Abstract
The storage proteins in wheat, particularly the high molecular weight glutenin subunits (HMW-GS), play crucial roles in the processing of flour and the quality of bread made from common wheat. These subunits are encoded by the Glu-A1, Glu-B1, and Glu-D1 loci. This study aimed to determine the composition of high molecular weight glutenin subunits in 89 Mongolian cultivars of spring wheat using KASP markers and sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The result showed the presence of the 5+10 allele at the Glu-D1 locus in Mongolian-developed varieties was confirmed through SDS-PAGE, providing evidence for the synthesis of high molecular weight glutenin subunits. The uniform presence of high molecular weight glutenin subunit alleles across the tested cultivars contributes valuable insights into the bread-making quality of Mongolian wheat varieties.
-
Key words: bread wheat, Glu-1 gene, high molecular weight glutenin subunit (HMW-GS), KASP molecular markers, SDS-PAGE
Introduction
The wheat (
T. aestivum L.) storage proteins (gliadin, glutenin) play a main role in the viscoelastic properties of bread dough (
Turner et al. 2004). The composition and amount of high molecular weight glutenin subunits in common wheat will determine the end-use quality, especially bread-making quality. Following molecular weight and amino acid sequences, the classification of gluten proteins involves two groups: those characterized by low molecular weight (10-70 kDa) and high molecular weight (80-130 kDa) (
Bietz et al. 1972). The most commonly used technique for analyzing high molecular glutenin subunits is SDS-PAGE, its reproducibility, cost-effective, rapid, and minimal sample requirement (
Bietz et al. 1975). Numerous genetic studies investigate wheat storage protein which makes up gluten, and have examined its functionality (
Shewry et al. 1997). However, comprehending the genetic basis of protein quantity is challenging. The regulation of grain protein involves a complex. The genetic system is further complicated by significant environmental factors, making genetic analysis notably difficult. HMW-GS are encoded by three loci (
Glu-A1, Glu-B1, Glu-D1) located in chromosomes 1A, 1B and 1D, respectively(
Payne 1987;
Sameri et al. 2008). Different reports indicate that subunits '5+10' (
Glu-D1) are reported to have a significant effect on dough strength by producing a greater proportion of large-sized polymer than subunits '2+12' (
Glu-D1) (
Buonocore et al. 1996;
Payne et al. 1987). In investigating wheat varieties across diverse regions, including Great Britain, Canada, Spain, Australia, Germany, and France discernible patterns have emerged. They identified certain subunits as either "good" or "bad". To measure the genetic impact on bakery quality related to high molecular weight glutenin subunits (HMW-GS), they introduced a scoring system Glu-1 quality score for common wheat varieties (
Payne et al. 1983).
Mongolian wheat cultivars are a significant crop for Mongolia's agricultural sector, but their genetic diversity and molecular characterization are not well understood. Improving yield and adaptability to changing environmental conditions requires knowledge of their genetic diversity. Previous studies have focused on their morphological and phenotypic characterization, but there is a lack of molecular data on their genetic diversity. This study aimed to address this knowledge gap by conducting a high molecular weight study of Mongolian wheat cultivars using KASP markers. The core objective of our research work was to inspect the glutenin subunits by KASP marker to detect Glu-1 gene alleles compared with the determination of subunits by SDS-PAGE polyacrylamide electrophoresis.
Materials and Methods
Wheat materials
The wheat material used included 10 spring soft wheat (T. aestivum L.) cultivars (Khalh-gol-1, Dakhan-131, Dakhan-172, Darkhan-34, Darkhan-74, Darkhan-166, Darkhan-193, Darkhan-144, Darkhan-181, Darkhan-212) which were chosen from the wheat breeding program at the Institute of Plant and Agricultural Sciences of Mongolia, 78 germplasm, which are Mongolian local wheat cultivars were retrieved from GenBank (Mongolian National Plant GenBank), and AGS2000 variety of USA.
Wheat cultivation growth chamber experimental design
Growth chamber experiments were conducted at the University of Georgia, Griffin Campus (33.26445975342215° N, -84.28409533175976° W) in the USA. 3 Seeds of each cultivar were planted in cones, which were in cone-tainer trays (Stuewe and Sons, Inc., Tangent, OR). Each cone-trainer tray was 30.48 cm×609.6 cm×17.15 cm and held 98 cones, with each cone being 3.81 cm in diameter at the largest point and approximately height is 10.16 cm. The cones were filled with Pro-mix growing mix (Pro-Mix Gardening, Quakertown, PA) and subjected to 14 hours days, 10 hours nights, and 18±3℃ in the greenhouse (
Zhao et al. 2020).
Five KASP markers already developed for
Glu-1 gene were selected from published article including Glu-Ax1/ x2*_SNP, Glu-Ax2_IND for
Glu-A1 gene, BX7OE_866_SNPBx13_SNP for
Glu-B1 gene,
Glu-D1d_SNP for
Glu-D1 gene (
Rasheed et al. 2016). Primers were ordered from Louisville city of USA of Eurofins Genomics LLC. DNA was extracted using a modified cetyltrimethylammonium bromide (CTAB) protocol (
Saghai-Maruuf et al. 1984) and then diluted to 20 ng/μL. Assays were tested in 96 well formats and set up as KASP master mixture 5.137 μL and 4.9 μL DNA. The following PCR program was used for KASP marker validation: 30℃ for 1 minute during the pre-read stage; 95℃ for 10 minutes during the preheating stage, followed by a touchdown program of 10 denaturation cycles at 95℃ for 20 seconds; annealing/extension at 61℃ for 1 minute with a 0.6-degree reduction every cycle; plus 30 cycles of 95℃ for 29 seconds and 56℃ for 1 minute. Temperature was reduced to 30℃ for 1 minute and 30 seconds during the post-read stage. Fluorescent signals were collected at the pre-read and post-read stages at 30°C. Real-time PCR was analyzed by using QuantStudio™ 3 Real-Time PCR System, 96-well, 0.2 mL (Thermo Fisher Scientific Inc. United states).
Forty grams of grains from each cultivar were milled by Brabander Quadrumat Junior laboratory miller to obtain a fine powder and used to analyze the composition of HMW-GS.
HMW-GS were extracted from wheat grains by using a modified method according to
Singh et al. (1991). Fine powder (30 mg) was extracted with 1 ml of 55% (v/v) propane-2-ol on an Eppendorf tube for 20 minutes with continuous mixing at 65℃, followed by centrifugation for 5 minutes at 10000 rpm. This step was repeated three times to remove the gliadin component. A pellet containing glutenin subunits fraction was resuspended in 0.1 mL a solution of 55% (v/v) propan-2-ol, 0.08 M tris (hydroxymethyl) aminomethane hydrochloric acid (Tris-HCl) pH 8.3 and 1%(w/v) dithiothreitol (DTT) as reducing agent and incubated for 30 minutes at 60°C with continuous mixing. After centrifugation for 5 minutes at 14000 rpm, supernatant with both HMW-GS and LMW-GS fractions, was transferred into a new Eppendorf tube. Acetone was then added up to 40% (v/v) to precipitate HMW-GS. The extracted fractions were then dried up at 40℃ and used for SDS-PAGE analyses.
SDS-PAGE was performed on Mini PROTEAN Tetra Cell (Bio-Rad, Hercules,CA) with 10% acrylamide gel (for HMW-GS). TGX™ FastCast™ Acrylamide starter Kit, 10% was ordered from Bio Rad LLC. Dried HMW-GS were suspended in 20 μL of double distilled water, then add 5 μL loading buffer containing 2% (w/v) SDS, 0.02% (w/v) bromophenol blue, 0.1% DTT, 0.05 M Tris-HCl pH 6.8 and 10% (v/v) glycerol. After boiling at 100°C for 5 minutes, a 10 μL sample was loaded into the gel. PageRuler™ Unstained Protein Ladder Marker (product #26630) range from 10 to 180 kDa was used for tracing HMW-GS. Electrophoresis separation was carried out at 40 mA. Gels were stained with Brilliant blue, fixed in 10% (v/v) acetic acid and 30% (v/v) methanol, and destained in 30% (v/v) methanol and 10% (v/v) acetic acid.
Data analysis
Microsoft Excel was used to calculate the allelic frequencies for each locus and for each subcollection, as well as to assess the genetic diversity (
He) and the effective number of alleles (
Ne) using the following formulas from Nei (
Masatoshi 1972).
Results
Result of genotyping
High molecular weight glutenin subunits are a group of storage proteins which are deposited in the endosperm of wheat during grain filling. The gene that controls their synthesis is located at three complex loci, one each on the long arms of chromosome
Glu-A1,
Glu-B1, and
Glu-D1(
Payne et al. 1983). HMW glutenin subunits at Ax1 and Ax2 at Glu-A1, Bx7OE at
Glu-B1, and 5+10 and 2+12 at
Glu-D1 locus by KASP markers, respectively were identified in 89 cultivars used in the study and are shown in
Table 1. The Ax1* subunit significantly enhances the rheological and physicochemical properties of dough, contributing to improved connectivity and uniformity of gluten microstructure, as well as increasing the content of disulfide bonds and foaming properties (
Junwei Zang 2023). At the locus, the most frequent subunit was Ax1* allele which is expressed in 44% of accessions, followed by subunit Ax2* in 29.0%. Among the analyzed cultivars, eight spring soft wheat varieties- Khalh-gol-1, Dakhan-131, Dakhan-172, Darkhan-34, Darkhan-193, Darkhan-144, Darkhan-181, Darkhan-212 showed an 80% presence of the Ax2* allele at the
Glu-A1 locus within Mongolian-developed wheat. In contrast, an analysis of 78 Mongolian local wheat cultivars revealed that the Ax2* allele occurred in only 22% of accessions, whereas the Ax1* allele was present in 49%. This genetic divergence highlights the predominance of the Ax1* allele in local varieties. Allele frequency differences indicate that selective breeding has a preferred Ax2* allele in cultivated varieties, enhancing agronomic traits. Conversely, the greater prevalence of the Ax1* allele in local cultivars may reflect adaptations to local environments.
Branlard et al (2001) found that the Ax2* allele has an identical impact on the strength and extensibility of dough compared to the Ax1* allele. This suggests that Ax2* performs at least as well as other beneficial alleles (
Branlard 2001).
The Bx7OE allele at the
Glu-B1 locus enhances dough strength and mixing properties through increased gluten protein expression. While it significantly improves dough strength under high nitrogen conditions, it does not notable affect loaf volume compared to lines without the allele (
Trittinger 2019). The Bx7OE allele, identified as an x-type subunit for the
Glu-B1 locus, had an allelic frequency of 7%. The Bx7OE allele was not identified in any of the Mongolian-developed varieties but was present in the local germplasm, specifically in varieties MK-307, MK-336, MK-354, MK-358, MK-916, and MK-1568. To improve the genetic quality of the developed varieties, it is essential to utilize these local varieties as breeding materials. Utilizing such genetic resources can enhance allelic diversity and potentially introduce desirable quality traits for future Mongolian wheat breeding programs.
At the
Glu-D1 locus, alleles 5+10 and 2+12 were identified with frequencies of 13% and 75%, respectively. The 5+10 allele, known for its positive contribution to dough strength, was predominant found in Mongolian-developed cultivars including Khalh-gol-1, Dakhan-131, Dakhan-172, Darkhan-34, Darkhan-74, Darkhan-166, Darkhan-193, Darkhan-144, Darkhan-181, and Darkhan-212. Additionally, it was present in other germplasms such as MK-287, Buritskay 34, NONAME, AGS2000, highlighting their potential value in breeding programs aimed at enhancing gluten strength and overall wheat quality (
Table 1). The study highlights the importance of integrating both local and developed wheat varieties in breeding programs. Utilizing diverse alleles can enhance dough quality and strength, improving the genetic quality of wheat and its adaptability to different growing conditions.
Table 2 shows genetic diversity at
Glu-A1 was 0.63 (
He). The Bx7OE allele, which x type subunit was identified for the
Glu-B1 locus, and the allelic frequency was 7%. At the
Glu-D1 locus, 5+10 and 2+12 alleles were identified, frequency was 13% and 73%, respectively. Nei index of genetic diversity at
Glu-D1 locus was 0.57 (
Table 2). The data reveal moderate genetic diversity in the wheat population, with the 2+12 allele at the
Glu-D1 locus being notably more prevalent.
Result of SDS-PAGE
Gluten proteins are categorized into two groups based on molecular weight and amino acid sequences: low molecular weight (10-70kDa) and high molecular weight (80-130 kDa) (
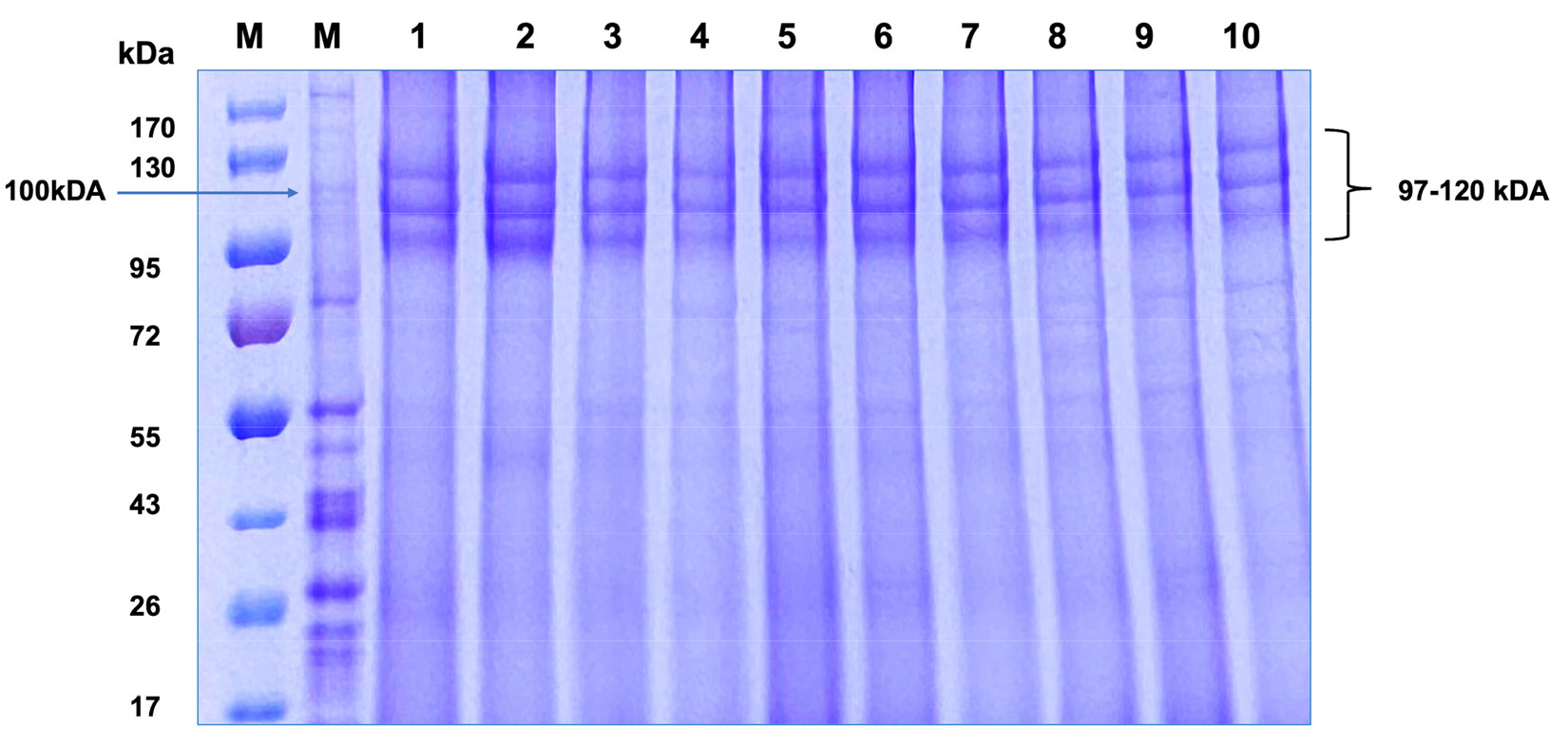
Bietz et al. 1972). The SDS-PAGE electrophoresis conducted on Mongolian-developed wheat cultivars (Khalh-gol-1, Dakhan-131, Dakhan-172, Darkhan-34, Darkhan-74, Darkhan-166, Darkhan-193, Darkhan-144, Darkhan-181 and Darkhan-212), guided by fundamental research in wheat genetics and protein analysis, determined distinct protein bands within the critical 97 to 120 kDa range (
Fig. 1). This result showed HMW-GS typical of the allelic configuration 5+10 at
Glu-D1 locus as compared with published data (
Sissons 2008). Multiple reports indicate that 5+10 (
Glu-D1) significantly enhances dough strength by producing a higher proportion of large-sized polymers compared to subunit 2+12(
Glu-D1) (
Buonocore et al. 1996;
Payne et al. 1987).
Discussion
Payne et al. revealed a correlation between gluten strength and the presence of certain HMW-GS measured by the SDS-sedimentation volume test. Accordingly, they designed a numeric scale to evaluate bread-making quality as a function of the described subunits (
Payne et al. 1987). It was determined that the allelic variation at the
Glu-D1 locus has a stronger impact on bread-making than the variation at the other
Glu-1 loci. In numerous studies, researchers have investigated the composition of High Molecular Weight Glutenin Subunits (HMW-GS) in collections of wheat cultivars from various countries (
Payne et al. 1983;
Shewry et al. 1997;
Singh et al. 1991;
Sissons 2008;
Turner et al. 2004).
The result of current research, High Molecular Weight Glutenin Subunits (HMW-GS) were studied in details. Mongolian10 cultivars (Khalh-gol-1, Dakhan-131, Dakhan-172, Darkhan-34, Darkhan-74, Darkhan-166, Darkhan-193, Darkhan-144, Darkhan-181, Darkhan-212), which developed the breeding program at the Institute of Plant and Agricultural Sciences of Mongolia showed Ax2* allele for Glu-A1 locus, null allele for Glu-B1 locus, and 5+10 for Glu-D1 locus indicating that all varieties have similar bread making quality alleles at HMW-GS loci. The Mongolian local cultivars, which 70 germplasms Ax2* and Ax1 alleles for Glu-A1 locus, Bx7OE allele for Glu-B1 locus, and 2+12 allele for Glu-D1 locus indicating that all lines have similar bread making quality alleles at HMW-GS loci.
The quality of wheat-derived dough is positively influenced by both Ax-1 and Ax2 alleles at the Glu-A1 locus, while Ax-null alleles have a detrimental impact on dough quality, as reported by Lukow O.M. in 1989.
In comparing Mongolian-developed varieties and Mongolian local cultivars a shared presence of beneficial HMW-GS alleles is evident. Specifically, the Ax2* allele at
Glu-A1 locus is common to both groups, suggesting a potential contribution to favorable dough quality. Moreover, the emphasis on specific alleles, such as Bx7OE for
Glu-B1 and 2+12 for
Glu-D1, in Mongolian local cultivars underscores a collective effort aimed at preserving or improving bread-making quality. Mongolian-developed wheat varieties exhibit enhanced bread-making quality due to the consist 5+10 allele at the
Glu-D1 locus, known for its favorable impact on dough quality (
Payne et al. 1987).
Genetic variation analysis across the
Glu-1 loci, encompassing
Glu-A1, Glu-B1 and
Glu-D1, revealed distinctive patterns in different studies. The present study demonstrated values of 0.63, 093, and 0.27 for these loci, resulting in average genetic variation was 0.71. This aligns with findings from a study on native wheat populations in the Zanjan Region, which reported values of 0.278, 0.8 and 0.586 for
Glu-A1, Glu-B1 and
Glu-D1, with an average genetic variation of 0.844 (
Sameri et al. 2008). Additionally, a separate investigation yielded values of 0.414, 0.532, and 0.648 for
Glu-A1, Glu-B1 and
Glu-D1, contributing to an average genetic variation of 0.536 (
Shiri et al. 2015). These diverse outcomes underscore the importance of considering regional variations and research methodologies in understanding the genetic landscape of glutenins in the wheat population.
Conclusion
Four KASP markers already developed we utilized for detecting high molecular weight glutenin genes encoded at the Glu-A1, Glu-B1, and Glu-D1 loci. These four KASP markers worked well with the DNA extracted using high throughput protocol, and Mongolian10 breeding populations, germplasm 78 lines and AGS2000 variety of USA were successfully genotyped with Institute of Plant Breeding, Genetics and Genomics, University of Georgia, Griffin Campus, Griffin, GA, United States. Therefore, these markers can be applied in the breeding program at the Institute of Plant and Agricultural Sciences of Mongolia.
Acknowledgments
This study was supported by Institute of Plant Breeding, Genetics and Genomics, University of Georgia, Griffin Campus, Griffin, GA, United States.
Fig. 1SDS-PAGE analysis of high molecular weight glutenin subunits (HMW-GS) in Mongolian-developed wheat varieties. The varieties analyzed are slots M. molecular weight marker, 1. Khalh-gol-1, 2. Dakhan-131, 3. Dakhan-172, 4. Darkhan-34, 5. Darkhan-74, 6. Darkhan-166, 7. Darkhan-193, 8. Darkhan-144, 9. Darkhan-181, 10. Darkhan-212.
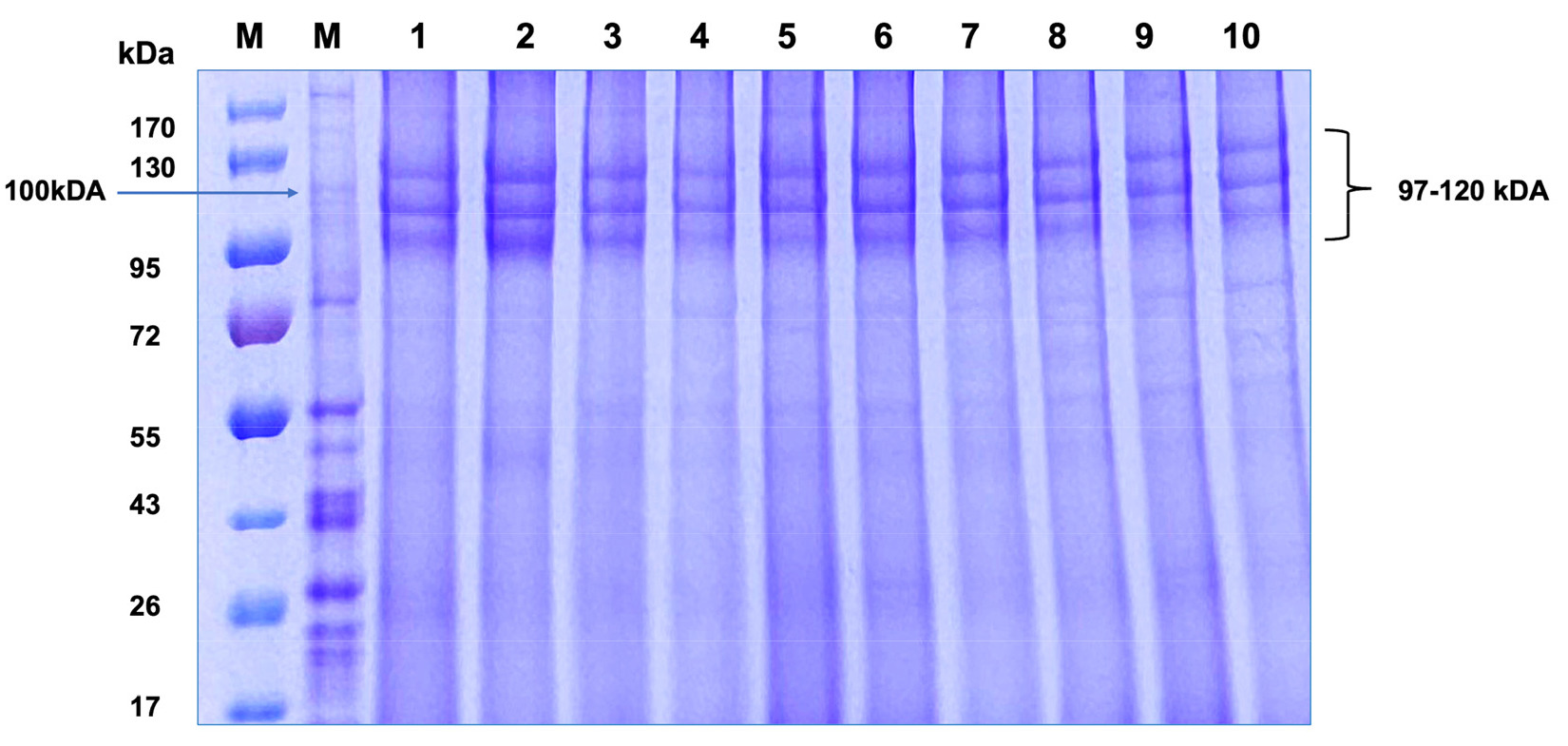
Table 1HMW-GS composition of Mongolian wheat cultivars.
Table 1
|
No |
Species |
Cultivar |
HMW-GS |
|
Alleles |
|
|
|
Glu-A1
|
Glu-B1
|
Glu-D1
|
Glu-A1
|
Glu-B1
|
Glu-D1
|
|
1 |
T.Aestivum |
Khalh-gol-1 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
2 |
T.Aestivum |
Dakhan-131 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
3 |
T.Aestivum |
Dakhan-172 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
4 |
T.Aestivum |
Darkhan-34 |
Ax2* |
- |
- |
|
b |
|
|
|
5 |
T.Aestivum |
Darkhan-74 |
- |
- |
- |
|
|
|
|
|
6 |
T. Aestivum |
Darkhan-166 |
- |
- |
5+10 |
|
|
|
d |
|
7 |
T. Aestivum |
Darkhan-193 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
8 |
T. Aestivum |
Darkhan-144 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
9 |
T. Aestivum |
Darkhan-181 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
10 |
T. Aestivum |
Darkhan-212 |
Ax2* |
- |
5+10 |
|
b |
|
d |
|
11 |
T. Aestivum |
MK-68 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
12 |
T. Aestivum |
MK-248 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
13 |
T. Compactum |
MK-260 |
- |
- |
- |
|
|
|
|
|
14 |
T. Compactum |
MK-263 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
15 |
T. Compactum |
MK-264 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
16 |
T. Aestivum |
MK-269 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
17 |
T. Aestivum |
MK-270 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
18 |
T. Aestivum |
MK-278 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
19 |
T. Aestivum |
MK-287 |
Ax1 |
- |
5+10 |
|
a |
|
b |
|
20 |
T. Aestivum |
MK-291 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
21 |
T. Aestivum |
MK-299 |
- |
- |
2+12 |
|
|
|
a |
|
22 |
T. Aestivum |
MK-307 |
- |
Bx7OE |
2+12 |
|
|
a |
a |
|
23 |
T. Aestivum |
MK-310 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
24 |
T. Aestivum |
MK-315 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
25 |
T. Aestivum |
MK-316 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
26 |
T. Aestivum |
MK-324 |
- |
- |
2+12 |
|
|
|
a |
|
27 |
T. Aestivum |
MK-325 |
- |
- |
2+12 |
|
|
|
a |
|
28 |
T. Aestivum |
MK-328 |
- |
- |
2+12 |
|
|
|
a |
|
29 |
T. Aestivum |
MK-331 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
30 |
T. Compactum |
MK-336 |
Ax1 |
Bx7OE |
2+12 |
|
a |
a |
a |
|
31 |
T. Aestivum |
MK-338 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
32 |
T. Compactum |
MK-349 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
33 |
T. Aestivum |
MK-350 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
34 |
T. Aestivum |
MK-354 |
Ax1 |
Bx7OE |
- |
|
a |
a |
|
|
35 |
T. Aestivum |
MK-358 |
Ax1 |
Bx7OE |
2+12 |
|
a |
a |
a |
|
36 |
T. Compactum |
MK-362 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
37 |
T. Aestivum |
MK-457 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
38 |
T. Aestivum |
MK-458 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
39 |
T. Aestivum |
MK-459 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
40 |
T. Aestivum |
MK-467 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
41 |
T. Aestivum |
MK-478 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
42 |
T. Aestivum |
MK-916 |
Ax1 |
Bx7OE |
- |
|
a |
a |
|
|
43 |
T. Aestivum |
MK-1140 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
44 |
T. Aestivum |
MK-1145 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
45 |
T. Aestivum |
MK-1565 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
46 |
T. Aestivum |
MK-1166 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
47 |
T. Compactum |
MK-1568 |
Ax2* |
Bx7OE |
2+12 |
|
b |
a |
a |
|
48 |
T. Aestivum |
MK-1582 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
49 |
T. Aestivum |
MK-1592 |
- |
- |
2+12 |
|
|
|
a |
|
50 |
T. Aestivum |
MK-4727 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
51 |
T. Aestivum |
MK-4730 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
52 |
T. Compactum |
MK-4732 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
53 |
T. Aestivum |
MK-4748 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
54 |
T. Compactum |
MK-5584 |
- |
- |
2+12 |
|
|
|
a |
|
55 |
T. Aestivum |
MK-5591 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
56 |
T. Aestivum |
MK-5592 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
57 |
T. Aestivum |
MK-5998 |
- |
- |
2+12 |
|
|
|
a |
|
58 |
T. Aestivum |
MK-5999 |
- |
- |
2+12 |
|
|
|
a |
|
59 |
T. Aestivum |
MK-6001 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
60 |
T. Aestivum |
MK-6007 |
- |
- |
2+12 |
|
|
|
a |
|
61 |
T. Aestivum |
MK-6008 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
62 |
T. Aestivum |
MK-6016 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
63 |
T. Aestivum |
MK-6019 |
- |
- |
2+12 |
|
|
|
a |
|
64 |
T. Compactum |
MK-6020 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
65 |
T. Aestivum |
MK-6026 |
- |
- |
2+12 |
|
|
|
a |
|
66 |
T. Aestivum |
MK-6368 |
- |
- |
2+12 |
|
|
|
a |
|
67 |
T. Aestivum |
MK-6370 |
- |
- |
- |
|
|
|
|
|
68 |
T. Aestivum |
MK-6371 |
Ax2* |
- |
- |
|
b |
|
|
|
69 |
T. Aestivum |
MK-6374 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
70 |
T. Aestivum |
MK-6378 |
- |
- |
2+12 |
|
|
|
a |
|
71 |
T. Aestivum |
MK-282 |
- |
- |
2+12 |
|
|
|
a |
|
72 |
T. Aestivum |
MK-340 |
- |
- |
2+12 |
|
|
|
a |
|
73 |
T. Aestivum |
MK-343 |
- |
- |
2+12 |
|
|
|
|
|
74 |
T. Aestivum |
MK-351 |
- |
- |
- |
|
|
|
|
|
75 |
T. Aestivum |
MK-1150 |
Ax2* |
- |
- |
|
b |
|
|
|
76 |
T. Aestivum |
MK-1564 |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
77 |
T. Compactum |
MK-1595 |
- |
- |
2+12 |
|
|
|
a |
|
78 |
T. Aestivum |
MK-4744 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
79 |
T. Compactum |
MK-5566 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
80 |
T. Aestivum |
MK-5580 |
- |
- |
2+12 |
|
|
|
a |
|
81 |
T. Aestivum |
MK-6376 |
Ax2* |
- |
2+12 |
|
b |
|
a |
|
82 |
T. Aestivum |
Buritskay 34 |
Ax2* |
- |
5+10 |
|
b |
|
b |
|
83 |
T. Durum |
Sondor |
Ax1 |
- |
- |
|
a |
|
|
|
84 |
T. Durum |
Khar Suwd |
Ax1 |
- |
- |
|
a |
|
|
|
85 |
T. Durum |
Caerulescens |
Ax1 |
- |
- |
|
a |
|
|
|
86 |
T. Compactum |
Creticum |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
87 |
T. Compactum |
Fetisowii |
Ax1 |
- |
2+12 |
|
a |
|
a |
|
88 |
T. Aestivum |
NONAME |
Ax1 |
- |
5+10 |
|
a |
|
b |
|
89 |
T. Aestivum |
AGS2000 |
Ax2* |
- |
5+10 |
|
b |
|
b |
Table 2HMW-GS composition of Mongolian wheat cultivars.
Table 2
|
Locus |
Primer |
Allele |
Relative frequency |
Genetic diversity (He) |
Average genetic variation |
|
Glu-A1
|
Glu-Ax1/x2*_SNP |
Ax1 |
0.29 |
0.63 |
0.71 |
|
Glu-Ax2_IND |
Ax2* |
0.44 |
|
|
Glu-B1
|
BX7OE_866_SNPBx13_SNP |
Bx7OE |
0.07 |
0.93 |
|
|
Glu-D1
|
Glu-D1d_SNP |
5+10 |
0.13 |
0.57 |
|
|
2+12 |
0.73 |
References
- Bietz JA, Shepard KW, Wall JS. 1975. Single-kernel analysis of glutenin: Use in wheat genetics and breeding. Cereal Chem. 52: 513-532.
- Bietz JA, Wall JS. 1972. Wheat gluten subunits: Molecular Weights determined by Sodium sulfate-polyacrylamide gel electrophoresis. Cereal Chem. 49: 416-430.
- Branlard G, Dardevet M. 2001. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica. 119: 59-67.
- Buonocore F, Caporale C, Lafiandra D. 1996. Purification and characterisation of high Mr glutenin subunit 20 and its linked y-type subunit from durum wheat. J. Cereal Sci. 23: 195-202.
- Junwei Zang Fl, Haocheng Sun, Jian Wang, Wenjing Duan, Yueming Yan. 2023. Wheat Glu-A1a encoded 1Ax1 subunit enhances gluten physicochemical properties and molecular structures that confer superior breadmaking quality. Int. J. Biol. Macromol. 225: 701-714.
- Masatoshi N. 1972. Genetic distnace between populations. Am. Nat. 106: 283-292.
- Payne PI. 1987. Genetics of wheat storage proteins and effect of allelic variation on bread making quality. Ann. Rev. Physiol. 38: 141-153.
- Payne PI, Lawrence GJ. 1983. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res. Commun. 11: 29-35.
- Payne PI, Nightingale MA, Krattiger AF, Holt LM. 1987. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric. 40: 51-65.
- Rasheed A, Wen W, Gao F. 2016. Development and validation of KASP assays for genes underpinning key economic traits in bread wheat. Springer-Verlag Berlin Heidelberg p.20..
- Saghai-Maruuf MA, Soliman KM, Jorgensen RA, Allard RW. 1984. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics: Proc. Natl. Acad. Sci. USA. 81: 8014-8018.
- Sameri H, Salavati A, Boushehri ASN, Yazdi-Samadi B. 2008. Identification of HMW Subunits in Iranian landrace wheat by using STS-PCR Method. In Proceedings of the 11th International Wheat Genetics Symposium. 24-29 August 2008, Brisbane, Qld., Australia..
- Shewry PR, Tatham AS. 1997. Disulphide bonds in wheat gluten proteins: J. Cereal Sci. 25: 207-227.
- Shiri Y, Solouki M, Frootan M. 2015. Analysis of allelic variation in HMW-Glu-1 gene blocks in Iranian wheat cultivars using ALP molecular markers. Ind. J. Sci. Technol. 8(S3): 111-117.
- Singh NK, Shepherd KW, Cornish GB. 1991. Rapid communication A simplified SDS-PAGE procedure for separating LMW subunits of slutenin. J. Cereal Sci. 14: 203-208.
- Sissons M. 2008. Role of durum wheat composition on the quality of pasta and bread. Food Global Science Book..
- Trittinger S. 2019. Wheat (Triticum aestivum) Kernel polymers as breeding targets for improved end-use quality. Oregon State University..
- Turner AS, Bradburne RP, Fish L, Snape JW. 2004. New quantitative trait loci influencing grain texture and protein content in bread wheat. J.Cereal Sci. 40: 51-60.
- Zhao L, Abdelsalam NR, Xu Y, Chen MS, Feng Y, Kong L, et al. 2020. Identification of two novel Hessian fly resistance genes H35 and H36 in a hard winter wheat line SD06165. Theor. Appl. Genet. 133: 2343-2353.