Abstract
Gene introgression that involves the transfer of favorable allelic diversity for broadening the genetic base of breeding materials is a powerful 'toolkit' for creating novel allelic combinations during plant sexual reproduction. It is a key factor playing an important role in plant breeding schemes by reintroducing genetic variation at selective sweeps or introgression of desirable traits required for the development of new varieties. Meiosis is a specialized cell division not only enables sexually reproducing organisms to reduce their genomic constituent by half, also provides indefinitely novel combinations of allelic diversity by reshuffling the parental genetic makeup. Crossing over that takes place during prophase-I facilitates the meiotically exchange of genetic materials between homolog pairs as well as their accurate segregations. It is tightly modulated and many intrinsic factors and extrinsic agents are associated with regulation of the process, however, the modulation of meiosis is possible. Although, advanced approaches such as CRIPR/Cas and Virus-Induced gene Silencing (VIGS) have opened new horizons for manipulation of meiotic recombination, distant hybridization could effectively influence the frequency and distribution of homologous (HR) and particularly homoeologous recombination (HeR). In this review, we provide a brief overview of the recent advances in the plant mechanisms for manipulation of HR and HeR employing distant hybridization.
-
Key words: Whole-genome duplication, Distant hybridization, Crossover, Homoeologous recombination, Selective sweep, CRIPR/Cas, Virus-Induced gene Silencing (VIGS)
INTRODUCTION
Gene introgression is a powerful 'toolkit' for creating novel allelic combinations during plant sexual reproduction (
Kopecký et al. 2010). It involves the transfer of favorable allelic diversity between or within crop species as well as sometimes between genera through wide hybridization followed by backcrossing (
Dreissig et al. 2020). It is a common phenomenon in plant breeding schemes playing an essential role in reintroducing genetic variation at selective sweeps or introgression of desirable traits from wild relatives into the crop gene pool required for the development of new varieties. Genetic variations on which artificial or natural selection could accomplish is the significant consequence of meiotic recombination that reshuffle parental genetic makeup. It is fundamental for both plant evolution and genetic improvement programs by providing indefinitely novel combinations of parental alleles through gene conversion and crossovers and ultimately affirming balanced segregation of homologous pairs (
Henderson and Bomblies 2021). Alien gene introgression, on the other hand, could result in accidental extensive genomic and epigenetic modifications in plant materials, paving the way for more diversity and artificial selections (
Wang et al. 2005). We will briefly cover recent progress in plants toward mechanisms controlling meiotic homologous (HR) and homoeologous recombinations (HeR) in this study. We will then discuss how whole-genome duplication (WGD) and distant hybridization could significantly improve HR and, in particular, HeR for plant breeding objectives.
DISTANT HYBRIDIZATION: AN APPLIED APPROACH FOR MANIPULATION OF MEIOTIC RECOMBINATION
Meiosis and recombination: a relationship
Meiosis is a two-round specialized cell division that follows only one round of DNA duplication and gives rise to haploid sex cells (gametes). Crossovers (COs) that take place during prophase-I dictates the meiotically accurate segregation of homologous pairs by generating physical connection between homologous chromosomes. However, at least one CO is obligatory for balanced segregation of chromosomes per homolog and per meiotic division, the event that is called “crossover assurance” (
Saito and Colaiácovo 2017). Sexual infertility and aneuploidy are common phenomena tightly associated with failure in COs formation in eukaryotes (
Xiong et al. 2011). Furthermore, COs formation directs meiotic recombination in prophase I where homologous chromosomes temporarily link and allow reciprocal exchange of genetic materials, the process that modifies two homologous non-sister chromatids and creates novel genetic combination of existing allelic variation (
Fig. 1, for review, see
Wang et al. 2021). Surprisingly, the frequency of local and genome-wide recombination rates (GWRRs) could influence the chance of disrupting linkage drag, the simultaneous introgression of deleterious and favorable alleles in wide crosses, and ultimately enhance the efficiency of subsequent selection toward desirable allelic variation (
Bohra et al. 2021). The lack or low rates of recombination along the chromosome is a serious problem making use of conventional breeding approaches including development of large segregating populations or employ-ment of several backcrossing generations to remove deleterious alleles. However, crossovers may disassociate desirable alleles on preferred haplotypes, which could negatively affect progeny fitness (Otto and Lenormand 2002). COs frequency, for example, has been shown to be negatively correlated with genome size, because of the elimination of long terminal repeat (LTR) retrotransposons (
Tiley and Burleigh 2015). In corn, approximately 25% of the genome may experience less than a one in a thousand probability of a CO per megabase (Mb) per recombinant inbred lines (RILs), which accounts for only 12% of all annotated gene space, imposing a significant linkage drag (
Rodgers-melnick et al. 2015). Increased crossover rates have been found to be 3.5-fold positively connected with higher mutation rates, and may occur more frequently in heterozygous people and locations close to hotspots (
Serra et al. 2018).
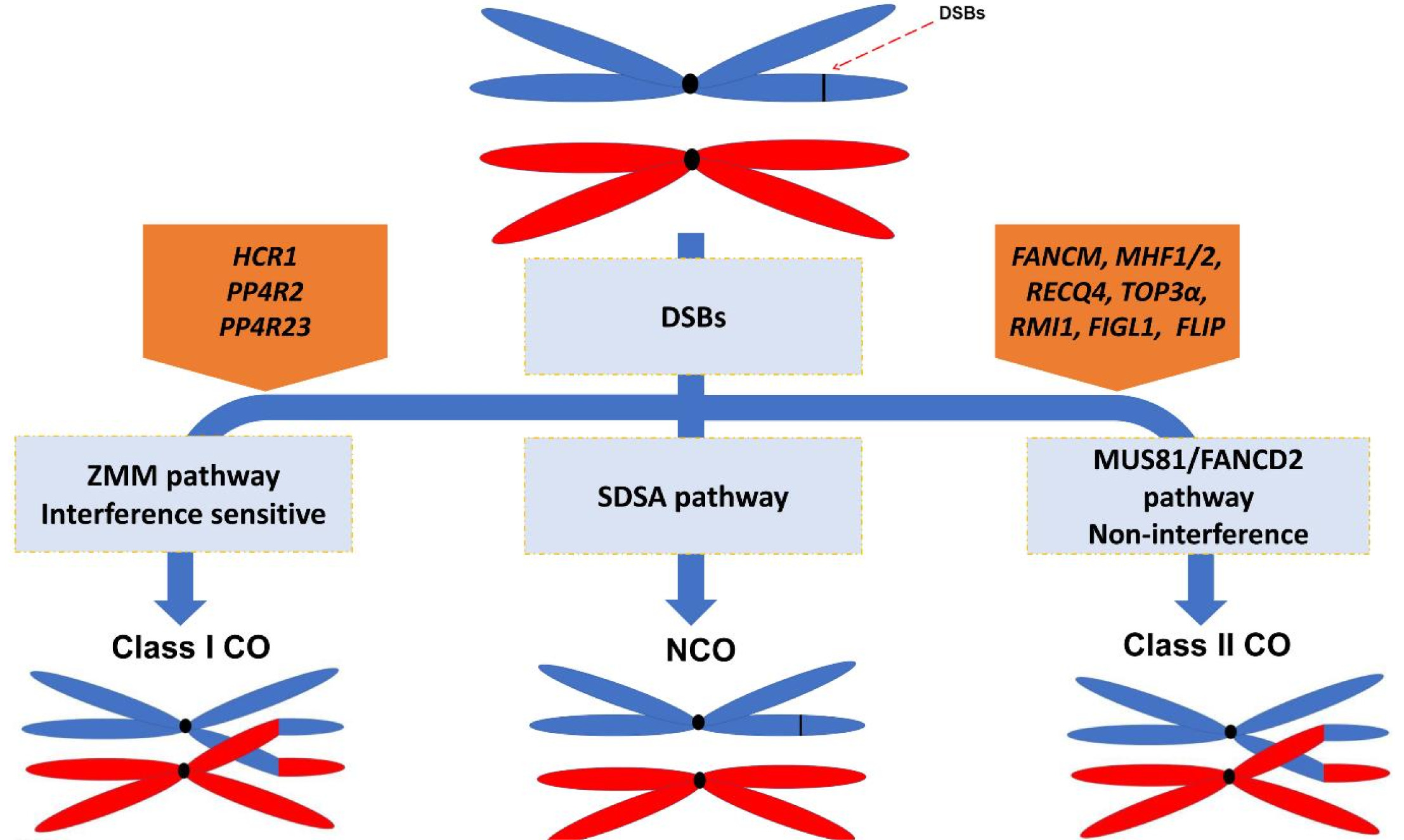
Meiotic recombination is initiated by generating hundreds of DNA double strand breaks (DSBs) that is precursor for COs. DSBs formation is catalyzed by an evolutionarily conserved topoisomerase-like protein complex consisting of SPORULATION-DEFICIENT 11 (SPO11) protein and several other joint proteins. There are two major mechanisms promoting the DSBs repair during meiotic recombination. The first mechanism is synthesis-dependent strand annealing (SDSA) which produces non-crossovers (NCOs). The other mechanism is homologous recombination which leads to COs or NCOs (
Wang and Copenhaver 2018). In
Arabidopsis thaliana, the majority of DSBs results in NCOs and maximum 4% to 7% of DSBs are repaired to COs (
Fig. 1) proposing the existence of mechanisms limiting COs number (
Ferdous et al. 2012).
At least two major pathways independently facilitate COs formation in plants. The first pathway repairs DSBs to “class I” COs that is also known interference sensitive COs. This pathway depends upon the activity of a group of protein collectively so-called ZMM pathway [Zipper (ZIP1), ZIP2, ZIP3, and ZIP4, Meiotic recombination 3 (Mer3), and MutS homologue 4 (MSH4) and MSH5] and MutL-homolog 1 (MLH1) and MLH3 and its formation consequently ensure an obligate crossover per homologs. Therefore, in zmm mutants such as hei10 of zip4, COs frequency has been significantly decreased. In plants, the majority of COs are formed through ZMM pathway. The second pathway results in “class II” COs that is interference insensitive and is not randomly distributed along the chromosome, mostly depends on the activity of endonuclease MUS81 (methyl methansulfonate and ultraviolet-sensitive gene clone 81) (
Anderson et al. 2014). In Arabidopsis and corn, DSBs are not uniformly distributed across the chro-mosomes and preferentially initiated in nucleosome-free regions and hypomethylated DNA sequences sites (
Choi et al. 2018). It is estimated that in the most living organisms, -25% of the genome monopolizes almost 80% of the COs (
Blary and Jenczewski 2019).
Although meiotic recombination is strictly regulated by several mechanisms and/or pathways (
Table 1), the pattern and overall number of COs exhibit high variations between and within eukaryotic species (
Haenel et al. 2018). Extrinsic environments like solar radiation, temperature (
Coulton et al. 2020), isothermality, and annual precipitation, epige-netic modifications, and genetic factors (e.g., genetic background as described in Brassica (
Mason et al. 2011) and banana (
Jeridi et al. 2011), sex as described in corn (
Kianian et al. 2018) and cassava (
Chan et al. 2022), and homology as proposed in potato (
Speranza et al. 2017)) are capable of profoundly influencing crossover position and frequency while remarkably advancing genetic improvement programs (
Dreissig et al. 2019). Furthermore, COs sites are unequally non-random distributed along the chromosome with a gradient from chromosome distal ends to pericen-tromeric regions (
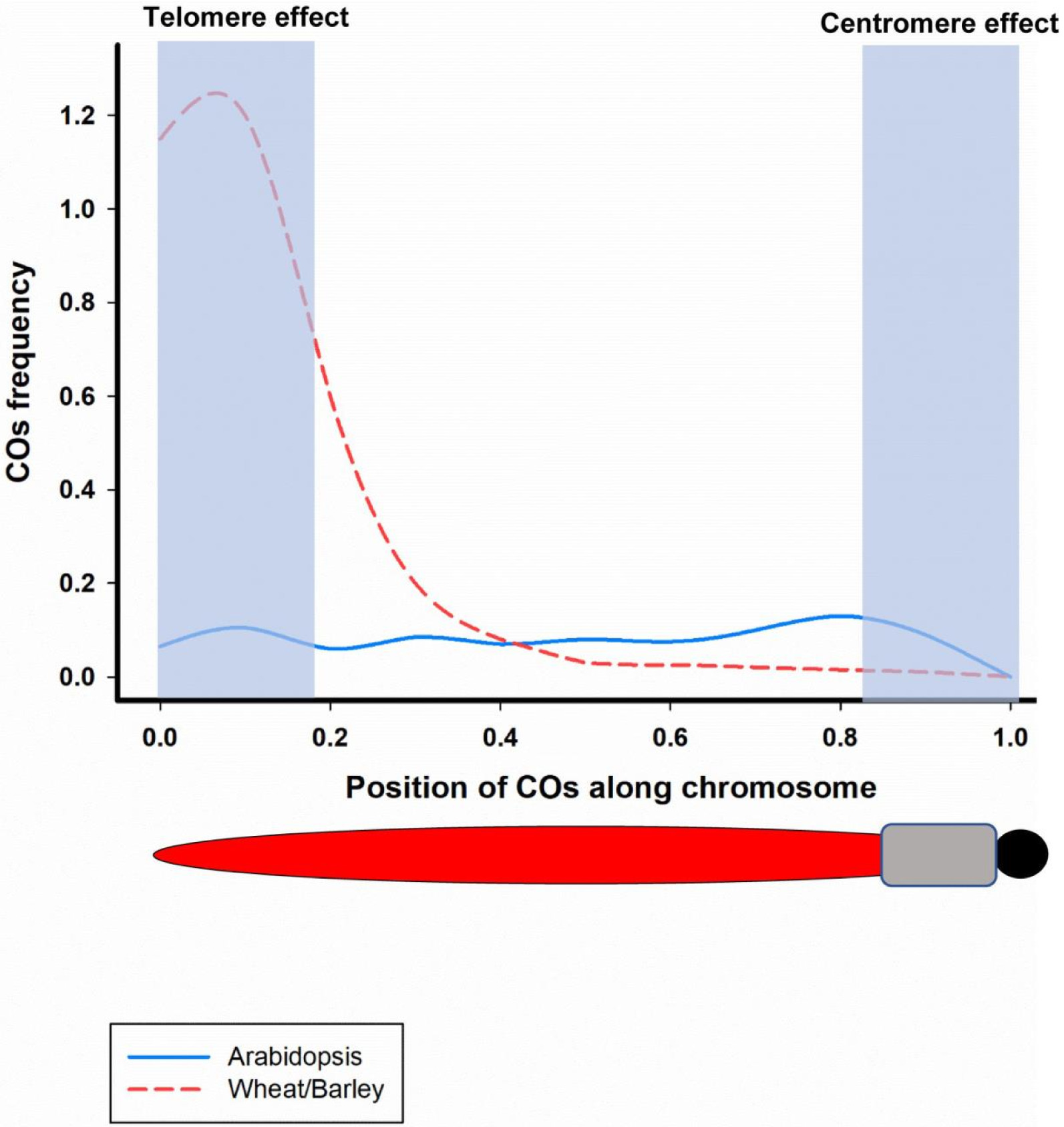
Fig. 2) (
Haenel et al. 2018) pointing out the effects of genomic and epigenetic features on meiotic recombination (
Yelina et al. 2015). In rice, suppression of CO at centromere and surrounding regions corresponds to almost 11% of chromosome total length (
Fayos et al. 2019). However, COs are highly repressed at centromeric regions of all species. It has been reported that the occurrence of COs adjacent to centromere could lead to improper chromosome segregation and sterility (
Wang and Copenhaver 2018). Notably, subtelomeric region of chromosomes that is proximal to telomer of chromosome contains recombination hotspots and fast-evolving gene determining plant adaptability and fitness (
Chen et al. 2018). Hypomethylated genomic regions containing functional genes reveal higher proportions of COs, while they might be far reduced or even suppressed in hypermethylated chromosomal regions or regions carry any divergent sequence and structural mutations (Rowan
et al. 2019).
Recombination is heterochromatic regions was expected to be positively associated with DNA demethylation. Loss of function in CG DNA methyltransferase1 (
Met1), required for maintaining methylation pattern in assistance with SWI/SNF chromatin remodeling protein Decreased DNA Methylation1 (
DDM1) during DNA duplication, could induce DNA hypomethylation, however, it could not elevate recombination initiation sites and chance of crossover in centromere proximal regions (
Melamed-bessudo and Levy 2012). However, an ultimate crossover distribution reshaped by
met1 and
ddm1 mutants has been observed within subtelomeric regions has been reported (
Choi et al. 2018). On the contrary, CHH (where H =A, T, or C) DNA methylation within sequences immediately adjacent to active genes could drive the crossover landscape much more efficient than CG and CHG methylation in corn (
Zhao et al. 2021).
Changes in ploidy level, also known as polyploidization or whole genome duplication (WGD), have accelerated the evolution of most seed plants (
Rey et al. 2021). The other most obvious result of polyploidy in plants is increased cell size, which could lead to larger plant parts. Although it has the potential to cause severe genomic instability, chromosomal rearrangements, gene losses, and changes in gene expression patterns, particularly when combined with hybridization, and thus lead to sexual sterility (
Schmickl and Yant 2021), there is growing evidence that WGD can promote introgressive hybridization to transfer desirable traits by introducing relatively small genomic segments from one species to another. However, the resulting genome instability in newly produced allopolyploids may afford opportunity for important genetic changes (
Chester et al. 2011). It may eventually contribute to greater growth performance and (
Carlos and Ramirez-Parra 2015). Additionally, employing existing genetic materials, WGD is a potent genomic technique that can restore hybrid sexual fertility (
Storme et al. 2020), develop novel phenotypic variety (
Wu et al. 2021), and vast ploidy variation (
Yin et al. 2020). WGD, on the other hand, is a one-of-a-kind evolutionary event that allows an entire organism to benefit from numerous homoeologous subgenomes.
Polyploid species, organisms possessing more than a diploid chromosome complement, fall into two distinct groups: autopolyploids, which carry three or more sets of the same genome originated from a common progenitor, and allopolyploids, in which interspecific hybridization of related species followed by WGD (
Rey et al. 2021). Allopolyploidization has played a crucial role in the evolution of various crop plants. Interspecific hybridization coupled with WGD has provided novel opportunities of incorporating various genomes at different ploidy levels. The world’s major field crops, including wheat, sugarcane, cotton, and rapeseed are among relatively novel allopolyploid crops experienced interspecific hybridization between related species or divergent populations coupled with WGD (
Bomblies et al. 2015). Allopolyploid crops mostly demon-strate higher heterosis, and better yield performance and adaptability across wider geographical contexts mostly due to fixation of heterozygosity in hybrid progenies (
Ozkan and Feldman 2001). Gaining extra gene dosages and extensive genomic changes (deletions, inversion, duplications, and translocations) that happen after divergent genomes merge are the main reason to explain how alloploids display novel allelic variations and better adaptability compared with their ancestors (
Zou et al. 2011). For example, in bread wheat, it has conveyed the benefits of heterosis and gene redundancy that enhances genetic variation, plasticity, and adaptability (
Wan et al. 2021). Gene redundancy could alleviate the negative effects of deleterious mutations arising from polyploidy and, thus promoting the evolution of the crop plants. However, the response to WGD may sometimes vary across different taxa and have negative influence on plant growth performance particularly in neo-autopolyploids.
WGD is usually followed by immediate evolutionary changes required for better adaptability and fitness of hybrids including increasing diploidization, gene conversion, transposition, and gene loss (
Tennessen et al. 2014). For example, upland cotton (
Gossypium hirsutum L.) is an allotetraploid crop plants that benefits from the different genomic constituents, however, an inversion on chromo-some A08 had a negative impact on meiotic recombination, thus resulting in reduced genetic variation in inversion containing regions (
Yang et al. 2019). Like
Tragopogon miscellus (
Chester et al. 2011), genomic rearrangement induced by allopolyploidization is the main source of allelic variation, however, it has led to minimum 5% gene loss across the genome through homoeologous recombi-nation in coffee (
Coffea arabica) (Diade
et al. 2016). Like allopolyploidy, segmental allopolyploidy may also result into loss of allelic variations as a consequence of homoeo-logous recombination between subgenomes (Leal-bertioli
et al. 2018). It occurs when chromosome belonging to subgenomes demonstrate partial homology leading to intermediate pairing behavior is another major evolu-tionary mechanism.
The increased rate of HR and HeR induced by auto- and allopolyploidization in a variety of plants including Arabidopsis (
Bomblies et al. 2015), wheat (
Sheikh et al. 2016), tomato (
Fuentes et al. 2022),
Brassica (
Pele et al. 2017), barley (
Scholz and Pendinen 2017), peanut (
Chu et al. 2021), grapevine (
Delame et al. 2019), rice (
Zhang et al. 2019), lily (
Barba-Gonzalez et al. 2005), and
Lolium perenne (
Harper et al. 2018) has been reported (
Table 2). Although WGD is a promising approach for modifying COs rate and distribution in crop species, the efficiency of this technique depends on various factors and varies from species to species. In addition, the mechanism underlying enhanced COs has not been well understood.
It is evident that domestication might has reduced hotspot abundance and, as a result, COs frequency and chromosomal patterning. However, there may be a way to use the genomic structure of wild relatives to induce recombination in domesticated crops without introducing new genes or desirable traits during selection sweeps, especially in the pericentromeric heterochromatin (
Fuentes et al. 2022). In practically all crop species, domestication has essentially achieved selection for beneficial genes situated in CO-rich regions of chromosomes, suggesting that the majority of allelic differences have not been fully utilized (
King et al. 2007). Regulation of HeR is a key component to ensure the success of distant hybridization that influences the genetic structure of next generations. It may be restricted or promoted by either a single gene in wheat (
Sheikh et al. 2016;
Copete-parada et al. 2021),
Aegilops speltoides (
Li et al. 2017),
Aegilops geniculata (
Koo et al. 2020), corn (
Zhao et al. 2021) or major QTL in Brassica (Higgins 2021) and multilocus interactions in polyploid cotton (
Jiang et al. 2000). However, induction of homoeologous recombinations between chromosomes of related subgenomes to transfer traits of interest into crop species should be followed by urgent reactivation of homoeologous recombinations repressing genes, such as
Ph1 in wheat, to prevent rapid removal of target DNA segment(s) resulting in diploid-like behavior of chromo-somes during meiosis. For example, ph1 mutant that lacks of functioning
Ph1 has shown massive increase in chromo-some pairing in distant hybridization of bread wheat with rye and
Ae. columnaris accounting for 37% and 50%, respectively (
Verma et al. 2016). However, cryptic alien gene introgression could induce significant mutations in the recipient genome and may enhance sequence variations in both coding and non-coding sequences and TEs (
Wang et al. 2005).
Distant hybridization is a cost-effective method for reintroducing beneficial genes at selective sweeps or introducing desirable alleles into the crop gene pool, both of which are required for the production of high-yielding, well-adapted varieties. In particular, hybrid sterility is a typical occurrence in remote hybridization, limiting its use in agricultural development projects. The most common cause of hybrid sterility, which results in imbalanced chromosome segregation and aneuploidy, is the suppression of crossovers between homoelogs with excessively different DNA sequences (
Bozdag et al. 2021).
Induction of HeR is not only an effective strategy to overcome hybrid sterility, also a key factor determining allelic diversity. Nonetheless, HeR is a major mechanism leading to gene presence/absence variation (PAV) pattern in major crop plants including
B. naups of which nearly 38% of genes have displayed PAV pattern (
Hurgobin et al. 2018). Wide hybridization particularly interspecific hybridization may give rise to some serious abnormalities affecting hybrids performance as reported in newly formed allotetraploid (
Cucumis ×
hytivus). Suppression of genes catalyzing chlorophyll biosynthesis,
HEMA1,
HEME2 and
POR, which causes delayed leaf maturation, may be attributed to gene loss/retention or non-reciprocal homoeologous recombination in interspecific hybridization (
Yu et al. 2018). Interestingly, the occurrence of homologous recombi-nation at upstream and/or downstream of coding sequences has been reported to potentially induce frequent HeR within gene bodies, thus creating intragenic recombination and forming novel chimeric genes (
Zhang et al. 2020). Homoeologous exchanges (HEs) not only alters gene copy number in allopolyploid and results to dosage-dependent changes gene expression, also may induce epistatic inter-actions between HEs loci. For example, in rice approxi-mately 32.8% of loci may demonstrate epistatic interaction giving rise to huge novel phenotypic variation (
Wu et al. 2021). It is also capable of causing genomic rearrangements to influence quantitative (polygenic) variation in
B. napus (
Stein et al. 2017) or even creating quantitative trait loci (QTLs) associated with major agronomic traits including disease resistance in tobacco (
Shi et al. 2022).
Although, the frequency and patterning of recombina-tion is under tight control, AABC, BBAC and CCAB allotriploid Brassicas are capable of overcoming this barrier by inducing higher ratio and wider distribution of recombination across the chromosomes or to reintroduce genetic variation at selective sweeps or introgress novel allelic variation from close species,
B. rapa and
B. oleracea, into modern
B. napus. (
Xu et al. 2019).
Enhanced recombination frequency has been reported in AAC hybrids allowing the introgression of more and smaller genomic segments from
B. rapa to
B. napus compared to AACC allotetraploid hybrids. Indeed, the trigenomic hybrids could be used as a bridge for introgressing desirable traits form either close or distant wild relatives into
Brassica gene pool (
Chen et al. 2011). However, the COs frequency in allotetraploid (AACC) has been reported to be intermediate between the diploid progenitors and allotriploid hybrids (
Leflon et al. 2010). The smaller the size of the introgressed segment, the higher the efficiency of gene introgression would be that is extremely beneficial for quick breakdown of the linkage disequilibrium. Interestingly, these AAC allotriploid hybrids could be also highly useful for characterization of favorable genes particularly in coldspots located in pericentromeric regions (
Boideau et al. 2021). Nonetheless, the very first meiotic division of newly synthesized B. napus has been proposed to play an important role in blending genomes and restructuring the allelic variations transmitted to progenies in different proportions meaningfully influenced by cytoplasmic background (
Nyarko and Mason 2021). However, the incidence of either homoeologous recombi-nations or chromosomal rearrangement events in the A and C genomes may result into loss of some alleles that could not question the advantageous of non-homologous exchange in
Brassica (Attri and Rahman 2018).
Despite this fact, repressing non-homologous recombi-nation in interspecific hybrids has significantly reduced the efficacy of mining novel allelic variations within genetic resources. Interspecific hybridization between tomato (
Solanum lycopersicum) and wild relative,
S. lycopersicoides, gave rise to severe decline in GWRRs (
Chetelat et al. 2000). Surprisingly, the allopolyploids have evolved an efficient system promoting the distribution of homologous recombination across the genome and subsequently ensuring genome stability and hybrid fertility. This mechanism leads to stable incorporation of desirable alleles from their close wild relatives into crops that is prerequisite for development of new varieties.
Newly synthesized AAC allotriploid Brassica hybrids owning the genomes of two different species,
B. napus and
B. rapa, presents a great opportunity for transfer small desirable genomic segments of
B. rapa into B. napus. Indeed, allotriploidy not only fundamentally
recombines B. napus allelic variations, but also significantly decrease the size of favorable QTLs of
B. napus. In other hand, the genomic structure of these allotriploids enables the manipulation of COs frequency (×3.7) and, preferentially, COs distribution across the chromosomes even in poor-CO regions (
Boideau et al. 2021). AAC allotriploid
Brassica has also demonstrated enhanced overall homoeologous recombination (1.7 to 3.4-fold) between C and A genomes relative to diploid progenitors mostly depending on the genetic background and the sex of meiosis. The event, surprisingly, was associated with considerable reduction in COs interference and tangible changes in the shape of recombination patterning. The COs distribution was also reported to move toward regions vicinal to centromere in male meiosis (
Pele et al. 2017). It could provide the chance of gene introgression and disruption of linkage drag best suited for plant breeding purposes alone.
The COs frequency mostly depends on the origin of the allopolyploid hybrids. The Pairing Regulator in B. napus (
PrBn) on chromosome 9 (C genome), and a large-effect QTL,
BnaPh1 (
BnaA9) on chromosome 9 (A genome), are responsible for modulating homoeologous chromosome associations in alloploid B. napus (
Higgins et al. 2021). Notably, HR frequency may not be affected by an allele alone, e.g.
PrBn allele in ArAnCC hybrids, proposing that the recombination could be mostly background karyotype dependent (
Soares et al. 2021). The characterization of the genes controlling homoeologous chromosome pairing or allosyndesis will be highly beneficial for better understanding of molecular mechanisms leading to efficient homoeologous recombination in allopolyploid plants.
Homoeologous recombination facilitated introgression of fertility restorer gene,
Rfo, from radish (
Raphanus sativus L.) into rapeseed genetic background through homoeologous recombination. However, extensively successive back-crossing was employed to enhance the female fertility of the introgressed lines (
Delourme et al. 1998). Similarly, boosted intergenomic homoeologous COs have been reported in trigeneric hybrids between
Aegilops tauschii,
Hordeum chilense, and
Secale cereale with H
chDR genomic constitution. The homoeologous COs considerably taken place in H
chDR hybrids, 46.5% and 32.4% for H.
chilense -Ae.
tauschii and Ae.
tauschii -
S. cereale genomes, respectively, demonstrating the potential application of distant hybridization for promotion of meiotic recombi-nation. Higher HeR rate may be attributable to the fact that some of these species/accessions may carry some particular alleles that play an important role in promoting or suppressing HeR. For example, rye possesses a polygenic system that modulates the homoelogous chromosme pairing between in rye-wheat hybrids. However, in trigeneric hybrids between
Triticum turgidum spp.
Durum,
H. chilense, and
S. cereale, hybrids with ABH
chR chromosome composition lack of COs between B genome with other genomes in ABH
chR hybrids may be attributed to the presence of
Ph1 genes restricting non-homologous recombination (
Rey et al. 2021).
Although wide hybridization proved to be an efficient technique to widen the genetic base of crop plants, it may give rise to some major issues in hybrids and subsequent generations. Due to the strong connection between beneficial and deleterious genes, linkage drag is an adverse limitation of exotic gene introgressions that has a negative impact on progenies’ adaptability and fitness. Clearly, the larger the alien segment size, the greater the risk of transmitting unfavorable allelic variation from low-adapted exotic genetic resources. For example, introduction of rye allelic variations (
Secale cereale) into wheat genetic background has negatively influenced its baking quality (
Kumlay et al. 2003).
Introgression of genes from distantly related species especially the tertiary genepool could be highly promising for enhancing homoeologous recombination in bread wheat (
Fig. 3). Newly synthesized allohexaploid wheat formed by crossing allotetraploid
T. turgidum and diploid
Ae. tauschii, has shown significant increase in meiotic recombination of the ancestral diploid D genome of synthetic wheat. It was reported that allopolyploidization not only promotes genetic recombination, but also unexpectedly increases segregation distortion in D genomes likely due to either sequence elimination or interchromosomal DNA exchange (
Wan et al. 2021).
Deletion/mutation of gene(s) restricting homoeologous recombination is an applied approach allowing the induction of intergenomic recombination between homoeologous pairs. Pavon monosomic for chromosome 5B that lacks
Ph1 allele is an efficient approach to stimulate homoeologous recombination between chromosomes of wheat and
Aegilops species or other related species for meticulous introgression of novel genetic variations (
Sheikh et al. 2016). There is growing evidences supporting the fact that suppression of
Ph1 locus on chromosomes B5L could be possibly achieved by employing chromosomes from wheat wild relatives including chromosome 5U in
Ae. umbellulata Zhuk and chromosome 5E in
Elytrigia genicula (Host)
Nevski. Although, wheat chromosomes share distal forma-tion of COs/chiasmata that is a very common event during meiosis,
ph1 mutants carrying
Hpp-
5Mg chromosome has shown highly increased homoeologous recombination distributed to even proximal regions with low COs frequency (
Koo et al. 2020).
TaZIP4-B2, located within the
Ph1 gene could significantly boost homoeologous COs in wide crosses of bread wheat with wild species. More recently, it was reported that growing
Tazip4-B2 ph1b-rye hybrids in Hogland solution with increased Mg
2+ supple-mentation could boost HeR from a mean of 1 CO to 7 COs per cell to up to 12 COs per cell (
Rey et al. 2018). Induction of homoeologous pairing has also led to successful introgression of useful allelic variations from
Ae. kotschyi into bread wheat prerequisite for developing varieties with higher grain Fe and Zn content. Monosomy of chromosome 5B in interspecific hybrids has led to nonhomologous recombination of wheat chromosomes with
Ae. kotschyi (
Verma et al. 2016). However, it was proposed that several generations of backcrossing should be inevitably accom-plished to ensure the genomic stability and introgression of compensating translocations into recipient genetic background as reported in crosses between
T. aestivum–Dasypyrum villosum hybrids (
Zhang et al. 2015).
Intergeneric hybridization in combination with chromo-somal duplication is another effective way to fundamen-tally incorporate the chromosome complements of key crops while gaining the benefits of their genetic advantages. It's a potential method for increasing CO ratios and frequency. Oilseed rape (
B. napus) genes were successfully introgressed into wild radish using this technique (
Adamczyk-chauvat et al. 2017).
Amblyopyrum muticum is a novel source of allelic diversity highly beneficial for improving major agronomic attributes of bread wheat. The species possesses two genes with two different variants inhibiting the activity of the bread wheat
Ph1 locus, thus allowing high frequency HeR to take place directly between chromosome pairs in different subgenomes in
Am. Muticum/wheat intergeneric hybrids (Dover and Riley 1972). In intergeneric hybrids of
Am. Muticum/wheat, significant HeR ratio between chromo-somes in
Am. Muticum T genome and wheat B and D genomes has been reported accounting for 21 and 18 recombination events, respectively, leading to gene introgressions from
Am. Muticum into wheat (
King et al. 2017). Moreover, the identification of two
Ph1 repressor alleles, 3S (
Su1-Ph1) and 7S (
Su2-Ph1) on chromosomes 3S and 7S of
Ae. speltoides, respectively, has opened novel possibilities for promoting HeR in wheat ×
Ae. speltoides hybrids (
Dvorak et al. 2006). Consequently, these loci could potentially affect chromosomal associations between homoelogs, ranging from 7.0 to 16.4 chiasmata per cell. The allele
Su1-Ph1 has been successfully merged into wheat genetic background, providing new opportunities for enhancing HeR for introgression of desirable traits. Adding a particular chromosome in wheat and developing unique chromosome substitution (CS) lines carry chro-mosome 5E from
Elytrigia genicula (
Dvorak 1987) or chromosome 5U from
Ae. umbellulate are other applicable approaches to suppress
Ph1 locus and induce HeR in wheat (
Koo et al. 2020). The repressive influence of a gene(s) on the functionality of
Ph genes and led to considerable change in HeR between 5M
g and 5D in CS lines with a chromosome 5M
g from Ae. geniculate has been also reported (
Liu et al. 2021). The higher HeR may be triggered by the existence of alien allele(s) located on the 5M
g chromosome either through inducing HeR associ-ations or suppressing the
Ph1 gene (
Svacina et al. 2020). Similarly,
ph1 deficient wheat lines have provided the possibility of gene introgression from other relative including
Lophopyrum elongatum through amphiploidy to bread wheat (
Xu et al. 2020).
Notably, despite the prevention of HeR, the
Ph1 gene probably does not influence homoeologs pairing as reported in wheat/barley hybrids (Calderón and Prieto 2021). Similarly, the gene
phKL posited in wheat landrace ‘Kaixianluohanmai’ (KL), not likely allelic to each of Ph1 genes, could give rise to enhanced homoeologous chromo-some associations in intergeneric hybrids between KL with
Ae. variabilis, rye, and
Psathyrostachys huashanica (
Hao et al. 2011).
The presence of
Ph1 gene is the most important barrier to HeR in wheat. Notably, the
Ph1 locus probably is responsible for detecting similarity of DNA sequences during early meiosis allowing the occurrence of crossover between highly similar sequences and ignoring it while the sequences show divergence (
Fan et al. 2021).
Rey et al. (2015) induced HeR in intergenric hybrids between wheat and cultivated barley (
H. vulgare) and a wild barley
H. chilense employing
ph1b mutant. However, the chromosome pairing induced by
ph1b mutant is very low and estimated to be nearly 4% in crosses between wheat and rye (Miller
et al.1994). It is the process of utmost importance to facilitate gene introgression and chromosomal translocation mediated by
ph1b mutant in such wide crosses to widen the genetic base of wheat. although, the chromosomal translocations take place randomly along the chromosomes, the ph1b mutant could manage chromosomal translocation to preferably occur at particular sites capable of inducing HeR (
Rey et al. 2015).
In intergeneric hybrids between
F. pratensis ×
Lolium multiflorum, HeR hotspots have shown to be unequally distributed along the chromosome pairs, with maximum incidence in intercalary chromosomal regions of the chromosome arms and minimum in centromeric and telomeric regions, thus, suggesting the feasibility of gene introgression from festuca into
L. multiflorum genomic background (
Kopecký et al. 2010). A high proportion of intergenomic recombination within chromosomes of tetraploid hybrids derived from a cross between barley and
H. bulbosum has been reported. Removal of wild barley,
H. bulbosum, facilitated the recovery of barley genome and production of diploid
HvHb introgression lines (
Scholz and Pendinen 2017). In grape vine
Vitis vinifera, there is a similar report demonstrating the influence of alien gene introgression from
V. rotundifolia on reshaping the recom-bination patterns. Although, meiotic recombination could influence HeR in grape vine, its efficiency mostly depend upon the size of introgressed segments (
Delame et al. 2019).
ADVANCED TECHNIQUES FOR MANAGING COS FREQUENCY AND DISTRIBUTION
It is now possible to induce HR and HeR through conventional techniques such as increased temperatures (
Modliszewski et al. 2018), phtoperiod (
Strelnikova et al. 2021), elevated ion concentration i.e. Mg
2+ (
Rey et al. 2018) along with the application of anti-COs mutant or suppressor genes such as
phb1 (
Rey et al. 2015),
phKL (
Hao et al. 2011),
Su1-Ph1 and
Su2-Ph1 (H.
Li et al. 2017). Virus Induced Gene Silencing and newly developed genome editing techniques such as CRISPR/Cas9 gene inactivation, Zinc-Finger Nucleases (ZFNs), Transcription Activator-Like Effector Nucleases (TALENs), and Targeting Induced Local Lesions IN Genomes (TILLING) approaches are promising approaches providing opportunities for efficient manipulation of meiotic recombination.
CRISPR/Cas9 technology has also the potential of inducing chromosomal inversions suitable for crop improve-ment to break or stabilize genetic linkages (
Rönspies et al. 2021;
Rehman et al. 2022). HeR that is recombination between two similar but nonidentical sequences will result in formation of a heteroduplex intermediate that carries mis-paired DNA. The mismatch repair system that depends on the activities of major genes e.g.
PMS1 and
MSH2 significantly decreases the likelihood of meiotic HeR (
Chambers et al. 1996). Nonetheless, RNAi- mediated Silencing of the
MSH2 gene in crop tomato has accompanied by enhanced meiotic recombination. CRSPR/Cas9 mediated inactivation of
RECQ4 (recq4 mutants), a strong class I COs repressor, has been resulted in 1.53-fold increase in homoelogous recombination in interspecific hybrids in tomato (de Maagd
et al. 2020). The application of this gene is not affected by heterozygosity of the plant genome making it potential candidate for development of hyper-recombinant plants (
Li et al. 2021). Furthermore,
Mieulet et al. (2018) found that
recq4 single mutant could increase COs frequency nearly three-fold in rice (
Oryza sativa), pea (
Pisum sativum) and tomato (
Solanum lycopersicum) making it a potential toll for enhancing COs in plants. More interestingly, CRISPR/Cas9 has been also exploited in hybrid rice to completely eliminate genetic interference by disrupting
ZEP1, a major gene encoding the central component of the meiotic synaptonemal complex (SC) and subsequently enhance meiotic recombination by about 1.01 to 3.57-fold (
Liu et al. 2021).
CONCLUSION
By reshuffling the parental genetic makeup, meiotic recombination plays a critical role in expanding the genetic base of crop species. It also makes it easier to introduce new allelic diversity or reintroduce genetic variety during selection sweeps, both of which are necessary for developing high-yielding, well-adapted cultivars. Although it is tightly regulated and numerous genes and QTLs, such as
RECQ4,
FANCM,
FIGL1,
MEICA1,
PrBn, and
BnaPh1 are involved in the regulation of meiotic recombinations, it is possible to manipulate the regulatory mechanism by exploiting intrinsic (
cis and
trans acting factors,
Table 1) or extrinsic factors (temperature, photoperiod conditions, or nutritional avail-ability) to induce target recombination.
Wide hybridization followed by WGD has proven to be an effective tool for reshaping the recombination patterning and frequency. There is mounting evidence that interspeci-fic and intergeneric hybridizations can result in significant inter-homeolog recombination. However, it is possible that it will cause a significant mutation, resulting in genomic instabilities like as meiotic instability, epigenetic shock, changed chromatin architecture, disrupted gene expression patterns, and hybrid fertility (
Wang et al. 2021). However, pre-adapted alleles must be introduced into rescued nascent hybrids in order to assure adaptation to WGD (
Marburger et al. 2020). Enhanced MRFs promote genome stability and proper chromosomal disjunction during meiosis, as well as increasing genetic variation in polyploids by reshuffling existing genetic variation (
Pecinka et al. 2011). Recently developed technologies, like as VIGS and, in particular, CRISPR/Cas9, have opened up new possibilities for manipulating genes that regulate meiotic recombination (
Rönspies et al. 2021). CRISPR/Cas mediated gene silencing would have a huge impact in inducing targeting HeR aiming to introduce desirable allelic diversity from wild relatives into crop plants, which would be a game-changing breakthrough. Next-generation sequencing (NGS) may also pave the path for the identification and regulation of homoeologous recombination genetic pro-cesses. However, identifying and/or manipulating allelic variations responsible for homoelogous chromosome pairing is a prerequisite for a significantly more precise control of HeR, allowing for the introduction of desirable genetic diversity from distant species into crop plants for breeding purposes via HeR.
-
CONFLICT OF INTEREST
The authors declare that they have no competing interests.
Fig. 1Pathways supressing COs frequency in plants. Restriction of COs frequency across the Arabidopsis genome results into approximately 11 and 2 COs per genome and chromosome each, repectively (see main text for explanation). During meiotic cell division, DSBs that are the COs precursors could be repaired to either COs (through class I or class II pathways) or NCOs (through SDSA pathway). The class I COs is suppressed through interaction of COs supressing gene, HCR1, with PP4 phosphatase complex to limit the interfering COs that is opposed to the pro-recombination kinases activity. The class II COs are suprssed through the activity of COs repressing elements (FANCM, FIGL1, RECQ4, TOP3a, and FLIP) that result in the prohibition of COs formation via class II COs formation pathway during strand invasion or D-loop formation (Choi, 2017; Li et al. 2021).
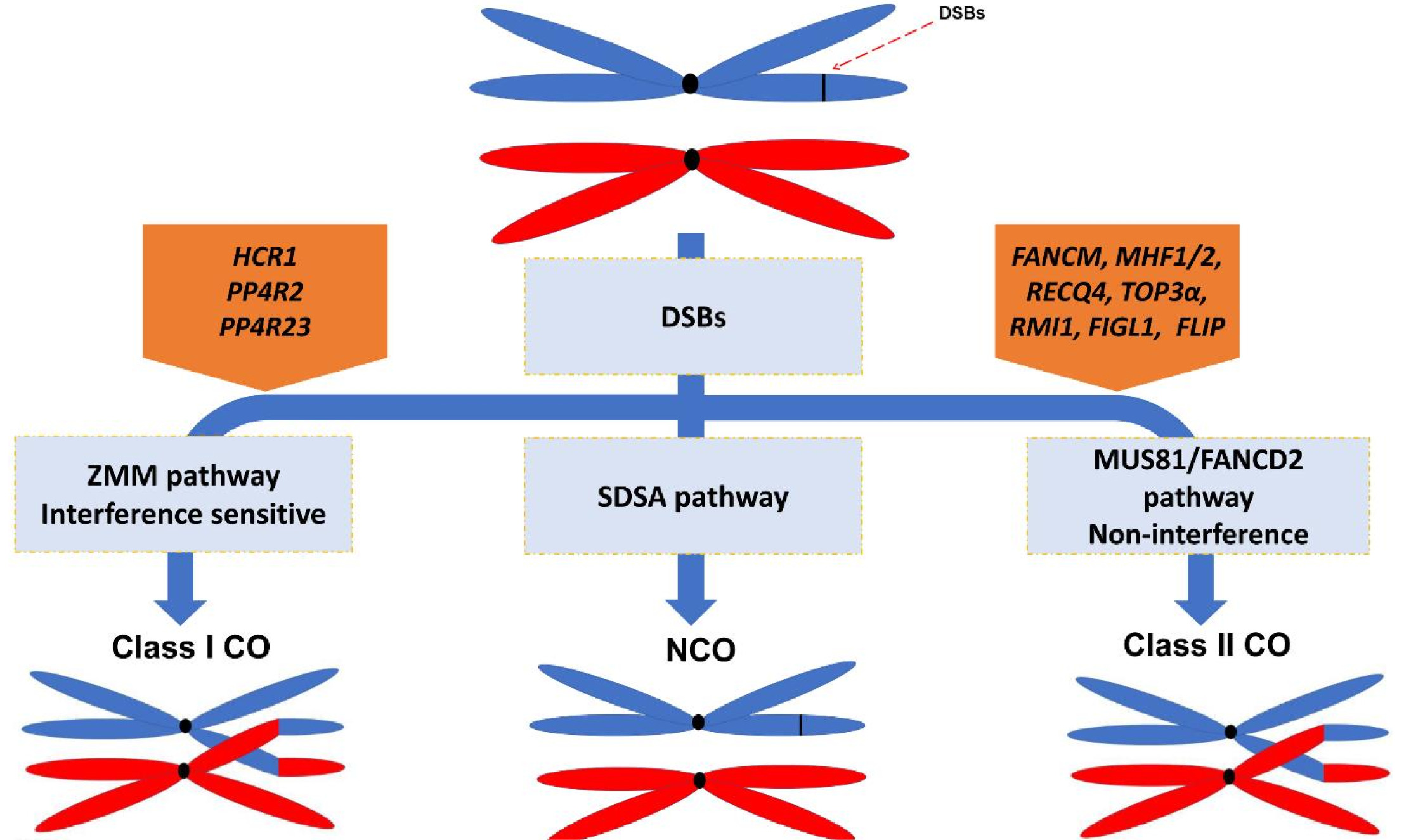
Fig. 2COs frequecny along a chromosome from distal end to centromere in Arabidopsis (blue line) and wheat/barley (red-dashed line) (Adopted from Kuo et al. 2021).
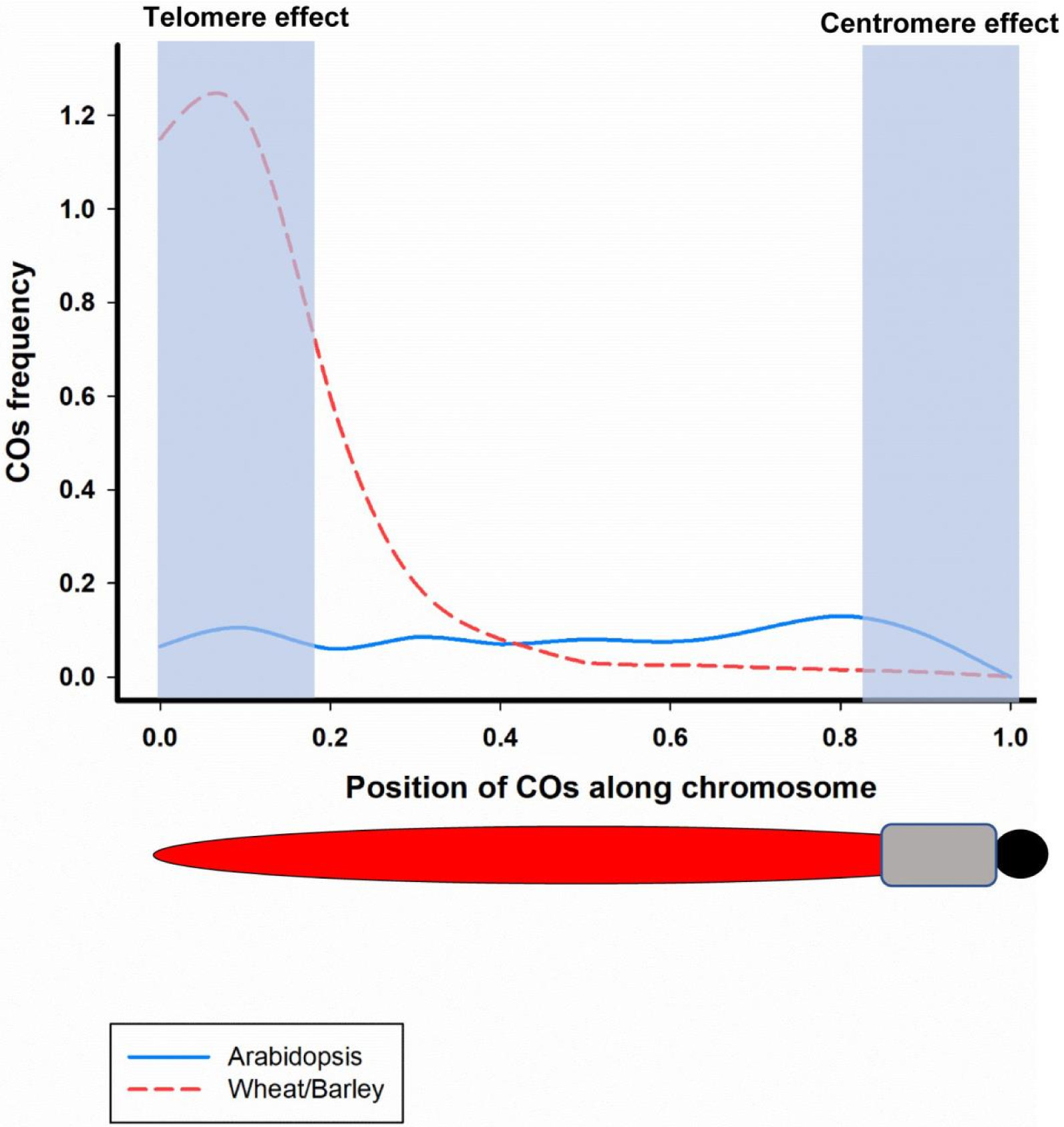
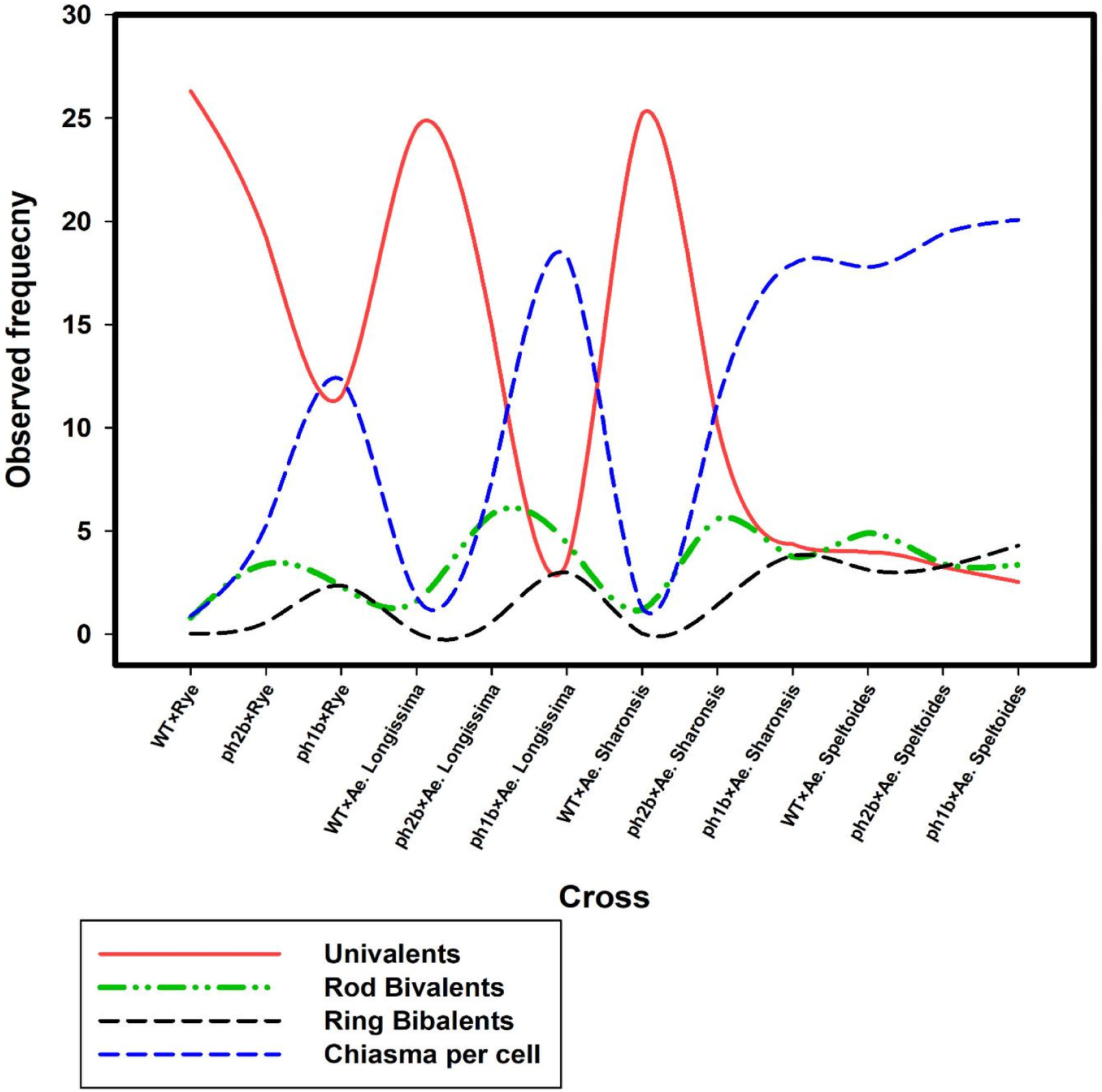
Fig. 3Homoeologous chromosomes association in metaphase I in different hybrid plants of wild-type wheat (WT) and lack of function pairing homoeologous (ph1b and ph2b mutants) mutants with wild close relatives (data adopted from Svacina et al. 2020).
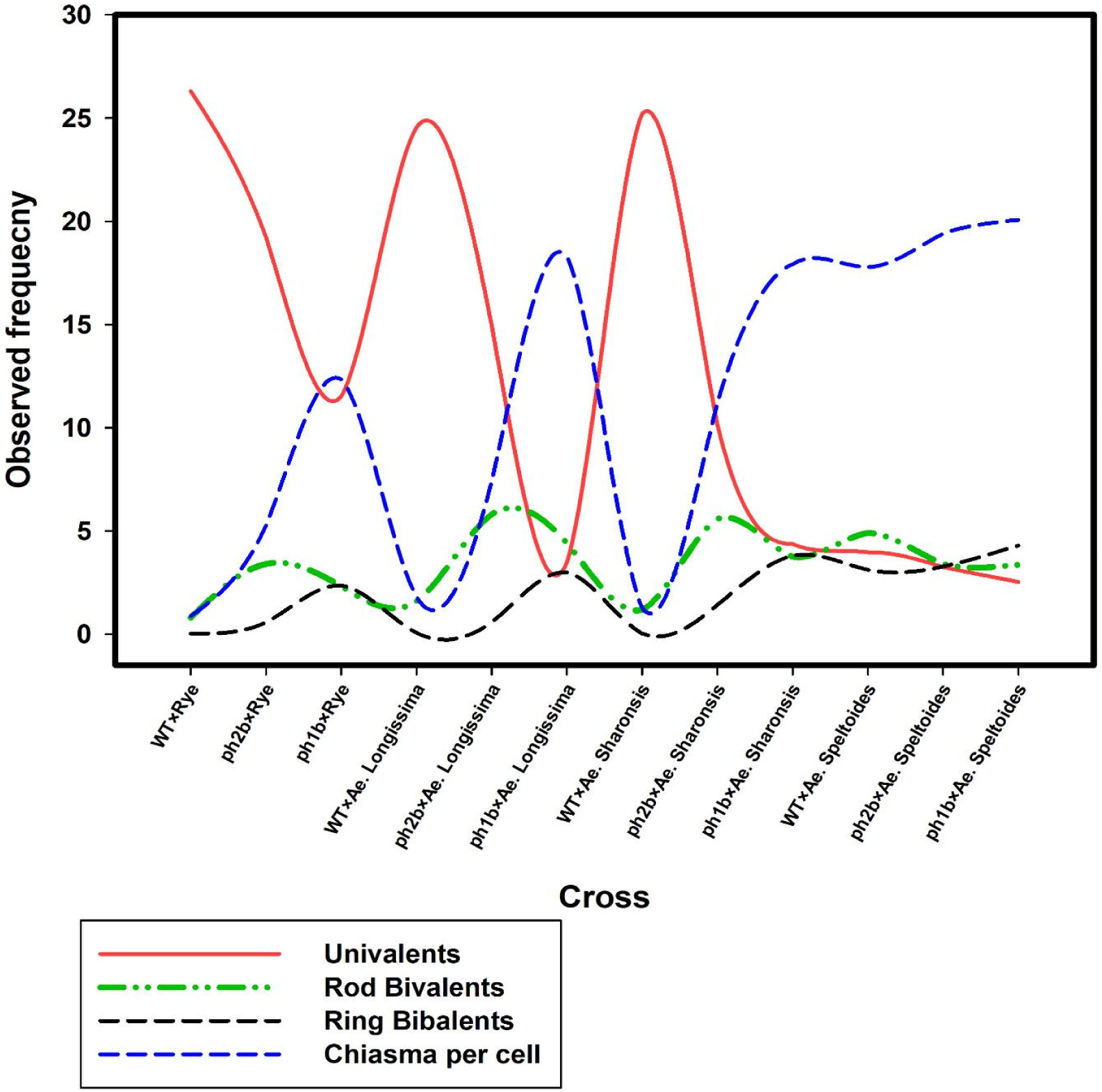
Table 1Crops orthologous genes to Arabidopsis FIGL1, RECQ4, FANCM, and CMT3 genes.
Table 1
|
Pathways/Complex |
Arabidopsis gene |
Orthologous in major crop plants |
Increased fold COs rate in mutant form |
Reference |
|
Crop species |
Gene/allele |
|
FIGL1 Helicase |
FIDGETIN-Like Protein 1 (FIGL1) |
Rice |
LOC9271031
|
2-fold increase in in both inbred and hybrid lines |
, Fernandes et al. 2018
|
|
Wheat |
AK331006
|
7.8-fold in double mutant hybrids (in combination with recq4) |
|
Corn |
LOC100193153
|
|
Tomato |
LOC101262887
|
|
Soybean |
LOC100789161, LOC100776024
|
|
RTR Complex |
RECQ4A
RECQ4B
|
Rice |
LOC_Os11g48090(A), C_Os04g35420(B)
|
3-fold in recq4 single mutant |
de Maagd et al. 2020; Serra et al. 2018
|
|
Wheat |
AK334643
|
5-fold in recq4a/recq4b double mutants hybrid context |
|
Corn |
LOC100274706
|
6-fold in recq4a/recq4b double mutants in inbred context |
|
Tomato |
LOC101260976
|
8.8-fold increase in recq4a recq4b fancm in inbred context |
|
Soybean |
LOC100800006, LOC100817867
|
4-fold increase in HEI10/recq4a/recq4b mutants |
|
Non-CG methylation |
CMT3
|
Rice |
OsCMT3a (LOC_Os10g01570), OsCMT3b (LOC_Os03g12570)
|
Increased COs in pericentromeric regions |
, Underwood et al. 2017
|
|
Wheat |
AK332918
|
|
Corn |
Zmet2 (GQ923937)
|
|
Tomato |
LOC101265056, LOC101267211
|
|
Soybean |
LOC100799480
|
|
Nuclear Core Complex (NC Complex) |
Fanconi anemia of complementation group M (FANCM)
|
Rice |
Os11g07870
|
3-fold in fancm mutant |
, Li et al. 2021; Fernandes et al. 2018; Blary et al. 2018; Girard et al. 2014; Copenhaver et al. 2012
|
|
Brassica
|
Brassica.E01856
|
8.8-fold in triple fancm/recq4a/b in inbred background |
|
Soybean |
Glyma.08G238200
|
∼2-fold in rice (Oryza sativa) and pea (Pisum sativum) hybrids |
|
Medicago truncatula
|
Medtr1g103260
|
|
Lettuce |
Lsat_1_v5_gn_8_141780.1
|
Table 2Techniques involved in enhancing COs frequency in major crops.
Table 2
|
Crop |
Technique |
Increased fold COs rate |
Reference |
|
Tomato |
Solanum lycopersicoides introgression lines and interspecific hybrids with L. pennellii hybrids
|
3 to 10-fold |
, Canady et al. 2006
|
|
Brassica napus
|
Haploid × euploid B. napus hybrids |
10 to 100-fold |
, Nicolas et al. 2007
|
|
Tomato |
MMR inhibitions (RNAi of SlMSH7, AtMSH2-DN) |
3.8 to 29.2% |
, Tam et al. 2011
|
|
Brassica napus
|
AAC hybrid with an extra chromosome 9 (C9) |
2.7-fold |
, Suay et al. 2014
|
|
Rice |
recq4 single mutant |
∼3-fold |
, Mieulet et al. 2018
|
|
Pea |
recq4 single mutant |
∼3-fold |
, Mieulet et al. 2018
|
|
Tomato |
recq4 single mutant |
∼3-fold |
, Mieulet et al. 2018
|
|
Hexaploid Wheat |
Hpp-5Mg/ph1bph1b genotypes double monosomic for T7BS.7S#3L/7B in wheat-Th. intermedium hybrids |
100-fold |
, Koo et al. 2020
|
|
Barley |
Hybridization between cultivated barley and wild barley accessions |
1.4-fold |
, Dreissig et al. 2020
|
|
Brassica napus
|
AAC allotriploid hybrids |
3.7-fold |
, Boideau et al. 2021
|
|
Hexaploid Wheat |
TaMSH7-3D loss-of-function in wheat/rye hybrids |
Up to 5.5-fold |
, Serra et al. 2021
|
|
Hexaploid wheat (newly synthetized) |
Hybrids of tetraploid Triticum turgidum and diploid Aegilops tauschii followed by WGD |
2.3-fold |
, Wan et al. 2021
|
|
Tomato |
CRISPR/Cas inactivation of RECQ4
|
1.53-fold |
de Maagd et al. 2020 |
|
Tetraploid wheat |
Virus-Induced gene Silencing (VIGS) of genes XRCC2 and FANCM
|
Up to 93% in the pericentromeric regions |
, Raz et al. 2021
|
|
Wheat |
TaMSH7-3D mutation in hexaploid wheat × Ae. variabilis hybrid |
5.5-fold |
, Soares et al. 2021
|
|
Barley |
Induced mutation in HvRECQL4
|
2-fold |
, Arrieta et al. 2021
|
References
- Adamczyk-chauvat K, Delaunay S, Vannier A, Thomas G, Eber F, et al. François C2017. Gene Introgression in Weeds Depends on Initial Gene Location in the Crop : Brassica napus - Raphanus raphanistrum Model. Genet.. 206: 1361-1372.
- Anderson LK, Lohmiller LD, Tang X, Hammond DB, Javernick L, Shearer L. 2014. Combined fluorescent and electron microscopic imaging unveils the specific properties of two classes of meiotic crossovers. Proc. Natl. Acad. Sci. U.S.A.. 111: 13415-13420.
- Arrieta M, Macaulay M, Colas I, Schreiber M, Shaw PD, Waugh R, et al. 2021. An induced mutation in HvRECQL4 increases the overall recombination and restores fertility in a barley HvMLH3 mutant background. Front. Plant Sci.. 12: 706560
- Attri R. RahmanH2018. Introgression of allelic diversity from genetically distinct variants of Brassica rapa into Brassica napus canola and inheritance of the B. rapa alleles. Crop Pasture Sci.. 69: 94-106.
- Barba-Gonzalez R, Ramanna MS, Visser RGF, Van Tuyl JM. 2005. Intergenomic recombination in F1 lily hybrids (Lilium) and its significance for genetic variation in the BC1 progenies as revealed by GISH and FISH. Genome. 48: 884-894.
- Blary A, Jenczewski E. 2019. Manipulation of crossover frequency and distribution for plant breeding. Theor. Appl. Genet.. 132: 575-592.
- Blary A, Gonzalo A, Eber F, Fourment J, et al. Bérard ABergès H2018. FANCM limits meiotic crossovers in Brassica crops. Front. Plant Sci.. 9: 368
- Bohra A, Kilian B, Sivasankar S, Caccamo M, Mccouch SR, Varshney RK. 2021. Reap the crop wild relatives for breeding future crops. Trends Biotechnol.. 40: 412-431.
- Boideau F, Pele A, Tanguy C, Trotoux G, Maillet L, Gilet M, et al. 2021. A Modified meiotic recombination in Brassica napus largely improves its breeding efficiency. Biol.. 10: 771
- Bomblies K, Higgins JD, Yant L. 2015. Meiosis evolves: adaptation to external and internal environments. New Phytol.. 208: 306-323.
- Bomblies K, Jones G, Franklin C, Zickler D, Kleckner N, Franklin C. 2016. The challenge of evolving stable polyploidy: could an increase in "crossover interference distance" play a central role? Chromosoma. 125: 287-300.
- Bozdag GO, Ono J, Denton JA, Karakoc E, Hunter N, Leu J, et al. 2021. Breaking a species barrier by enabling hybrid recombination. Curr. Biol.. 31(4): R180-R181.
- Prieto P. Calderón MCReyMMartín A2018. Homoeologous chromosomes from two Hordeum species can recognize and associate during meiosis in wheat in the presence of the Ph1 Locus. Front. Plant Sci.. 9: 585
- Canady MA, Ji Y, Chetelat RT. 2006. Homeologous recombination in Solanum lycopersicoides introgression lines of cultivated tomato. Genet.. 174: 1775-1788.
- Carlos DP, Ramirez-Parra E. 2015. Whole genome duplications in plants : an overview from Arabidopsis. J. Exp. Bot.. 66(22): 6991-7003.
- Chambers SR, Hunter N, Louis EJ, Borts RH. 1996. The mismatch repair system reduces meiotic homeologous recombination and stimulates recombination-dependent chromosome loss. Mol. Cell. Biol.. 16: 6110-6120.
- Chan AW, Villwock SS, Williams AL, Jannink J. 2022. Sexual dimorphism and the effect of wild introgressions on recombination in cassava (Manihot esculenta Crantz) breeding germplasm. G3-Genes Genome Genet.. 12: jkab372
- Chen NWG, Thareau V, Ribeiro T, Magdelenat G, Ashfield T, Innes RW, et al. 2018. Common bean subtelomeres are hot spots of recombination and favor resistance gene evolution. Front. Plant Sci.. 9: 1185
- Chen S, Nelson MN, Jenczewski E, Li Z, Mason AS, et al. Chèvre AM2011. Trigenomic bridges for Brassica improvement. Crit. Rev. Plant Sci.. 30: 524-547.
- Chester M, Gallagher JP, Symonds VV, Veruska A, Mavrodiev EV, Leitch AR, et al. 2011. Extensive chromosomal variation in a recently formed natural allopolyploid species, Tragopogon miscellus (Asteraceae). Proc. Natl. Acad. Sci. U.S.A.. 109: 1176-1181.
- Chetelat RT, Meglic V, Cisneros P. 2000. A genetic map of tomato based on BC1 Lycopersicon esculentum × Solanum lycopersicoides reveals overall synteny but suppressed recombination between these homeologous genomes. Genet.. 154: 857-867.
- Choi K, Zhao X, Tock AJ, Lambing C, Underwood CJ, Hardcastle TJ, et al. 2018. Nucleosomes and DNA methylation shape meiotic DSB frequency in Arabidopsis thaliana transposons and gene regulatory regions. Genome Res.. 28: 532-546.
- Chu Y, Bertioli D, Levinson CM, Stalker HT, Holbrook CC, Ozias-Akins P. 2021. Homoeologous recombination is recurrent in the nascent synthetic allotetraploid Arachis ipae. G3-Genes Genomes Genet.. 11(4): jkab066
- Copenhaver G, Knoll A, Girard C. 2012. FANCM limits meiotic crossovers related papers. Science. 336: 1588-1590.
- Copete-parada A, Palomino C, Cabrera A. 2021. Development and characterization of Wheat- Agropyron cristatum Introgression lines induced by gametocidal genes. Agron.. 11: 277
- Coulton A, Burridge AJ, Edwards KJ. 2020. Examining the effects of temperature on recombination in wheat. Front. Plant Sci.. 11: 230
- Danilova TV, Zhang G, Friebe B. LiuW2016. Homoeologous recombination-based transfer and molecular cytogenetic mapping of a wheat streak mosaic virus and Triticum mosaic virus resistance gene Wsm3 from Thinopyrum intermedium to wheat. Theor. Appl. Genet.. 130: 549-556.
- de Maagd RA, Loonen A, Chouaref J, Pele A, Meijer-Dekens F, Fransz P, et al. 2020. CRISPR/Cas inactivation of RECQ4 increases homeologous crossovers in an interspecific tomato hybrid. Plant Biotechnol. J.. 18: 805-813.
- Delame M, Prado E, Blanc S, Robert G, Christophe S, Mestre P, et al. 2019. Introgression reshapes recombination distribution in grapevine interspecific hybrids. Theor. Appl. Genet.. 132: 1073-1087.
- Delourme R, Foisset N, Horvais R, Barret P, Champagne G, Cheung WY, et al. 1998. Characterisation of the radish introgression carrying the Rfo restorer gene for the Ogu -INRA cytoplasmic male sterility in rapeseed (Brassica napus L.). Theor. Appl. Genet.. 97: 129-134.
- Ding M, Chen ZJ. 2018. Epigenetic perspectives on the evolution and domestication of polyploid plant and crops. Curr. Opin. Plant Biol.. 42: 37-48.
- Dreissig S, Mascher M, Heckmann S. 2019. Variation in recombination rate is shaped by domestication and environmental conditions in barley. Mol. Biol. Evol.. 36: 2029-2039.
- Dreissig S, Maurer A, Pillen K, Sharma R, Milne L, Flavell AJ. 2020. Natural variation in meiotic recombination rate shapes introgression patterns in intraspecific hybrids between wild and domesticated barley. New Phytol.. 228: 1852-1863.
- Dvorak J, Deal KR, Luo MC. 2006. Discovery and mapping of wheat Ph1 suppressors. Genet.. 174: 17-27.
- Dvorak J. 1987. Chromosomal distribution of genes in diploid Elytriga elongata that promote or suppress pairing of wheat homoeologous chromosomes. Genome. 29: 34-40.
- Fan C, Hao M, Jia Z, Neri C, Chen X, Chen W, et al. 2021. Some characteristics of crossing over in induced recom-bination between chromosomes of wheat and rye. Plant J.. 105: 1665-1676.
- Fayos I, Mieulet D, Petit J, Meunier AC, Nicolas A, Guiderdoni E. 2019. Engineering meiotic recombination pathways in rice. Plant Biotechol. J.. 17: 2062-2077.
- Ferdous M, Higgins JD, Osman K, Lambing C, Roitinger E, Franklin FCH, et al. 2012. Inter-homolog crossing-over and synapsis in Arabidopsis meiosis are dependent on the chromosome axis protein AtASY3. PLoS Genet.. 8: e1002507
- Fernandes BJ, Lloyd AH. Séguéla-arnaud MLarchevêque C2018. Unleashing meiotic crossovers in hybrid plants. Proc. Natl. Acad. Sci. U.S.A.. 115(10): 2431-2436.
- Fuentes RR, de Ridder D, van Dijk ADJ, Peters SA. 2022. Domestication shapes recombination patterns in tomato. Mol. Biol. Evol.. 39: msab287
- Girard C, Crismani W, Froger N, Mazel J, Lemhemdi A, Horlow C, et al. 2014. FANCM-associated proteins MHF1 and MHF2, but not the other Fanconi anemia factors, limit meiotic crossovers. Nucleic Acids Res.. 42: 9087-9095.
- Haenel Q, Laurentino TG, Roseti M, Berner D. 2018. Meta‐analysis of chromosome‐scale crossover rate variation in eukaryotes and its significnace to evolutionary genomics. Mol. Ecol.. 27: 2477-24.
- Hao M, Luo J, Yang M, Zhang L, Yan Z, Yuan Z. 2011. Comparison of homoeologous chromosome pairing between hybrids of wheat genotypes Chinese Spring ph1b and Kaixian-luohanmai with rye. Genome. 54: 959-964.
- Harper J, Gasior D, Mathews R, Thomas A, Evans C, King J, et al. 2018. An investigation of genotype-phenotype association in a festulolium forage grass population containing genome-spanning Festuca pratensis chromosome segments in a Lolium perenne background. PLoS ONE. 13(11): e0207412
- Henderson IR, Bomblies K. 2021. Evolution and plasticity of genome-wide meiotic recombination rates. Annu. Rev. Genet.. 55: 23-43.
- Higgins EE, Howell EC, Armstrong SJ, Parkin IAP. 2021. A major quantitative trait locus on chromosome A9, BnaPh1, controls homoeologous recombination in Brassica napus. New Phytol.. 229: 3281-3293.
- Hurgobin B, Golicz AA, Bayer PE, Chan CK, Tirnaz S, Dolatabadian A, et al. 2018. Homoeologous exchange is a major cause of gene presence/absence variation in the amphidiploid Brassica napus. Plant Biotechnol. J.. 16: 1265-1274.
- Jeridi M, Bakry F, Escoute J, Fondi E, Jeridi M, Ferchichi A. 2011. Homoeologous chromosome pairing between the A and B genomes of Musa spp. revealed by genomic in situ hybridization. Ann. Bot.. 108: 975-981.
- Jiang CX, Chee PW, Draye X, Morrell PL, Smith CW, Paterson AH. 2000. Multilocus interactions restrict gene introgression in interspecific populations of polyploid Gossypium (Cotton). Evol.. 54(3): 798-814.
- Kianian PMA, Wang M, Simons K, Ghavami F, He Y, Dukowic-schulze S, et al. 2018. High-resolution crossover mapping reveals similarities and differences of male and female recombination in maize. Nat. Commun.. 9: 2370
- King J, Armstead IP, Donnison SI, Roberts LA, Harper JA, Skøt K, et al. 2007. Comparative analyses between Lolium/Festuca introgression lines and rice reveal the major fraction of functionally annotated gene models is located in recombination-poor/very recombination-poor regions of the genome. Genet.. 177: 597-606.
- King J, Grewal S, Yang C, Hubbart S, Scholefield D, Ashling S, et al. 2017. A step change in the transfer of interspecific variation into wheat from Amblyopyrum muticum. Plant Biotechol. J.. 15: 217-226.
- Koo D, Friebe B, Gill BS. 2020. Homoeologous recombi-nation: a novel and effi cient system for broadening the genetic variability in wheat. Agron.. 10: 1059
- Loureiro J. Kopecký DHavránková MBartoš JKopecká JDoleže J2010. Physical distribution of homoeologous recombination in individual chromosomes of Festuca pratensis in Lolium multiflorum. Cytogenet. Genome Res.. 129: 162-172.
- Kumlay AM, Baenziger PS, Gill KS, Shelton DR, Graybosch RA, Lukaszewski AJ, et al. 2003. Understanding the effect of rye chromatin in bread wheat. Crop Sci.. 43: 1643-1651.
- Kuo P, Lambing C. Da ines O2021. Rewiring meiosis for crop improvement. Front. Plant Sci.. 12: 708948
- Leal-Bertioli SCM, Godoy IJ, Santos JF, Doyle JJ, Abernathy BL, et al. Guimarães PM2018. Segmental allopolyploidy in action increasing diversity through polyploid hybridi-zation and homoeologous recombination. Am. J. Bot.. 105(6): 1053-1066.
- Leflon M, Grandont L, Eber F, Huteau V, Coriton O, Chelysheva L, et al. 2010. Crossovers get a boost in brassica allotriploid and allotetraploid hybrids. Plant Cell. 22: 2253-2264.
- Li A, Geng S, Zhang L, Liu D, Mao L. 2015. Making the bread: insights from newly synthesized allohexaploid wheat. Mol. Plant.. 8: 847-859.
- Li F, Jin C, Zhang L, Wang J. 2021. Hyper-recombinant plants: an emerging field for plant breeding. Crit. Rev. Plant Sci.. 40: 446-458.
- Li H, Deal KR, Luo MC, Ji W, Distelfeld A, Dvorak J. 2017. Introgression of the Aegilops speltoides Su1-Ph1 Suppressor into Wheat. Front. Plant Sci. 8: 1-13.
- Li X, Yu M, Bolanos-Villegas P, Zhang J, Ni D, Ma H, et al. 2021. Fanconi anemia ortholog FANCM regulates meiotic crossover distribution in plants. Plant Physiol.. 186: 344-360.
- Liu C, Cao Y, Hua Y, Du G, Liu Q, Wei X, et al. 2021. Concurrent disruption of genetic interference and increase of genetic recombination frequency in hybrid. Front. Plant Sci.. 12: 757152
- Marburger S, Monnahan P, Seear PJ, Martin SH, Koch J, Paajanen P, et al. 2020. Interspecific introgression mediates adaptation to whole genome duplication. Nat. Commun.. 2019: 1-11.
- Mason AS, Nelson MN, Castello M, Yan G, Cowling WA. 2011. Genotypic effects on the frequency of homoeologous and homologous recombination in Brassica napus × B. carinata hybrids. Theor. Appl. Genet.. 122: 543-553.
- Melamed-bessudo C, Levy AA. 2012. Deficiency in DNA methylation increases meiotic crossover rates in euchromatic but not in heterochromatic regions in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 109(16). E981-E988.:
- Mieulet D, Aubert G, Bres C, Klein A, Droc G, Vieille E, et al. 2018. Unleashing meiotic crossovers in crops. Nat. Plants.. 4: 1010-1016.
- Miller DE, Hawley RS. 2017. Dispatches meiotic recombi-nation : taking the path less traveled. Curr. Biol.. 27(1): R26-R28.
- Modliszewski JL, Wang H, Albright AR, Lewis SM, Bennett AR, Huang J, et al. 2018. Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana. PLoS Genet.. 14: e1007384
- Nicolas SD, Le Mignon G, Eber F, Coriton O, Monod H, Clouet V, et al. 2007. Homeologous Recombination Plays a Major Role in Chromosome Rearrangements That Occur During Meiosis of Brassica napus Haploids. Genet.. 175: 487-503.
- Nyarko CA, Mason AS. 2021. Genetics chromosome pairing: sequence similarity or genetic control ? Trends Genet.. 38: 419-421.
- Lenormand T. OttoSP2002. Resolving the paradox of sex and recombination. Nat. Rev. Genet.. 3: 252-261.
- Ozkan H, Feldman M. 2001. Genotypic variation in tetraploid wheat affecting homoeologous pairing in hybrids with Aegilops peregrina. Genome 44. 1000-1006.:
- Pecinka A, Fang W, Levy AA, Scheid OM. RehmsmeierM2011. Polyploidization increases meiotic recombination frequency in Arabidopsis. BMC Biol. 9: 24
- Pele A, Falque M, Trotoux G, Jousseaume T, Dechaumet S, Gilet M, et al. 2017. Amplifying recombination genome-wide and reshaping crossover landscapes in Brassicas. PLoS Genet.. 13: e1006794
- Prakash S, Chopra VL. 2021. Reconstruction of allopolyploid Brassicas through non- homologous recombination : introgression of resistance to pod shatter in Brassica napus. Genet. Res. 56: 1-2.
- Raz A, Dahan-meir T, Melamed-bessudo C, Leshkowitz D, Levy AA. 2021. Redistribution of Meiotic Crossovers Along Wheat Chromosomes by Virus-induced gene silencing. Front. Plant Sci. 11: 635139
- Rehman F, Gong H, Bao Y, Zeng S, Huang H, Wang Y, et al. 2022. CRISPR gene editing of major domestication traits accelerating breeding for Solanaceae crops improvement. Plant Mol. Biol.. 108: 157-173.
- Rey M, Ram C, Mart AC. 2021. Wheat, rye, and barley genomes can associate during meiosis in newly synthesized trigeneric hybrids. Plants 10. 113.:
- Rey MD, Prieto P, Lanteri S, Vega JM. Calderón MC2015. The use of the ph1b mutant to induce recombination between the chromosomes of wheat and barley. Front. Plant Sci.. 6: 1-9.
- Hayta S, Harwood W, Shaw P, et al. Rey María-doloresMartínACSmedleyM2018. Magnesium Increases homoeologous crossover frequency during meiosis in ZIP4 (Ph1 Gene) Mutant wheat-wild relative hybrids. Front. Plant Sci.. 9: 1-12.
- Rodgers-melnick E, Bradbury PJ, Elshire RJ, Glaubitz JC, Acharya CB. 2015. Recombination in diverse maize is stable, predictable, and associated with genetic load. Proc. Natl. Acad. Sci. U.S.A.. 112: 3823-3828.
- Dorn A, Schindele P, Puchta H. Rönspies M2021. CRISPR-Cas-mediated chromosome engineering for crop impro-vement and synthetic biology. Nat. Plants.. 7: 566-573.
- Saito TT. Colaiácovo MP2017. Regulation of crossover frequency and distribution during meiotic recombination. Cold Spring Harb. Symp. Quant. Biol.. 82: 223-234.
- Schmickl R, Yant L. 2021. Adaptive introgression: how polyploidy reshapes gene flow landscapes. New Phytol.. 230: 457-461.
- Scholz M, Pendinen G. 2017. The Effect of homoeologous meiotic pairing in tetraploid Hordeum bulbosum L. × H. vulgare L. hybrids on alien introgressions in offspring. Cytogenet. Genome Res.. 150: 139-149.
- Serra H, Lambing C, Griffin CH, Topp SD, Nageswaran DC. 2018. Massive crossover elevation via combination of HEI10 and recq4a recq4b during Arabidopsis meiosis. Proc. Natl. Acad. Sci. U.S.A.. 115(10): 2-7.
- Serra H, Svacina R, Baumann U, Whitford R, Sutton T, et al. Bartoš J2021. Ph2 encodes the mismatch repair protein MSH7-3D that inhibits wheat homoeologous recombination. Nat. Commun.. 12: 41467
- Sheikh I, Sharma P, Verma SK, Kumar S. 2016. Characterization of interspecific hybrids of Triticum aestivum × Aegilops sp. without 5B chromosome for induced homoeologous pairing. J. Plant Biochem. Biotechnol.. 117: 117-120.
- Shi R, Jin J, Nifong JM, Shew D, Lewis RS. 2022. Homoeologous chromosome exchange explains the creation of a QTL affecting soil-borne pathogen resistance in tobacco. Plant Biotechnol. J.. 20: 47-58.
- Soares NR, Mollinari M, Oliveira GK, Pereira GS, Lucia M, Vieira C. 2021. Meiosis in polyploids and implications for genetic mapping: a review. Genes. 12: 1517
- Speranza P, Gaiero P, Mazzella C, Vilaro F, Jong HD. 2017. Pairing analysis and in situ Hybridisation reveal autopolyploid-like behaviour in Solanum commersonii × S. tuberosum (potato) interspecific hybrids. Euphytica. 213: 137
- Stein A, Coriton O, Rousseau-gueutin M, Samans B, Schiessl SV, Obermeier C, et al. 2017. Mapping of homoeologous chromosome exchanges influencing quantitative trait variation in Brassica napus. Plant Biotechnol. J.. 15: 1478-1489.
- Storme ND, Geelen D, Meeus S, Vallejo-marı M. 2020. Effect of Whole-Genome Duplication on the Evolutionary Rescue of Sterile Hybrid Monkeyflowers. Plant Commun.. 1: 100093
- Strelnikova SR, Krinitsina AA, Komakhin RA. 2021. Effective RNAi-mediated silencing of the mismatch repair msh2 gene induces sterility of tomato plants but not an increase in meiotic recombination. Genes. 12: 1167
- Suay L, Zhang D, Lod M, Huteau V, Coriton O, Szadkowski E, et al. 2014. Crossover rate between homologous chromosomes and interference are regulated by the addition of specific unpaired chromosomes in Brassica. New Phytol.. 201: 645-656.
- Svacina R, Sourdille P, Kopecky D, Bartos J. 2020. Chromo-some pairing in polyploid grasses. Front. Plant Sci.. 11: 1056
- Tam SM, Hays JB, Chetelat RT. 2011. Effects of suppressing the DNA mismatch repair system on homeologous recombination in tomato. Theor. Appl. Genet.. 123: 1445-1458.
- Tennessen JA, Govindarajulu R, Ashman TL, Liston A. 2014. Evolutionary origins and dynamics of octoploid strawberry linkage maps. Genome Biol. Evol.. 6(12): 3295-3313.
- Tiley GP, Burleigh JG. 2015. The relationship of recombi-nation rate, genome structure, and patterns of molecular evolution across angiosperms. BMC Evol. Biol.. 15: 194
- Underwood CJ, Choi K, Lambing C, Zhao X, Serra H. 2017. Epigenetic activation of meiotic recombination near Arabidopsis thaliana centromeres via loss of H3K9me2 and non-CG DNA methylation. Genome Res.. 28(4): 519-531.
- Verma SK, Kumar S, Sheikh I, Sharma P. 2016. Induced homoeologous pairing for transfer of useful variability for high grain Fe and Zn from Aegilops kotschyi into wheat. Plant Mol. Biol. Rep.. 34: 1083-1094.
- Wan H, Li J, Ma S, Yang F, Chai L, Liu Z, et al. 2021. Allopolyploidization increases genetic recombination in the ancestral diploid D genome during wheat evolution. Crop J.. 10(3): 743-753.
- Wang L, Cao S, Chen ZJ. 2021. Altered chromatin architecture and gene expression during polyploidization and domestication of soybean. Plant Cell 33(5). 1430-1446.:
- Wang OM, Dong ZY, Zhang ZJ, Lin XY, Shen Y, Zhou D, et al. 2005. Extensive de novo genomic variation in rice induced by introgression. Genetics. 170: 1945-1956.
- Wang Y, van Rengs WM, Waznul M, Mohd A, Underwood CJ. 2021. Meiosis in crops: from genes to genomes. J. Exp. Bot.. 72(18): 6091-6109.
- Wang Y, Copenhaver GP. 2018. Meiotic Recombination: mixing it up in plants. Annu. Rev. Plant Biol.. 69: 1-33.
- Wu Y, Lin F, Zhou Y, Wang J, Sun S, Wang B, et al. 2021. Genomic mosaicism due to homoeologous exchange generates extensive phenotypic diversity in nascent allopolyploids. Natl. Sci. Rev.. 8: nwaa277
- Xiong Z, Gaeta RT, Pires JC. 2011. Homoeologous shuffling and chromosome compensation maintain genome balance in resynthesized allopolyploid Brassica napus. Proc. Natl. Acad. Sci. U.S.A.. 108: 7908-7913.
- Xu C, Huang Q, Ge X, Li Z. Id2019. Phenotypic, cytogenetic, and molecular marker analysis of Brassica napus intro-gressants derived from an intergeneric hybridization with Orychophragmus. PLoS ONE. 14: 1-16.
- Xu J, Wang L, Deal KR, Zhu T, Ramasamy RK, Cheng M, et al. 2020. Genome ‑ wide introgression from a bread wheat × Lophopyrum elongatum amphiploid into wheat. Theor. Appl. Genet.. 133: 1227-1241.
- Yang Z, Ge X, Yang Z, Qin W, Sun G, Wang Z, et al. 2019. Extensive intraspecific gene order and gene structural variations in upland cotton cultivars. Nat. Commun.. 10: 2989
- Yelina NE, Lambing C, Hardcastle TJ, Zhao X, Santos B, Henderson IR. 2015. DNA methylation epigenetically silences crossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis. Genes Dev.. 29: 2183-2202.
- Yin L, Zhu Z, Luo X, Huang L, Li Y, Mason AS, et al. 2020. Genome-wide duplication of allotetraploid brassica napus produces novel characteristics and extensive ploidy variation in self-pollinated progeny. G3-Genes Genome Genet.. 10: 3687-3699.
- Yu X, Wang X, Hyldgaard B, Zhu Z, Zhou R, Kjær KH, et al. 2018. Allopolyploidization in Cucumis contributes to delayed leaf maturation with repression of redundant homoeologous genes. Plant J.. 94: 393-404.
- Zhang R, Hou F, Feng Y, Zhang W, Zhang M, Chen P. 2015. Characterization of a Triticum aestivum - Dasypyrum villosum T2VS - 2DL translocation line expressing a longer spike and more kernels traits. Theor. Appl. Genet.. 128: 2415-2425.
- Zhang Z, Fu T, Liu Z, Wang X, Xun H, Li G, et al. 2019. Extensive changes in gene expression and alternative splicing due to homoeologous exchange in rice segmental allopolyploids. Theor. Appl. Genet.. 132: 2295-2308.
- Zhang Z, Gou X, Xun H, Bian Y, Ma X, Li J, et al. 2020. Homoeologous exchanges occur through intragenic recom-bination generating novel transcripts and proteins in wheat and other polyploids. Proc. Natl. Acad. Sci. U.S.A.. 117: 14561-14571.
- Zhao M, Ku J, Liu B, Yang D, Yin L, Ferrell TJ, et al. 2021. The mop1 mutation affects the recombination landscape in maize. Proc. Natl. Acad. Sci. U.S.A.. 118(7): 1-8.
- Zou J, Fu D, Gong H, Qian W, Xia W, Pires JC, et al. 2011. De novo genetic variation associated with retrotransposon activation, genomic rearrangements and trait variation in a recombinant inbred line population of Brassica napus derived from interspecific hybridization with Brassica rapa. Plant J.. 68: 212-224.