Abstract
Squash (Cucurbita moschata D.) is an economically important vegetable of the Cucurbitaceae family. The genetic purity of commercial hybrid seed is crucial for the success of hybrid seed production. The molecular markers like single nucleotide polymorphism (SNP) can efficiently and cost-effectively distinguish the genetic differences among F1 hybrid cultivars. Therefore, in this study, we used ‘Fluidigm SNP Genotyping’ assay using 27 SNPs to distinguish and purity analysis of registered commercial F1 hybrid cultivars and F1 breeding lines of squash. Of these, eight SNP markers, including CMo-A01, CMo-A02, CMo-A04, CMo-A05, CMo-A12, CMo-A16, CMo-A20 and CMo-A25 can successfully identified heterozygotes from the registered commercial F1 hybrid squash cultivars with 100% accuracy and partial contamination was detected for F1 hybrid squash breeding lines which resulted due to outcrossing. Moreover, the HRM analysis of a registered commercial F1 hybrid cultivar ‘Parangsae’ with CMo-A03 SNP marker showed 96.30-100% purity of the cultivar. Our results suggest that the ‘Fluidigm SNP Genotyping’ technology could be a rapid and cost-effective method for cultivar differentiation and genetic purity analysis of F1 hybrids and squash cultivars.
-
Key words: Molecular marker, Genetic purity test, Squash, SNP, Fluidigm assay, High-throughput genotyping, F1 hybrid, Cultivar
INTRODUCTION
Squash (
Cucurbita moschata D., 2
n = 2× = 40) is an important vegetable crop of the Cucurbitaceae family. As per archaeological remains, this crop was domesticated from southern Mexico and coastal Peru (Feriol
et al. 2004;
Jacobo-Valenzuela et al. 2011). It is an essential component in human diet in different parts of the world. In Korea, squash ranked second position in the area cultivated under Cucurbitaceous crops (
Kim et al. 2016). The fruit pulp and seed are used for human consumption (
Gomes et al. 2020;
Mashitoa et al. 2021). The fruit pulp of squash is the richest source of carotenoids, lutein, vitamin C, dietary fiber, phenolic compounds and minerals, including K, Ca, P, Mg and Cu (
Jacobo-Valenzuela et al. 2011). Besides, it has various culinary uses as a food ingredient in different preparations, including breads, pasties, soups etc. (
Guiné et al. 2012). Moreover, squash has many health promoting properties, including anti-diabetic, anti-carcinogenic, and anti-microbial activities (
Yadav et al. 2010;
Kim et al. 2016).
The genetic purity assessment of F
1 hybrid seed is essential for successful adoption of hybrid seed technology (
Nandakumar et al. 2004). There are many commercial F
1 hybrid squash cultivars available in the seed market of the world including Korea. Therefore, commercial seed producers need genetic purity tests to ensure quality seeds. Traditionally, grow-out-trials (GOTs) are carried out to test the seed genetic purity, which time consuming and labor expressive while molecular markers can potentially dis-criminate the genetic variation among F
1 hybrid cultivars with a low cost (
Yashitola et al. 2002;
Ye et al. 2013;
Pattanaik et al. 2018). Recently, molecular markers have already been proven as an effective approach for detection of genetic variations between F
1 commercial cultivars (
Jamali et al. 2019;
Kim et al. 2021). For example, simple sequence repeat (SSR) markers are successfully used for differentiation of F
1 cultivars in several agricultural crops, including onion (
Tsukazaki et al. 2006), asparagus (
Caruso et al. 2008), watermelon (
Kwon et al. 2010), radish (Lee
et al. 2015), pear (
Pierantoni et al. 2004), melon (Kwon and Hong, 2014), pumpkin (
Sim et al. 2015), and tomato (
Phan et al. 2016), sunflower (Pallavi,
et al. 2011). However, SSR marker has a limitation for high-throughput genotyping due to a large number of sample and markers (Guichoux
et al. 2011). On the other hand, the next-generation sequencing technology allows large-scale detection of genetic variants, including single nucleotide polymorphisms (SNPs) (
Serba et al. 2019;
Kishor et al. 2021). Now-a-days, SNPs are considered to be the most commonly used powerful genetic markers in plant breeding for detection of genetic differences among various crop cultivars due to their co-dominant inheritance, high abundance in the genome and high- throughput genotyping (
Gupta et al. 2008;
Chung et al. 2014). Therefore, SNP based genotyping technology like Fluidigm SNP array has been developed for effective exploration of genetic differences in the commercial F
1 hybrid cultivars, including pumpkin (
Nguyen et al. 2020), melon (
Kishor et al. 2020), watermelon (
Park et al. 2022), chili pepper (
Kim et al. 2017), tomato (Kim
et al. 2020), pearl millet (
Serba et al. 2019). In this study, we developed SNP type assays for a purity test of F
1 cultivars/breeding line of squash for use in 27 Fluidigm SNP array which will be useful in developing a high-throughput SNP type assays for variety identification in commercial F
1 hybrid squash cultivars.
MATERIALS AND METHODS
Plant materials
Seven F
1 hybrid breeding lines (SQ001, SQ002, SQ003, SQ004, SQ005, SQ006, and SQ007) and seven commercial F
1 cultivars (PMR Teunteunae, DN-A, DN-B, Nongwooae, Jinhanae, Parangsae, and Doksuri) of squash were collec-ted from Republic of Korea were used for validation of the newly developed SNP type assays in this study (
Table 1). A total of 751 individuals of the parangsae cultivars were used for F
1 hybrid seed purity testing.
The genomic DNA was extracted from the young leaves of each squash commercial F1 hybrid cultivars and F1 hybrid breeding lines using the ‘DNeasy Plant Mini Kit’ (Qiagen, Wilmington, USA) according to the manufacturer’s instructions. A Nanodrop spectrophotometer was used to determine the concentration and purity of the extracted DNA samples (Thermo Scientific, Wilmington, USA) and then diluted to a concentration of 20 ng/mL with nuclease- free water.
Fluidigm SNP assay
The SNP information of was collected from ‘Cucurbit Genomics Database (CuGenDB)’ available at
http:// cucurbitgenomics.org. For genotyping, seven F
1 hybrid breeding lines and seven commercial F
1 cultivars were used for hybrid seed purity testing (
Table 1). The Fluidigm SNP assay sets used in this study were synthesized by Macrogen, Korea (
Table 2). The Fluidigm assay was performed using the platform ‘Fluidigm EP1 genotyping’ (Fluidigm, USA) according to the manufacturer’s protocol. The SNPs were then called using the ‘Fludigidm EP1 Genotyping’ analysis software (version 4.8.1).
SNPs were detected using high resolution melting (HRM) analysis combined with a 3’-blocked and unlabeled oligonucleotide probe (HybProbe) specific to the SNP site. The HRM primers used in this study were ordered from Macrogen (
Table 3). After that, using a LightCycler 96 instrument, the gDNA was used for HRM analysis (Roche, Mannheim, Germany) according to the manufacturer’s instructions. HRM was carried out in 10 mL reaction mixtures containing 1 mL at 5ng/mL DNA, 0.1 mL of forward primer (10 pmol), 0.5 mL of reverse primer (10 pmol), 0.5 mL of probe (10 pmol), 0.3 mL of SYTO9 fluorescent dye (Invitrogen, USA), 5 mL of HS prime LP premix (GENETBIO, Korea), and 2.6 mL of DDW. HRM conditions were include three steps, first step is pre- incubation (300 seconds initial pre-incubation at 95℃), second step is 40 cycle of 3 step amplification (95℃ for 10 seconds, 64℃ to 56℃ for 15 seconds under touchdown and 72℃ for 15 seconds), and the last step is HRM reading (last step is four readings per ℃ at the final step after 60 seconds at 95℃, 60 seconds at 40℃, and 1 at 97℃). HRM data was analyzed using LightCycler 96 software (version 1.1) with a 0.2 positive/negative threshold level and 100 percent discrimination for delta Tm and curve shape.
RESULTS
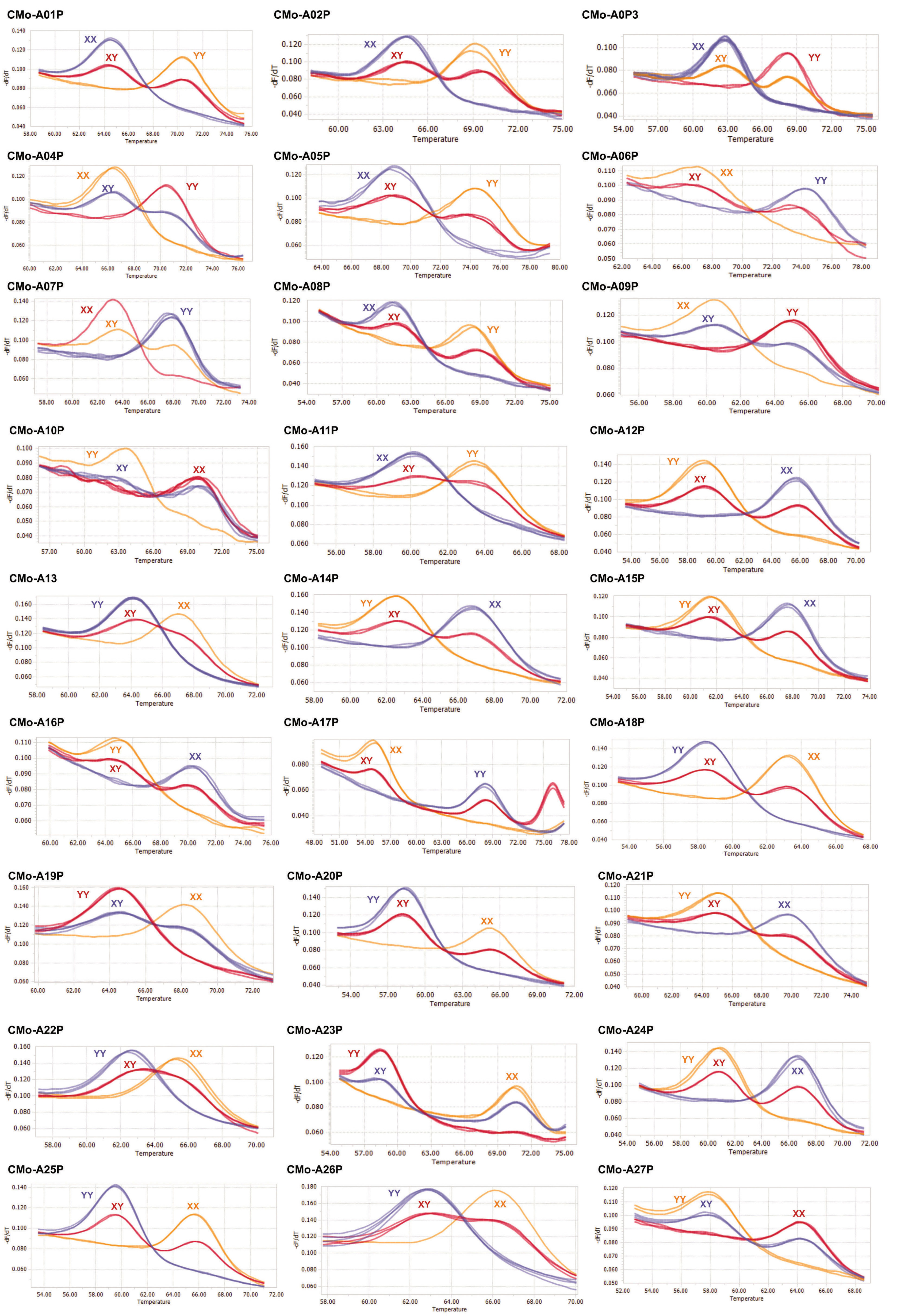
We analyzed a total of 192 primer sets for the ‘Fuidigm SNP type’ assays and 29 of which had successful DNA amplification (
Fig. 1,
Table 2). The red, green, and blue dots represent XX (fluorescence of only the FAM dye), XY (both FAM and HEX dyes), YY (only HEX dye), respec-tively. The SNP markers and their chromosomal position of 27 successful SNPs are presented in
Table 2. The primers and probe sequences of each SNP marker are present in
Table 3. The marker type of each SNP type assay was divided into homozygote reference SNP (XX), homo-zygote alternative SNP (YY) and heterozygotes (XY) (
Fig. 1 and
Table 4). Twenty-nine SNP markers were further checked by the high resolution melting (HRM) analysis (
Fig. 2) and 27 SNP markers can distinguish three different types (
Table 4), including homozygote reference SNP (XX), homozygote alternative SNP (YY) and heterozygotes (XY). Eight SNP markers (CMo-A01, CMo-A02, CMo- A04, CMo-A05, CMo-A12, CMo-A16, CMo-A20 and CMo-A25) accurately distinguished hetero-zygotes (XY) from the registered commercial F
1 hybrid squash cultivars while partially heterozygotes (XY) and partially homozy-gotes (XX and YY) from the analyzed squash breeding lines (
Table 4). Three SNP markers (CMo-A06, CMo-A11 and CMo-A13) were distinguished all the registered com-mercial F
1 hybrid squash cultivars either as homozygote reference allele (XX) or homozygote alternate allele (YY) but partially heterozygotes (XY) and partially homozygote reference allele (XX) from the squash breeding lines. However, CMo-A08 successfully identified most of the registered commercial F
1 cultivars as heterozygotes (XY) except ‘Doksuri’ and CMo-A24 iden-tified the heterozygotes except ‘Doksuri’ and ‘PMR Teun-teunae’.
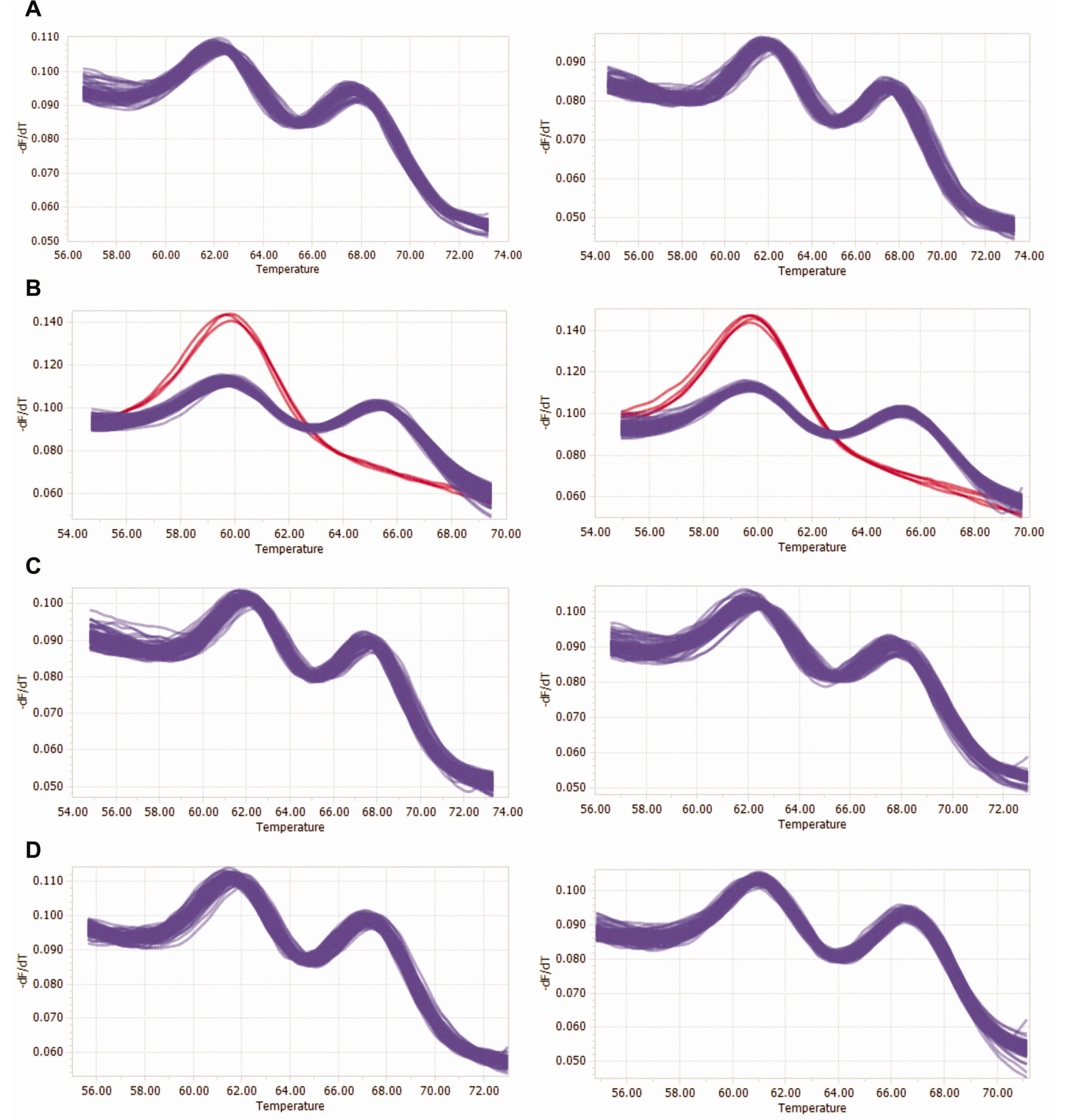
Further, the genetic purity of the registered commercial F
1 hybrid cultivar was determined by HRM analysis using CMo-A03 SNP marker (
Fig. 3). Four lots of individuals from the registered commercial F
1 hybrid cultivar ‘Paransae’ were used to determine genetic purity (
Table 5). The HRM analysis results revealed that LOT1 (186 heterozygotes out of 186 individuals), LOT3 (187 heterozygotes out of 187 individuals) and LOT4 (189 heterozygotes out of 189 individuals) showed 100% purity whereas LOT2 (182 heterozygotes out of 189 individuals) had 96.3% purity (
Table 5).
DISCUSSION
The genetically pure seed crop cultivar is the key criteria for commercial cultivars including F
1 hybrids. We aimed to develop SNP marker set for rapid and high-throughput genotyping system to identify and genetic purity analysis of squash cultivars based on ‘Fluidigm SNP Genotyping’ assay. The ‘Fluidigm SNP Genotyping’ assay is a cost- effective platform for distinguishing commercial crop cul-tivars (
Park et al. 2022). The genetic purity of commercial F
1 hybrids and cultivars is of great importance for the success of plant breeding programs (
Kishor et al. 2020).
In this study, we analyzed seven registered commercial F
1 hybrid cultivars and seven squash breeding lines using 27 SNP markers based on ‘Fluidigm SNP Genotyping’ assay to evaluate the genetic purity (
Table 1 and
5). The results showed that all the tested SNP markers had successful DNA amplification in both the eight registered commercial F
1 hybrid cultivars and the seven breeding lines of squash (
Fig. 1 and
Table 3). Moreover, all of those markers were analyzed using HRM analysis and all were amplified (
Fig. 2). Similar results on HRM-based SNP markers for genetic purity analysis of cultivars/species were reported in melon and
Capsicum species (An
et al. 2010;
Jeong et al. 2010). Even though total 27 SNP markers were tested, eight SNP markers (CMo-A01, CMo-A02, CMo-A04, CMo-A05, CMo-A12, CMo-A16, CMo-A20 and CMo-A25) successfully identified hetero-zygotes (XY) from the registered commercial F
1 hybrid squash cultivars with 100% accuracy (
Table 3). However, in case of squash breeding lines, partially heterozygotes (XY) and partially homozygotes alleles (XX and YY) were identified which suggested that those F
1 hybrid squash breeding lines were contaminated due to outcrossing. Similar results were previously reported in several crop species (
Pattanaik et al. 2018;
Kishor et al. 2020). Then again, HRM analysis of a registered commercial F
1 cultivar ‘Parangsae’ with CMo-A03 SNP marker also revealed 96.30-100% purity of the cultivar (
Fig. 3 and
Table 4). Our results suggested that ‘Fluidigm SNP Genotyping’ platform could be an effective approach for purity testing and identification of registered commercial squash F
1 hybrids cultivars and breeding lines.
STATEMENT
All our experiments complied with local and national regulations.
-
CONFLICT OF INTEREST
The authors declare that they have no competing interests.
-
FUNDING
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Digital Breeding Transformation Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA) (Project number 322071-03) and Sunchon National University Research Fund in 2020 (Grant no. 2020-0192).
-
AUTHOR’S CONTRIBUTIONS
I.-S.N. conceived the study. J.-G.P. and J.-E.H. con-ducted the experiments and analyzed the data. M.A.R. wrote and revised the final version of the manuscript. All authors read the final version and approved the manuscript.
Fig. 1Scatter plots of 27 ‘Fluidigm SNP Genotyping’ assays. Red, blue and green dots indicated XX (fluorescence of the only FAM dye), XY (both FAM and HEX dyes) and YY (only HEX dye) types, respectively.

Fig. 2HRM curve profiles of 27 SNP primers (CMo-A01P – A27P). XX, YY and XY indicate reference SNP, alternative SNP and heterozygotes, respectively.
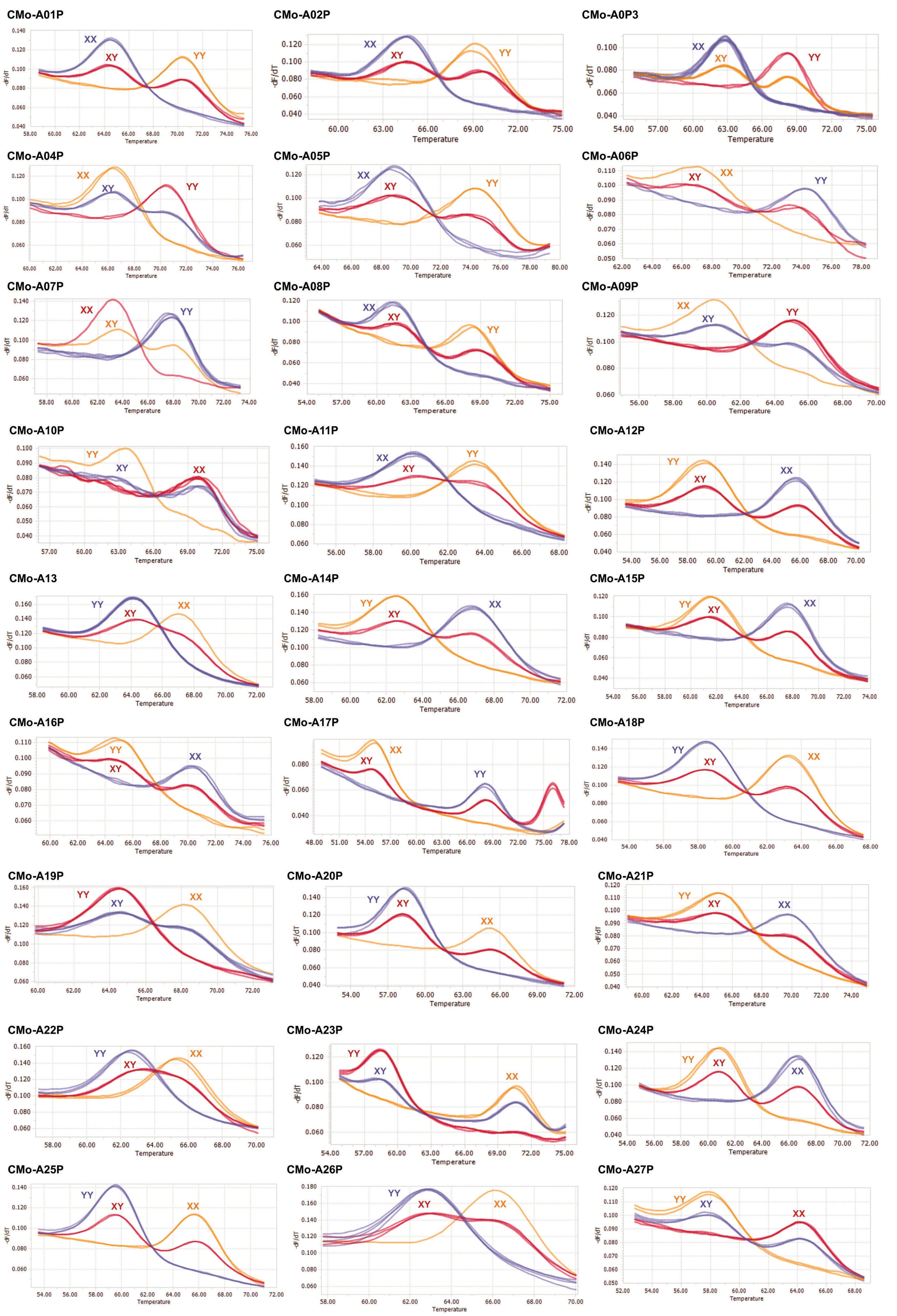
Fig. 3HRM curve profiles for purity test of ‘Parangsae’ F1 cultivars using CMo-A03 marker. (A) LOT1 of ‘Parangsae’, (B) LOT2, (C) LOT3 and (D) LOT4. Red and blue curves indicate genotypes of parental and F1, respectively.
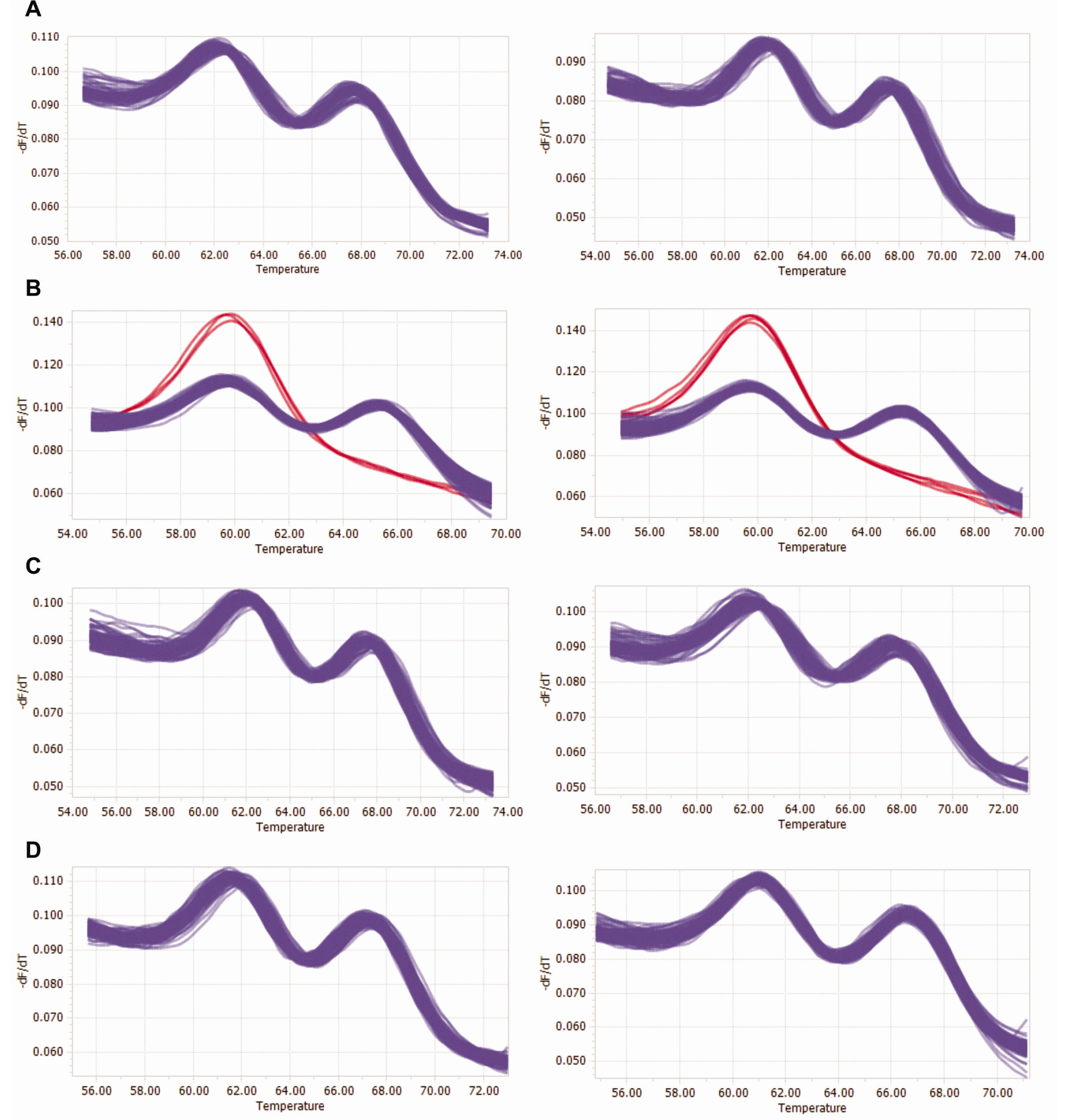
Table 1List of F1 hybrid breeding lines and commer-cial F1 cultivars of squash used in this study.
Table 1
|
No. |
Cultivars |
|
1 |
SQ001 |
|
2 |
SQ002 |
|
3 |
SQ003 |
|
4 |
SQ004 |
|
5 |
SQ005 |
|
6 |
SQ006 |
|
7 |
SQ007 |
|
8 |
PMR Teunteunae |
|
9 |
DN-A |
|
10 |
DN-B |
|
11 |
Nongwooae |
|
12 |
Jinhanae |
|
13 |
Parangsae |
|
14 |
Doksuri |
Table 2List of SNPs used in ‘Fluidigm SNP Genotyping’ assays.
Table 2
|
SNP Marker Name |
Chromosome |
Location (bp) |
SNP [Ref/Alt] |
SNP (color of dye)a)
|
|
CMo-A01 |
1 |
3,318,442 |
…CAA[A/G]CCA… |
A(F):G(H) |
|
CMo-A02 |
1 |
9,837,040 |
…TCT[G/A]AGG… |
G(F):A(H) |
|
CMo-A03 |
2 |
8,873,265 |
…CTC[A/G]GCA… |
A(F):G(H) |
|
CMo-A04 |
2 |
10,378,972 |
…GAG[G/A]ATT… |
G(F):A(H) |
|
CMo-A05 |
3 |
5,724,100 |
…AGC[G/C]CCG… |
G(F):C(H) |
|
CMo-A06 |
3 |
6,119,169 |
…AGG[T/C]TTA… |
T(F):C(H) |
|
CMo-A07 |
4 |
8,389,880 |
…CTC[G/A]ATT… |
G(F):A(H) |
|
CMo-A08 |
4 |
20,447,172 |
…TCC[C/T]TCA… |
C(F):T(H) |
|
CMo-A09 |
6 |
3,712,655 |
…TTG[G/A]GTC… |
G(F):A(H) |
|
CMo-A10 |
6 |
11,115,368 |
…TTC[C/T]CTC… |
C(F):T(H) |
|
CMo-A11 |
8 |
2,798,704 |
…AGG[T/G]TAA… |
T(F):G(H) |
|
CMo-A12 |
8 |
6,252,977 |
…AAA[C/A]AAA… |
C(F):A(H) |
|
CMo-A13 |
10 |
6,646,259 |
…GCT[C/T]GAT… |
C(F):T(H) |
|
CMo-A14 |
11 |
4,451,324 |
…GCC[G/A]AAC… |
G(F):A(H) |
|
CMo-A15 |
13 |
8,296,803 |
…GAA[C/T]GCG… |
C(F):T(H) |
|
CMo-A16 |
15 |
1,475,799 |
…TGT[G/C]CAA… |
G(F):C(H) |
|
CMo-A17 |
15 |
9,704,446 |
…GGG[T/C]GTA… |
T(F):C(H) |
|
CMo-A18 |
16 |
1,782,015 |
…TAT[G/A]TAG… |
G(F):A(H) |
|
CMo-A19 |
16 |
3,290,808 |
…TTG[G/T]TGC… |
G(F):T(H) |
|
CMo-A20 |
16 |
8,018,335 |
…GGG[G/T]AAA… |
G(F):T(H) |
|
CMo-A21 |
17 |
2,812,397 |
…GAC[G/A]GTC… |
G(F):A(H) |
|
CMo-A22 |
17 |
10,502,431 |
…TCC[C/T]GTA… |
C(F):T(H) |
|
CMo-A23 |
18 |
1,876,608 |
…CAG[G/A]TCT… |
G(F):A(H) |
|
CMo-A24 |
18 |
3,666,538 |
…TTT[C/G]ATG… |
C(F):G(H) |
|
CMo-A25 |
18 |
10,701,307 |
…TCT[C/G]ACT… |
C(F):G(H) |
|
CMo-A26 |
20 |
3,466,354 |
…TGC[C/A]AGA… |
C(F):A(H) |
|
CMo-A27 |
20 |
4,075,437 |
…CTA[G/A]GCT… |
G(F):A(H) |
Table 3List of primer and probe sequences used in HRM assay of SNPs.
Table 3
|
HRM Marker |
Primer (5’-3’) |
|
Forward |
Reverse |
Probea)
|
|
CMo-A01P |
AGTCTTCAGTGCCAACGGTGATTC |
ATGGGTTTTGGAGGAGATTCTTATC |
AGATTGGCAGGCAAACCATGAGCAT |
|
CMo-A02P |
ATTGCCAAAATGCCATTAAGTAAGC |
GTTGTAGTTTTAGCTGCTGCTCTCA |
CTGCAAATCTTACCTCTGAGGTGTTT |
|
CMo-A03P |
ATTATAGAACATGATATGCTGCCCAC |
TCCCTTACGCTATCACTTGTTTGTTAA |
ACATTCACAAATCTCAGCATCATCC |
|
CMo-A04P |
GTCCACCAACCCATTATGCTTTGAA |
AAGAGGCAATAGTGGAAGAACTTG |
AGCAAGGTGGAGAGGATTGTTTCCA |
|
CMo-A05P |
TGAGCAGCTCTTCAACCTCTTCGAA |
TGCTGCAGTTTCAAATTGGGTTTCA |
AGGAATGTACCAGCAGCGCCGAGA |
|
CMo-A06P |
AGCAGCAATGTTGGCAGTGGCAAA |
TTCCCATGTTGACCTCAATGTCCT |
AAGGCACCAATGCCTAAACCTCGCA |
|
CMo-A07P |
GATGAAGTTTCTTCAAGAACTAGTCC |
GACATAGTTAATGATTTCGTCTAGCA |
TCACATCAATTCGCTCGATTAAAGTT |
|
CMo-A08P |
TTGAAGTCCATGCAGCCCTTGTT |
CTGTTCTTCAGCCTCAATGTGGAT |
AGACCATTTCTCTCCCTCAATCACT |
|
CMo-A09P |
TGAAACTGTGTAAACTGGCTGCTCT |
CATGTTGGATTTAAGAAATGGAAGAAG |
TTCAAGAGGAATTGGGTCTGAAGAA |
|
CMo-A10P |
CAATTGGAGAAAGGGTTTCGCG |
AGCAGCAGCGGAATGAAGTTGA |
TTGAAGAACTCGATTCCCTCGAGAG |
|
CMo-A11P |
CTTATATGTGCATGGAGAAACGGC |
CATGAGAGAAATGATGAAAGGACAG |
TCCCTAAAACCTTAACCTCTTTCAG |
|
CMo-A12P |
GCGGTTGTTACTCATATAATGAGAAG |
GAGCATCTGCAGCAAGTTCTCTC |
TGTTTTCCCTTTGTTTGGGGTTGTT |
|
CMo-A13P |
GAGGTTGCACATCGGCTAGGT |
CTGCAGCAAGACCTATAGGATTTGA |
GAGAAGAGGCTTGATCCACAAAGTT |
|
CMo-A14P |
ATGTCAAATAAATCTGTCTCGACGC |
TGGAATGGATAATCTAGAGCTACAG |
TCTGGAAATGCCGAACTTTTGATACT |
|
CMo-A15P |
CGGCATTGTCGAGAGATATCGA |
ATCGTGAAGAACTCCATAATGGCT |
ACCGATAAGCTTCGCATTCCTGTTG |
|
CMo-A16P |
TGTACTTTCATGGAAGCTGGCGTT |
TGACGGTTCGGAGGTTGGAGA |
TACGGAAGATGTGCAAAGACGCCAT |
|
CMo-A17P |
GTGATATAATTCCAGTAATTTGCAGC |
TTGCCCATTCTATTGCAGCTATAG |
TTCATCCTACGCCCATCGGTTCAT |
|
CMo-A18P |
TCTTGATCTCTTCCATTCTGGATC |
GATGAAGCAAGCGAAATTGCTACA |
TCTACGTCTTATATAGATTCTGACACG |
|
CMo-A19P |
GCAGCTATCTAAGAAGGCTAAATATC |
ATGATTCGAAGAAACTGCCGAATTAGT |
TCCTTCTTTGGTTGGTGCAATGTGC |
|
CMo-A20P |
CCGAAACTCAACGTCAAATAATGTG |
ACGGAGAGCGAAGGGCTAAATTT |
TCAAGTCTTTTCCCCACCATCCAAT |
|
CMo-A21P |
TGATTGCGAAATAGTCTTTCGTTGC |
TCTTGTTCAGCGTTCGAGTATCGA |
GACTCGACGACGGTCACGATGA |
|
CMo-A22P |
TGCTTATCAGAGTGGCATTTATTCTG |
GAGCTTAGTAAGAATGGTGATTAGAAC |
AGACATACCGTCCTGTACAGTAAGA |
|
CMo-A23P |
GAAGCTTACAAACGGGTATGCAGA |
ATATGTAGTGTTCTTGATAGTGTTAAT |
GTGACAAAGACGCTGCCATTGTCTT |
|
CMo-A24P |
CTCACTTAAGAGGATCCAGGGTG |
TGCCTTGAACACCAATGTTGCCTT |
CATGACAGCATGAAAAGCATTCTCTG |
|
CMo-A25P |
TACGAGTCGCATTTCTTGACCG |
ACGAAGTTAGCACAGTAATAGTCATC |
TTAGGTCATTTCTGACTTTGATGCTAG |
|
CMo-A26P |
CTGTCAACTGTTTGAATACTCGGG |
GTACCTAACAGTTGAAGAATCTCCA |
TGGACCAGAAATGCAAGAAATGGAG |
|
CMo-A27P |
CATGTGAGCTGCTGTGGACGATT |
ACAAGCGAAAGACTTCATAGCAGATAT |
AGGAATAGATCTAAGCTGAATGGCT |
Table 4Validation of ‘Fluidigm SNP Genotyping’ assay developed for the purity test.
Table 4
|
Assay |
SQ001 |
SQ002 |
SQ003 |
SQ004 |
SQ005 |
SQ006 |
SQ007 |
PMR
Teun-teunae |
DN-A |
DN-B |
Nong-wooae |
Jin-hanae |
Parang-sae |
Dok-suri |
|
CMO-A01 |
YY |
YY |
XY |
XY |
XY |
YY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A02 |
YY |
YY |
XY |
XY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A03 |
YY |
YY |
XY |
YY |
YY |
YY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A04 |
XY |
YY |
XY |
XY |
XY |
YY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A05 |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A06 |
XX |
XY |
XY |
XY |
XX |
XY |
XY |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
|
CMO-A07 |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
YY |
XY |
|
CMO-A08 |
YY |
YY |
XY |
XY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XX |
|
CMO-A09 |
XX |
XX |
XY |
XY |
XY |
XX |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A10 |
XY |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A11 |
YY |
YY |
XY |
YY |
YY |
YY |
XY |
YY |
YY |
YY |
YY |
YY |
YY |
YY |
|
CMO-A12 |
XX |
XX |
XY |
XY |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A13 |
XX |
XX |
XY |
XX |
XX |
XX |
XY |
XX |
XX |
XX |
XX |
XX |
XX |
XX |
|
CMO-A14 |
XX |
XX |
XX |
XY |
XX |
XX |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A15 |
XY |
YY |
YY |
XY |
YY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
YY |
YY |
|
CMO-A16 |
XY |
XX |
XX |
XY |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A17 |
XY |
YY |
XY |
XY |
XY |
YY |
XX |
XY |
XY |
XY |
XY |
XY |
XX |
XX |
|
CMO-A18 |
XY |
YY |
YY |
XY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XX |
XX |
|
CMO-A19 |
YY |
YY |
YY |
YY |
YY |
YY |
YY |
YY |
XY |
XY |
XY |
XY |
YY |
YY |
|
CMO-A20 |
XY |
YY |
YY |
YY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A21 |
YY |
YY |
XY |
XY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XX |
|
CMO-A22 |
XY |
XX |
XY |
XY |
XY |
XX |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
XX |
|
CMO-A23 |
XY |
XY |
YY |
YY |
YY |
YY |
XX |
YY |
XY |
XY |
XY |
XY |
XY |
YY |
|
CMO-A24 |
XY |
XY |
XX |
XX |
XX |
XX |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XX |
|
CMO-A25 |
XY |
XX |
XX |
XY |
XY |
XX |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
XY |
|
CMO-A26 |
XX |
XX |
XY |
XX |
XX |
XX |
XY |
XX |
XX |
XX |
XX |
XX |
XX |
XY |
|
CMO-A27 |
YY |
YY |
XY |
YY |
YY |
YY |
XY |
YY |
XY |
XY |
XY |
XY |
XY |
XY |
Table 5Results of purity test in ‘Parangsae’ cultivars.
Table 5
|
Cultivar |
Lots |
No. of indivi-duals |
SNP marker |
Heterozygote (F1) |
Purity (%) |
|
Parangsae |
LOT1 |
186 |
CMo-A03 |
186 |
100 |
|
LOT2 |
189 |
CMo-A03 |
182 |
96.3 |
|
LOT3 |
187 |
CMo-A03 |
187 |
100 |
|
LOT4 |
189 |
CMo-A03 |
189 |
100 |
References
- Caruso M, Federici CT, Roose ML. 2008. EST-SSR markers for asparagus genetic diversity evaluation and cultivar identification. Mol. Breed.. 21(2): 195-204.
- Chung H, Jeong YM, Mun JH, Lee SS, Chung WH, Yu HJ. 2014. Construction of a genetic map based on high- throughput SNP genotyping and genetic mapping of a TuMV resistance locus in Brassica rapa. Mol. Genet. Genom.. 289(2): 149-160.
- Ferriol M, Pico B, de Cordova PF, Nuez F. 2004. Molecular diversity of a germplasm collection of squash (Cucurbita moschata) determined by SRAP and AFLP markers. Crop Sci. 44(2):pp. 653-664.
- Gomes RS, de Almeida CF, Chagas RR, Júnior RM, Fara JS, da Silva DJH. 2020. Winter squash (Cucurbita moschata D.) Displays Promising Nutritional Aspects in Fruits, Seeds and in the Seed Oil. J. Plant Biochem. Physiol.. 8: 248.
- Guichoux E, Lagache L, Wagner S, Chaumeil P, Léger P, Lepais O, et al. 2011. Current trends in microsatellite genotyping. Mol. Ecol. Resour.. 11(4): 591-611.
- Guiné RPF, Henrriques F, João Barroca M. 2012. Mass Transfer Coefficients for the Drying of Pumpkin (Cucurbita moschata) and Dried Product Quality. Food Bioproc. Tech.. 5: 176-183.
- Gupta PK, Rustgi S, Mir RR. 2008. Array-based high- throughput DNA markers for crop improvement. Heredity. 101(1): 5-18.
- Jacobo-Valenzuela N, Zazueta-Morales JJ, Gallegosinfante JA. 2011. Chemical and physicochemical characterization of winter squash (Cucurbita moschata D.) Notulae Botanicae. Horti. Agrobotanici.. 9: 34-40.
- Jacobo-Valenzuela N, Maróstica-Junior MR, de Jesús Zazueta-Morales J, Gallegos-Infante JA. 2011. Physico-chemical, technological properties, and health-benefits of Cucurbita moschata Duchense vs A Review. . Food Res. Int.. 44(9): 2587-2593.
- Jamali SH, Cockram J, Hickey LT. 2019. Insights into deployment of DNA markers in plant variety protection and registration. Theor. Appl. Genet.. 132(7): 1911-1929.
- Jeong HJ, Jo YD, Park SW, Kang BC. 2010. Identification of Capsicum species using SNP markers based on high resolution melting analysis. Genome. 53(12): 029-1040.
- Kim DK, Seo SG, Kwon SB, Park YD. 2016. Development of RAPD and SCAR markers related to watermelon mosaic virus and zucchini yellow mosaic virus resistance in Cucurbita moschata. Hortic. Environ. Biotechnol.. 57(1): 61-68.
- Kim H, Yoon JB, Lee J. 2017. Development of fluidigm SNP type genotyping assays for marker-assisted breeding of chili pepper (Capsicum annuum L.). Hortic. Sci. Technol.. 35(4): 465-479.
- Kim M, Jung JK, Shim EJ, Chung SM, Park Y, Lee GP, et al. 2021. Genome-wide SNP discovery and core marker sets for DNA barcoding and variety identification in commercial tomato cultivars. Sci. Hortic.. 276: 109734
- Kishor DS, Noh Y, Song WH, Lee GP, Jung JK, Shim EJ, et al. 2020. Identification and purity test of melon cultivars and F1 hybrids using fluidigm-based snp markers. Hortic. Sci. Technol.. 38(5): 686-694.
- Kishor DS, Noh Y, Song WH, Lee GP, Park Y, Jung JK, et al. 2021. SNP marker assay and candidate gene identifi-cation for sex expression via genotyping-by-sequencing- based genome-wide associations (GWAS) analyses in Oriental melon (Cucumis melo L. var. makuwa). Sci. Hortic.. 276: 109711
- Kwon YS, Oh YH, Yi SI, Kim HY, An JM, Yang SG, et al. 2010. Informative SSR markers for commercial variety discrimination in watermelon (Citrullus lanatus). Genes Genom.. 32(2): 115-122.
- Lee ON, Park HY. 2017. Assessment of genetic diversity in cultivated radishes (Raphanus sativus) by agronomic traits and SSR markers. Sci. Horti.. 223: 19-30.
- Mashitoa FM, Shoko T, Shai JL, Slabbert RM, Sultanbawa Y, Sivakumar D. 2021. Influence of different types of drying methods on color properties, phenolic metabolites and bioactivities of pumpkin leaves of var. Butternut squash (Cucurbita moschata Duchesne ex Poir). Front. nutr. 8:
- Nandakumar N, Singh AK, Sharma RK, Mohapatra T, Prabhu KV, Zaman FU. 2004. Molecular fingerprinting of hyb-rids and assessment of genetic purity of hybrid seeds in rice using microsatellite markers. Euphytica. 136(3): 257-264.
- Nguyen NN, Kim M, Jung JK, Shim EJ, Chung SM, Park Y. 2020. Genome-wide SNP discovery and core marker sets for assessment of genetic variations in cultivated pumpkin (Cucurbita sp.). Hortic. Res. 7: pp. 121-
- Pallavi HM, Gowda R, Shadakshari YG, Bhanuprakash K, Vishwanath K. 2011. Identification of SSR markers for hybridity and seed genetic purity testing in sunflower (Helianthus annuus L.). Helia. 34(54): 59-66.
- Park JY, Jang YJ, Jung JK, Shim EJ, Sim SC, Chung SM. 2022. Development of SNP Markers for the Identification of Commercial Korean Watermelon Cultivars Using Fluidigm Genotyping Analysis. Hortic. Sci. Technol.. 40(1): 75-84.
- Pattanaik A, Reddy DCL, Ramesh S, Aswath C. 2018. Comparison of traditional grow-out test and DNA-based PCR assay to estimate F1 hybrid purity in cauliflower. Curr. Sci.. 115: 2095-2102.
- Phan NT, Kim MK, Sim SC. 2016. Genetic variations of F1 tomato cultivars revealed by a core set of SSR and InDel markers. Sci. Hortic.. 212: 155-161.
- Pierantoni L, Cho KH, Shin IS, Chiodini R, Tartarini S, Dondini L. 2004. Characterisation and transfera-bility of apple SSRs to two European pear F1 populations. Theor. Appl. Genet. 109(7):pp. 1519-1524.
- Pierantoni L, Cho KH, Shin IS, Chiodini R, Tartarini S, Dondini L, et al. 2019. Genetic diversity, population structure, and linkage disequilibrium of pearl millet. Plant Genome. 12(3): 180091
- Sim SC, Hong JH, Kwon YS. 2015. DNA profiling of commercial pumpkin cultivars using simple sequence repeat polymorphisms. Hortic. Environ. Biotechnol.. 56(6): 811-820.
- Tsukazaki H, Fukuoka H, Song YS, Yamashita KI, Wako T, Kojima A. 2006. Considerable heterogeneity in commer-cial F1 varieties of bunching onion (Allium fistulosum) and proposal of breeding scheme for conferring variety traceability using SSR markers. Breed. Sci.. 56(3): 321-326.
- Yadav M, Jain S, Tomar R, Prasad GBKS, Yadav H. 2010. Medicinal and biological potential of pumpkin: an updated review. Nutr. Res. Rev.. 23(2): 184-190.
- Yashitola J, Thirumurugan T, Sundaram RM, Naseerullah MK, Ramesha MS, Sarma NP, et al. 2002. Assessment of purity of rice hybrids using microsatellite and STS markers. Crop Sci.. 42(4): 1369-1373.
- Ye S, Wang Y, Huang D, Li J, Gong Y, Xu L, et al. 2013. Genetic purity testing of F1 hybrid seed with molecular markers in cabbage (Brassica oleracea var. capitata). Sci. Hortic.. 155: 92-96.