Abstract
The GeBP gene family, known for its GLABROUS1 enhancer-binding proteins, represents a standard transcription factor characterized by an unconventional leucine-zipper motif. The constituents of this family possess a core DNA-affinity domain. This protein is crucial for controlling plant growth and development. GeBP genes have been analyzed and defined only in some Gramineae crops, Arabidopsis, rice and soybean. However, a comprehensive examination of the role of the GeBP genes in Brassica rapa has yet to be conducted. This study used genome-wide characterization to identify twenty BrGeBP genes and accessible RNA sequencing data to evaluate each gene's expression. Based on phylogenetic research, we grouped 114 GeBP proteins from five different plant species into six groups. Generally, groupings of BrGeBP genes with comparable motifs and exon-intron distribution were conserved. The twenty BrGeBP genes were found on six of the ten B. rapa chromosomes, and one paralog showed segmental duplication. The varied protein motifs and gene structures showed that the GeBP genes had various roles in plants. The functions of BrGeBP genes are numerous and include expressions responsive to specific tissues, stress, and hormones according to cis-elements analysis. Furthermore, expression analysis using RNA sequencing revealed that the genes BrGeBP3, BrGeBP9, and BrGeBP20 showed substantially increased expression in different organs. These findings will aid in elucidating the possible roles and traits of BrGeBP genes in the B. rapa biological cycle and related species.
-
Key words: GeBP gene family, Transcription factor; Brassica rapa, Genome-wide analysis, Expression analysis
Introduction
Transcription factors (TFs) are key in controlling the stress response and developmental processes. Under stress, TFs in plants frequently connect the cis-acting component to alter gene expression, eventually preventing or minimizing plant injury. A focused study on TFs has identified their function in stress (abiotic and biotic stress), metabolism (primary and secondary metabolism), and plant development (from seed germination to fruit development). For instance,
NF-Y,
AP2,
TCP,
MYB,
WRKY,
GRF, and
SPL TFs play significant roles in the ability of plants to withstand stress. In contrast,
NAC,
GRF, and
SPL TFs are critical for root development, flower development, seed maturation, and plant transformation (
Samad et al. 2017). One of the newest plant-specific transcription factor families, the glabrous-enhancer-binding protein (
GeBP) was discovered and extracted from
Arabidopsis for the first time in 2003 (
Curaba et al. 2003).
Plant-specific gene regulators with a central DNA-affinity domain are the
GeBP gene family (
Curaba et al. 2003;
Ma et al. 2021). The
GeBP can control the presence of epidermal projections by altering the expression profile of the early gene
GLABROUS1 (
GL1), which was reported to govern the commencement of epidermal hair formation in
Arabidopsis (
Perazza et al. 1998). According to a study of conserved motifs, the
Arabidopsis GeBP sequencing showed a leucine zipper region and a basic amino acid region. This evidence indicated that GeBP might be a part of the
bZIP gene regulatory family. However, it was discovered that there are more than nine residues between these two conserved domains, which does not fit the typical criteria of a bZIP protein (
Huang et al. 2021;
Jakoby et al. 2002). A typical bZIP conserved domain's basic structure is N-x7-R/ K-x9-L-x6-L-x6-L, which separates the Leu zipper domain from the alkaline domain by nine residues. Researchers have concentrated on the parallels between the GeBP/GeBP-LIKE development of a distinct clade in
Arabidopsis and an unexplained extra-endian invariant area (
Curaba et al. 2003). Plants can adjust to complicated environmental changes because their root systems are extremely pliable while developing (
Khare et al. 2017). Abiotic stresses such as heavy metal pollution at sub-lethal levels cause plants to demonstrate avoidance behavior in the form of altered root architecture (
Clemens 2006).
Arabidopsis thaliana responds to cadmium (Cd) by rapidly inducing the
GeBP-LIKE 4 (
GPL4) transcription factor, which has been recognized as a root development inhibitor. When plants are subjected to hazardous metals, GPL4 reduces the concentrations of reactive oxygen species (ROS), slows down root growth. This condition settled the roots in the uncontaminated areas (
Khare et al. 2017). Moreover, GPL4 root avoidance is necessary for
Arabidopsis to respond to excessive concentrations of zinc (Zn) and copper (Cu), and to respond to Cd stress.
A putative membrane protein is encoded by the
Arabidopsis gene known as
CONSTITUTIVE EXPRESSOR OF PATHOGENESIS-RELATED GENES5 (
CPR5), which also has pleiotropic roles, notably in pathogen reactions, cell expansion, cell proliferation, and cellular death (
Bowling et al. 1994;
Brininstool et al. 2008). In 2011, Gilles Vachon et al. discovered a connection between
CPR5 and the
GeBP gene family. In the
CPR5 pathway,
GeBP/
GPLs controlled several genes (
Perazza et al. 2011). The transcriptome profile of the
gebp gpl 1, 2, 3 triple mutant and the transgenic overexpression line of GPL2 has allowed researchers to determine the role of the
GeBP/
GPLs genes in cell wall metabolism and defense responses. CPR5 partially controls a portion of the
GeBP members.
GeBP/
GPLs are
PR gene repressors because the mutant has elevated gene expression levels for the pathogen-responsive signaling genes
PR1 and
PR5. According to a recent study, the
Arabidopsis GeBP-like transcription factor VFP4 interacted with the
Agrobacterium F-box effector VirF and its plant operative equivalent VBF (
García-Cano et al. 2018).
Brassica crops serve as sources of human nutrition and offer chances to study genome evolution. Brassica vegetables primarily include
Brassica rapa and
Brassica oleracea.
B. rapa consists of several subspecies, such as Chinese cabbage (
B. rapa ssp.
pekinensis), non-heading Chinese cabbage (
B. rapa ssp.
chinensis), and turnip (
B. rapa ssp.
rapifera). It is a diploid species (AA, 2n = 20) and serves as one of the progenitors of key allotetraploid oilseed crops, including
B. juncea (AABB, 2n = 36) and
B. napus (AACC, 2n = 38) (
Nagaharu et al. 1935).
B. rapa varieties are valuable for enhancing the genetic diversity of these domesticated species. With a short life cycle, minimal growth requirements, and sensitivity to environmental factors,
B. rapa (like Wisconsin Fast Plants) and other rapid-cycling Brassicas are widely used in education and research. One of the rare subspecies of
Brassica rapa is Chinese cabbage (
Brassica rapa ssp.
pekinensis) (
Song et al. 2014). Its variation Chiifu-401-42 genome has undergone sequencing (
Wang XiaoWu et al. 2011). While more than 40,000 proteins have been described, the sequences were organized based upon their significant commercial importance and tight ties to
Arabidopsis (
Wang XiaoWu et al. 2011). The haploid genome of
Brassica rapa is approximately 529 Mb, making it much larger than the 125 Mb haploid genome of
Arabidopsis (
Johnston et al. 2005).
Arabidopsis and
Brassica share a common ancestor, having diverged around 14.5-20.4 million years ago (
Bowers et al. 2003). Comparative genetic mapping has shown collinear chromosome segments within the Brassicaceae family and conserved linkage patterns between
Arabidopsis and
Brassica (
Acarkan et al. 2000a;
Acarkan et al. 2000b;
Bohuon et al. 1998;
Lukens et al. 2003). This makes
B. rapa a valuable model for studying the genomics and evolutionary history of Brassica species.
Consequently, a genome-wide investigation was carried out to identify the B. rapa GeBP gene family. The 20 GeBP genes that were discovered underwent additional examination for their protein sequences, gene characterization, structural organization, conserved motif analysis, chromosomal localization, phylogeny, promoter cis-elements, and the investigation of gene expression using transcriptomic data. These bioinformatic investigations will offer valuable insights into the role of BrGeBP genes in response to abiotic stresses and provide essential genetic resources to support biological breeding efforts.
Materials and Methods
GeBP genes in Brassica rapa: genome-wide identification
To do a comprehensive genomic analysis of the
GeBP gene family, we extracted 20
GeBP genes of
B. rapa. An HMM test was performed to verify their identities using the HMMER web server (
Potter et al. 2018). All sequences, including coding DNA (CDS), protein, and genomic sequences of
B. rapa, were downloaded from the BRAD database (
Cheng et al. 2011). iTAK database (
Zheng et al. 2016), and Ensembl Plants database (
Kersey et al. 2016) were used to confirm the sequences found. To find out about molecular characteristics including length, molecular weight (Mw), iso-electric point (pI), and grand average of hydropathicity (GRAVY) of GeBP proteins, ProtParam tool (
Gasteiger et al. 2005) was employed. Ensembl Plants database (
Kersey et al. 2016) was utilized to discover the Open Reading Frame (ORF) of
GeBP genes. The subcellular localization of the identified proteins was anticipated using Cell-PLoc 2.0 (
Chou et al. 2010). The SMART conserved domain search tool (
Letunic et al. 2018) was used to anticipate the site of the GeBP domain in twenty
BrGeBP genes. The protein sequences of
Arabidopsis (20),
B. oleracea (18),
B. napus (36), and Radish (20) (
Supplementary Data File S1) were collected from iTAK databases (
Zheng et al. 2016) (
Table 1).
To investigate the evolutionary connections of GeBP family proteins among the five-plant species, phylogenetic analysis of GeBP proteins of
Arabidopsis thaliana,
B. rapa,
B. oleracea,
Raphanus sativus, and
B. napus was performed. The phylogenetic tree was assembled employing MEGA7.0 by the Neighbor-Joining (NJ) method (
Saitou et al. 1987) considering the parameter of pair-wise deletion and 1000 bootstrap replications. A synteny map was generated using the Circoletto database (
http://tools.bat.infspire.org/circoletto) to display the similarities among the protein sequences of
Brassica rapa,
Brassica oleracea,
Brassica napus, and
Arabidopsis thaliana.
The BrGeBP protein motifs were analyzed using MEME suite 5.1.0 (
Bailey et al. 2009). The pre-defined analysis settings were a minimum width of 6 and a maximum width of 50, with a maximum of 10 motifs each. The exon-intron distribution of the CDSs of
GeBP genes was analyzed using the gene structure display server (GSDS) web tool (
Hu et al. 2015).
The chromosomal positions, including start-end coordinates and chromosome numbers, of the 20
BrGeBP genes were compiled from the Ensembl Plants database (
Kersey et al. 2016) and the MapGene2Chrom web v2 tool (
Chao et al. 2015) was employed to plot their locations on all 10 chromosomes of
B. rapa. To determine the gene duplication of
BrGeBP genes, an NCBI BLAST search (
Zhang et al. 1997) was carried out based on the proportion of query cover percentage among 20
BrGeBP genes.
The PlantCARE web-based tool (
Lescot et al. 2002) was used to come up with a list of possible cis-regulatory elements of 20
BrGeBP genes. Each cis-element was categorized based on its association with stress response, hormonal regulation, growth and development. The frequency of each element was recorded for each
BrGeBP gene, and the data were systematically organized in Microsoft Excel. To improve visualization, conditional formatting was applied to highlight values, providing a clear and intuitive representation of the variations in cis-element distribution across genes. Cis-regulatory elements were found in the area between each gene's start codon and the 2000 bp upstream sequence (
Wu et al. 2012). Such upstream regions that modulate target genes contain cis-elements coupled to transcription factors (
Fang et al. 2008).
BrGeBP RNA-sequencing data was retrieved from the Expression Atlas database (
https://www.ebi.ac.uk/gxa/ home) (
Petryszak et al. 2016).
Tong et al. (2013) had previously gathered and processed that data. Six types of tissues such as leaf, stem, flower, silique, root, and callus were examined (
Tong et al. 2013). The transcript quantity is measured in fragments per kilobase of exon model per million mapped reads (FPKM) values. Microsoft Office Excel created a heat map for the
BrGeBP genes using these values.
Results
GeBP Genes in Brassica rapa: genome-wide identification
A total of 20
GeBP genes were identified and according to their gene id, genes were named as
BrGeBP1 to
BrGeBP20 where 'B' for
Brassica and 'r' for
rapa (
Table 2). To establish the veracity of the conserved domains, the SMART conserved domain search tool (
Letunic et al. 2018) was used to verify their presence. DUF573 domains were found in 17 out of 20 genes (
Table 2). Three candidate genes (
BrGeBP16,
BrGeBP19, and
BrGeBP20) were eliminated due to their incomplete domain structure. The protein length varied from 299aa, found in both
BrGeBP11 and
BrGeBP12 (shortest protein) to 640aa found in
BrGeBP10 (longest protein) (
Table 2). These proteins have molecular weights that range from 33.63 kda (
BrGeBP11) to 72.81 kda (
BrGeBP10). The theoretical pI values ranged from 4.56 (
BrGeBP4) to 8.95 (
BrGeBP2). Five proteins (
BrGeBP2,
BrGeBP3,
BrGeBP6,
BrGeBP7, and
BrGeBP10) have pI values above 7, demonstrating that they were basic, while the proteins generated by other
GeBP genes were acidic (pI values <7) (
Table 2). The CDS of
GeBP genes had a wide range of lengths, ranging from 735 bp (
BrGeBP14) to 1923 bp (
BrGeBP10) (
Table 1). In addition, the GRAVY values differed from -0.263 to -1.087 (
Table 2). The GRAVY scores for all of these proteins were negative, illustrating that they were all hydrophilic in nature. In the subcellular location, 19 proteins were expected to be nuclear and one protein (
BrGeBP2) was predicted to be endoplasmic reticulum (
Table 2). It can be deduced that GeBP proteins have a wide range of physiochemical characteristics.
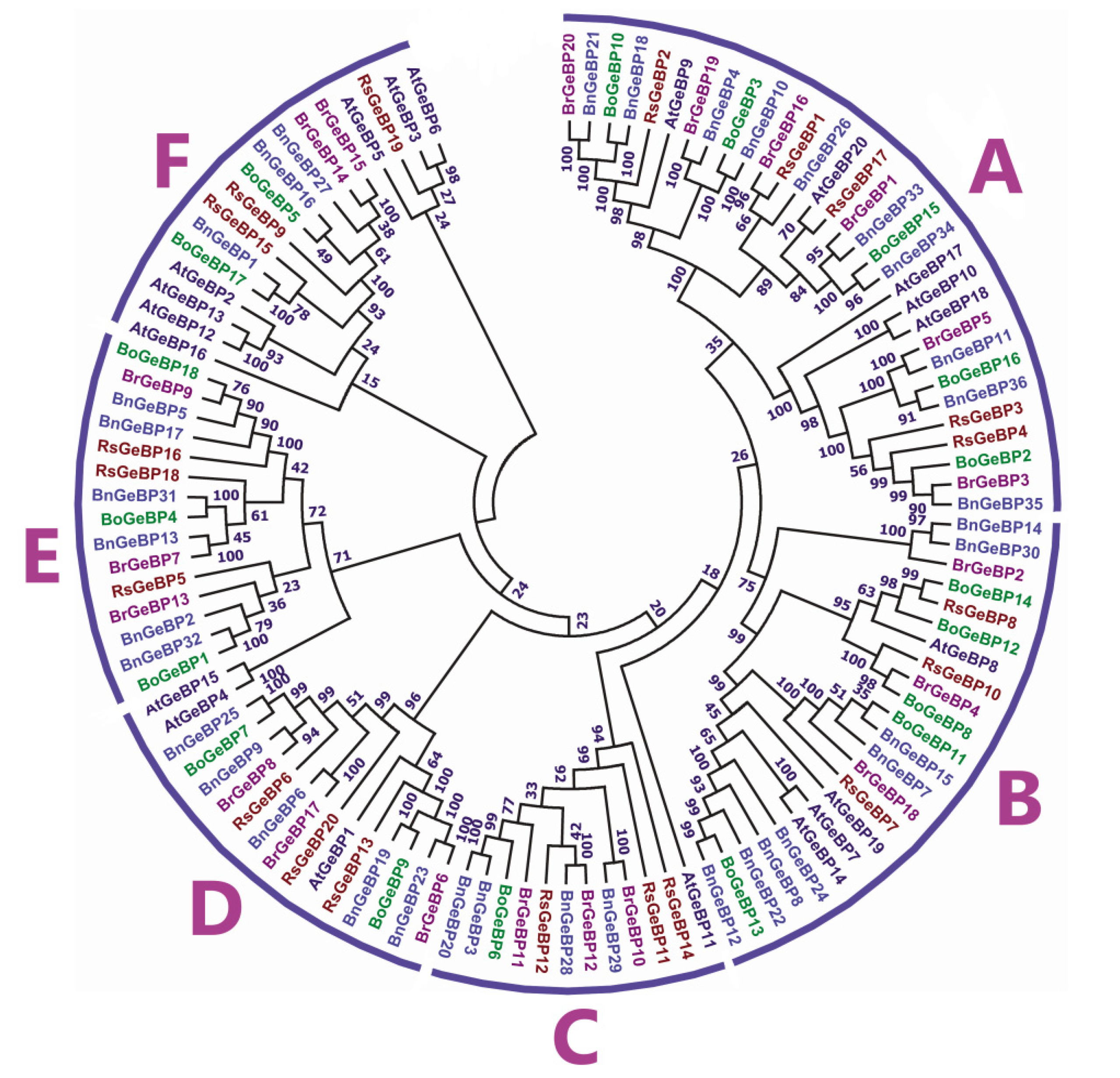
A phylogenetic tree was built to gain insight into the evolutionary associations among GeBP family proteins from 114 amino acid sequences of
B. rapa (20),
B. oleracea (18),
B. napus (36),
Arabidopsis thaliana (20), and
Raphanus sativus (20) (
Fig. 1). While majority of the nodes had more believable bootstrap values, certain nodes' bootstrap values were low due to low species identity. The GeBP proteins from different species were clustered into six groups, marked as A, B, C, D, E, and F (
Fig. 1). Among these, group A comprised 31 members which is 27% of the total
GeBP genes, group B comprised 23 members which is 20% of the total
GeBP genes, group C contained 12 members which is 10% of the total
GeBP genes, succeeded by group D (16 members), group E (15 members), and group F (17 members) which are 13% of the total
GeBP genes (
Fig. 1). Moreover, the
GeBP genes from the dicot were closely grouped in the phylogenetic tree.
The maximum number of
BrGeBP was found in group A, which had six members, and the lowest count was found in group F with two members (
Fig. 1). In the tree, orthologous pairs represented the majority of BrGeBP proteins (i.e. BrGeBP20/BnGeBP21, BrGeBP19/ BnGeBP4, BrGeBP16/RsGeBP1, BrGeBP1/BnGeBP33, BrGeBP5/ BnGeBP11, BrGeBP3/ BnGeBP35, BrGeBP4/ BoGeBP8, BrGeBP10/ BnGeBP29, BrGeBP12/ BnGeBP28, BrGeBP6/ BnGeBP23, BrGeBP17/ BnGeBP6, BrGeBP8/ BnGeBP9, BrGeBP7/ BnGeBP13, and BrGeBP9/ BoGeBP18) (
Fig. 1). On the other hand, only one paralogous pair of BrGeBP proteins was detected in the tree (i.e., BrGeBP14/ BrGeBP15) (
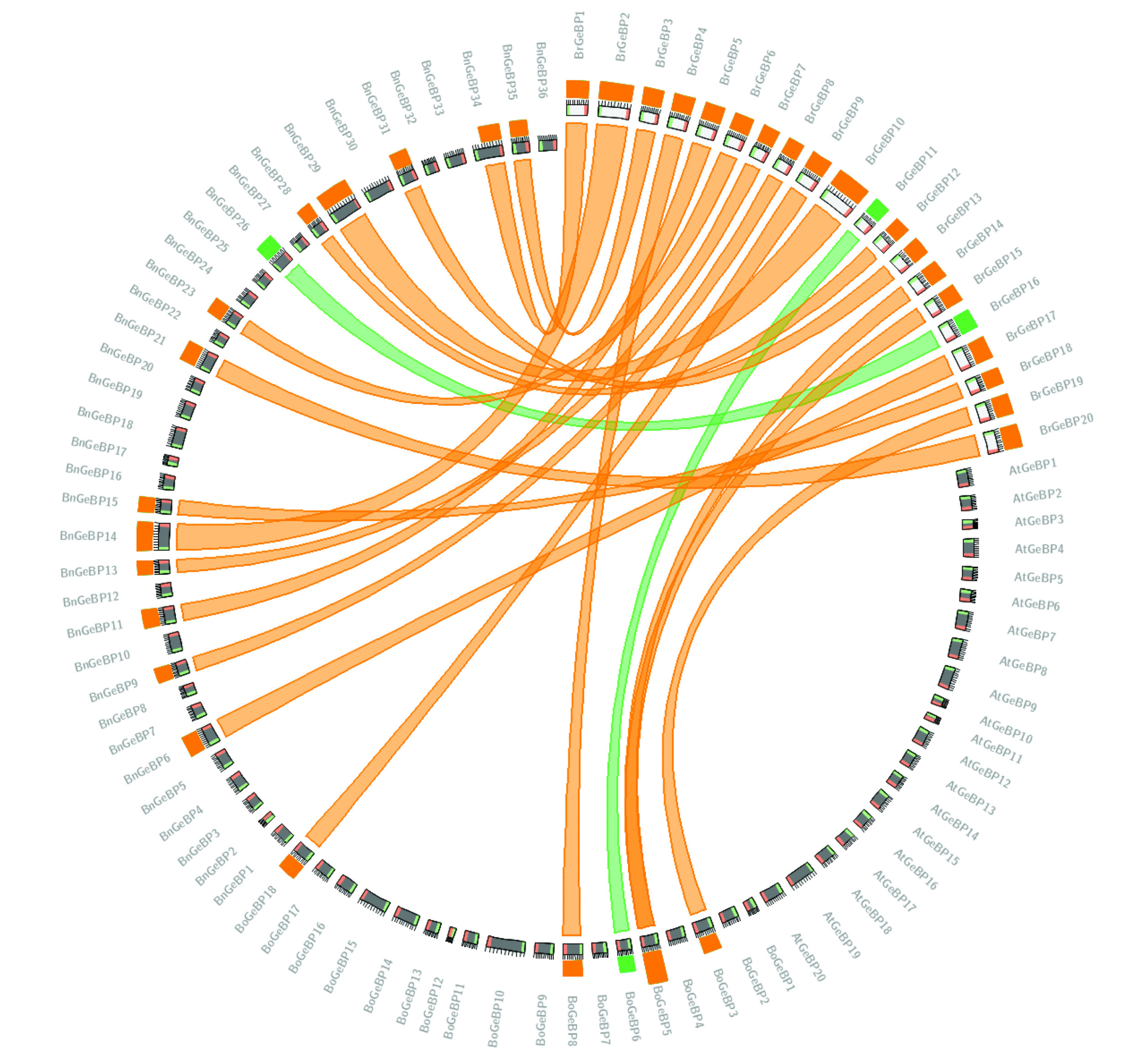
Fig. 1). Furthermore, the synteny analysis explored the close physical proximity of genes on a chromosome and the syntenic relationships between BrGeBP proteins and those of related species. The green ribbon connecting BrGeBP11/BoGeBP6 and BrGeBP16/BnGeBP26 indicates a sequence similarity of ≤75% (
Fig. 2). In contrast, the remaining orthologous pairs, linked by orange ribbons, represent higher sequence similarities of up to ≤99.99%. Some examples include BrGeBP1/BnGeBP34, BrGeBP2/BnGeBP14, BrGeBP3/BnGeBP35, and BrGeBP4/BoGeBP8 (
Fig. 2). This demonstrates that
Brassica rapa shares a close syntenic connection with other
Brassica species.
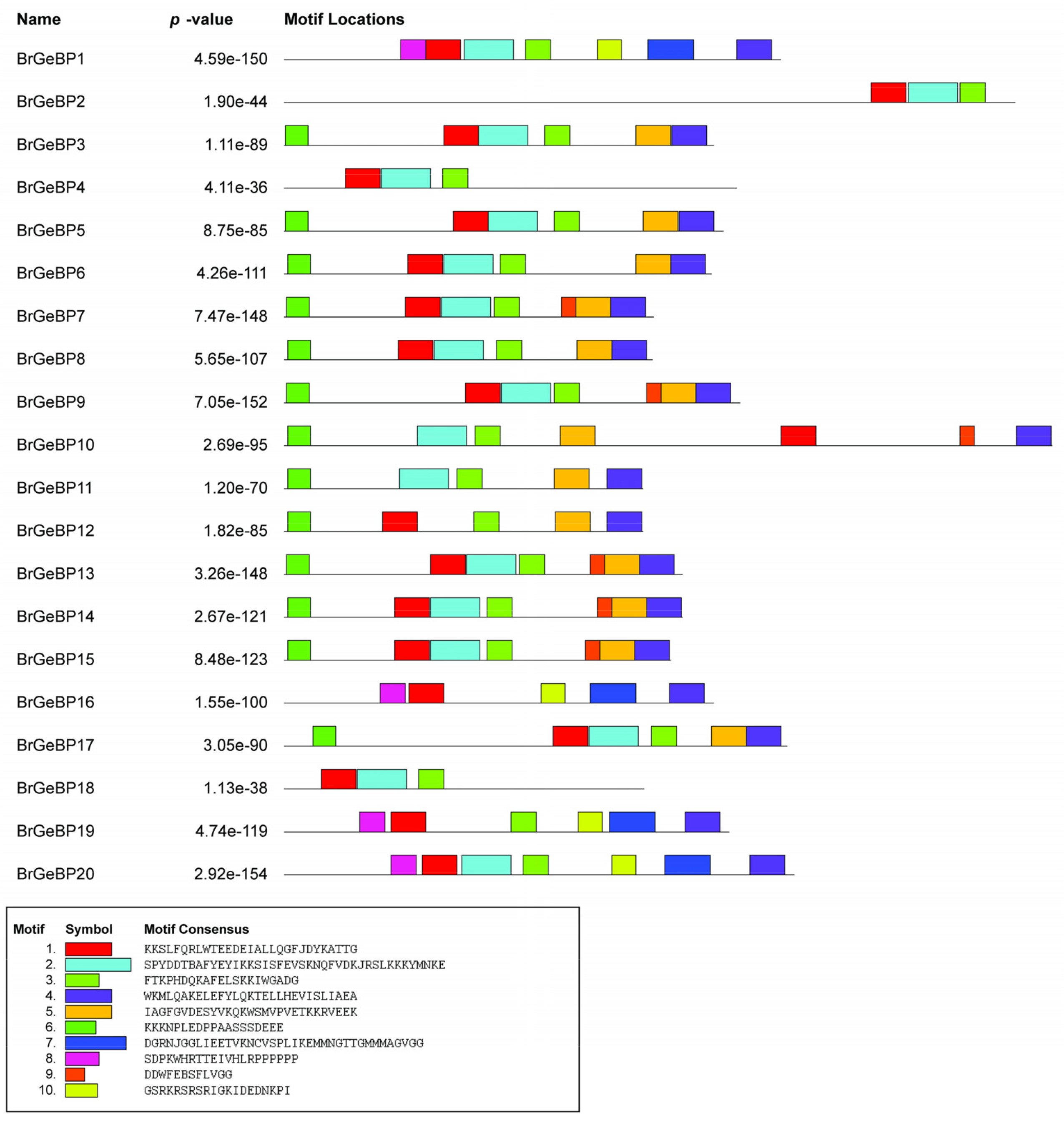
To more thoroughly explore the variation of the
BrGeBP genes, we conducted the motif analysis of the BrGeBP proteins. There was diversity in the motif distribution among the 20
GeBP genes. Ten conserved motifs were discovered through the MEME online server-based motif analysis. From
Fig. 3, we noticed that motif 1 and motif 3 were present in 19 proteins which were the most common motifs among 10. On the other hand, motifs 5 and 6 were observed on 7 proteins whereas motifs 2 and 4 were absent on 3 proteins (
Fig. 3). Besides that,
BrGeBP2 and
BrGeBP18 contained only 3 motifs among 10 motifs (
Fig. 3). Members of the evolutionary tree that shared the same group have similar motif compositions (
Figs. 1,
3). Group A contained 8 motifs including motifs 1, 3, 4, 5, 6, 7, 8, and 10 whereas motif 4 was absent in group B and motif 10 was absent in group F (
Figs. 1,
3). The motif distribution verified that the BrGeBP proteins had not changed over time.
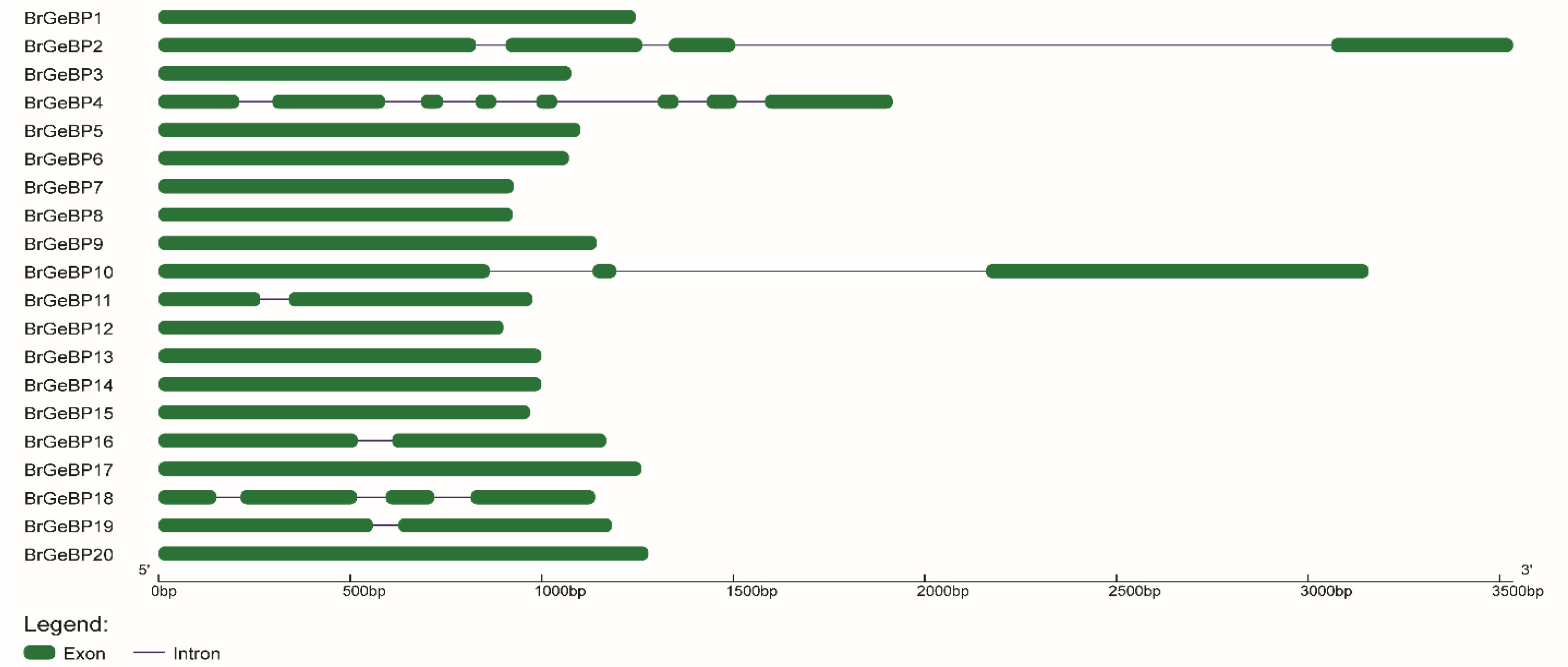
It is well-accepted that the evolution of multigene families is driven by structural variation within genes. One of the crucial structural elements that play a role in the evolution of a given gene is the distribution of exons and introns. Comparable exon-intron distributions suggest functional commonality. Additionally, exonization, insertion/deletion, and addition or removal of exon/intron are three major types of processes that contribute in diverse ways to structural differences in the exon-intron architecture. The exon-intron distribution of each
BrGeBP gene was inferred to obtain a clearer perspective of the structural variation of the
GeBP genes in
B. rapa (
Fig. 4). There were only seven
BrGeBP genes that comprised introns in their coding regions which ranged from one to seven.
BrGeBP4 contained the highest seven introns whereas
BrGeBP11 contained only one intron (
Fig. 4). Most individuals of a group that were linked to one another had nearly identical exon-intron arrangements, either in terms of intron counts or exon dimensions. Group D, E, and F contained only exons which was the equivalent distribution of exons with varying lengths (
Fig. 4). However, compared to other members of group A,
BrGeBP16 and
BrGeBP19 have a varied number of exons and introns (
Fig. 4).
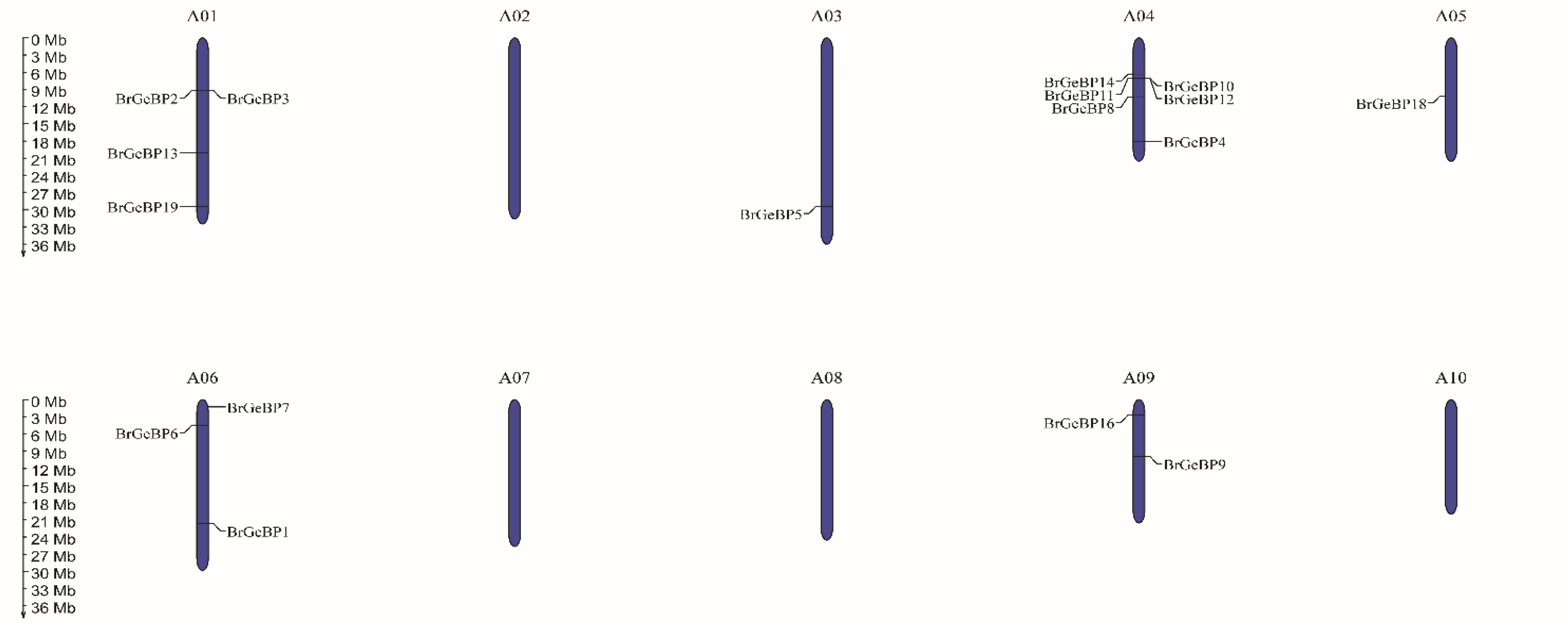
The physical location of each gene was identified after the 20
BrGeBP genes were mapped onto the 10 chromosomes of
B. rapa (
Fig. 5). On six of the ten chromosomes,
BrGeBP genes were disproportionally distributed. The most
BrGeBP genes were found on chromosome A04, which had six of them followed by chromosomes A01 (four genes) and A06 (three genes). Chromosome A09 contained two genes, while chromosomes A03 and A05 contained only one gene (
Fig. 5). On the other hand, there were no genes found on the rest of chromosomes A02, A07, A08, and A10 (
Fig. 5). Besides that, the chromosomal locations of
BrGeBP15,
BrGeBP17, and
BrGeBP20 were scaffolded which was not identified specifically (unknown).
The sequence identity among the 20 GeBP proteins of
B. rapa ranged from 22.14% (between BrGeBP7 and BrGeBP19) to 93.37% (between BrGeBP14 and BrGeBP15) (
Supplementary Table S1). If two genes were found in an area of 100 kb of a chromosome and spaced by only four or fewer other genes, they were considered as tandemly duplicated genes (
Wang et al. 2015). Genes were considered to be segmentally duplicated if their percentage of query cover and identity was ≥ 80 (
Kong et al. 2013). Depending on the degree of identity and query coverage (more than 80%) of the
BrGeBP genes, a single pair of paralogous segmentally duplicated
BrGeBP genes was discovered (
Supplementary Table S1). One gene was found on chromosome A04 and another gene was scaffolded (
Fig. 5). According to the evolutionary relationships depicted in
Fig. 1, the duplicated gene pairs are members of the same group. None of the genes had any tandem duplication.
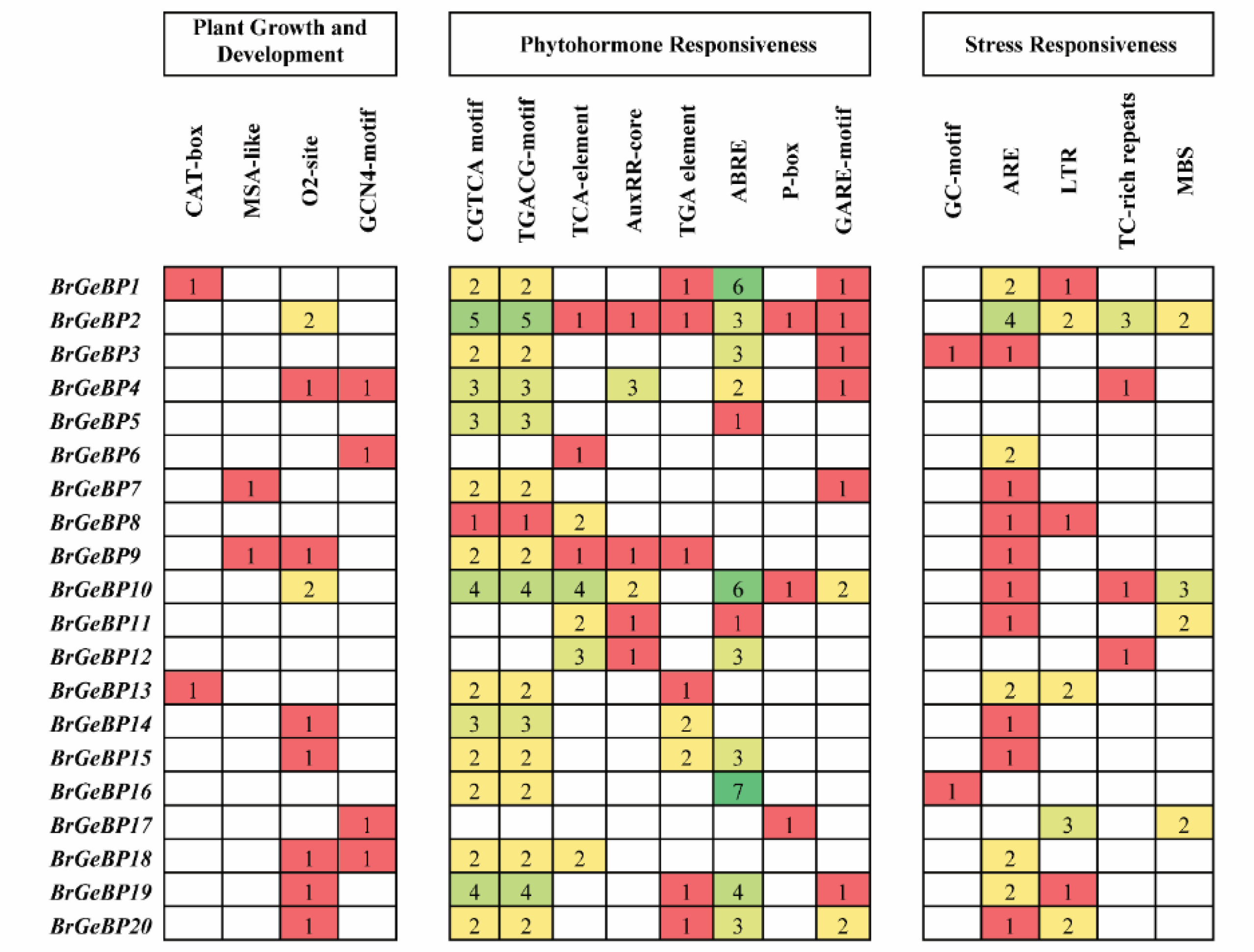
During all stages of a plant's life cycle,
cis-elements (CEs) are crucial for the transcriptional control of genes. CEs were discovered in the promoter areas of the 20
GeBP genes in
B. rapa to better comprehend the transcriptional control mechanisms. Plant growth and development, phytohormone responsiveness, and stress responsiveness were the three kinds of responses among 20 CEs (
Supplementary Table S2). Here, we found four CEs in response to growth and development (
Fig. 6). CAT-box (for meristem expression) was found on two genes, MSA-like (for cell cycle regulation) was found on two genes, O2-site (for controlling zein metabolism) was present in nine genes, and GCN4-motif (for expressing endosperm) was present in four genes (
Fig. 6). Five cis-elements responded to stress: MBS (MYB binding site involved in drought-inducibility) was present in four different
BrGeBP genes, LTR (low-temperature responsiveness) was present in seven genes, TC-rich repeats were present in four genes, GC-motif was present in two
BrGeBP genes, and ARE (anaerobic induction) was present in 15
BrGeBP genes that responded to both defense and stress (
Fig. 6). Additionally, we discovered different hormone-sensitive cis-elements: salicylic acid (TCA-element) in eight genes, auxin (TGA-element) in eight genes, abscisic acid (ABRE) in 12 genes, and gibberellin (GARE-motif) in eight genes (
Fig. 6). Furthermore, the majority of the
BrGeBP genes in the same phylogenetic group have comparable cis-acting components in their promoter zones, indicating a potential role for this gene family in growth and resilience to stress (
Figs. 1,
6).
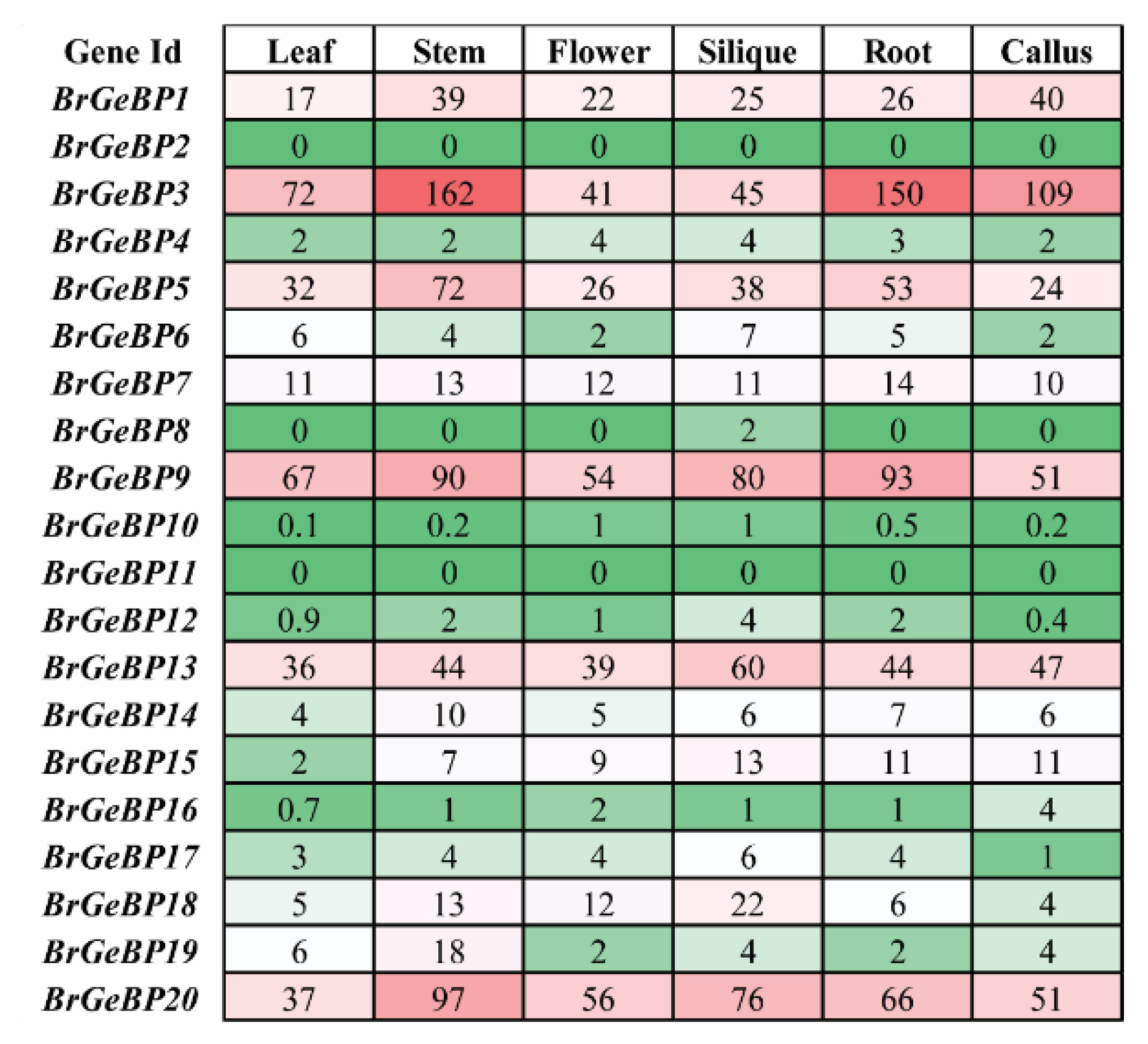
A gene family's tissue-specific expression profile can reveal information about its putative developmental functions. To figure out which
GeBP genes are involved in
B. rapa's tissue or organ development regulation, the expression levels of the
BrGeBP in six tissues (leaf, stem, flower, silique, root, and callus) were investigated based on FPKM values derived from RNA-sequencing data in
B. rapa that Tong et al. (2013) had previously examined. According to the findings, six genes (
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13, and
BrGeBP20) were expressed at varying levels in all the tissues (
Fig. 7). Mostly
BrGeBP3,
BrGeBP9, and
BrGeBP20 showed notably higher expression levels among the twenty genes. Some genes have distinct expression patterns in different tissues to some extent, for example-
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13,
BrGeBP18, and
BrGeBP20 prominently expressed during the reproductive stage, particularly in flower tissue (
Fig. 7). The transcript levels of seven genes, namely-
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13, and
BrGeBP20 were prominently expressed in leaves, eight genes (
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13,
BrGeBP19, and
BrGeBP20) in stems, seven genes (
BrGeBP1, BrGeBP3, BrGeBP5, BrGeBP9, BrGeBP13, BrGeBP18, and
BrGeBP20) in siliqua, Six genes in roots (
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13, and
BrGeBP20), and six genes (
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13, and
BrGeBP20) in callus compared to other genes (
Fig. 7). In contrast, two genes (
BrGeBP2 and
BrGeBP11) were not expressed in any organs out of six (
Fig. 7). Besides that,
BrGeBP10 was very weakly expressed in the different organs examined (
Fig. 7). In
B. rapa tissues, we discovered that the majority of the individuals belonging to the same phylogenetic grouping showed a comparable expression profile (
Figs. 1,
7). In particular, the genes from group A exhibited greater expression, and group C showed lower expression compared to other groups (
Figs. 1,
7).
Discussion
The opportunity to customize the mechanisms and functions of genes associated with plant growth, development, and stress tolerance is expanding as genome sequencing initiatives advance more rapidly. A useful method for examining gene function, structure, and evolution is now the study of the gene family. The ensuing findings imply that
GeBP will support the hormone cascade as a TF. First, the GeBP protein connects to the cis-acting region of the
GLABROUS1 gene, a MYB gene involved in determining epidermal cells, which is controlled by the cytokinin hormone GA (
Gan et al. 2007;
Oppenheimer et al. 1991;
Perazza et al. 1998). Second, the KNOTTED1 homeodomain (
KNOX) family gene BREVIPEDICELLUS (
BP) (
Curaba et al. 2004), which positively controls the apical meristem of the shoot contains the cytokinin pathway, positively influences the transcript levels of
GeBP (SAM) (
Jasinski et al. 2005;
Yanai et al. 2005).
Arabidopsis, rice, apple, moso bamboo and soybean have all undergone a preliminary investigation of the
GeBP gene family. Therefore, it is feasible and essential to characterize the
GeBP genes in
B. rapa at the genome level.
The number of
GeBP genes in
B. rapa was slightly lesser than in rapeseed (36) but equivalent to that of
Arabidopsis (20) and Radish (20), which was higher than that of cabbage (18). In addition, the genome size of
B. rapa (283.8 Mb) (
Wang XiaoWu et al. 2011), and
Arabidopsis (125 Mb) (
Hu et al. 2011), but the genome size of cabbage is about 530 Mb (
Liu et al. 2014) which is higher than
B. rapa. So, the size of the crop genome is unrelated to the quantity of
GeBP genes.
From the phylogenetic analysis, it was revealed that the evolutionary ties between the BrGeBP and BnGeBP proteins were found to be closer in contrast to other crop GeBP proteins (
Fig. 1). This result implied that
B. rapa and
B. napus might have shared a functional progenitor and evolved from them. In the tree, several paralogous and orthologous relationships were found. The term "paralog" refers to identical genes that have evolved within a single species and were due to gene duplication, which causes the creation of new genes with novel functions (
Gabaldón et al. 2013). Instead, following a speciation event, orthologs diverged yet still perform similarly to the ancestor gene's function (
Gabaldón et al. 2013). As one paralog gene pair of BrGeBP proteins was identified, indicating highly functional similarity (
Fig. 1). The evolutionary tree of BrGeBP proteins and the similarity in gene structure and motifs among each category provided more evidence for the links between the proteins.
Besides the function in gene family evolution, we examined the conserved motifs. Surprisingly, out of 20 GeBP proteins, 19 of them had motif 1 and motif 3 demonstrating the potential importance of these motifs in regulating target gene expression by transcripts during evolution (
Fig. 3). Since the bulk of the BrGeBP proteins sharing the same group contained comparable patterns, gene expansion was likely the cause of their creation. The diverse patterns of motif distribution among the various groups revealed the factors of differentiating functions in
GeBP genes throughout their evolutionary background. In addition, the gene structure divergence offers additional proof for analyzing phylogenetic connections. As a result, each member of the
B. rapa GeBP family's exon/intron structure and evolutionary connections were examined (
Figs. 1,
4). The consistent exon-intron distribution across various groups revealed that these genes had undergone substantial evolutionary conservation. Moreover, the functional divergence of
BrGeBP may be explained by variations in intron number.
Genetic material is multiplied throughout selection and alterations, mostly by instances of gene duplication and divergence (Pickett et al. 1995). Tandem and segmental duplication, together with transpositional events, are the primary reasons for the gene family growth in plants (
Cannon et al. 2004;
Li et al. 2014). These include, segmental duplication is more prevalent among them because the majority of plants are polyploids with duplicate chromosomes and are diploidized (
Cannon et al. 2004). In this investigation, no pair of genes' tandem duplications have been discovered, demonstrating that the growth of the
GeBP gene family in
B. rapa has been mostly driven by segmental duplication as opposed to tandem duplication (
Supplementary Table S1). The phylogenetic tree revealed that overlapping genes belonged to the same groupings, demonstrating the occurrences of group-specific gene duplications during evolution.
It may be possible to learn more about the roles of
BrGeBP genes at various plant developmental stages by studying the promoters in the upstream area of genes owing to their significant functions in regulating gene expression throughout development or in the environment (
Zhu et al. 2014). They are crucial switches at the molecular level that contribute to the regulation of the transcription of genes and operate a vast network of genes engaged in a range of biological functions, such as the effects of stress on development (
Zhang et al. 2005). There are numerous cis-acting factors in plants that are connected to stress (
Saeediazar et al. 2014). In this study, every
BrGeBP gene contains a single or many phytohormones cis-elements listed below, which increases the likelihood that each of the
BrGeBP genes has functions related to stress tolerance (
Fig. 6). These elements include gibberellin, auxin, salicylic acid, abscisic acid, and methyl jasmonate (
Fig. 6). To respond to stressors, phytohormones assemble many regulatory cascades as intracellular transmitters (
Wolters et al. 2009). The gene promoter regions contained cis-elements that respond to stress hence describing their purpose in environmental adaptation and abiotic stress.
It has been proposed that transcription factors are important regulators in controlling how genes express themselves in response to different environmental stressors (
Chen et al. 2021). Twenty
BrGeBP gene expression profiles from various tissues were examined to determine the potential roles of these genes (
Fig. 7). The RNA-seq analysis revealed that three different expression patterns were found among the twenty
BrGeBP genes, comprising tissue-specific expression patterns, constitutive expression, and no expression (
Fig. 7). Two
BrGeBP genes (
BrGeBP2 and
BrGeBP11) were not expressed in any of the tissues analyzed, posing the possibility that they are pseudogenes or that they are only produced during particular developmental stages or under particular circumstances that were not examined in our research.
BrGeBP3 is predominantly expressed in stem, siliqua, and callus tissues, indicating involvement in the growth and development of stem and siliqua, and also engaged in the formation of roots and/or the intake of nutrients and water (
Fig. 7). The
BrGeBP3 gene, identified as being associated with drought tolerance (
Wang et al. 2023), can be targeted to enhance nutrient uptake and improve resilience to drought conditions. Similar results obtained from
BrGeBP9 and
BrGeBP20 show that these genes can be harnessed to develop cabbage and related crops with enhanced adaptability, increased yield, and greater stress tolerance using advanced breeding techniques such as gene editing. Additionally,
BrGeBP genes can beleveraged in biotechnological applications, where their targeted modification could enhance oilseed yield, nutritional quality, and stress resilience, benefiting industries reliant on
Brassica crops. As the six genes (
BrGeBP1,
BrGeBP3,
BrGeBP5,
BrGeBP9,
BrGeBP13, and
BrGeBP20) showed preferential expression in all the tissues (leaf, stem, flower, silique, root, and callus), they could play a variety of regulatory roles in
B. rapa's growth and development (
Fig. 7). In addition,
GeBP genes can control how plant epidermal cells grow (
Jing et al. 2002). In rice,
OsGeBP genes can react to various stimuli or hormones to control the downstream target genes (
Huang et al. 2021). In soybeans,
GmGeBP4 may be involved in the control of
CPR5-dependent mechanisms that result in the development of trichomes (
Liu et al. 2022). Overexpression of
MdGeBP3 demonstrated increased sensitivity to cytokinin in apples, while its ectopic expression in
Arabidopsis thaliana reduced drought resistance (
Liu et al. 2023). In moso bamboo, 12 of 16
PeGeBP's showed higher expression in tissues with trichomes compared to tissues lacking them, suggesting these genes help control trichome formation (
Shan et al. 2020). Silencing
CsGeBP4 using a new virus-induced gene silencing method in tea plants stopped trichome formation, suggesting that
CsGeBP4 is necessary for this process (
Zhou et al. 2023). Future applications of these findings include the implementation of marker-assisted selection, CRISPR/Cas9-based genome editing, and transgenic approaches to develop stress-resilient and high-yielding crop varieties.
Conclusion
The identification, characterization, and expression profiling of the GeBP gene family in B. rapa across the entire genome are described in this research. In this investigation, we discovered a GeBP gene family in B. rapa with twenty BrGeBP genes spread over six of the ten chromosomes. We performed a thorough genome-wide study of the GeBP gene family to investigate their possible involvement in anatomical formation and stress adaptation. The BrGeBP genes were also evaluated for their expected gene arrangements, preserved regions, locations on chromosomes, instances of duplication, evolutionary separation, and syntenic connections. We then categorized them using phylogenetic analysis. Additionally, we found various types of development-related and hormone-sensitive cis-acting elements in the putative BrGeBP gene promoters, demonstrating the gene family's involvement in various biological processes. Finally, based on the expression pattern of the BrGeBP genes, we hypothesized their roles. These findings lay the foundation for additional research into the biological roles of each GeBP gene in plants.
Fig. 1Phylogenetic analysis of GeBP proteins from B. rapa, B. oleracea, B. napus, Arabidopsis thaliana, and Raphanus sativus. With 1000 bootstrap replicates, the neighbor joining method was used in MEGA-7 to create the phylogenetic tree.
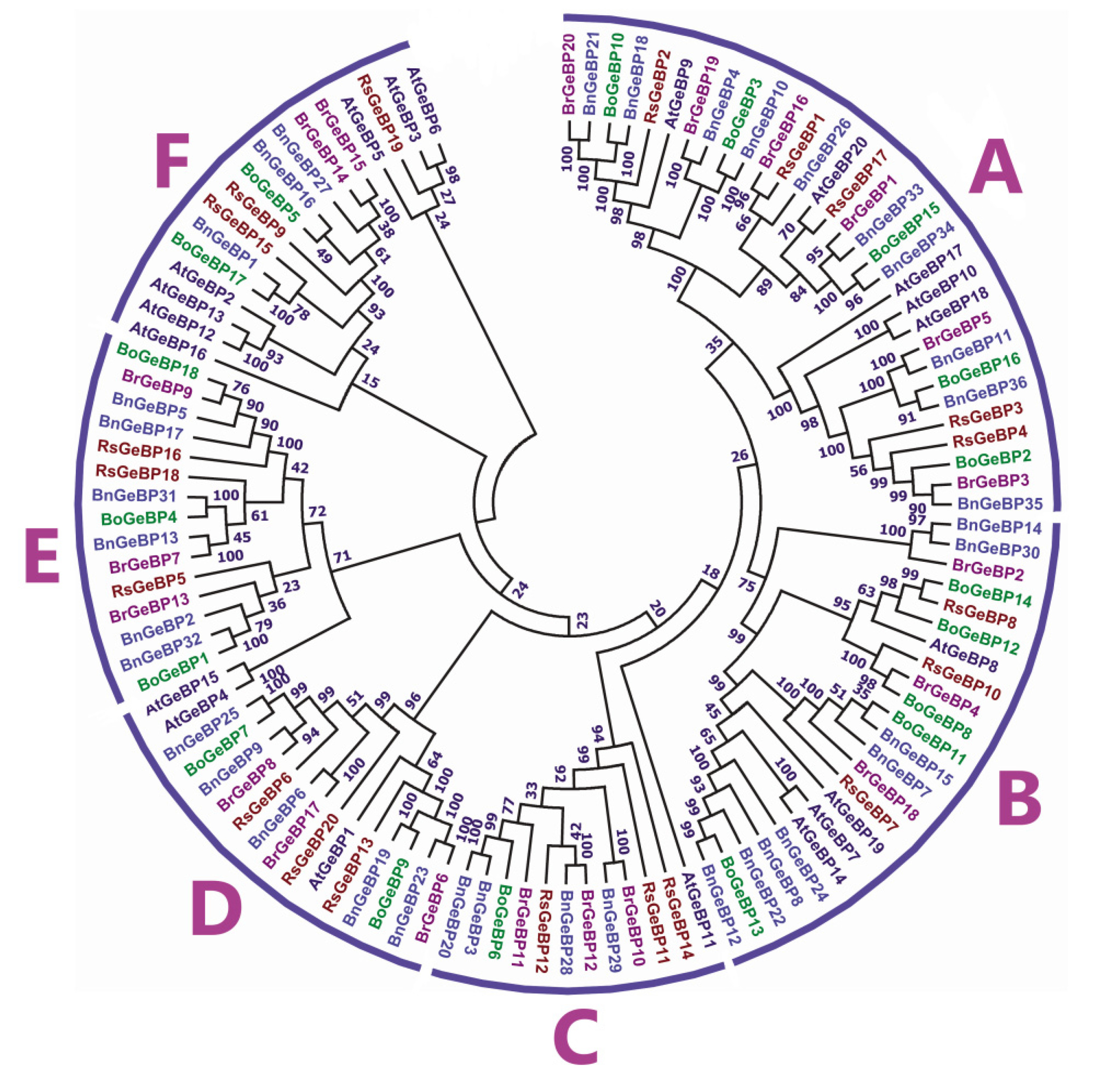
Fig. 2The syntenic relationship among Brassica rapa, Brassica oleracea, Brassica napus, and Arabidopsis thaliana is depicted using green and orange colors to represent the highest sequence similarity between the genes of these crops.
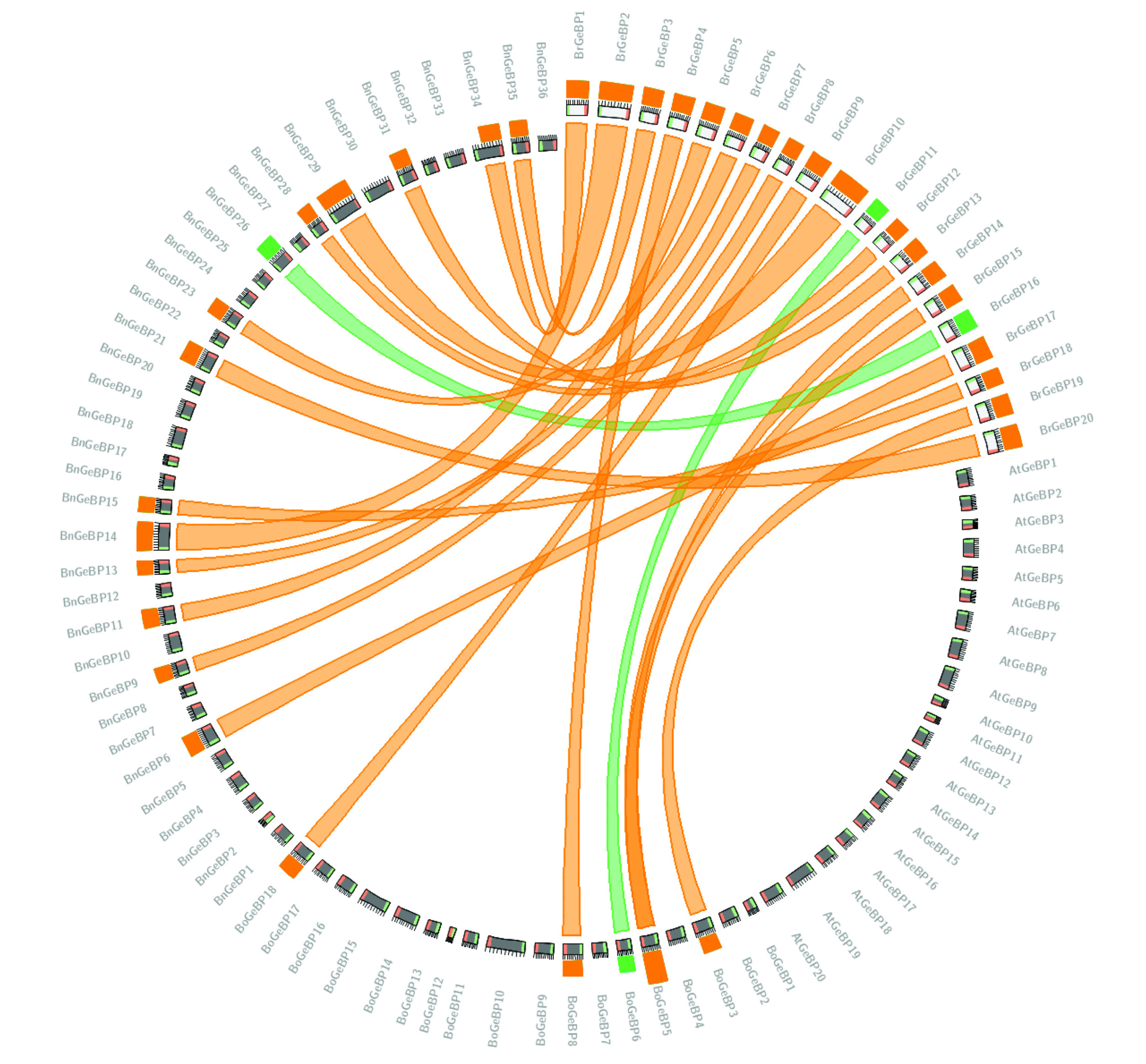
Fig. 310 conserved motifs were found with the aid of the MEME-suite tool. The different colored boxes indicate different types of motifs.
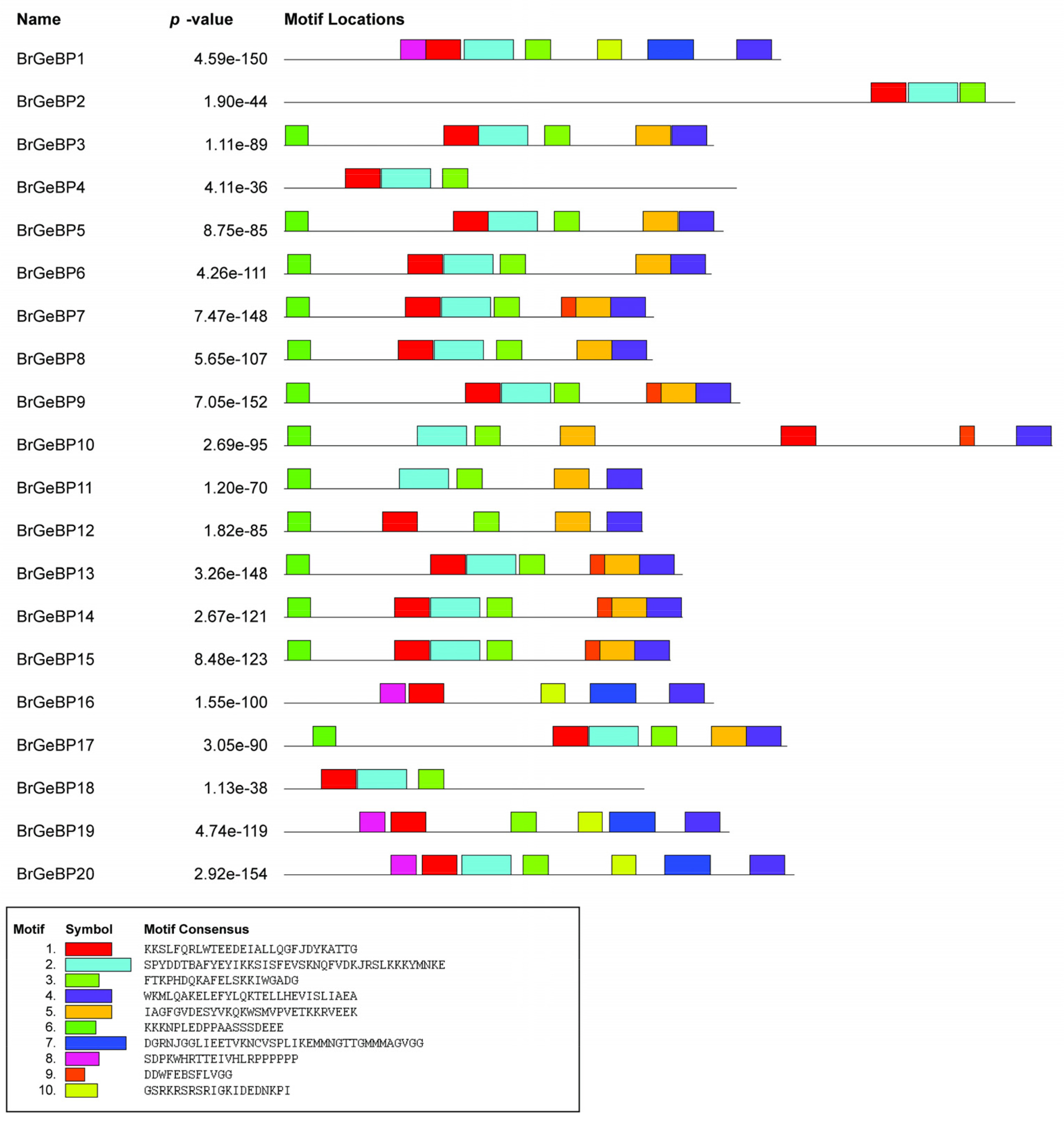
Fig. 4Exon-intron arrangement of GeBP genes was forecasted from GSDS 2.0. This structural arrangement was generated using gene and CDS sequences.
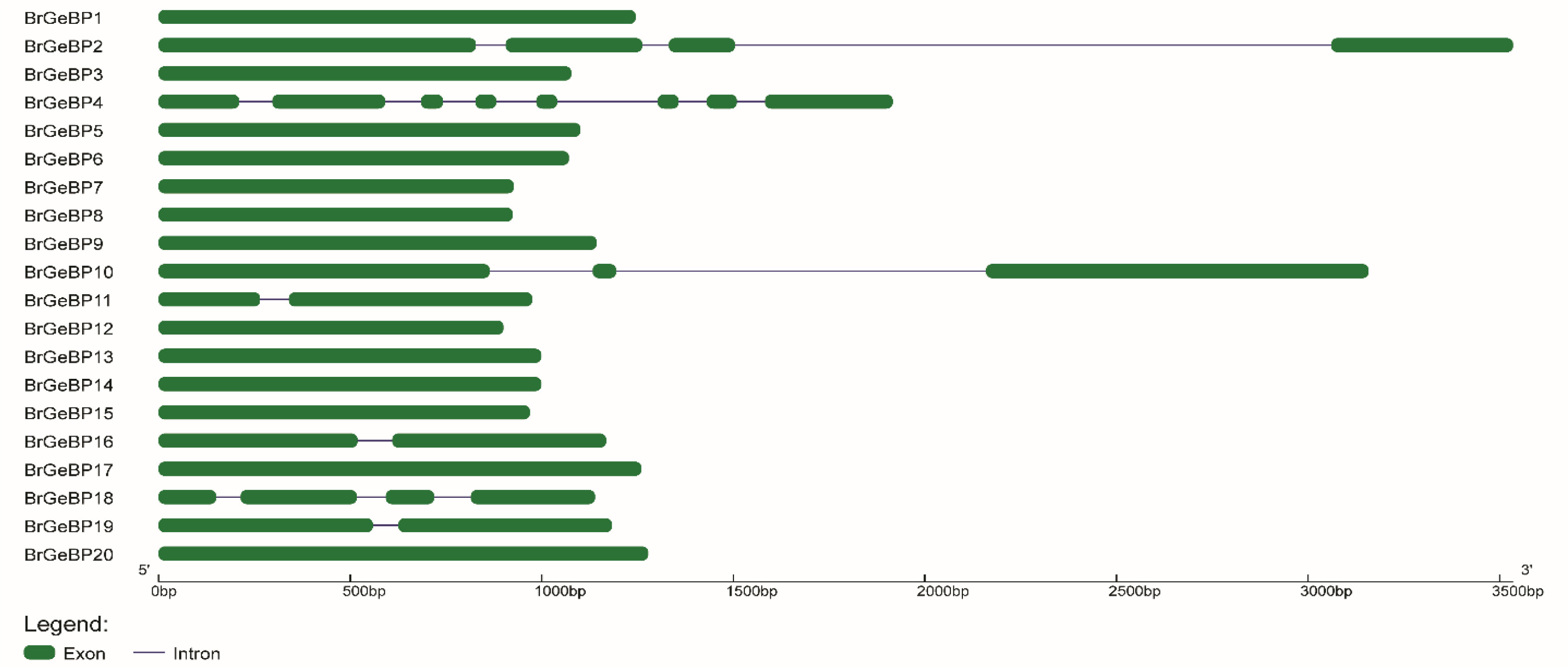
Fig. 5Localization of the GeBP gene family on the chromosome of B. rapa. The relative length of a chromosome serves as a measure of its size. Each chromosome has a number at the top that corresponds to its chromosome. Megabases (Mb) is the scale on the left side of the image that was used to estimate the sizes of the chromosomes and the locations of the genes.
Fig. 6Cis-regulatory elements were detected in the promoter region of BrGeBP genes. Numerical values indicate the number of cis-regulatory elements in each gene. These elements are non-coding DNA regions that control the neighboring genes' transcription.
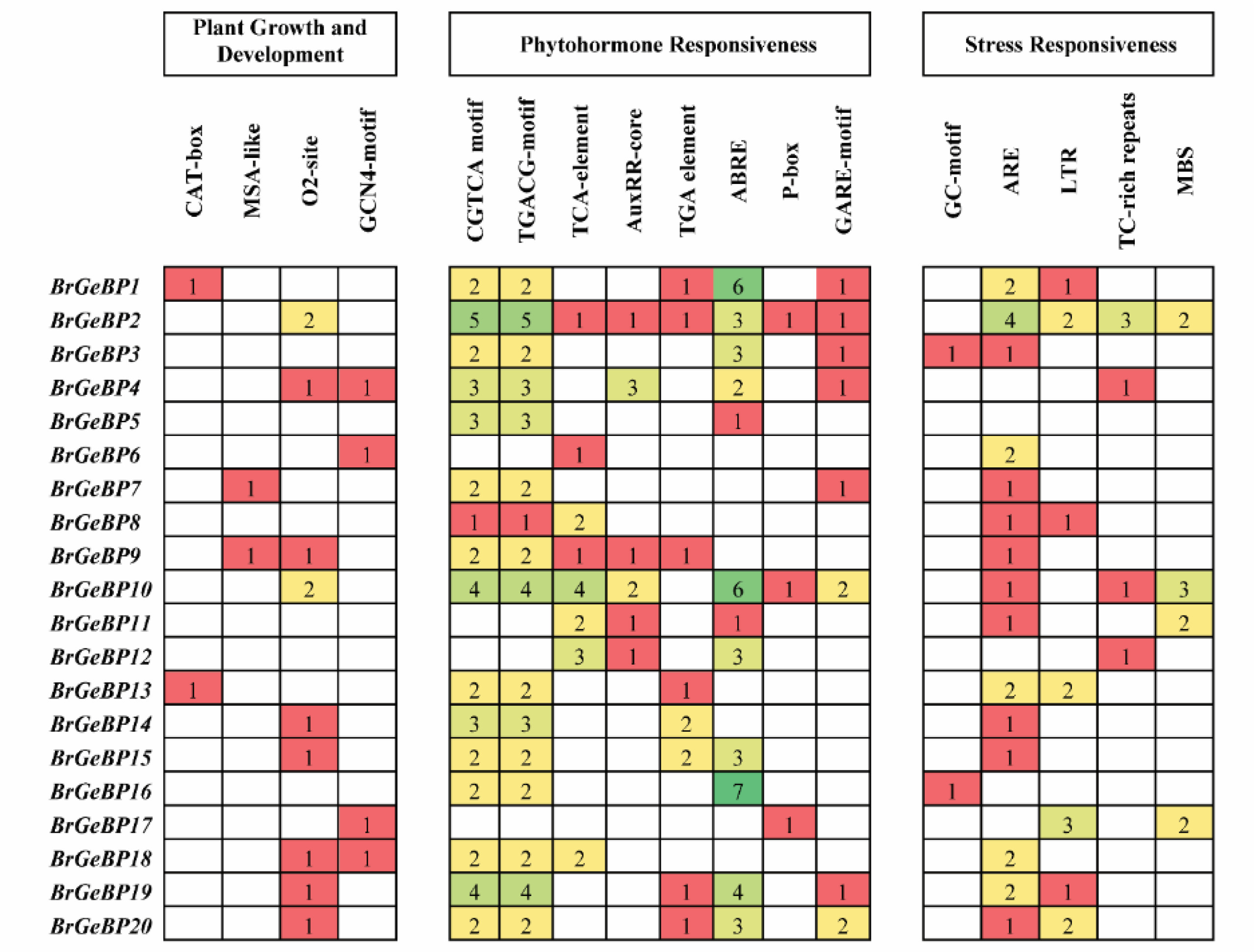
Fig. 7Expression of the BrGeBP genes in specific organs. Leaf, stem, flower, siliqua, root, and callus were present. The heat map was generated using the conditional formatting option in Microsoft Excel. The colors red, white, and green indicate high, moderate, and low levels of gene expression, respectively.
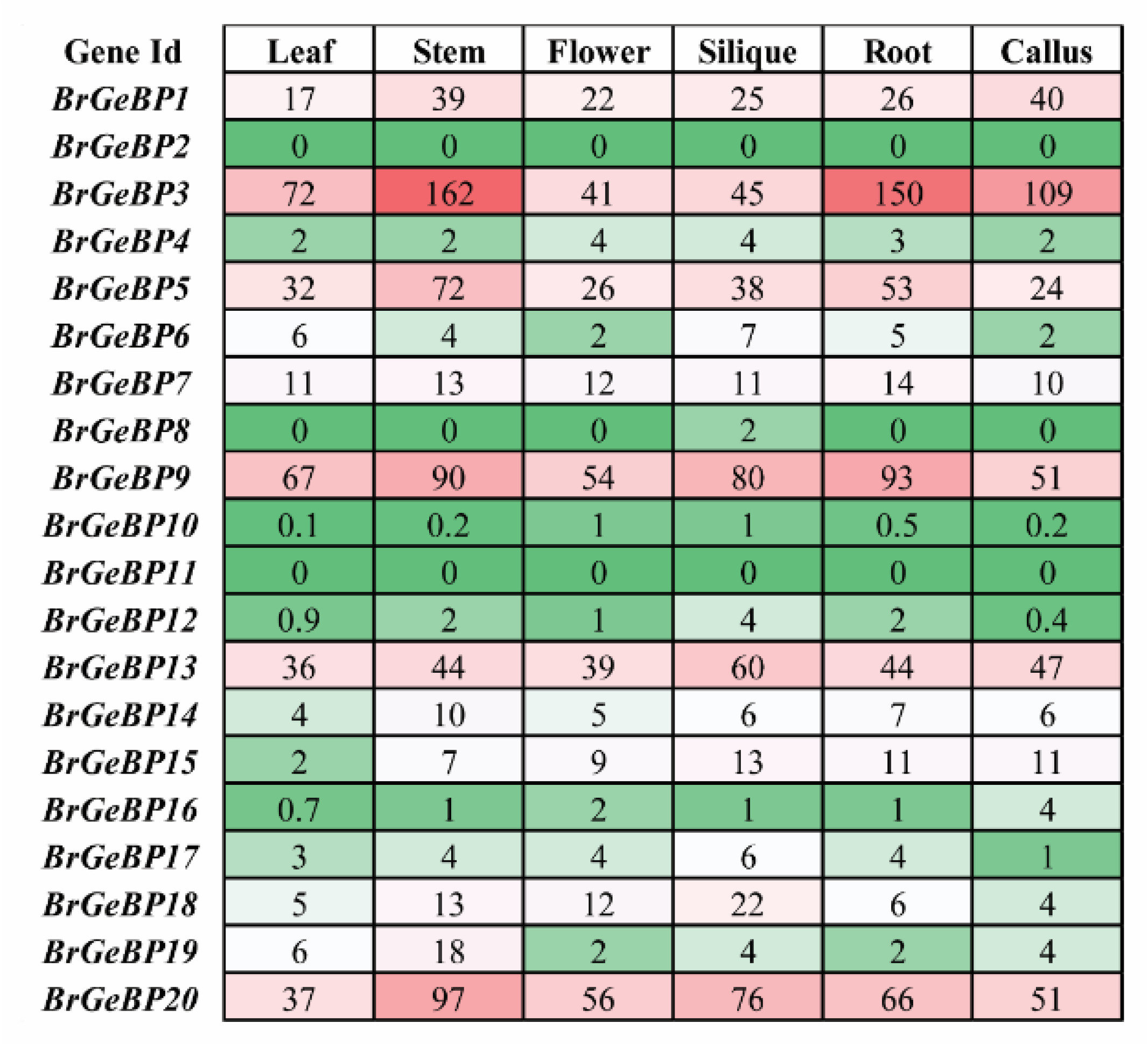
Table 1A comprehensive summary table consolidating data sources
Table 2Comprehensive details on the Brassica rapa BrGeBP genes and associated proteins
Table 2
|
Gene name |
Gene ID |
Chromosome location |
Protein
length |
Mol. wt.
(kDa) |
pI |
GRAVY |
ORF
(bp) |
Subcellular
localization |
Domain
DUF573 |
|
|
Number |
Start |
End |
Length |
|
BrGeBP1
|
Bra009986 |
A06 |
19087069 |
19088313 |
26271742 |
414 |
44.96 |
4.87 |
-0.587 |
1245 |
Nuclear |
123-219 |
|
BrGeBP2
|
Bra013842 |
A01 |
8042900 |
8046434 |
28605136 |
609 |
69.62 |
8.95 |
-0.263 |
1830 |
ER |
494-581 |
|
BrGeBP3
|
Bra013873 |
A01 |
8185027 |
8186103 |
28605136 |
358 |
39.31 |
7.53 |
-0.785 |
1077 |
Nuclear |
138-235 |
|
BrGeBP4
|
Bra017244 |
A04 |
15879572 |
15881487 |
18965343 |
377 |
43.16 |
4.56 |
-1.020 |
1134 |
Nuclear |
56-150 |
|
BrGeBP5
|
Bra019183 |
A03 |
25857483 |
25858583 |
31715688 |
366 |
40.52 |
5.72 |
-0.924 |
1101 |
Nuclear |
146-243 |
|
BrGeBP6
|
Bra019847 |
A06 |
4034905 |
4035975 |
26271742 |
356 |
39.16 |
7.66 |
-0.945 |
1071 |
Nuclear |
108-198 |
|
BrGeBP7
|
Bra021317 |
A01 |
21515533 |
92151645 |
28605136 |
308 |
34.58 |
8.89 |
-1.001 |
927 |
Nuclear |
106-193 |
|
BrGeBP8
|
Bra025445 |
A04 |
9053857 |
9054780 |
18965343 |
307 |
34.64 |
5.61 |
-0.892 |
924 |
Nuclear |
100-195 |
|
BrGeBP9
|
Bra027078 |
A09 |
8683856 |
8684998 |
18965343 |
380 |
42.10 |
5.15 |
-1.017 |
1143 |
Nuclear |
156-243 |
|
BrGeBP10
|
Bra028105 |
A04 |
6123535 |
6126692 |
18965343 |
640 |
72.81 |
8.98 |
-1.087 |
1923 |
Nuclear |
89-177,
419-508 |
|
BrGeBP11
|
Bra028106 |
A04 |
6127732 |
6128706 |
18965343 |
299 |
33.63 |
6.17 |
-0.957 |
900 |
Nuclear |
85-162 |
|
BrGeBP12
|
Bra028108 |
A04 |
6131824 |
6132723 |
18965343 |
299 |
33.91 |
5.91 |
-0.945 |
900 |
Nuclear |
87-176 |
|
BrGeBP13
|
Bra031402 |
A01 |
17604367 |
17605365 |
28605136 |
332 |
36.94 |
5.09 |
-1.055 |
999 |
Nuclear |
127-214 |
|
BrGeBP14
|
Bra032717 |
A04 |
5586257 |
5587255 |
18965343 |
332 |
36.61 |
5.31 |
-0.683 |
735 |
Nuclear |
97-187 |
|
BrGeBP15
|
Bra034488 |
Scaffold |
356960 |
357928 |
0 |
322 |
35.63 |
5.35 |
-0.668 |
969 |
Nuclear |
97-187 |
|
BrGeBP16
|
Bra036116 |
A09 |
2403139 |
2404306 |
18965343 |
358 |
38.40 |
4.69 |
-0.570 |
1077 |
Nuclear |
- |
|
BrGeBP17
|
Bra038583 |
Scaffold |
250721 |
251980 |
0 |
419 |
46.35 |
5.08 |
-1.013 |
1260 |
Nuclear |
229-324 |
|
BrGeBP18
|
Bra039500 |
A05 |
8899379 |
8900517 |
18965343 |
300 |
34.16 |
5.71 |
-0.870 |
903 |
Nuclear |
36-130 |
|
BrGeBP19
|
Bra040116 |
A01 |
25918538 |
25919720 |
28605136 |
371 |
40.83 |
5.17 |
-0.700 |
1116 |
Nuclear |
- |
|
BrGeBP20
|
Bra040416 |
Scaffold |
97200 |
98477 |
0 |
425 |
46.79 |
4.84 |
-0.638 |
1278 |
Nuclear |
- |
References
- Acarkan A, Roßberg M, Koch M, Schmidt R. 2000. Comparative genome analysis reveals extensive conservation of genome organisation for Arabidopsis thaliana and Capsella rubella. Plant. J. 23: 55-62.
- Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, et al. 2009. MEME suite: tools for motif discovery and searching. Nucleic Acids Res. 37(Suppl_2): W202-W208.
- Bohuon E, Ramsay L, Craft J, Arthur A, Marshall D, Lydiate D, et al. 1998. The association of flowering time quantitative trait loci with duplicated regions and candidate loci in Brassica oleracea. Genetics. 150: 393-401.
- Bowers JE, Chapman BA, Rong J, Paterson AHJN. 2003. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature. 422: 433-438.
- Bowling SA, Guo A, Cao H, Gordon AS, Klessig DF, Dong X. 1994. A mutation in Arabidopsis that leads to constitutive expression of systemic acquired resistance. The Plant Cell. 6: 1845-1857.
- Brininstool G, Kasili R, Simmons LA, Kirik V, Hülskamp M, Larkin JC. 2008. Constitutive Expressor Of Pathogenesis-related Genes5 affects cell wall biogenesis and trichome development. BMC Plant Biol. 8: 1-11.
- Cannon SB, Mitra A, Baumgarten A, Young ND, May G. 2004. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 4: 1-21.
- Chao JT, Kong YZ, Wang Q, Sun YH, Gong DP, Lv J, et al. 2015. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Yi chuan= Hereditas. 37: 91-97.
- Chen Y, Shen J, Zhang L, Qi H, Yang L, Wang H, et al. 2021. Nuclear translocation of OsMFT1 that is impeded by OsFTIP1 promotes drought tolerance in rice. Mol Plant. 14: 1297-1311.
- Cheng F, Liu S, Wu J, Fang L, Sun S, Liu B, et al. 2011. BRAD, the genetics and genomics database for Brassica plants. BMC Plant Biol. 11: 1-6.
- Chou KC, Shen HB. 2010. Cell-PLoc 2.0: an improved package of web-servers for predicting subcellular localization of proteins in various organisms. Nat. Sci. 2: 1090
- Clemens S. 2006. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie. 88: 1707-1719.
- Curaba J, Herzog M, Vachon G. 2003. GeBP, the first member of a new gene family in Arabidopsis, encodes a nuclear protein with DNA-binding activity and is regulated by KNAT1. Plant J. 33: 305-317.
- Curaba J, Moritz T, Blervaque R, Parcy F, Raz V, Herzog M, et al. 2004. AtGA3ox2, a key gene responsible for bioactive gibberellin biosynthesis, is regulated during embryogenesis by LEAFY COTYLEDON2 and FUSCA3 in Arabidopsis. Plant Physiol. 136: 3660-3669.
- Fang Y, You J, Xie K, Xie W, Xiong L. 2008. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genomics. 280: 547-563.
- Gabaldón T, Koonin EV. 2013. Functional and evolutionary implications of gene orthology. Nat. Rev. Genet. 14: 360-366.
- Gan Y, Liu C, Yu H, Broun P. 2007. Integration of cytokinin and gibberellin signalling by Arabidopsis transcription factors GIS, ZFP8 and GIS2 in the regulation of epidermal cell fate. Development. 134: 2073-2081.
- García-Cano E, Hak H, Magori S, Lazarowitz SG, Citovsky V. 2018. The Agrobacterium F-box protein effector VirF destabilizes the Arabidopsis GLABROUS1 enhancer/binding protein-like transcription factor VFP4, a transcriptional activator of defense response genes. MPMI. 31: 576-586.
- Gasteiger E, Hoogland C, Gattiker A, Duvaud Se, Wilkins MR, Appel RD, et al. 2005. Protein identification and analysis tools on the ExPASy server. The proteomics protocols handbook. 571-607..
- Hu B, Jin J, Guo AY, Zhang H, Luo J, Gao G. 2015. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics. 31: 1296-1297.
- Hu TT, Pattyn P, Bakker EG, Cao J, Cheng J-F, Clark RM, et al. 2011. The Arabidopsis lyrata genome sequence and the basis of rapid genome size change. Nat. Genet. 43: 476-481.
- Huang J, Zhang Q, He Y, Liu W, Xu Y, Liu K, et al. 2021. Genome-wide identification, expansion mechanism and expression profiling analysis of GLABROUS1 enhancer-binding protein (GeBP) gene family in Gramineae crops. Int. J. Mol. Sci. 22: 8758
- Jakoby M, Weisshaar B, Dröge-Laser W, Vicente-Carbajosa J, Tiedemann J, Kroj T, et al. 2002. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 7: 106-111.
- Jasinski S, Piazza P, Craft J, Hay A, Woolley L, Rieu I, et al. 2005. KNOX action in Arabidopsis is mediated by coordinate regulation of cytokinin and gibberellin activities. Curr. Bioly. 15: 1560-1565.
- Jing HC, Sturre MJ, Hille J, Dijkwel PP. 2002. Arabidopsis onset of leaf death mutants identify a regulatory pathway controlling leaf senescence. Plant J. 32: 51-63.
- Johnston JS, Pepper AE, Hall AE, Chen ZJ, Hodnett G, Drabek J, et al. 2005. Evolution of genome size in Brassicaceae. Ann. Bot. 95: 229-235.
- Kersey PJ, Allen JE, Armean I, Boddu S, Bolt BJ, Carvalho-Silva D, et al. 2016. Ensembl Genomes 2016: More genomes, more complexity. Nucleic Acids Res. 44(D1): D574-D580.
- Khare D, Mitsuda N, Lee S, Song WY, Hwang D, Ohme-Takagi M, et al. 2017. Root avoidance of toxic metals requires the GeBP-LIKE 4 transcription factor in Arabidopsis thaliana. New Phytol. 213: 1257-1273.
- Kong X, Lv W, Jiang S, Zhang D, Cai G, Pan J, et al. 2013. Genome-wide identification and expression analysis of calcium-dependent protein kinase in maize. BMC Genomics. 14: 1-15.
- Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, et al. 2002. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 30: 325-327.
- Letunic I, Bork P. 2018. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 46(D1): D493-D496.
- Li Z, Jiang H, Zhou L, Deng L, Lin Y, Peng X, et al. 2014. Molecular evolution of the HD-ZIP I gene family in legume genomes. Gene. 533: 218-228.
- Liu RX, Li HL, Qiao ZW, Liu HF, Zhao LL, Wang XF, et al. 2023. Genome-wide analysis of MdGeBP family and functional identification of MdGeBP3 in Malus domestica. Environ. Exp. Bot. 208: 105262
- Liu S, Liu Y, Liu C, Zhang F, Wei J, Li B. 2022. Genome-wide characterization and expression analysis of GeBP family genes in soybean. Plants. 11: 1848
- Liu S, Liu Y, Yang X, Tong C, Edwards D, Parkin IA, et al. 2014. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 5: 3930.
- Lukens L, Zou F, Lydiate D, Parkin I, Osborn TJG. 2003. Comparison of a Brassica oleracea genetic map with the genome of Arabidopsis thaliana. Genetics. 164: 359-372.
- Ma C, Chen Q, Wang S, Lers A. 2021. Downregulation of GeBP-like α factor by MiR827 suggests their involvement in senescence and phosphate homeostasis. BMC Biol. 19: 90
- Nagaharu U, Nagaharu NJJJB. 1935. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 7: 389-452.
- Oppenheimer DG, Herman PL, Sivakumaran S, Esch J, Marks MD. 1991. A myb gene required for leaf trichome differentiation in Arabidopsis is expressed in stipules. Cell. 67: 483-493.
- Perazza D, Laporte F, Balagué C, Chevalier F, Remo S, Bourge M, et al. 2011. GeBP/GPL transcription factors regulate a subset of CPR5-dependent processes. Plant Physiol. 157: 1232-1242.
- Perazza D, Vachon G, Herzog M. 1998. Gibberellins promote trichome formation by up-regulating GLABROUS1 in Arabidopsis. Plant Physiol. 117: 375-383.
- Petryszak R, Keays M, Tang YA, Fonseca NA, Barrera E, Burdett T, et al. 2016. Expression Atlas update-an integrated database of gene and protein expression in humans, animals and plants. Nucleic Acids Res. 44(D1): D746-D752.
- Pickett FB, Meeks-Wagner DR. 1995. Seeing double: appreciating genetic redundancy. Plant. Cell. 7: 1347
- Potter SC, Luciani A, Eddy SR, Park Y, Lopez R, Finn RD. 2018. HMMER web server: 2018 update. Nucleic Acids Res. 46(W1): W200-W204.
- Saeediazar S, Zarrini HN, Ranjbar G, Heidari P. 2014. Identification and study of cis regulatory elements and phylogenetic relationship of TaSRG and other salt response genes. JBES. 5: 1-5.
- Saitou N, Nei M. 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4: 406-425.
- Samad AF, Sajad M, Nazaruddin N, Fauzi IA, Murad AM, Zainal Z, et al. 2017. MicroRNA and transcription factor: key players in plant regulatory network. Front. Plant Sci. 8: 565
- Shan X, Yang K, Shi J, Zhu C, Gao Z. 2020. Genome-wide identification and expression analysis of GeBP transcription factor gene family in moso bamboo. J. Nanjing For. Univ. 44: 41.
- Song XM, Huang ZN, Duan WK, Ren J, Liu TK, Li Y, et al. 2014. Genome-wide analysis of the bHLH transcription factor family in Chinese cabbage (Brassica rapa ssp. pekinensis). Mol. Genet. Genomic. 289: 77-91.
- Tong C, Wang X, Yu J, Wu J, Li W, Huang J, et al. 2013. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genomics. 14: 1-10.
- Wang R, Wu X, Wang Z, Zhang X, Chen L, Duan Q, et al. 2023. Genome-wide identification and expression analysis of BrGeBP genes reveal their potential roles in cold and drought stress tolerance in Brassica rapa. Int. J. Mol. Sci. 24: 13597
- Wang XiaoWu WX, Wang HanZhong WH, Wang Jun WJ, Sun RiFei SR, Wu Jian WJ, Liu ShengYi LS, et al. 2011. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 43: 1035-1039.
- Wang Y, Feng L, Zhu Y, Li Y, Yan H, Xiang Y. 2015. Comparative genomic analysis of the WRKY III gene family in populus, grape, Arabidopsis and rice. Biol. Direct. 10: 1-27.
- Wolters H, Jürgens G. 2009. Survival of the flexible: hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 10: 305-317.
- Wu J, Peng Z, Liu S, He Y, Cheng L, Kong F, et al. 2012. Genome-wide analysis of Aux/IAA gene family in Solanaceae species using tomato as a model. Mol. Genet. Genomics. 287: 295-311.
- Yanai O, Shani E, Dolezal K, Tarkowski P, Sablowski R, Sandberg G, et al. 2005. Arabidopsis KNOXI proteins activate cytokinin biosynthesis. Curr. Biol. 15: 1566-1571.
- Zhang W, Ruan J, Ho T-hD, You Y, Yu T, Quatrano RS. 2005. Cis-regulatory element based targeted gene finding: genome-wide identification of abscisic acid-and abiotic stress-responsive genes in Arabidopsis thaliana. Bioinformatics. 21: 3074-3081.
- Zhang Z, Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25: 3389-402.
- Zheng Y, Jiao C, Sun H, Rosli HG, Pombo MA, Zhang P, et al. 2016. iTAK: a program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol. Plant. 9: 1667-1670.
- Zhou H, Zhou W, Yao X, Zhao Q, Lu L. 2023. Genome--wide investigation and functional analysis reveal that CsGeBP4 is required for tea plant trichome formation. Int. J. Mol. Sci. 24: 5207
- Zhu Y, Wu N, Song W, Yin G, Qin Y, Yan Y, et al. 2014. Soybean (Glycine max) expansin gene superfamily origins: segmental and tandem duplication events followed by divergent selection among subfamilies. BMC Plant Biol. 14: 1-19.