Abstract
Grain yield in wheat is mostly governed prior to flowering during primordia development. Mature primordia development, floret fertility, and maximum grain number per spikelet are the three most important characteristics of spike development. The genes involved in primordia formation, on the other hand, have remained unclear. Here, we demonstrate molecular characterization of Triticum aestivum MARCH9 (TraesCS7A01G283700) which encodes RING domain, might play a positively regulating primordium and floret development until anthesis stages. TaMARCH9 expression level was highly induced at differentiation stages (WS 3-6) in Yeongkwang cultivar. TaMARCH9 exhibited ubiquitination activities and was localized to the plasma membrane using a transient expression system in tobacco. Moreover, overexpression of TaMARCH9 in Arabidopsis plant displayed early flowering phenotypes compared to wild-type. Some of flowering related genes such as FLC, MAF, FT and SOC were highly induced in TaMARCH9- overexpressing Arabidopsis plants. Our finding suggested that TaMARCH9 may act a key role in the formation of floret primordium development and/or flowering time in wheat.
-
Key words: Wheat, RING-type E3 ligase, Floret, Primordia, Flowering, Arabidopsis
INTRODUCTION
In annual crop, spikelets are typically begun during the initial reproductive phases, while primordia form inside the spikelets and determine spike fertility later in the repro-ductive phase (
Ochagavía et al. 2018). In contrast to the development of leaves and spikelets, when all initiated organs are visible in fully grown plants, floret development involves a period of floret initiation prompted by a phase of floret death (Kim and Khan,
et al. 2021). Generally, 6-12 floret primordia are formed in each spikelet; however, most degenerate before reaching the anthesis stage (
Sibony and Pinthus 1988). The proper rate of this mortality and the onset of floret mortality are most likely related to the amount of assimilates available to the developing juvenile spike (
González et al. 2003;
Guo et al. 2016). As a result of the floret's production and degeneration, the potential grain number is determined during the pre-anthesis stage (
Slafer and Savin 1994). Moreover, the most of these processes occur while the spikes are connected alongside the culm during the pre-anthesis stage; the quantity of grains is related to the number of fertile florets that emerge during stem elongation (
Ferrante et al. 2010,
2013). The number of fertile florets is usually determined by the number of remaining florets rather than the total number of florets produced (
Ferrante et al. 2013). Thus, the total number of spikelet on each spike and the number of fertile florets per spikelet typically determine the wheat grain yield potential (
Li et al. 2019).
In
Arabidopsis, the time to bolting or the rosette leaf number when the first floral bud emerges are two often utilized macroscopic markers of phase shift in reproductive phase or floral transitions (
Pouteau and Albertini 2009). However, in temperate grasses such as wheat (
Tritium aestivum) and barley (
Hordeum vulgare), most phases of reproductive development, such as blooming, occur within the leaf sheath and can be scored most efficiently when the shoot is dissected.
Waddington et al. (1983) produced a quantified index for wheat and barley development based on carpel and shoot morphogenesis. This scale is associated with the development of advanced floret primordium and/or floret. In wheat and barley, flowering is generally scored as heading, however the several stages of shoot apex development differ in their sensitivity to environmental stimuli and are influenced by unique genetic variables; hence, floral transition and flowering time may not be linked or separated (
Digel et al. 2015).
Flowering time and/or maturity are determined by an extensive network of genetic regulators in plants that res-pond to external stimuli and endogenous genes (
Kong et al. 2018). The cross-talk network between VERNALIZATOIN genes such as VRN1, VRN2, VRN3, and VRN-D4 in wheat and barley consolidates vernalization and photo-period pathway-related flowering time (
Li et al. 2020). However, in
Arabidopsis, the flowering regulatory network modulates the MADS-domain transcription factor Flowering LOCUS C (
FLC), which influences large numbers of target genes and is also known as a fundamental regulator of flowering.
In all eukaryotic cells, the ubiquitin (Ub) 26S pro-teasome system is a critical major mechanism for protein degradation (
Cho et al. 2006). This system degrades a large number of proteins in the cytoplasm and nucleus and is important in the regulation of cell metabolism such as cell differentiation, endocytosis, embryogenesis, senescence, hormone responses, and pathogen defense (
Smalle and Vierstra 2004;
Frugis and Chua 2002). In particular, the ubiquitination system required E1 (activating enzyme), E2 (conjugating enzyme), and E3 (ubiquitin ligase). An E3 ligase is an essential enzyme that recognizes and controls substrate proteins (Morreale and Walden 2016). The RING (Really Interesting New gene)-type is one of the E3 ligases, and it consists of eight zinc-coordinating C (Cys) and H (His) residues (in the order C C-X2-C-X(9–39)-C-X(1–3)-H-X(2–3)-Cys-X2-C-X(4–8)-C-X2-C (X can be any amino acid) that forms a complex two zinc atoms (
Saurin et al. 1996). Many RING E3 ligases have been found to be involved in the regulation of plant growth and develop-ment, hormone signaling, and responses to biotic and abiotic stresses (
Hwang et al. 2003;
Cho et al. 2006;
Park et al. 2018; Kim and Khan
et al. 2021; Kim and
Kim et al. 2021). Despite extensive research into the function of flowering related pathways in other plants, little known about the physiological process underlying E3 ligase, particularly in wheat.
In this study, we demonstrate the function of Wheat Membrane Associated Ring-CH-Type Finger 9, TaMARCH9, whose expression level was highly increased from differen-tiation to the end of floret initiation stages (WS3-6). The plasma membrane localized TaMARCH9 protein possessed E3 ligase activities. Moreover, transgenic TaMARCH9 in Arabidopsis plants had earlier bolting phenotype than wild type plant.
MATERIALS AND METHODS
Plant material and growth conditions
The wheat cultivar (T. aestivum L., cv. Yeongkwang) seeds were vernalized for 50 days in a dark room at 4°C. At the end of vernalization, the plants were transferred to soil pots in a greenhouse at 25°C under 16 hours/8 hours light/dark photoperiod conditions. Wheat primordia tissues at various developmental stages (WS1.5 to 6). were collected for analysis. As previously mentioned, each stage of wheat primordia and floret measurement and sampling informa-tion was clarified by Kim and Khan et al. 2021.
Gene expression studies
To investigate the relative expression level, total RNA extraction, cDNA synthesis, and quantitative Real-time PCR (qRT-PCR) were used. TRIzol
Ⓡ (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from 40 to 50 wheat primordia tissues at various developmental stages (WS1.5 to 6). First-strand cDNA was created from the total RNA (2 mg) using the Power cDNA Synthesis kit (iNtRON Biotechnology, Seongnam, Gyeonggi, Korea) following the manufacturer’s instructions. The qRT-PCR findings were examined using a CFX96 real-time PCR (Bio-Rad, Hercules, CA, USA). As an internal control,
TaActin was used.
Supplementary Table S1 contains a list of all primers used in this work.
To determine the subcellular localization of
TaMARCH9,
TaMARCH9 was cloned into green fluorescent protein (GFP)-vector (pMDC43) that containing
CaMV 35S pro-moter using LR reaction (Invitrogen, Carlsbad, CA, USA). The constructs generated with 35S:
TaMARCH9-GFP and 35S:GFP (empty) were applied to polyethylene glycol- transfected wheat protoplasts for transient experiments. Subcellular localization was performed as previously described by Kim (
Kim et al. 2021). Fluorescent signals were conducted by fluorescence microscopy (LSM 700; Carl Zeiss, Jena, Germany).
To examine the ubiquitination activities of TaMARCH9, maltose binding protein (MBP) tagged vector (pMAL-C-5X) was constructed. TaMARCH9-MBP was expressed by E. coli strain (BL21) and purified by amylose resin (NEB, Ipswich, MA, USA). TaMARCH9-MBP protein were co-incubated 3 hours in absence or presence of E1, E2, ubiquitin, and ubiquitination reaction butter [1 M Tris-HCl, pH 7.5; 40 mM ATP; 100 mM MgCl2; 40 mM dithiothreitol (DTT)) at 30°C degree. Anti-MBP (NEB, Ipswich, MA, USA) and anti-Ubi (Sigma-Aldrich, St Louis, MO, USA) were used to immunoblot the reaction products.
Overexpression of TaMARCH9 in Arabidopsis
Arabidopsis (ecotype type Columbia-0) was used to generate
TaMARCH9 overexpressing plants.
Arabidopsis flowers were soaked with
Agrobacterium tumefaciens cells (GV3101) with recombinant vectors (
TaMARCH9-GFP) suspended in 5% sucrose and 0.05 percent Silwet-L77 (Sigma-Aldrich, St Louis, MO, USA). All of the steps involved in
Arabidopsis transformation were carried out exactly as described previously (
Ho et al. 2022). For transgenic plant selection, T
1 generation seeds were allowed to grow on MS medium with 50 mg/L hygromycin. Independent T
3 overexpression lines (OE1, OE2) were used in this study.
RESULT
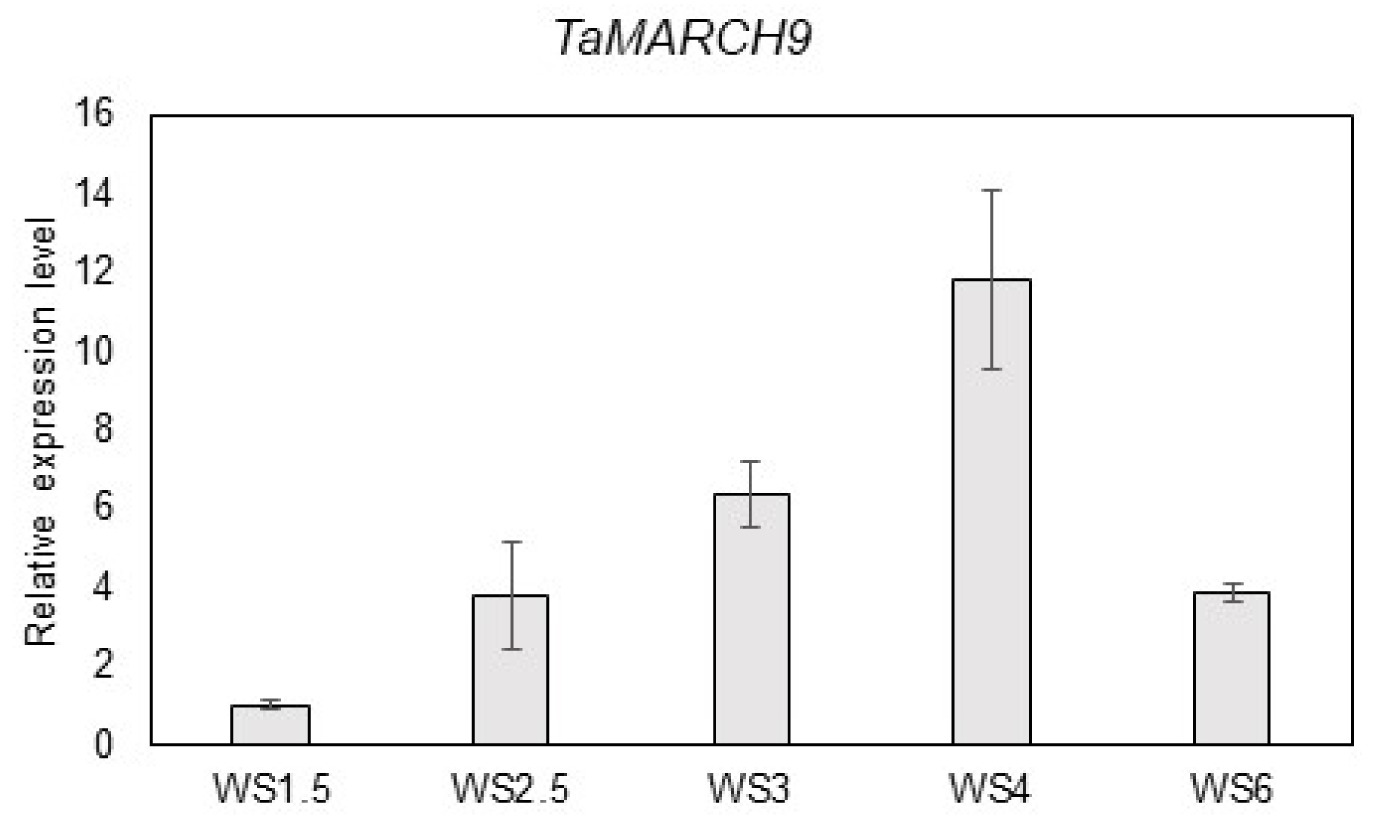
Expression analysis of TaMARCH9 at floret development stages in wheat
In previous study, the expression level of 34 wheat RING type E3 ligase protein that were selected based on domain analysis were investigated under floret primordia development stages (Kim and Khan
et al. 2021). Of these, we verified the transcription pattern of 34 wheat RING type E3 ligase under primordia developmental stages. Among them, a transcript level of
TraesCS7A02G283700 (
TaMARCH9) was highly induced at reproductive development phases. After the initiation of reproductive phase,
TaMARCH9 was up-regulated from WS 3 to WS 6 (about 4 to 12-fold) (
Fig. 1).
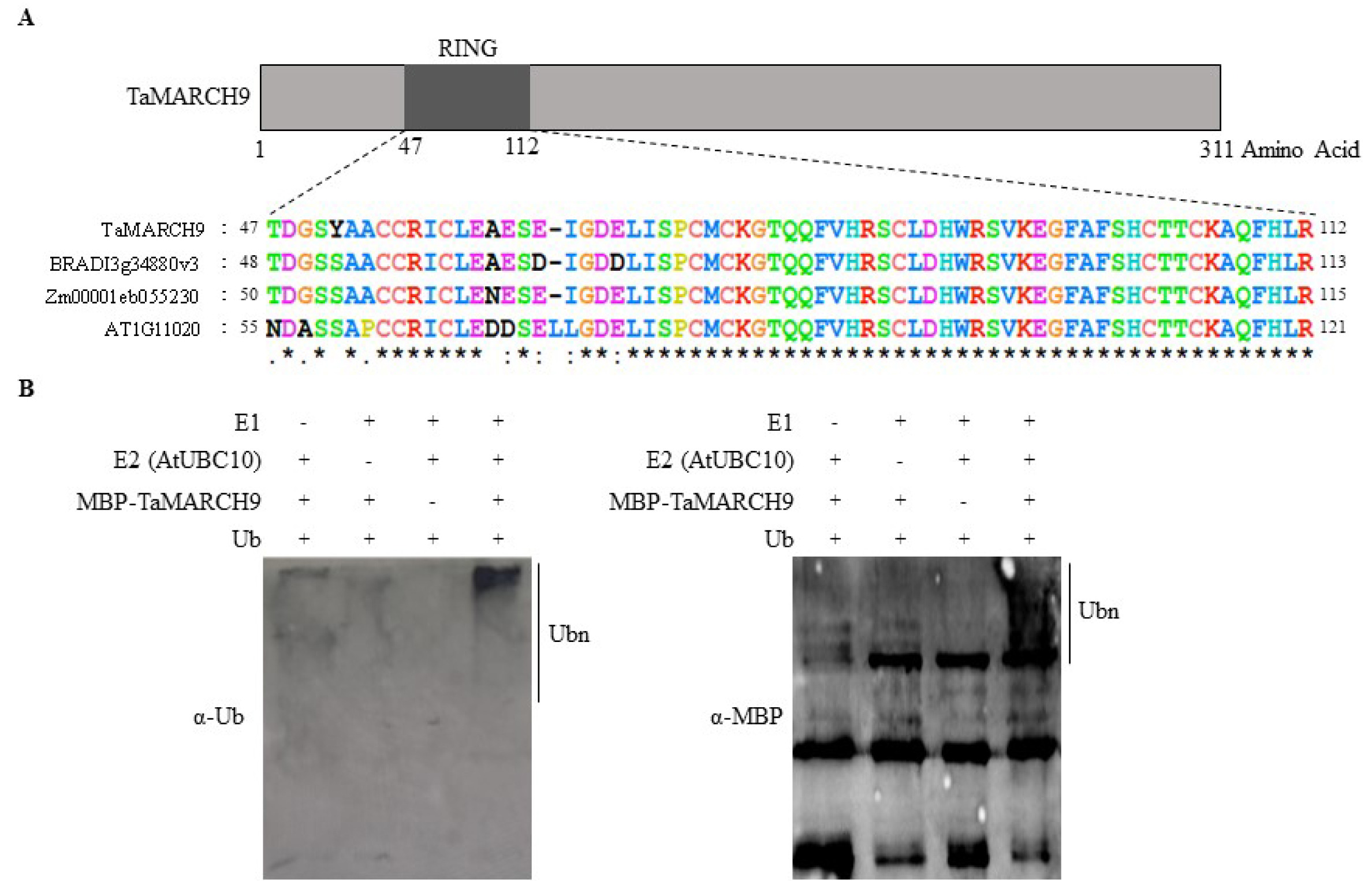
To characterize the
TaMARCH9, we designed gene specific primers based on
Triticum aestivum L. in the NCBI sequence database to isolate the
TaMARCH9 (
Supplementary Table 1). Se-quence analysis revealed that the full-length of
TaMARCH9 protein encodes 311 amino acid residues, which contains RING-CH-type- domain (
Fig. 2A). Multiple sequence align-ment indicated that
TaMARCH9 had a high identity with
Brachypodium distachyon MARCH9 (BRADI_ 3g34880v3) (92%) over the protein size as well as a similar level of identity with
Zea mays RING type E3 ligase protein (Zm00001eb055230) (87%) (
Supplementary Fig. 1A).
To determine the ubiquitination activities of
TaMARCH9, we constructed MBP-tagged
TaMARCH9 recombinant pro-tein. Ubiquitination chains were observed in the presence of all of these components with MBP-tagged
TaMARCH9, according to immunoblot analysis using both anti-Ub and anti-MBP, respectively (
Fig. 2B).
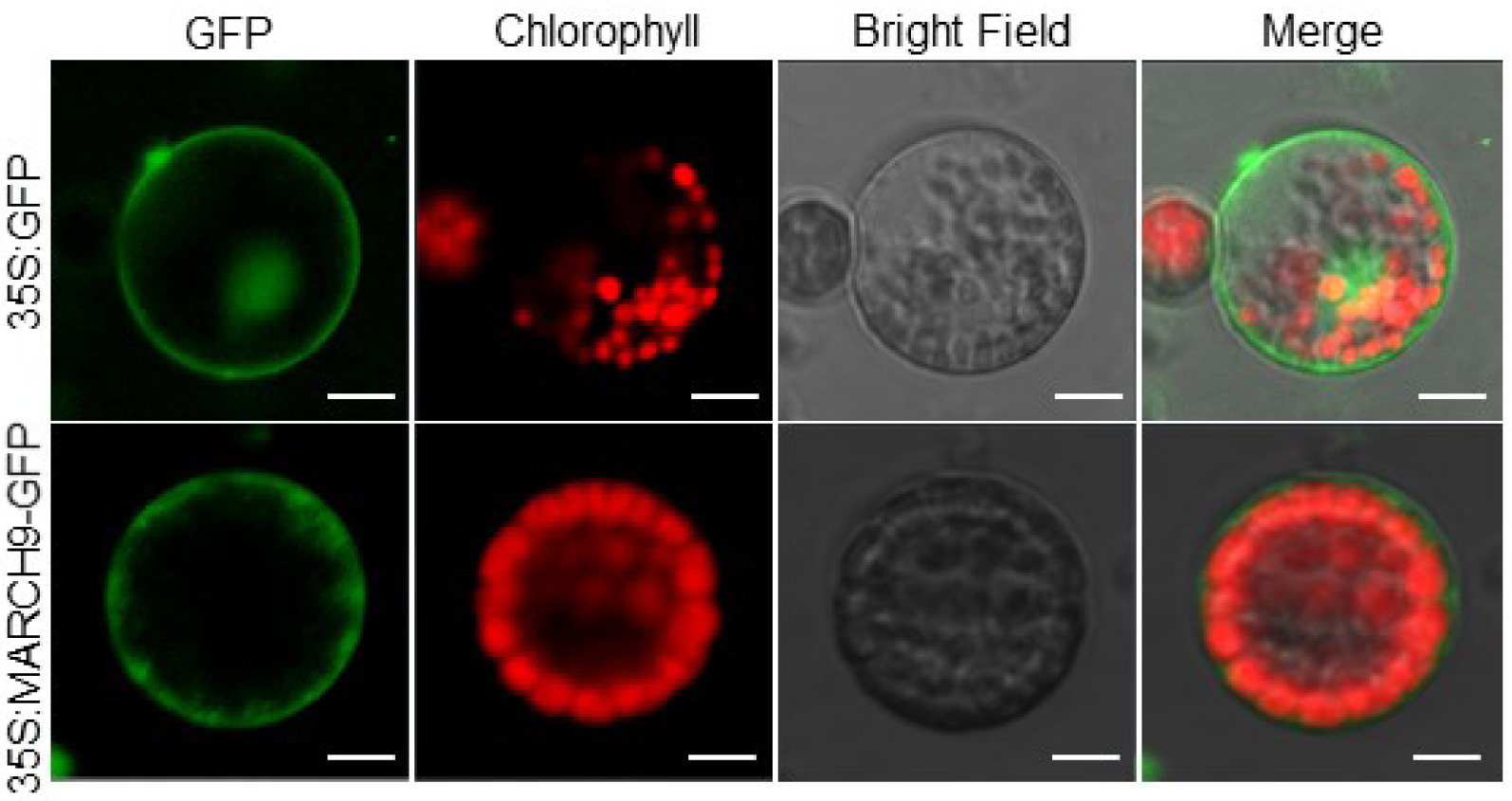
In order to explore the transient expression of
TaMARCH9 protein, the full length
TaMARCH9 gene was fused into it with GFP to generate 35S:
TaMARCH9-GFP. In wheat protoplast, the GFP tagged
TaMARCH9 protein was tran-siently expressed using transfection methods.
TaMARCH9 was mainly observed on plasma-membrane, whereas the empty GFP protein was appeared in the nucleus as well as plasma-membrane (
Fig. 3).
Since
TaMARCH9 transcription level was shown to be considerably increased during the differentiation stage (WS 3 to 6), we predicted that
TaMARCH9 would be connected to flowering time. Moreover, the cauliflower mosaic virus 35S constitutive promoter (CaMV35S) is a foundational promoter that could be found in a variety of cell types, tissues, and organs in plants (
Li et al. 2011). To examine the phenotype of
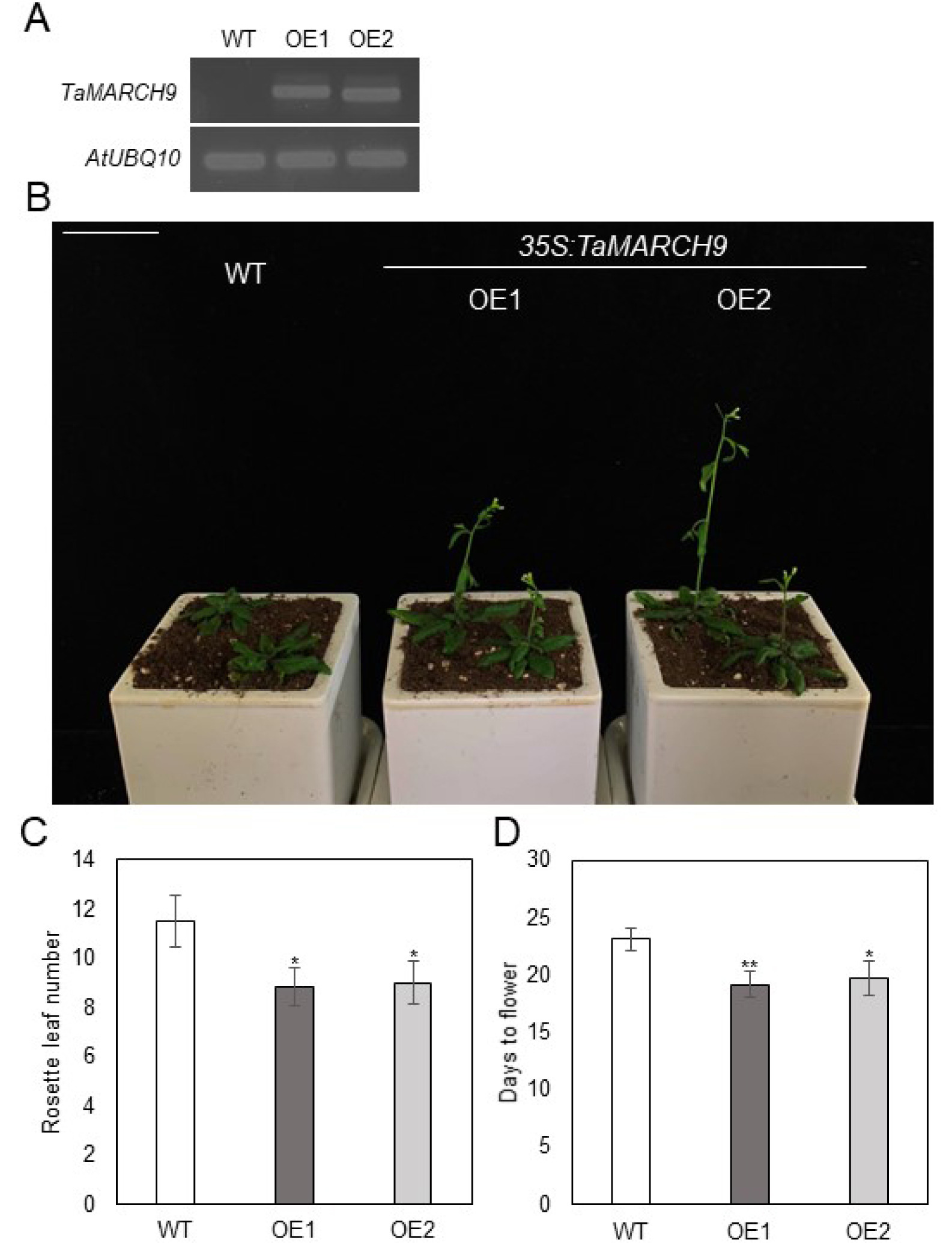
TaMARCH9, CaMV35S was used to create transgenic
Arabidopsis plants that overex-pressed
TaMARCH9. Two independent T
3 transgenic lines with considerably enhanced level of
TaMARCH9 tran-script level were chosen for further investigation (
Fig. 4A). During our investigation for phenotypic difference, we found that under the normal growth condition (16 hours light/8 hours dark), the
TaMARCH9 overexpression lines (OE1, 2) displayed early flowering phenotype (
Fig. 4B). In this regard, we also recorded rosette leaf number and flowering time to determine the impact of
TaMARCH9 overexpression on phase transitions in
Arabidopsis. Com-pare to wild type (WT),
TaMARCH9 OE1, 2 showed decreased rosette leaf number and reduced time to flowering (
Fig. 4B, C).
TaMARCH9 OE was therefore able to accelerate
Arabidopsis flowering.
Fig. 4 shows the phenotypic differences in flowering time between the WT and
TaMARCH9 OE in
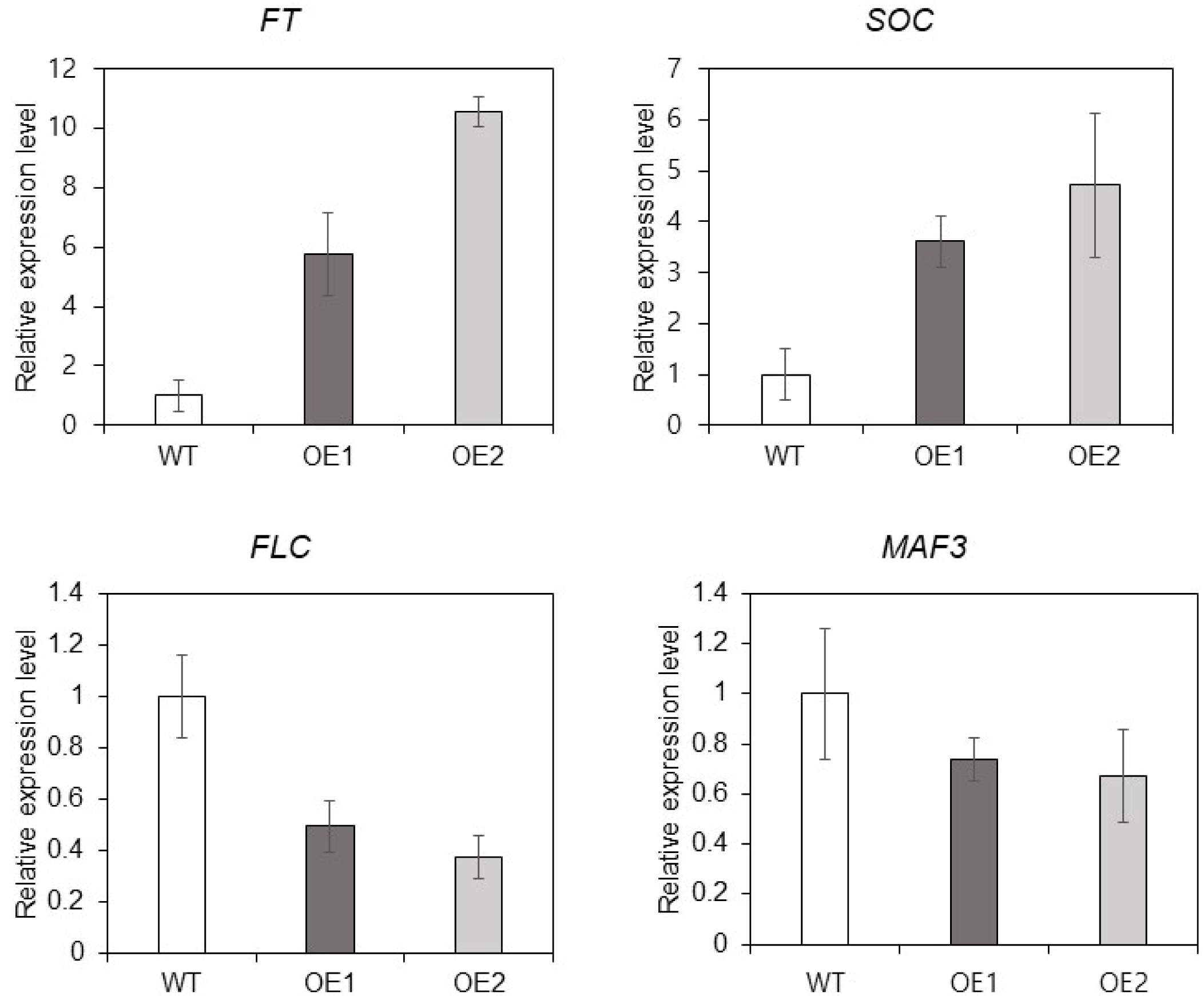
Arabidopsis. As rosette leaf number and bolting time was significantly reduced in OE lines, we hypothesized that
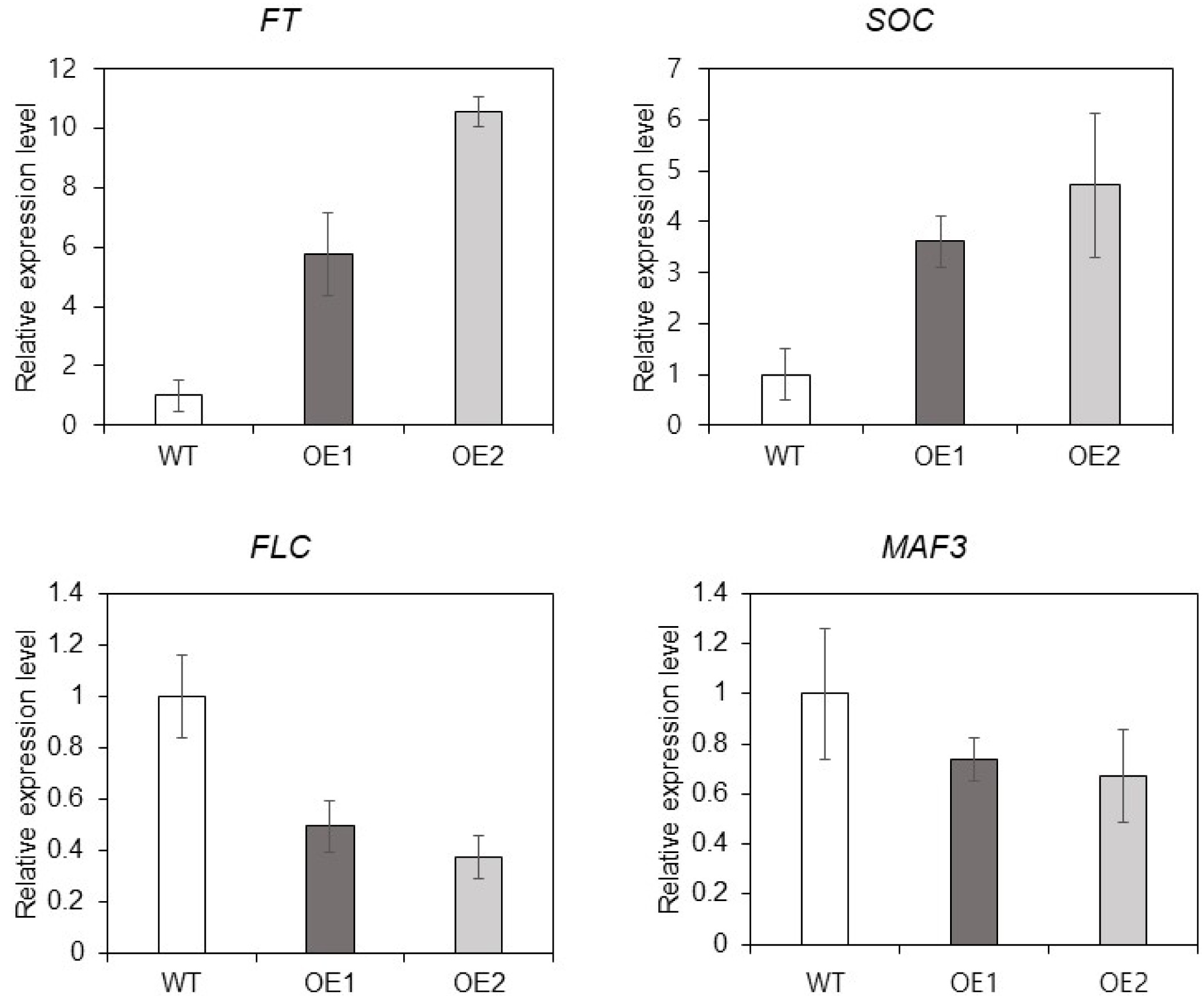
TaMARCH9 might be explained by autonomous pathways. To check the expression of flowering-time induced genes, we confirmed some of floral accelerate genes (
FT and
SOC) and repressor genes (
FLC and
MAF3). The transcript levels of floral accelerator genes (
FT and
SOC), was considerably increased (about 3 to 10-fold) in the OE lines (
Fig. 5). On the other hands, the expression of floral repressors genes (
FLC and
MAF3) was reduced in OE lines (
Fig. 5). These findings showed that
TaMARCH9 was implicated in the transcrip-tion levels of flowering time-related genes in response to flowering.
DISCUSSION
Flowering time and shorter periods of vegetative stages might become particularly significant for wheat produc-tivity in condition of abiotic stress since they can reduce harsh conditions during the sensitive flowering and post- anthesis grain filling periods (
Shavrukov et al. 2017). As a result, early flowering is a useful method for producing improved wheat cultivars.
Previous studies verified that different types of E3 ligases were highly induced at floral transition period (Kim and Khan
et al. 2021;
Ho et al. 2022). Similarly, our finding reveals that the
TaMARCH9 was highly up-regulated at reproductive stages, causing it to be involved in flowering time in wheat. During its life cycle,
Arabidopsis passes through at least two phase transitions: vegetative and reproductive (
Bäurle and Dean 2006). The vegetative phase transition is characterized by a shift from the juvenile to the adult vegetative stage, which is generally recognized by the production of leaves with abaxial trichomes; in contrast, the substantial vegetative to reproductive floral transition, is recognized by bolting, flowering, and seed set for the next generation (
Bäurle and Dean 2006;
Li et al. 2011).
Endogenous regulator that are independent of the photoperiod, temperature, and gibberellin pathways are referred to as the autonomous pathway for regulating flowering. All of the genes in the autonomous pathway suppress
FLC expression.
FLC is a core floral repressor that prevents the induction of flowering during vegetative growth by blocking the expression of floral activators such as
FT and SUPPRESSOR OF OVEREXPRESSION OF CO1 (
SOC1) (
Srikanth and Schmid 2011). The inhibition of
FLC promotes activation of
FT and
SOC expression level (
Michaels et al. 2005). In addition, the additional members of the
FLC gene family, denoted
MAFs (
MAF1-5), can operate as floral repressors (
Ratcliffe et al. 2001). Similarly, early flowering phenotype
TaMARCH9 showed that the mRNA level of
FLC and the transcription level of
FT and
SOC were reversed (
Fig. 5).
Various type of E3 ligases, which contains RING, U-box, SCF complex domain, regulate plant external and internal stimuli (
Hwang et al. 2003;
Li et al. 2012;
Ho et al. 2022). Moreover, E3 ligase encoded proteins such as RING, U-box, F-box type has been linked to the regulation of reproductive phase development, including flowering time, maturity, reproductive organ development (
Cao et al. 2008;
Li et al. 2012;
AN et al. 2017;
Hong et al. 2020). This study demonstrates that
TaMARCH9 protein encoded RING domain called RING-CH-type at its N-terminal side (
Fig. 2A). The
TaMARCH9 exhibited ubiquitination chains when combined with E1 and E2 enzymes and also was localized in plasma-membrane (
Fig. 2B,
3). Plasma-mem-brane in plant cells serves key functions in response to transporting solution and sensory transductions (
Sussman and Harper 1989).
Supplemental Material
ACKNOWLEDGEMENTS
This work was supported by a grant from the Korea University and Basic Science Research Program through the National Research Foundation of Korea, which is funded by the Ministry of Science, ICT & Future Planning (grant number 2017R1D1A1B06030349).
Fig. 1Expression pattern of TaMARCH9 was regulated different stage of primordia development. (A) TaMARCH9 transcripts quantified by qRT-PCR in wheat primordia samples (WS: Waddington scale). Each bar represents the mean ± SD of three inde-pendent experiments.
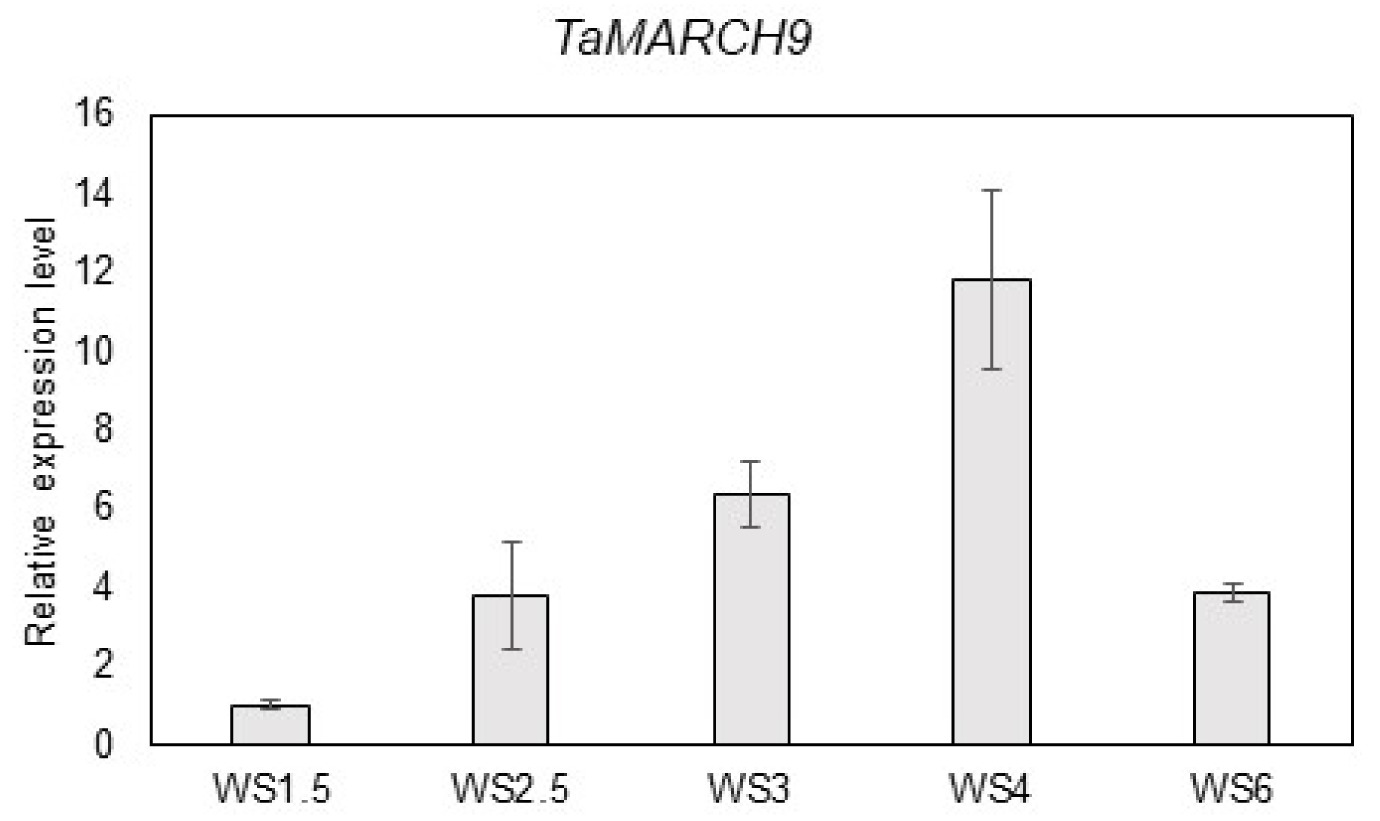
Fig. 2Domain analysis in TaMARCH9 and ubiquitination assay. (A) Schematic diagram of full-length TaMARCH9 protein. The box depicts the coding region and the solid lines represent the 5’- and 3’-untranslated regions. (B) In vitro ubiquitination assay with TaMARCH9. The ubiquitination reaction mixture contained E1 (Human), E2 (Arabidopsis), maltose-labeled protein TaMARCH9 (MBP-TaMARCH9), ubiquitin, and ATP. Polyubiquitin chains were visualized via immunoblotting using an anti-ubiquitin and anti-MBP antibody.
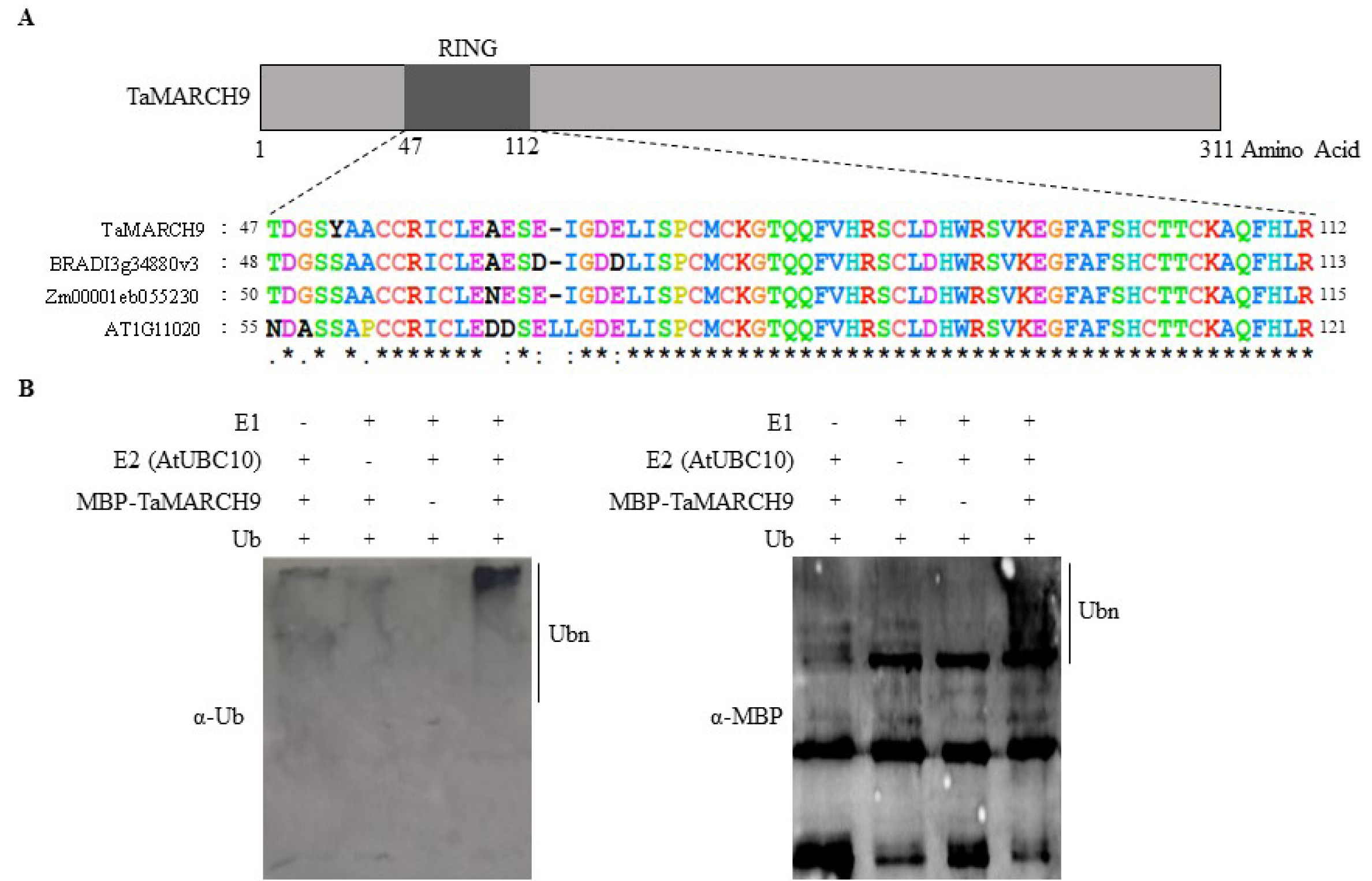
Fig. 3Subcellular localization of TaMARCH9. Subcellular localization of 35S:GFP-TaMARCH9 in wheat pro-toplasts. Bars = 10 mm. The fluorescence signal was detected using a confocal laser-scanning micro-scope.
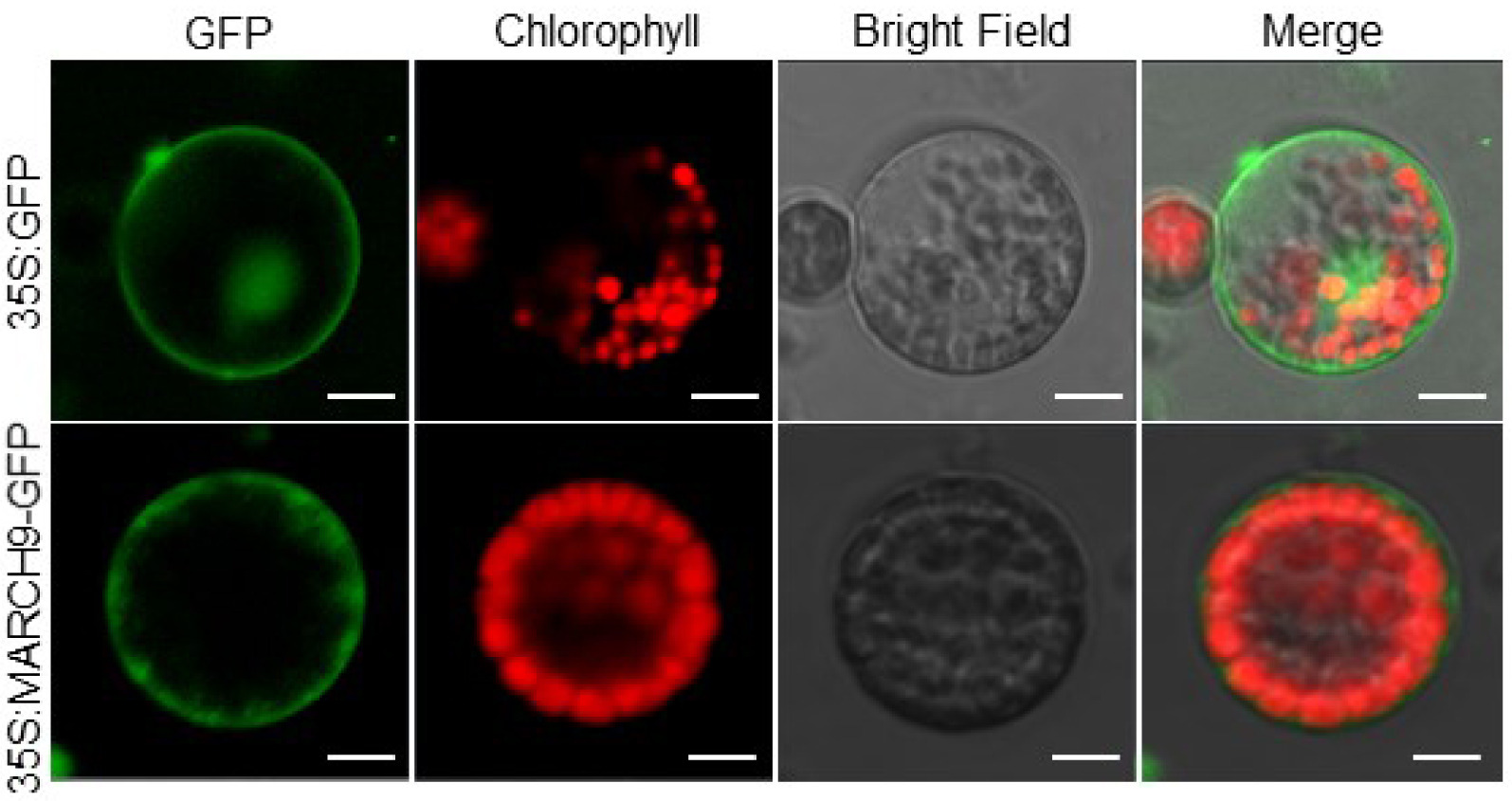
Fig. 4Early flowering phenotype of TaMARCH9 in Arabidopsis. (A) Semi-quantitative RT-PCR of TaMARCH9 overexpressing Arabidopsis (B) Flower-ing phenotype of the transgenic plants overex-pressed TaMARCH9. The WT and 35S:TaMARCH9- GFP (OE) plants in a chamber (LD, 16 hours light/8 hours dark photoperiod at 22°C/18°C). Bars = 3.5 cm. Rosette leaf number at bolting (C) Days of opening of first flower (D) in the WT and OE line in plants. Data represent the mean ± SD of ten biological replicates, and asterisks re-present significant differences (*P value < 0.05, and **P value < 0.01 Student’s t-test).
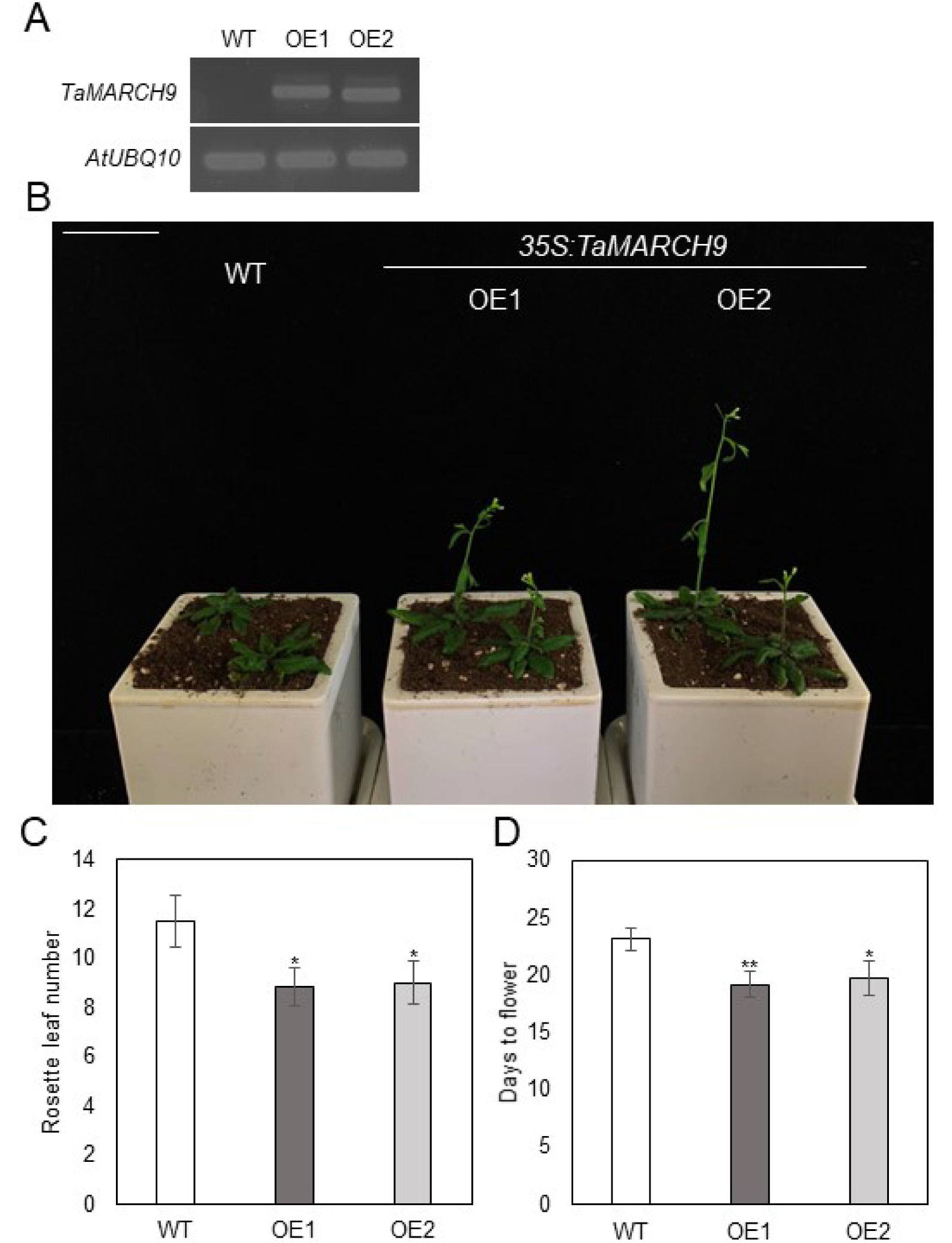
Fig. 5Expression analysis of TaMARCH9 in Arabidopsis. Expression pattern of predicted downstream genes in the WT and OE1, 2 plants.
References
- AN J, LIU X, SONG L, YOU C, WANG X, HAO Y. 2017. Functional Characterization of the Apple RING E3 Ligase MdMIEL1 in Transgenic Arabidopsis. Hortic. Plant J.. 3(2): 53-59.
- Dean C. Bäurle I2006. The Timing of Developmental Transitions in Plants. Cell. 125(4): 655-664.
- Cao Y, Dai Y, Cui S, Ma L. 2008. Histone H2B monoubiquiti-nation in the chromatin of Flowering Locus C regulates flowering time in Arabidopsis. Plant Cell. 20(10): 2586-2602.
- Cho SK, Chung HS, Ryu MY, Park MJ, Lee MM, Bahk YY, et al. 2006. Heterologous expression and molecular and cellular characterization of CaPUB1 encoding a hot pepper U-box E3 ubiquitin ligase homolog. Plant Physiol.. 142(4): 1664-1682.
- Digel B, Pankin A, von Korff M. 2015. Global transcriptome profiling of developing leaf and shoot apices reveals dis-tinct genetic and environmental control of floral transi-tion and inflorescence development in barley. Plant Cell. 27(9): 2318-2334.
- Ferrante A, Savin R, Slafer GA. 2010. Floret development of durum wheat in response to nitrogen availability. J. Exp. Bot.. 61(15): 4351-4359.
- Ferrante A, Savin R, Slafer GA. 2013. Is floret primordia death triggered by floret development in durum wheat? J. Exp. Bot.. 64(10): 2859-2869.
- Frugis G, Chua NH. 2002. Ubiquitin-mediated proteolysis in plant hormone signal transduction. Trends Cell Biol.. 12(7): 308-311.
- Slafer GA, Miralles DJ. González FG2003. Grain and floret number in response to photoperiod during stem elonga-tion in fully and slightly vernalized wheats. Field Crops Res.. 81(1): 17-27.
- Guo Z, Slafer GA, Schnurbusch T. 2016. Genotypic variation in spike fertility traits and ovary size as determinants of floret and grain survival rate in wheat. J. Exp. Bot.. 67(14): 4221-4230.
- Ho J, Woo K, Jung J, Seok M, Chan K, Ko S, et al. 2022. Molecular characterization of wheat floret development- related F-box protein (TaF-box2): Possible involvement in regulation of Arabidopsis flowering. Physiol. Plant.. 174(2): e13677
- Hong MJ, Kim JB, Seo YW, Kim DY. 2020. F-box genes in the wheat genome and expression profiling in wheat at different developmental stages. Genes. 11(10): 1-16.
- Hwang WW, Venkatasubrahmanyam S, Ianculescu AG, Tong A, Boone C, Madhani HD. 2003. A conserved RING finger protein required for histone H2B monoubiquitination and cell size control. Mol. Cell. 11(1): 261-266.
- Kim JH, Khan IU, Kim MS, Seo YW. 2021. Functional characterization of wheat histone H2B monoubiquitination enzyme TaHUB2 in response to vernalization in Keumkang (Triticum aestivum L.). J. Plant Physiol.. 16(1): 93-103.
- Kim JH, Khan IU, Lee CW, Kim DY, Jang CS, Lim SD, et al. 2021. Identification and analysis of a differentially expressed wheat RING-type E3 ligase in spike primordia development during post-vernalization. Plant Cell Rep.. 40(3): 543-558.
- Kim JH, Kim MS, Prasad D, Jung WJ, Seo YW. 2021. Molecular characterization of the wheat putative proline- rich protein TaELF7 and its involvement in the negative regulation of Arabidopsis flowering. J. Plant Physiol.. 262: 153439
- Kong L, Lu S, Wang Y, Fang C, Wang F, Nan H, et al. 2018. Quantitative trait locus mapping of flowering time and maturity in soybean using next-generation sequencing- based analysis. Front. Plant Sci.. 9: 995
- Li C, Lin H, Chen A, Lau M, Jernstedt J, Dubcovsky J. 2019. Wheat VRN1, FUL2 and FUL3 play critical and redun-dant roles in spikelet development and spike determinacy. Development. 146(14): dev175398
- Li W, Ahn IP, Ning Y, Park CH, Zeng L, Whitehill JGA, et al. 2012. The U-Box/ARM E3 ligase PUB13 regulates cell death, defense, and flowering time in Arabidops. Plant Physiol.. 159(1): 239-250.
- Li W, Wang Z, Li J, Yang H, Cui S, Wang X, et al. 2011. Overexpression of AtBMI1c, a polycomb group protein gene, accelerates flowering in arabidopsis. PLoS One.. 6(6): e21364
- Li Y, Xiong H, Guo H, Zhou C, Xie Y, Zhao L, et al. 2020. Identification of the vernalization gene VRN-B1 res-ponsible for heading date variation by QTL mapping using a RIL population in wheat. BMC Plant Biol.. 20(1): 1-15.
- Michaels SD, Himelblau E, Sang YK, Schomburg FM, Amasino RM. 2005. Integration of flowering signals in Winter-annual Arabidopsis. Plant Physiol.. 137(1): 149-156.
- Prieto P, Savin R, Griffiths S, Slafer G. Ochagavía H2018. Dynamics of leaf and spikelet primordia initiation in wheat as affected by Ppd-1a alleles under field conditions. J. Exp. Bot.. 69(10): 2921-2631.
- Park YC, Chapagain S, Jang CS. 2018. A Negative Regulator in Response to Salinity in Rice: Oryza sativa Salt-, ABA- and Drought-Induced RING Finger Protein 1 (OsSADR1). Plant Cell Physiol.. 59(3): 575-589.
- Pouteau S, Albertini C. 2009. The significance of bolting and floral transitions as indicators of reproductive phase change in Arabidopsis. J. Exp. Bot.. 60(12): 3367-3377.
- Ratcliffe OJ, Nadzan GC, Reuber TL, Riechmann JL. 2001. Regulation of flowering in Arabidopsis by an FLC homologue. Plant Physiol.. 126(1): 122-132.
- Saurin AJ, Borden KLB, Boddy MN, Freemont PS. 1996. Does this have a familiar RING? Trends Biochem. Sci.. 21(6): 208-214.
- Shavrukov Y, Kurishbayev A, Jatayev S, Shvidchenko V, Zotova L, Koekemoer F, et al. 2017. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant. 8: 1950
- Sibony M, Pinthus MJ. 1988. Floret initiation and develop-ment in spring wheat (Triticum aestivum L.). Ann. Bot.. 61(4): 473-479.
- Slafer GA, Savin R. 1994. Source-sink relationships and grain mass at different positions within the spike in wheat. Field Crops Res.. 37(1): 39-49.
- Smalle J, Vierstra RD. 2004. the Ubiquitin 26S Proteasome Proteolytic Pathway. Annu. Rev. Plant Biol.. 55(1): 555-590.
- Srikanth A, Schmid M. 2011. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci.. 68(12): 2013-2037.
- Sussman MR, Harper JF. 1989. Molecular biology of the plasma membrane of higher plants. The Plant cell. 1(10): 953
- Waddington SR, Cartwright PM, Wall PC. 1983. A quantitative scale of spike initial and pistil development in barley and wheat. Ann. Bot.. 51(1): 119-130.