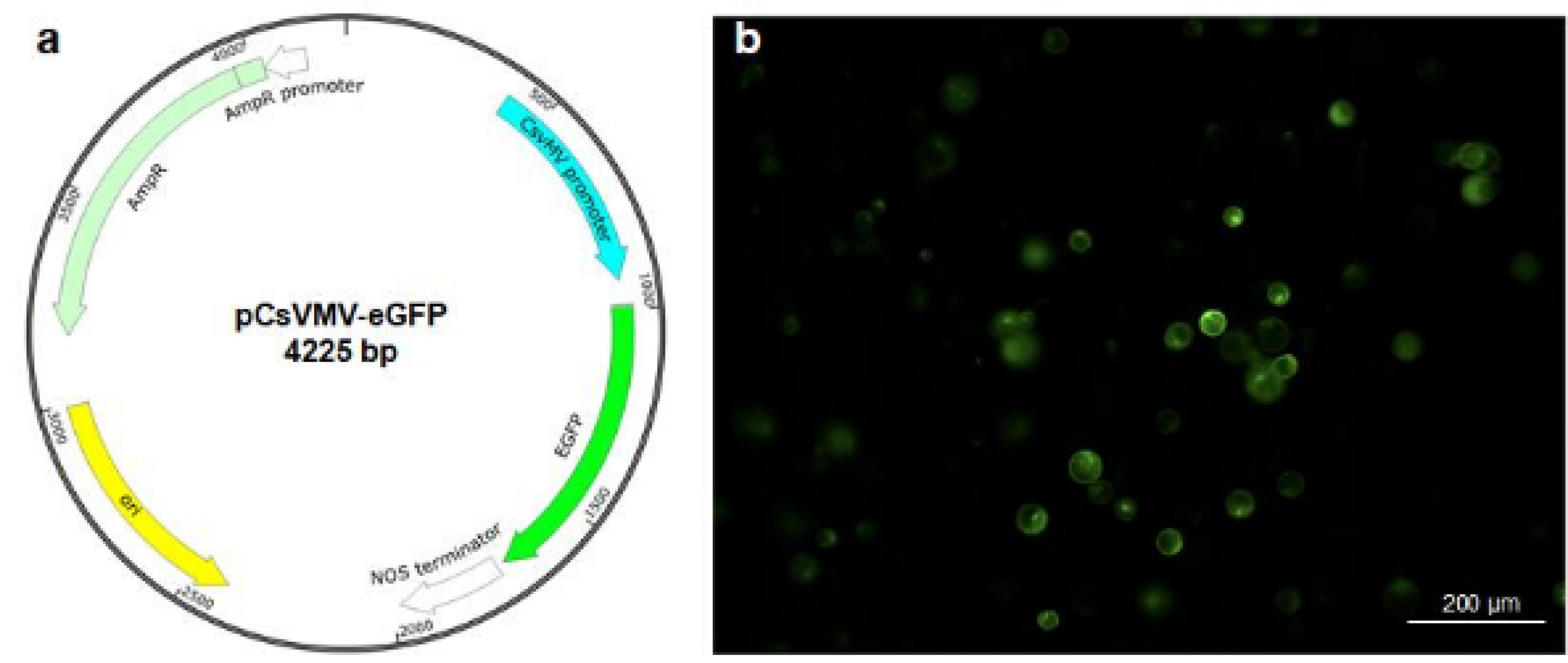
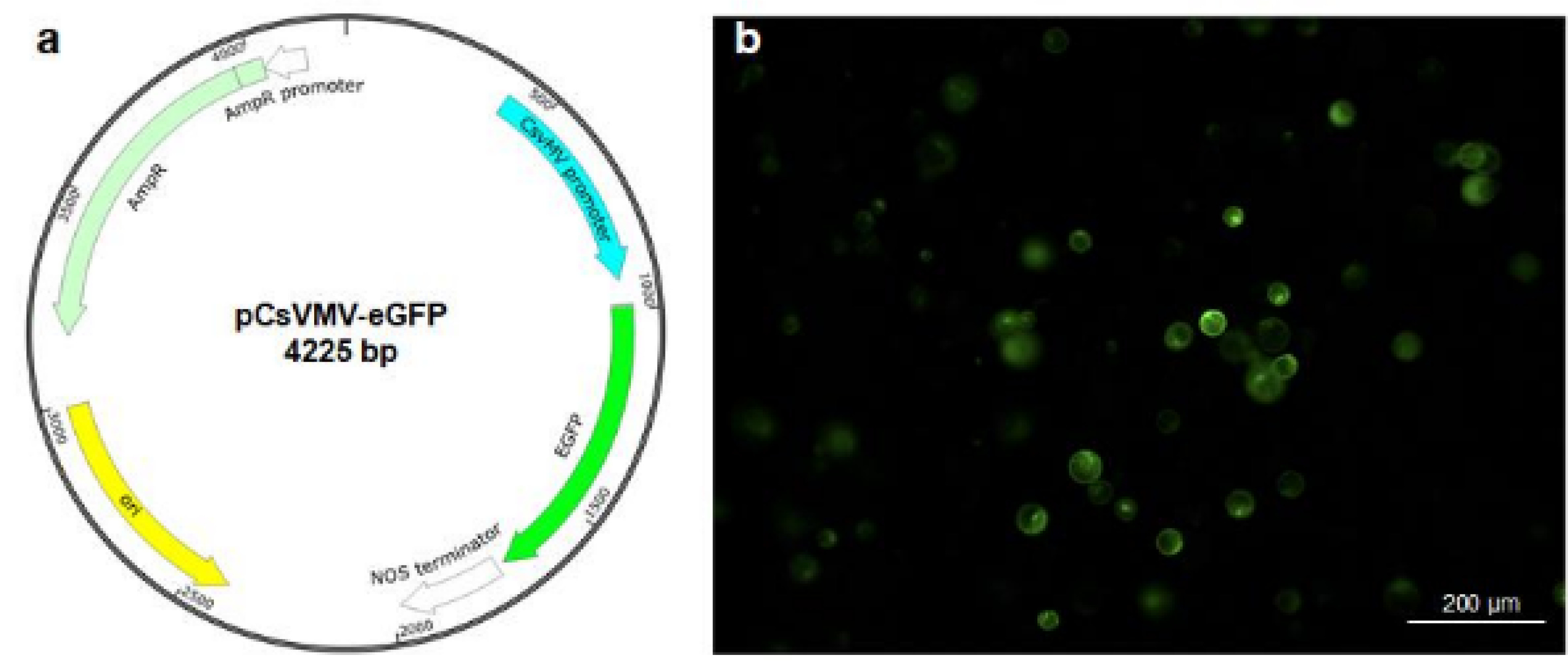
Yoo et al. 2007). To perform protoplast transfection, protoplasts were transfected with a vector carrying the enhanced green fluorescent protein (eGFP) gene under the control of the cassava vein mosaic virus promoter (pCsVMV-eGFP) (
Samac et al. 2004;
Verdaguer et al. 1998)(
Fig. 4a). Both the number of protoplasts and the concentration of plasmid DNA are critical factors influencing transfection efficiency (
Burris et al. 2016;
Kang et al. 2020). In Arabidopsis and other plant species, 10~20 µg of plasmid DNA and 2 × 10
5 protoplasts are typically used. Consequently, we employed 20 µg of plasmid DNA and 2 × 10
5 protoplasts, using 40% PEG 4000 (PEG with 4000 molecular weight) solution for the transfection. These results showed that intact protoplasts expressing GFP fluorescence were observed 8 h after transfection (
Fig. 4b), indicating that the transfection protocol commonly used in other plants, such as Arabidopsis, is effective in milk thistle.
Discussion
Sterilization is a critical step in
in vitro culture to eliminate fungal and microbial contamination, ensuring the acquisition of high-quality, sterile explant material for tissue culture. In this study, we compared with several disinfectants such as, ethanol, sodium hypochlorite, and chlorine gas for milk thistle seeds sterilization. The type and size of the seeds significantly influenced the effectiveness of the disinfectants. Both sterilization duration and the choice of disinfectant had a notable impact on the sterilization outcome. While ethanol, sodium hypochlorite, and chlorine gas showed disinfecting effects, their sterilization efficiencies were relatively low when used individually, suggesting the need for their combination with other agents in milk thistle seed sterilization (
Fig. 1). Previous studies have shown that hydrogen peroxide treatment is effective for surface sterilization and seed germination in watermelon and maize (
Duval et al. 2000;
Lizárraga-Paulín et al. 2013). In
Brassica oleracea seeds, contamination was completely eliminated with a 3% H₂O₂ treatment for 30 minutes (
Sanna et al. 2022). However, a 1 h treatment with H₂O₂ alone had no significant sterilizing effect on milk thistle seeds (
Fig. 1). In contrast, the combined treatment with detergent, ethanol, and H₂O₂ was most effective for surface sterilization, consistent with results reported in cotton (
Barampuram et al. 2014).
For protoplast preparation, different plant species—and even different organs within the same plant—require specific enzyme compositions (
Zhao et al. 2016). Previous studies reported that an enzyme mix of 0.02% macerozyme, 0.5% cellulase, and 0.05% pectolase for 16–18 h was suitable for protoplast isolation from milk thistle leaves and suspension cells. For callus-derived protoplasts, a combination of 0.8% macerozyme, 3.0% cellulase, and 0.1% pectolase was found to be more effective (
Hetz et al. 1995b;
Si-qing et al. 1990). To date, there have been no reports on the use of a three-enzyme mixture comprising Viscozyme
® L, Celluclast
® 1.5 L, and Pectinex
® Ultra SP-L for protoplast isolation in milk thistle. In our study, treatment with 2% Viscozyme
® L, 1% Celluclast
® 1.5 L, and 1% Pectinex
® Ultra SP-L for 2 h resulted in a protoplast yield of 1.0 × 10⁵ with approximately 90% viability.
For PEG-mediated transfection, 40% PEG 4000 is the most commonly used, and we found this method is effective in milk thistle. However, the transformation efficiency achieved with the 40% PEG 4000 was limited to 7–10% in
Chirita pumila protoplasts (
Wang et al. 2022). Electroporation has also been employed for delivering plasmid DNA and ribonucleoproteins (RNPs) into plant cells (
Dhir et al. 1991;
Lee et al. 2020;
Subburaj et al. 2023). However, standardized protocols for electroporation-based delivery of plasmids and RNPs into milk thistle protoplasts remain limited.
Although shoot regeneration from callus and leaves has been previously reported (
Hetz et al. 1995b;
Rady et al. 2018;
Si-qing et al. 1990), these protocols have yet to be updated or optimized for applications such as genome editing or transient gene expression. Therefore, the seed sterilization and protoplast isolation methods developed in this study provide a foundational platform for
in vitro culture and protoplast-based transformation in milk thistle. Further optimization of transfection and shoot regeneration protocols from protoplasts will be essential to enable functional gene analysis and CRISPR/Cas9-mediated genome editing in this species.
Acknowledgments
This research was supported by Kyungpook National University Development Project Research Fund, 2022.
Fig. 1Effect of different sterilization procedures on milk thistle seeds.
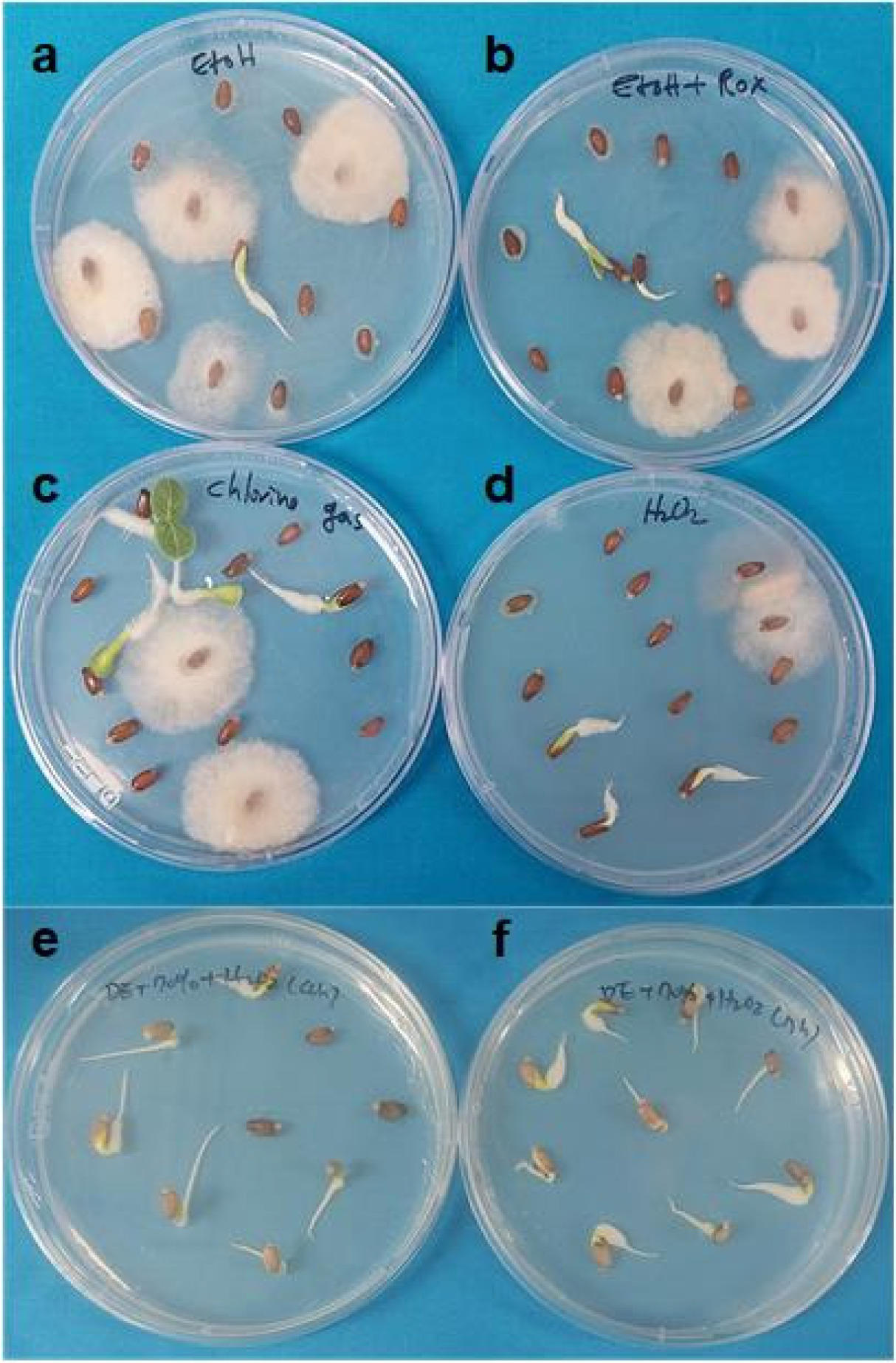
Fig. 2Protoplast isolation from healthy young milk thistle leaves.
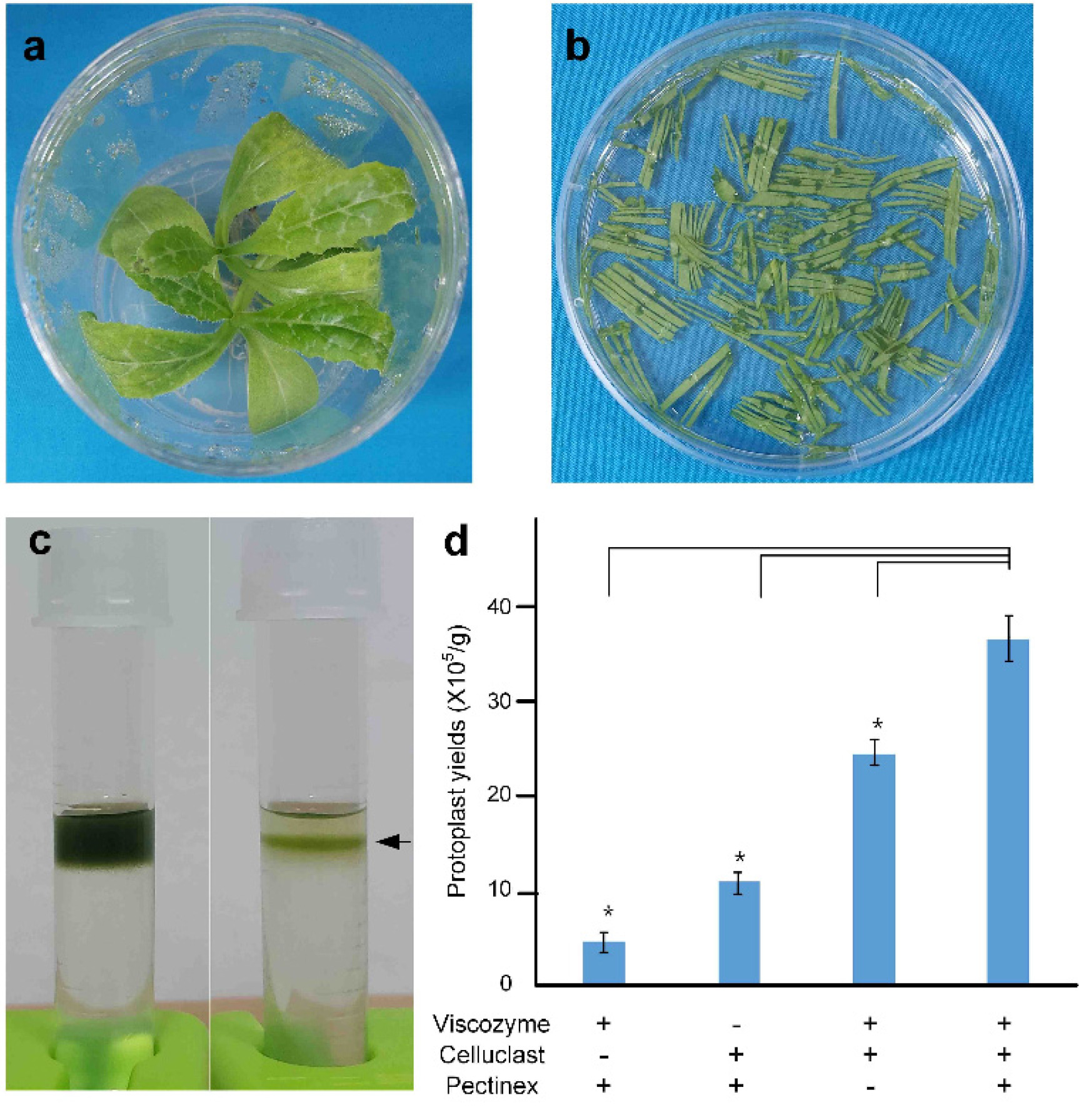
Fig. 3Effect of different digestion durations (1 h, 2 h, 3 h, and 4 h) on protoplast yield and viability.
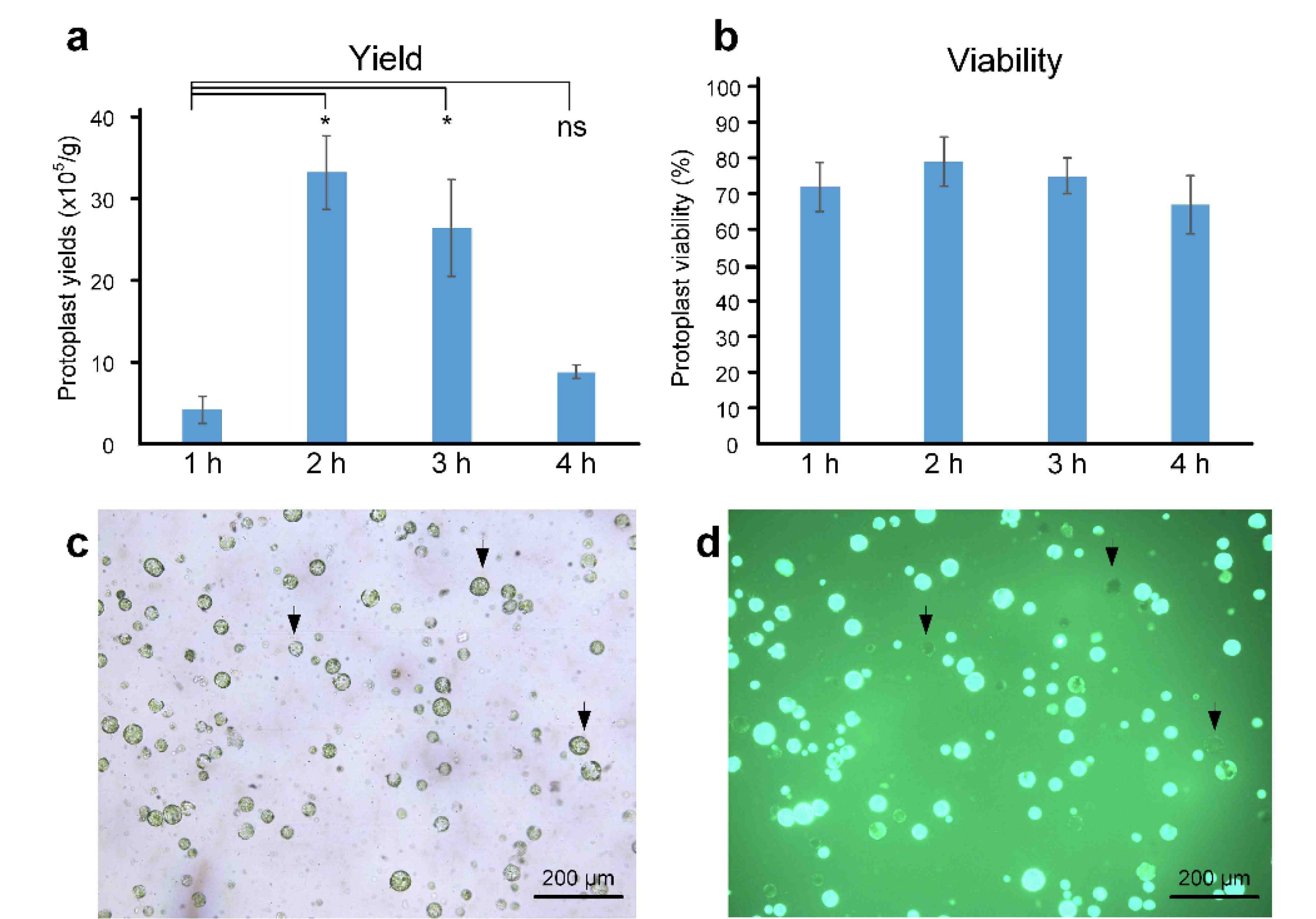
Fig. 4Polyethylene glycol-mediated transfection assay in milk thistle protoplasts.
References
- Abenavoli L, Izzo AA, Milić N, Cicala C, Santini A, Capasso R. 2018. Milk thistle (Silybum marianum): A concise overview on its chemistry, pharmacological, and nutraceutical uses in liver diseases. Phytotherapy Research. 32(11): 2202-2213.
- Andrzejewska J, Martinelli T, Sadowska K. 2015. Silybum marianum: non‐medical exploitation of the species. Annals of Applied Biology. 167(3): 285-297.
- Barampuram S, Allen G, Krasnyanski S. 2014. Effect of various sterilization procedures on the in vitro germination of cotton seeds. Plant Cell, Tissue and Organ Culture (PCTOC). 118(1): 179-185.
- Bart R, Chern M, Park C-J, Bartley L, Ronald PC. 2006. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods. 2(1): 13
- Bhuyan SJ, Kumar M, Ramrao Devde P, Rai AC, Mishra AK, Singh PK, et al. 2023. Progress in gene editing tools, implications and success in plants: a review. Frontiers in Genome Editing. 5: 1272678
- Bijak M. 2017. Silybin, a major bioactive component of milk thistle (Silybum marianum L. Gaernt.)-Chemistry, bioavailability, and metabolism. Molecules. 22(11): 1942
- Burris KP, Dlugosz EM, Collins AG, Stewart CN Jr, Lenaghan SC. 2016. Development of a rapid, low-cost protoplast transfection system for switchgrass (Panicum virgatum L.). Plant Cell Reports. 35(3): 693-704.
- Cabral J, Fevereiro P, Novais J, Pais M. 1984. Comparison of immobilization methods for plants cells and protoplasts. Presented at Enzyme Engineering: Proceedings of the International Enzyme Engineering Conference..
- Chen J, Miao Z, Kong D, Zhang A, Wang F, Liu G, et al. 2024a. Application of CRISPR/Cas9 technology in rice germplasm innovation and genetic improvement. Genes. 15(11): 1492
- Chen K, Wang Y, Zhang R, Zhang H, Gao C. 2019. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annual Review of Plant Biology. 70(1): 667-697.
- Chen W, Zhao X, Huang Z, Luo S, Zhang X, Sun W, et al. 2024b. Determination of Flavonolignan Compositional Ratios in Silybum marianum (Milk Thistle) Extracts Using High-Performance Liquid Chromatography. Molecules. 29(13): 2949
- Clough SJ, Bent AF. 1998. Floral dip: a simplified method for Agrobacterium‐mediated transformation of Arabidopsis thaliana. The Plant Journal. 16(6): 735-743.
- Coy MR, Abbitt SE, Frank MJ. Wang K., Zhang F., 2022. Protoplast Isolation and Transfection in Maize. editors. Protoplast Technology. Methods in Molecular Biology. vol 2464: Humana. New York, NY: pp. 91-104.
- Dhir SK, Dhir S, Sturtevant AP, Widholm JM. 1991. Regeneration of transformed shoots from electroporated soybean (Glycine max (L.) Merr.) protoplasts. Plant Cell Reports. 10(2): 97-101.
- Drouet S, Tungmunnithum D, Lainé É, Hano C. 2020. Gene expression analysis and metabolite profiling of silymarin biosynthesis during milk thistle (Silybum marianum (L.) Gaertn. ) fruit ripening. International Journal of Molecular Sciences. 21(13): 4730
- Duval JR, NeSmith DS. 2000. Treatment with hydrogen peroxide and seedcoat removal or clipping improve germination ofGenesis' triploid watermelon. HortScience. 35(1): 85-86.
- Elwekeel A, AbouZid S, Sokkar N, Elfishway A. 2012. Studies on flavanolignans from cultured cells of Silybum marianum. Acta Physiologiae Plantarum. 34(4): 1445-1449.
- Emadi SA, Rahbardar MG, Mehri S, Hosseinzadeh H. 2022. A review of therapeutic potentials of milk thistle (Silybum marianum L.) and its main constituent, silymarin, on cancer, and their related patents. Iranian Journal of Basic Medical Sciences. 25(10): 1166.
- Hasanloo T, Khavari-Nejad R, Majidi E, Ardakani MS. 2008. Flavonolignan production in cell suspension culture of Silybum marianum. Pharmaceutical Biology. 46(12): 876-882.
- Hetz E, Liersch R, Schieder O. 1995a. Genetic investigations on Silybum marianum and S. eburneum with respect to leaf colour, outcrossing ratio, and flavonolignan composition. Planta Medica. 61(1): 54-57.
- Hetz E, Perales EH, Liersch R, Schieder O. 1995b. Plant regeneration from mesophyll and suspension protoplasts of Silybum marianum. Planta Medica. 61(6): 554-557.
- Jeong YY, Lee H-Y, Kim SW, Noh Y-S, Seo PJ. 2021. Optimization of protoplast regeneration in the model plant Arabidopsis thaliana. Plant Methods. 17(1): 21
- Kang GH, Ko Y, Lee JM. 2025. Enhancing virus-mediated genome editing for cultivated tomato through low temperature. Plant Cell Reports. 44(1): 22
- Kang HH, Naing AH, Kim CK. 2020. Protoplast isolation and shoot regeneration from protoplast-derived callus of Petunia hybrida cv. Mirage rose. Biology. 9(8): 228
- Lee J, Moon S, Jang S, Lee S, An G, Jung K-H, et al. 2020. OsbHLH073 negatively regulates internode elongation and plant height by modulating GA homeostasis in rice. Plants. 9(4): 547
- Li S, Cong Y, Liu Y, Wang T, Shuai Q, Chen N, et al. 2017. Optimization of Agrobacterium-mediated transformation in soybean. Frontiers in Plant Science. 8: 246
- Liu L, Zhang H. 2023. Milk thistle oil extracted by enzyme-mediated assisted solvent extraction compared with n-hexane and cold-pressed extraction. Molecules. 28(6): 2591
- Lizárraga-Paulín E-G, Miranda-Castro S-P, Moreno-Martínez E, Lara-Sagahón A-V, Torres-Pacheco I. 2013. Maize seed coatings and seedling sprayings with chitosan and hydrogen peroxide: their influence on some phenological and biochemical behaviors. Journal of Zhejiang University Science B. 14(2): 87-96.
- Malekzadeh M, Mirmazloum S, Mortazavi S, Panahi M, Angorani H. 2011. Physicochemical properties and oil constituents of milk thistle (Silybum marianum Gaertn. cv. Budakalászi) under drought stress. Journal of Medicinal Plants Research. 5(8): 1485-1488.
- Marceddu R, Dinolfo L, Carrubba A, Sarno M, Di Miceli G. 2022. Milk thistle (Silybum Marianum L.) as a novel multipurpose crop for agriculture in marginal environments: A review. Agronomy. 12(3): 729
- Martinelli T. 2019. Identification of milk thistle shatter‐resistant mutant lines with altered lignocellulosic profile for the complete domestication of the species. Crop Science. 59(5): 2119-2127.
- Murashige T, Skoog F. 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum. 15(3): 473-497.
- Patil GB, Stupar RM, Zhang F. Wang K., Zhang F., 2022. Protoplast Isolation, Transfection, and Gene Editing for Soybean (Glycine max). editors. Protoplast Technology. Methods in Molecular Biology. vol 2464: Humana. New York, NY: pp. 173-186.
- Peluola C, Hoesel S, Crutcher F. 2020. Chlorine gas is an effective alternative to sterilize carnation leaves for Fusarium spp. identification. Journal of Microbiological Methods. 170: 105841
- Rady M, Saker M, Matter M. 2018. In vitro culture, transformation and genetic fidelity of Milk Thistle. Journal of Genetic Engineering and Biotechnology. 16(2): 563-572.
- Rambaldi A, Jacobs BP, Iaquinto G, Gluud C. 2005. Milk thistle for alcoholic and/or hepatitis B or C virus liver diseases. Cochrane Database of Systematic Reviews. (2): 1-18..
- Rosner L, Harrington J, Dreesen D, Murray L. 2003. Hydrogen peroxide seed scarification of New Mexico collections of Ribes cereum. Seed Science and Technology. 31(1): 71-81.
- Samac DA, Tesfaye M, Dornbusch M, Saruul P, Temple SJ. 2004. A comparison of constitutive promoters for expression of transgenes in alfalfa (Medicago sativa). Transgenic Research. 13(4): 349-361.
- Sanna M, Gilardi G, Gullino ML, Mezzalama M. 2022. Evaluation of physical and chemical disinfection methods of Brassica oleracea seeds naturally contaminated with Xanthomonas campestris pv. campestris. Journal of Plant Diseases and Protection. 129(5): 1145-1152.
- Si-qing L, Qi-gui C. 1990. Callus formation from protoplasts and plant regeneration from tissue culture of Silybum marianum Gaertn. Journal of Integrative Plant Biology. 32(1): 19-25.
- Sorokin A, Yadav NS, Gaudet D, Kovalchuk I. 2020. Transient expression of the β-glucuronidase gene in Cannabis sativa varieties. Plant Signaling & Behavior. 15(8): 1780037
- Speciale A, Muscarà C, Molonia MS, Cimino F, Saija A, Giofrè SV. 2021. Silibinin as potential tool against SARS‐Cov‐2: In silico spike receptor‐binding domain and main protease molecular docking analysis, and in vitro endothelial protective effects. Phytotherapy Research. 35(8): 4616-4625.
- Stajič E, Kunej U. 2023. Optimization of cabbage (Brassica oleracea var. capitata L.) protoplast transformation for genome editing using CRISPR/Cas9. Frontiers in Plant Science. 14: 1245433
- Subburaj S, Agapito-Tenfen SZ. 2023. Establishment of targeted mutagenesis in soybean protoplasts using CRISPR/Cas9 RNP delivery via electro− transfection. Frontiers in Plant Science. 14: 1255819
- Tolangi P, Shim J, Sumabat RM, Kim S, Park H-S, Do Kim K, et al. 2024. The genetics and genomics of milk thistle: unlocking its therapeutic potential through modern breeding and biotechnological innovations. Applied Biological Chemistry. 67(1): 115
- Verdaguer B, de Kochko A, Fux CI, Beachy RN, Fauquet C. 1998. Functional organization of the cassava vein mosaic virus (CsVMV) promoter. Plant Molecular Biology. 37(6): 1055-1067.
- Wang J, Wang Y, Lü T, Yang X, Liu J, Dong Y, et al. 2022. An efficient and universal protoplast isolation protocol suitable for transient gene expression analysis and single-cell RNA sequencing. International Journal of Molecular Sciences. 23(7): 3419
- Yao D, Zhou J, Zhang A, Wang J, Liu Y, Wang L, et al. 2023. Advances in CRISPR/Cas9-based research related to soybean [Glycine max (Linn.) Merr] molecular breeding. Frontiers in Plant Science. 14: 1247707
- Yoo S-D, Cho Y-H, Sheen J. 2007. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nature Protocols. 2(7): 1565-1572.
- Zhang X, Liu M, Wang Z, Wang P, Kong L, Wu J, et al. 2024. A review of the botany, phytochemistry, pharmacology, synthetic biology and comprehensive utilization of Silybum marianum. Frontiers in Pharmacology. 15: 1417655
- Zhao F-L, Li Y-J, Hu Y, Gao Y-R, Zang X-W, Ding Q, et al. 2016. A highly efficient grapevine mesophyll protoplast system for transient gene expression and the study of disease resistance proteins. Plant Cell, Tissue and Organ Culture (PCTOC). 125(1): 43-57.