Abstract
Doubled haploid (DH) technology enables the generation and evaluation of new plant genotypes in ≤ 2 years, which is in stark contrast to the 6-7 years required for conventional breeding. Recently, a few proteins including MATRILINEAL (MTL) were found to trigger haploid induction in monocot plants. However, MTL function in dicot plants remains unclear. Here, we investigate the effects of Arabidopsis PATATIN-RELATED PHOSPHOLIPASE 2A (pPLA-IIa), a maize MTL homolog, on the expression of genes that modulate pollen development and fertilization in Arabidopsis. Quantitative real-time PCR analysis showed that transcript levels of four pollen tube growth-related genes and six pollen guidance- or reception-related genes were increased in the ppla-iia T-DNA insertion mutant compared with the wild type. In addition, transcript levels of four mitosis- and meiosis-related genes, two hormone-related genes, and a calcium pump-encoding gene were also upregulated in the ppla-iia mutant, whereas expression levels of genes encoding a synergid-secreted peptide (AtLURE1.1) and calmodulin-activated Ca2+-ATPase ion pump 9 (ACA9) protein were downregulated in the ppla-iia mutant compared with the wild type. Taken together, these results suggest that AtLURE1.1 and ACA9 play positive roles in pPLA-IIa-mediated fertilization. Thus, pPLA-IIa controls fertilization through the modulation of pollen germination and pollen tube growth and guidance in Arabidopsis.
-
Key words: pPLA-IIa, Haploid, Patatin-like phophpolipase 2A, Pollen tube, Synergid, Fertilization
INTRODUCTION
Phospholipase A2 (PLA) enzymes participate in numerous developmental and physiological processes through the production of free fatty acid and lysophos-pholipid signaling molecules in animals and plants (
Meijer and Munnik 2003;
Shimizu et al. 2006). PLA proteins are classified into four different groups: secreted PLA2 (sPLA2), calcium-independent PLA2 (iPLA2), cytosolic PLA2 (cPLA2), and patatin-like PLA2 (pPLA2) (
Six and Dennis 2000).
The pPLA proteins contain a patatin domain. In
Arabidopsis thaliana, the pPLA family is divided into three subfamilies: pPLA-I, pPLA-II (a, b, g, d, e), and pPLA-II (a, b, g, d). The pPLA-I subfamily proteins contain a leucine-rich enzymatic domain at the C-terminal end. The catalytic domains of pPLA-I and pPLA-II subfamily proteins contain 6-7 exons, similar to the animal iPLA2s (
Scherer et al. 2010). The pPLA-III subfamily proteins possess only one intron and harbor a unique feature in the patatin domain, which distinguishes them from other patatin domain-containing enzymes (
Scherer et al. 2010).
In plants, pPLA-II was first described as a lipid acyl hydrolase and potato tuber storage protein (
Senda et al. 1996;
Shewry 2003). The Arabidopsis genome encodes ten pPLA-II proteins (
Ryu 2004). The activity of pPLA proteins is rapidly induced by different environmental signals, and the reaction products (polyunsaturated fatty acids and lysolipids) function as secondary messengers to regulate distinct proteins or downstream processes (
Scherer 1996;
Meijer and Munnik 2003). In many plants, pPLAs are induced upon infection to help control the spread of disease, whereas in mammals, pPLAs are mostly involved in lipid metabolism and turnover.
Recently, a sperm-specific pPLA2, known as MATRILINEAL (MTL), was shown to trigger haploid induction in maize (
Zea mays) and rice (
Oryza sativa) (
Gilles et al. 2017;
Kelliher et al. 2017;
Liu et al. 2017;
Yao et al. 2018). Haploid seed production is an important breeding approach, as it helps achieve complete homozygosity within a single generation using the doubled haploid (DH) technology (
Dwivedi et al. 2015). However, Arabidopsis
ppla-iia mutants did not produce haploid seeds (
Kwak et al. 2021). Instead, Arabidopsis
ppla-iia mutants flowered later than wild-type plants under long day conditions (
Kwak et al. 2021).
Nevertheless, the function of pPLA2-II in reproduction, specifically pollen germination, pollen tube growth, pollen reception, and fertilization, has not yet been identified. Therefore, in this study, we characterized the role of Arabidopsis pPLA-IIa, a homolog of maize MTL, in pollen development and fertilization. The expression patterns of various pollen development- and fertilization- related genes were investigated in the ppla-iia T-DNA insertion mutant. We found that only AtLURE1.1 and ACA9 genes were downregulated in the ppla-iia mutant, whereas other genes were upregulated, indicating that AtLURE1.1 and ACA9 act as positive regulators of pPLA-IIa-mediated pollen development and fertilization.
MATERIALS AND METHODS
Plant materials and growth conditions
Arabidopsis thaliana ecotype Columbia (Col-0; wild type [WT]) and T-DNA insertion knockout mutant ppla-iia (SAIL_172_F04) were used in this study. The At2g26560 T-DNA insertion mutant ppla-iia was obtained from the Arabidopsis Biological Resource Center at Ohio University. To grow plants in vitro, seeds were surface- sterilized in commercial bleach, containing 5% sodium hypochlorite and 0.1% Triton X-100, for 10 minutes, and then rinsed five times in sterilized water. The surface- sterilized seeds were cold-stratified in the dark at 4℃ for 3 d, and then sown on Murashige and Skoog (MS) medium containing 1% sucrose and 0.8% agar (pH 5.7). To grow plants in soil, seeds were directly sown into sterile vermiculite. Plants were grown in a growth chamber at 22℃ under long-day (LD) photoperiod (16 hours light/8 hours dark).
Quantitative real-time RT-PCR analysis
Expression levels of
ANXUR1 (
ANX1),
ANX2,
CHROMOSOME TRANSMISSION FIDELITY 7 (
AtCTF7),
AtMYB97,
RAPID ALKALINIZATION FACTOR 4 (
AtRALF4),
ATUNIS1 (
AUN1),
BRASSINOSTEROID INSENSITIVE 1 (
BRI1),
centromere-specific histone 3 (
CENH3),
Centromeric Protein C (
CENPC),
CLAVATA3/EMBRYO SURROUNDING REGION-RELATED PEPTIDE 45 (
CLE45),
CORONATINE INSENSITIVE1 (
COI1),
DUO POLLEN1 (
DUO1),
LOST IN POLLEN TUBE GUIDANCE 1 (
LIP1),
LIP2,
LEUCINE-RICH REPEAT EXTENSIN 10 (
LRX10),
MALE DISCOVERER1 (
MDIS1),
OMISSION OF SECOND DIVISION1 (
OSD1),
POLLEN RECEPTOR-LIKE KINASE 1 (
PRK1), and
Rho of plant1 (
ROP1) genes were examined by quantitative real-time RT-PCR in WT and
ppla-iia mutant plants grown under LD conditions, as described above. Total RNA was isolated from the flowers of WT and
ppla-iia plants, and mRNA levels were analyzed as previously described (
Park et al. 2011). Quantitative real-time RT-PCR was performed using gene-specific primers (
Table 1).
UBQ10-specific primers were included in each reaction as an internal control. All reactions were performed three times with three independent RNA samples.
RESULTS
Genes encoding pollen tube growth regulators were upregulated in the ppla-iia mutant
In previous study, we found that pPLA-IIa loss upregulated the floral repressor
FLOWERING LOCUS C (
FLC) expression and downregulated a long intronic noncoding RNA
COLDAIR expression, resulting in late flowering of the
ppla-iia mutants (
Kwak et al. 2021;
Fig. 1). However, there was no big difference in the growth between WT and
ppla-iia mutants at germination and vegetative stages (
Fig. 1). In addition,
ppla-iia mutants did not produce abnormal seeds (
Kwak et al. 2021). Therefore, we focused on the analysis of the expression of various genes involved in fertilization including pollen and pollen tube growth in WT and
ppla-iia plants.
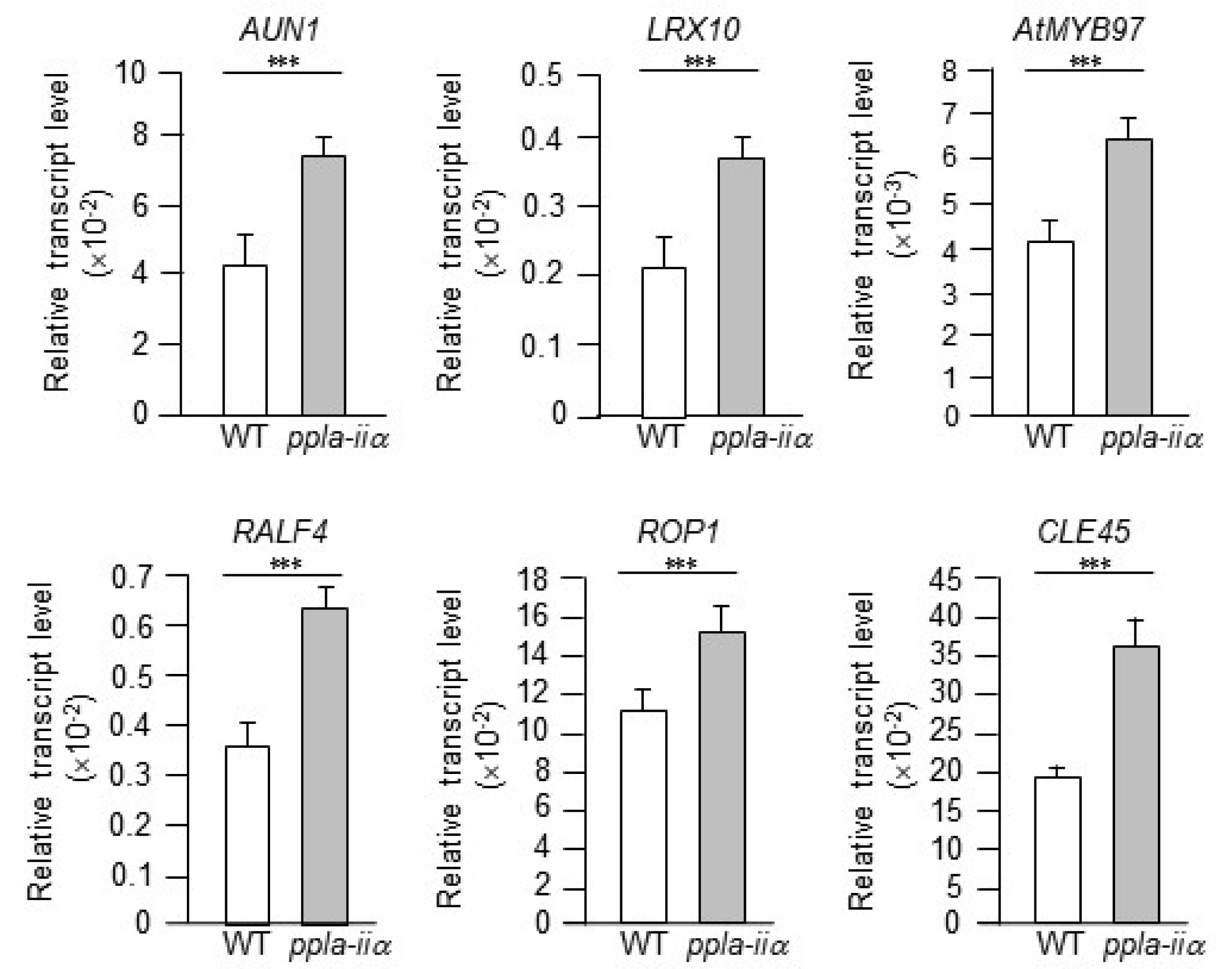
We first examined the expression levels of six genes,
AtRALF4,
AUN1,
LRX10,
AtMYB97,
ROP1, and
CLE45, involved in pollen tube growth. The RALF family of proteins include cysteine-rich peptides and the ligands of receptor-like kinases of the
Catharanthus roseus receptor- like kinase 1 (RLK1)-like (CrRLK1L) subclass. AtRALF4 strongly inhibits pollen germination (
Morato et al. 2014), and its function depends on pollen-expressed LRX family proteins, which play a critical role in cell wall development (
Mecchia et al. 2017). AUN1 is a phosphatase that acts as a nucleocytoplasmic negative regulator of tip growth. A recent study showed that AUN1
D94N and AUN1
H127A proteins, harboring substitutions of key amino acid residues in the conserved catalytic domain, function as dominant amorphic variants that repress pollen tube growth (
Franck et al. 2018). LRXs are chimeric proteins that contain an LRR and an EXT domain (
Baumberger et al. 2003). The
LRX genes are grouped into two classes, depending on their spatial expression patterns; the first group of LRXs is expressed in vegetative tissue (
Zhou et al. 1992), while the second group is expressed specifically in pollen (
Baumberger et al. 2003). LRX10 belongs to the second group, and the Arabidopsis
lrx10 mutant produces abnormal pollen tubes, with an irregular deposition of callose and pectin (
Sede et al. 2018).
AtMYB97 is a R2R3-MYB transcription factor and is mainly expressed in mature pollen grains and pollen tubes (
Liang et al. 2013). Mutation of the
MYB97 gene causes the overgrowth of pollen tubes in the embryo sac, disrupts the discharge of sperm cells into the embryo sac, and significantly alters the expression of a group of pollen-expressed genes in mature pollen grains (
Liang et al. 2013). Rop1 is a small G-protein molecular switch that regulates pollen germination and tube growth (
Kost et al. 1999;
Li et al. 1999;
Fu et al. 2001). Actin assembly in the pollen tip requires AtRop1 activation (
Fu et al. 2001). CLE45 is a small peptide composed of 90 amino acids that prolongs pollen tube growth at high temperatures. The
CLE45 gene was preferentially expressed in the stigma within the pistil at 22℃. However, upon the temperature shift to 30℃, the expression of
CLE45 expanded into the transmitting tract, along which the pollen tubes elongate.
Based on these reports, we examined the expression levels of
AtRALF4,
AUN1,
LRX10,
AtMYB97, and
ROP1 in the flowers of WT and
ppla-iia mutant plants. The
ppla-iia mutant showed approximately 2-fold higher expression of
AtRALF4,
AUN1,
LRX10, and
AtMYB97 and approximately 1.5-fold higher expression of
ROP1 than the WT (
Fig. 2).
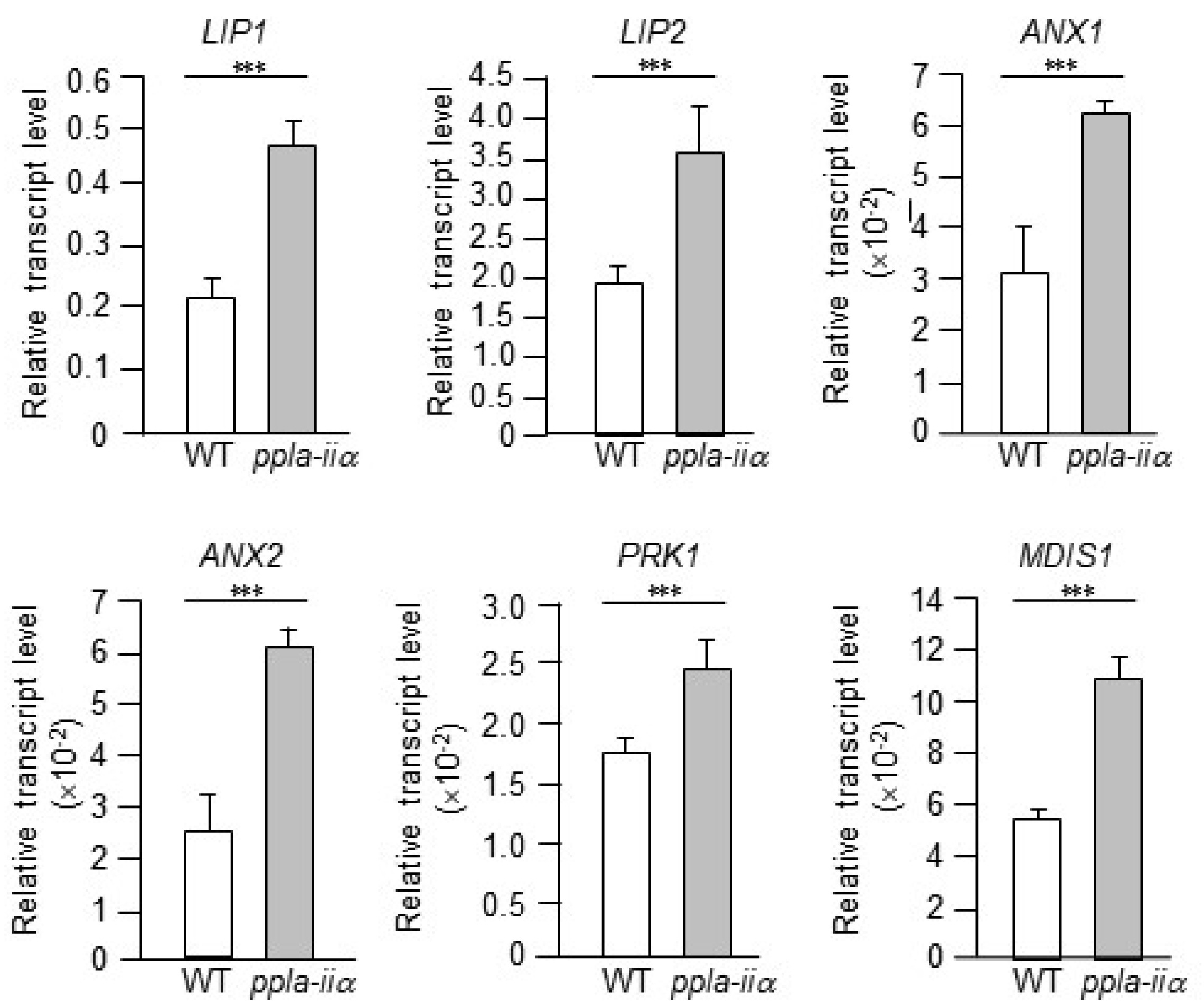
Next, we checked the expression levels of six genes,
LIP1,
LIP2,
ANX1,
ANX2,
PRK1, and
MDIS1, involved in pollen guidance or reception. LIP1 and LIP2 are RLKs that localize to the pollen tube tip plasma membrane (
Liu et al. 2013). The
lip1 lip2 double mutant is defective in pollen tube guidance triggered by AtLURE1. Interestingly, LIPs are not direct receptors of pollen because they lack an extracellular domain. Genes encoding RLKs such as ANXUR1/2 (ANX1/2) (
Boisson-Dernier et al. 2009;
Miyazaki et al. 2009) and MDIS1/2 (
Wang et al. 2016) show the highest expression in mature pollen grains and/or pollen tubes, and play important roles in pollen tube development. The anx1 anx2 double mutant is sterile because pollen burst immediately after germination (
Boisson-Dernier et al. 2009;
Miyazaki et al. 2009).
On the contrary,
ANX1/2 overexpression lines show defective pollen tube growth due to the overaccumulation of cell wall material and invagination of the plasma membrane (Boisson-Dernier
et al. 2013). Plants expressing kinase-dead dominant negative MDIS1 (MDIS1
DN) variant protein exhibit decreased micropylar guidance and fertilization efficiency, indicating that MDIS1
DN interferes with pollen tube guidance (
Wang et al. 2016). PRK-encoding genes were first identified in petunia (
Petunia ×
atkinsiana; PRK1) (
Lee et al. 1996) and tomato (
Solanum lycopersicum; SlPRK1/2) (
Muschietti et al. 1998). Although similar in structure, PRK1 and SlPRK1/2 perform different functions. PRK1 plays a role in microspore development (
Lee et al. 1996), while LePRK1/2 participate in pollen tube growth and/or pollen–pistil interactions (
Zhang et al. 2008;
Salem et al. 2011;
Gui et al. 2014). In Arabidopsis, AtPRK1 and AtPRK3 are predicted to function as the receptors of the AtLURE1.2 peptide. In addition, AtPRK3 is involved in pollen tube growth, plant fertility, and plant reproduction (
Takeuchi and Higashiyama 2016).
In this study, we examined the expression levels of
LIP1,
LIP2,
ANX1,
ANX2,
MDIS1, and
PRK1 in the flowers of WT and
ppla-iia plants. Compared with the WT, transcript levels of
LIP1,
LIP2,
ANX1,
ANX2, and
MDIS1 were approximately 2-fold higher in the
appla2 mutant, while that of
PRK1 was approximately 1.5-fold higher in
ppla-iia mutant flowers (
Fig. 3).
Research in
Torenia fournieri showed that
LURE genes, which encode defensin-like cysteine-rich polypeptides, were specifically expressed in the synergid cell to attract the pollen tube (
Okuda et al. 2009). Consistent with this finding, the injection of morpholino antisense oligomers targeting
LURE impaired pollen tube attraction (
Okuda et al. 2009). Homologs of TfLURE peptides have been identified in Arabidopsis, namely, AtLURE1 (
Higashiyama 2010;
Takeuchi and Higashiyama 2012). AtLURE1 peptides also attract the pollen tube; however, knockdown of
AtLURE1 only partially impaired the precise guidance of the pollen tube around the ovule (
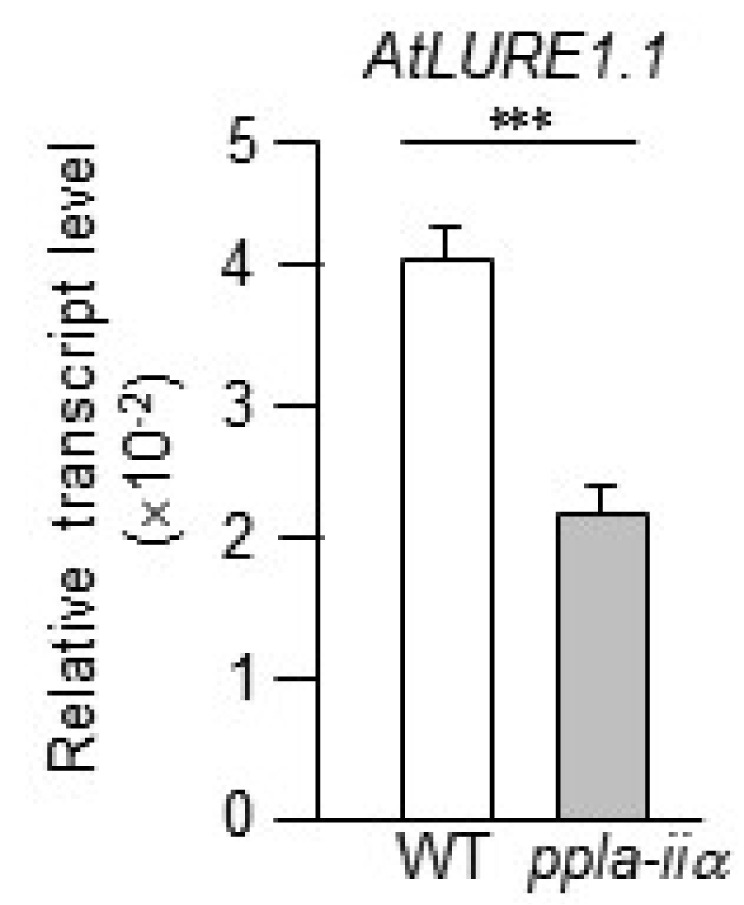
Takeuchi and Higashiyama 2012). Therefore, to determine the effect of pPLA-IIa on the expression of
AtLURE1.1, we compared the expression level of
AtLURE1.1 between WT and
ppla-iia mutant flowers. Results showed that the expression level of
AtLURE1.1 was approximately 2-fold lower in the
ppla-iia mutant than in the WT (
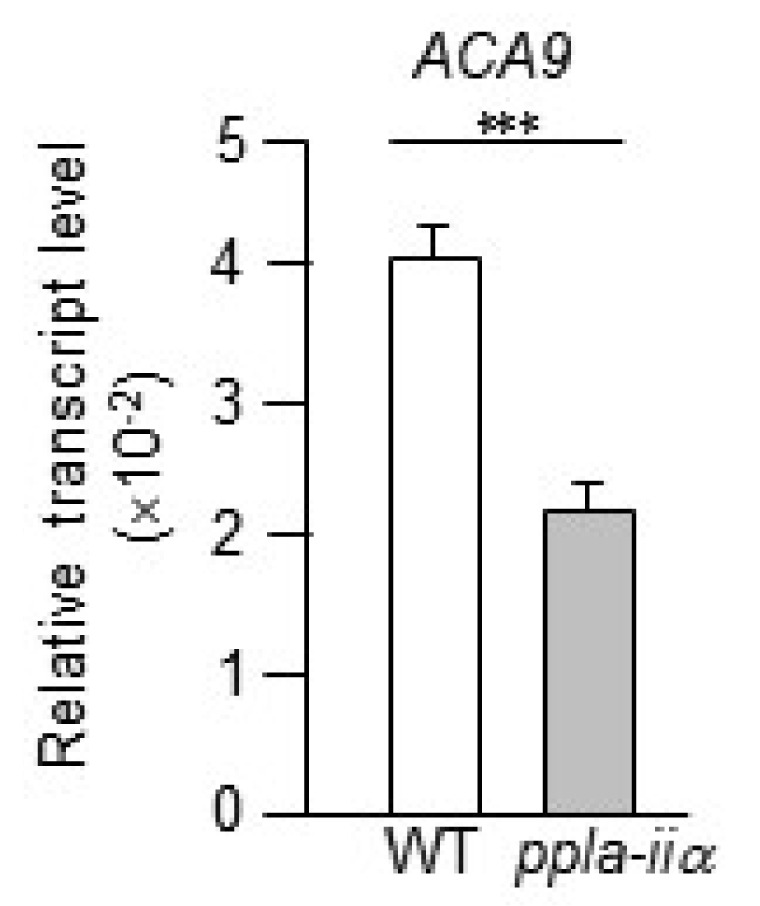
Fig. 4).
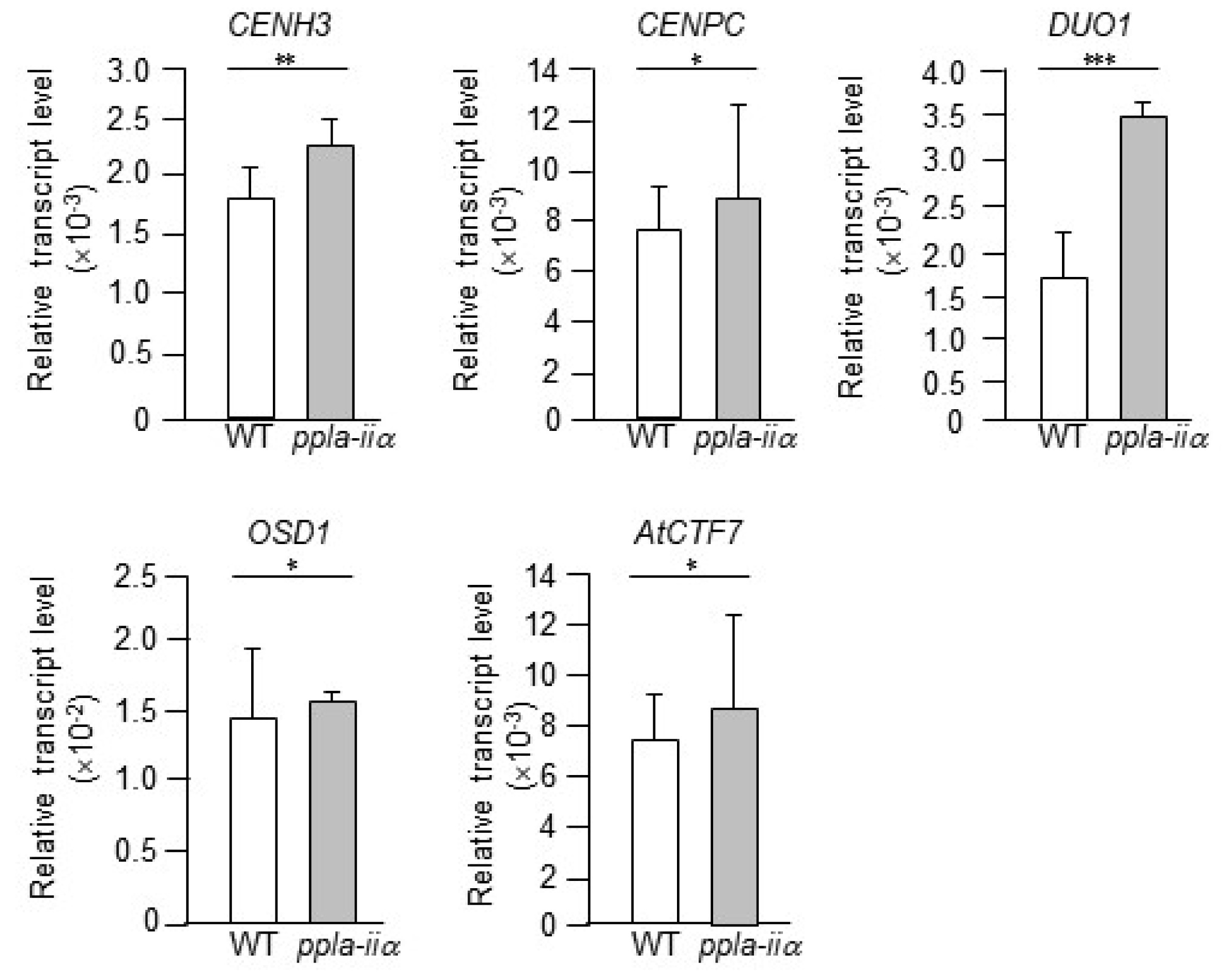
Proper mitosis and meiosis are important for cell division and equal separation of chromosomes between daughter cells. Here, we examined the expression levels of five mitosis- and meiosis-related genes, CENH3, CENPC, DUO1, AtCTF7, and OSD1, in WT and ppla-iia mutant flowers.
In Arabidopsis, haploid lines could be obtained by modifying CENH3 (
Ravi and Chan 2010;
Ravi et al. 2010). In addition, mutagenizing the histone fold domain of CENH3 causes excellent haploid induction upon pollination by wild-type pollen, although the growth and fertility of the DH are affected (
Kuppu et al. 2020). CENPC is an essential kinetochore component required for accurate cell division in yeast and mammals. Arabidopsis CENPC proteins display very similar characteristics and localization patterns as their homologs in other plant species (
Dawe et al. 1999;
Ogura et al. 2004;
Du et al. 2010). In addition,
CENPC knockdown has been reported to cause cell cycle arrest (
Burgos-Rivera and Dawe 2012) as well as chromosome misalignment and segregation defects during mitosis (
Shono et al. 2015). DUO1 is an R2R3-MYB transcription factor that plays a key role in the development of the male germline in Arabidopsis (
Durbarry et al. 2005;
Rotman et al. 2005). DUO1 is responsible for the transcriptional control of genes that comprise a regulatory network and encode mitotic cyclins required for entry into pollen mitosis II and other proteins required for sperm cell adhesion and fertilization (
Brownfield et al. 2009;
Borg et al. 2011;
Mori et al. 2014). OSD1 harbors three putative cell cycle-related domains and is involved in the transition from meiosis I to meiosis II (
Cromer et al. 2012). The
osd1 mutants fail to enter the second meiotic division in both male and female gametophytes, resulting in functional 2n gametes and tetraploid progeny (d’Erfurth
et al. 2009).
In addition, Arabidopsis OSD1 protein acts as a mitotic APC/C inhibitor (
Iwata et al. 2011) and consequently promotes meiotic progression (
Cromer et al. 2012). AtCTF7 functions as an acetyltransferase. While the
ctf7 T-DNA insertion mutant exhibits poor anther development and sterility and defective chromosome segregation during mitosis (
Bolaños-Villegas et al. 2013), RNA interference (RNAi) lines of AtCTF7 display chromosome fragmentation and the loss of sister chromatid cohesion during meiosis (
Singh et al. 2013), indicating that AtCTF7 is required for both mitosis and meiosis. We thus examined the effect of pPLA-IIa on the expression of these genes using total RNA isolated from the flowers of WT and
ppla-iia plants. The results of qRT-PCR analysis showed that the expression level of
DUO1 was increased by approximately 2-fold, whereas
CENH3,
CENPC,
AtCTF7, and
OSD1 were upregulated only slightly in the
ppla-iia mutant compared with the WT (
Fig. 5).
Calcium ions (Ca
2+) are implicated in many physiological processes, including pollen tube tip growth and fertili-zation. The gene encoding calmodulin (CaM)-activated Ca
2+-ATPase ion pump 9 (ACA9) is expressed primarily in pollen, and the ACA9 protein is localized to the plasma membrane (
Schiøtt et al. 2004). Disruption of the ACA9 gene decreases the growth rate and growth potential of pollen tubes and increases the rate of abortion (
Schiøtt et al. 2004), indicating that ACA9 functions as a key regulator of pollen tube growth and fertilization. In this study, qRT-PCR analysis revealed that the expression of
ACA9 was downregulated in
ppla-iia mutant flowers by approximately 2-fold compared with WT flowers (
Fig. 6).
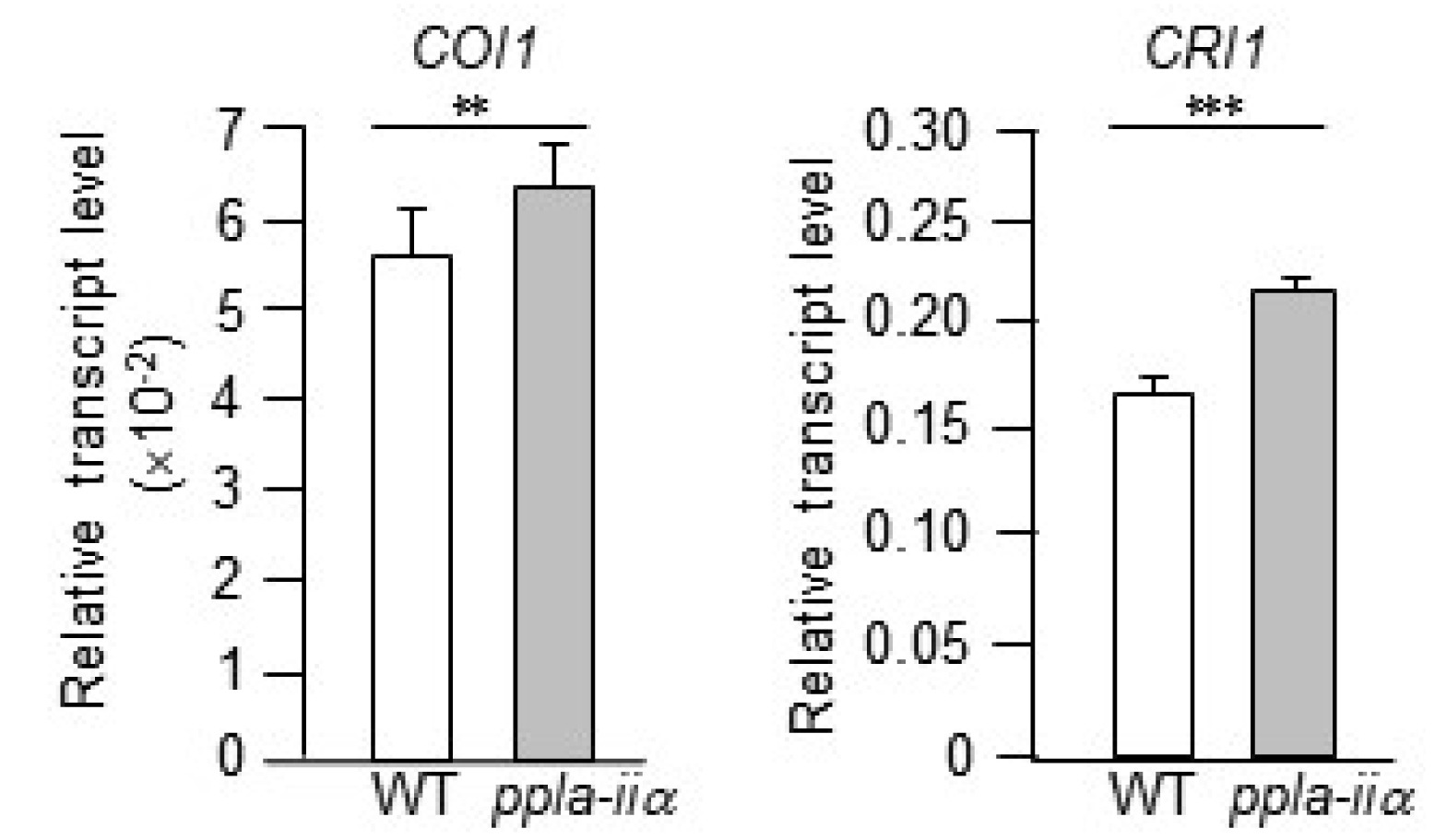
Jasmonate (JA) signaling is essential for several en-vironmental stress responses and reproductive develop-ment in many plant species. In Arabidopsis, mutations in genes encoding JA biosynthesis enzymes and JA receptors result in sporophytic male sterility, characterized by the failure of stamen filament elongation, severe delay in anther dehiscence, and decline in pollen viability (
McConn and Browse 1996). AtCOI1 is an F-box protein and an essential component of the JA co-receptor complex (
Sheard et al. 2010). The coi1 null mutant is male sterile and does not produce seeds because of non-germinating pollen, anthers dehiscence, and insufficient elongation of stamen filaments (
Feys et al. 1994;
Xie et al. 1998).
Brassinosteroids (BRs) are a class of polyhydroxysteroids, which are perceived by the cell surface receptor BRI1. BRs regulate a wide range of physiological processes, including plant growth, development, and immunity, and promote stem elongation and cell division (
Grove et al. 1979;
Bajguz 2007). The
bri1 mutant produces only appro-ximately 20% pollen per anther compared with the wild type, and fails to release pollen upon anther dehiscence (
Ye et al. 2010). Expression analysis of
COI1 and
BRI1 in this study showed that both genes were slightly upregulated in
ppla-iia mutant flowers compared with WT flowers (
Fig. 7).
DISCUSSION
Therefore, in this study, we attempted to identify the possible function of Arabidopsis pPLA-IIa during pollen growth, pollen tube development, and fertilization. We isolated the
ppla-iia T-DNA insertion mutant, and examined the expression of genes involved in pollen growth and pollen tube development, and fertilization by qRT-PCR. We first analyzed the expression of genes involved in pollen tube growth, as it is the first step in the fertilization process (
Kost et al. 1999;
Li et al. 1999; Baumberger
et al. 2001;
Fu et al. 2001;
Baumberger et al. 2003;
Liang et al. 2013;
Morato et al. 2014;
Mecchia et al. 2017;
Franck et al. 2018;
Sede et al. 2018). Unexpectedly, the expression of these genes was upregulated in the
ppla-iia mutant (
Fig. 2). Then, we examined the expression of pollen receptor genes because pollen guidance and reception are important for normal fertilization (
Lee et al. 1996;
Muschietti et al. 1998;
Zhang et al. 2008;
Boisson-Dernier et al. 2009;
Miyazaki et al. 2009;
Salem et al. 2011; Boisson-Dernier
et al. 2013;
Liu et al. 2013;
Gui et al. 2014;
Takeuchi and Higashiyama 2016;
Wang et al. 2016). Like pollen tube growth-related genes, the expression of pollen receptor genes was also upregulated in the
ppla-iia mutant (
Fig. 3). Because pollen tube attraction is an important step in the fertilization process (
Wilhelmi and Preuss 1997;
Higashiyama and Hamamura 2008;
Punwani and Drews 2008; Higashiyama
et al. 2009;
Okuda et al. 2009;
Takeuchi and Higashiyama 2012), we analyzed the expression of a gene encoding a synergid-secreted protein and found that its expression was downregulated in the
ppla-iia mutant (
Fig. 4).
At the beginning of this research, we speculated that the expression of all analyzed genes would be downregulated in the
ppla-iia mutant because reduced expression of these genes can cause abnormal or reduced fertilization, thus increasing DH production. However, only the genes involved in pollen tube attraction and Ca
2+ pumping were downregulated in the
ppla-iia mutant, while four other types of genes were upregulated in the mutant. Our data suggest that several proteins are involved in the regulation of pollen tube growth and fertilization, although pPLA-IIa is the major player in both these processes. Very recently, we found that haploid seeds were not produced in
ppla-iia mutants (Kwak
et al. 2012). Therefore, our data also suggest that the haploid production mechanism in monocots is different from that in dicots because only the loss of pPLA-IIa induces the production of haploid seeds in maize and rice (
Gilles et al. 2017;
Kelliher et al. 2017;
Liu et al. 2017;
Yao et al. 2018). Further analyses using
ppla-iia mutants and lines carrying mutations in pPLA-IIa homologous genes are required for elucidating the roles of various types of phospholipases including pPLAs in pollen development, fertilization, and haploid production.
ACKNOWLEDGEMENTS
This work was supported by National Research Foundation of Korea Grant funded by the Korean Government (Project No. 2021R1A2C1003446). This work was also carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01567701)” Rural Development Administration, Republic of Korea.
Fig. 1Phenotypes of WT and ppla-iia mutant plants. Seeds of WT and ppla-iia mutants were germinated on MS media or in the soil and then photographed at germination, vegetative growth and flowering stages.

Fig. 2Evaluation of the effect of pPLA-IIa on the expression of pollen tube growth-regulating genes. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and expression levels of AtRALF4, AUN1, LRX10, AtMYB97, and ROP1 were analyzed by real-time qRT-PCR using gene-specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
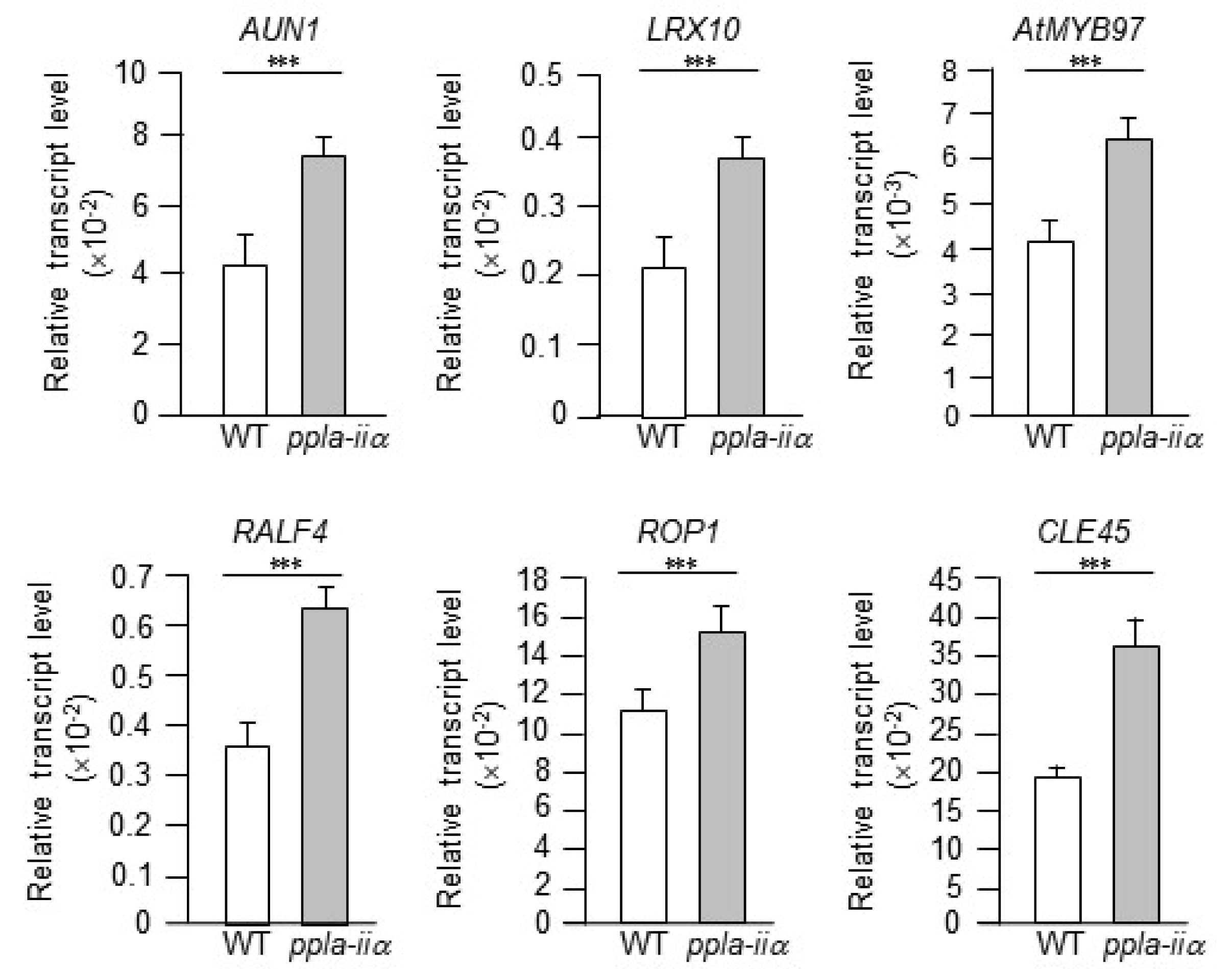
Fig. 3Analysis of the effect of pPLA-IIa on the expression of genes encoding pollen receptors. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and expression levels of LIP1, LIP2, ANX1, ANX2, PRK1, and MDIS1 were analyzed by real-time qRT-PCR using gene-specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
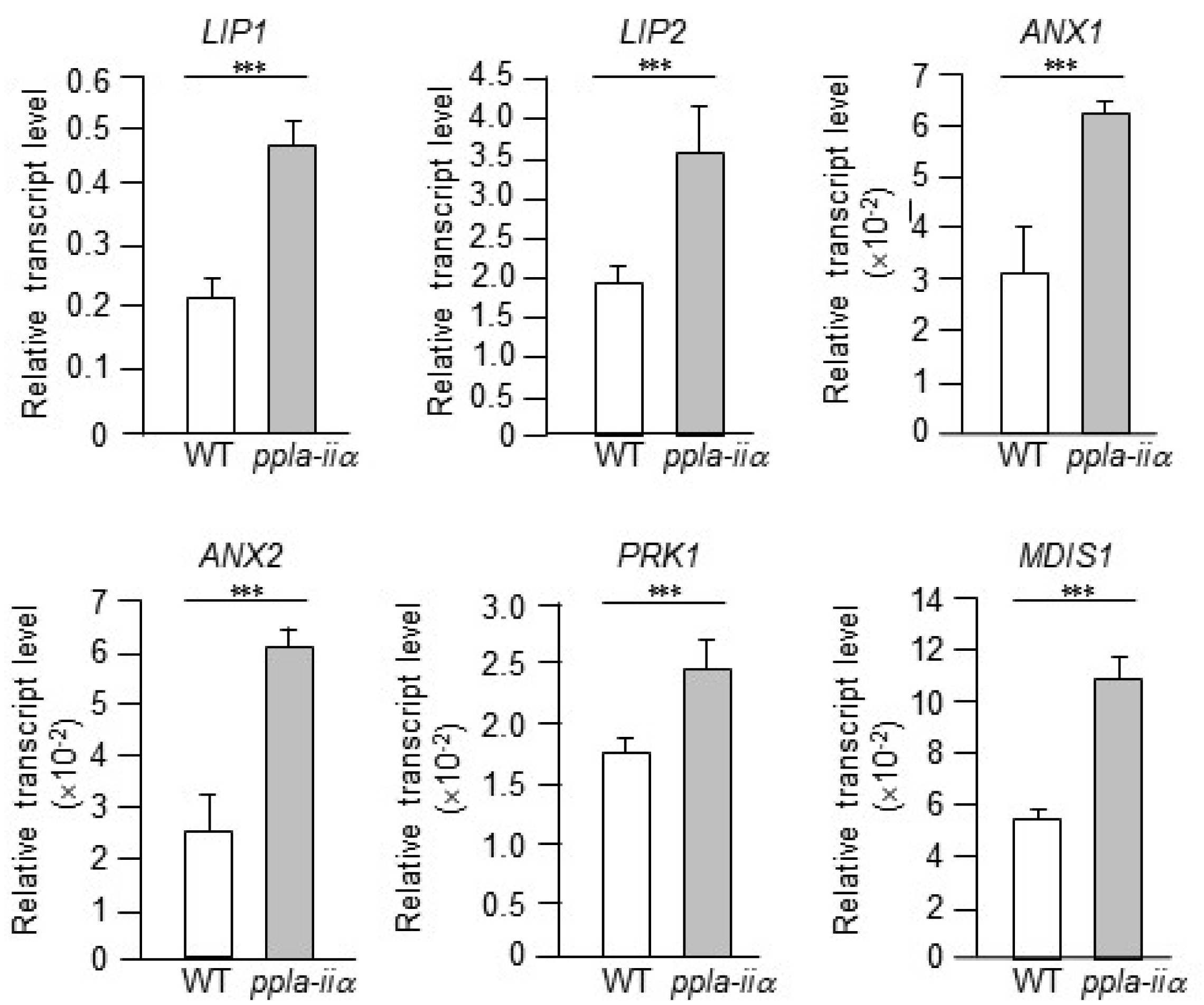
Fig. 4Examination of the effect of pPLA-IIa on the expression of the AtLURE1.1 gene, which encodes a synergid-secreted protein. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and the expression level of AtLURE1.1 was examined by real-time qRT-PCR using gene- specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
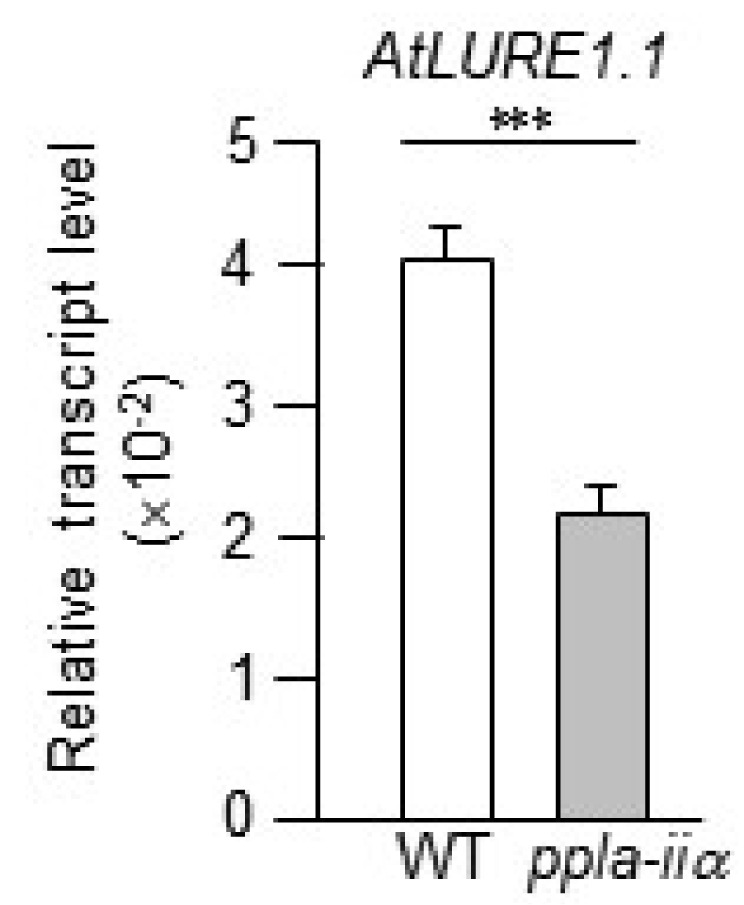
Fig. 5Examination of the effect of pPLA-IIa on the expression of cell division-related genes. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and expression levels of CENH3, CENPC, CTF7, and DUO1 were examined by real-time qRT-PCR using gene-specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (*P < 0.1, **P < 0.01, ***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
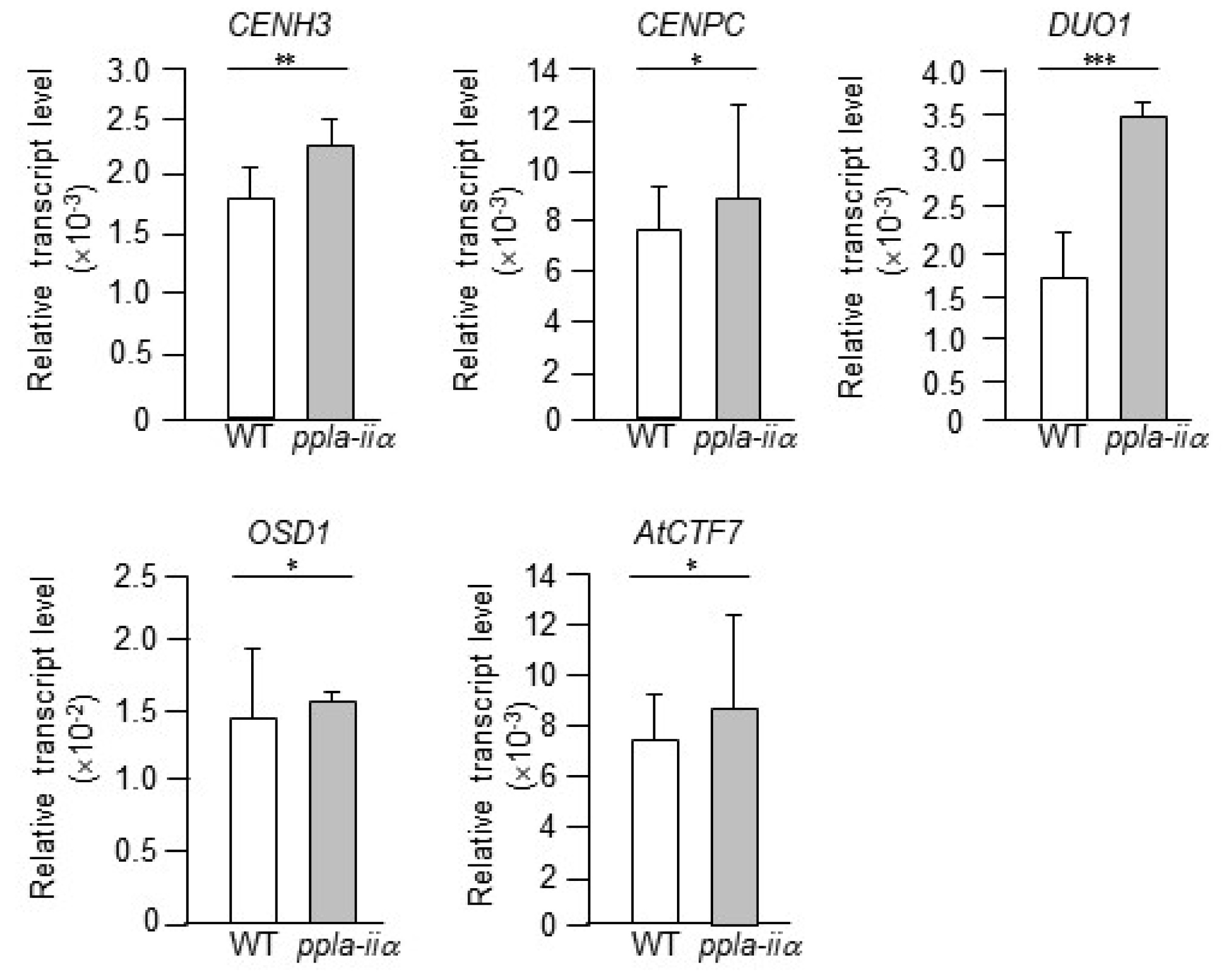
Fig. 6Analysis of the effect of pPLA-IIa on the expression of ACA9, which encodes a Ca2+ pump. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and the expression level of ACA9 was examined by real-time qRT-PCR using-specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
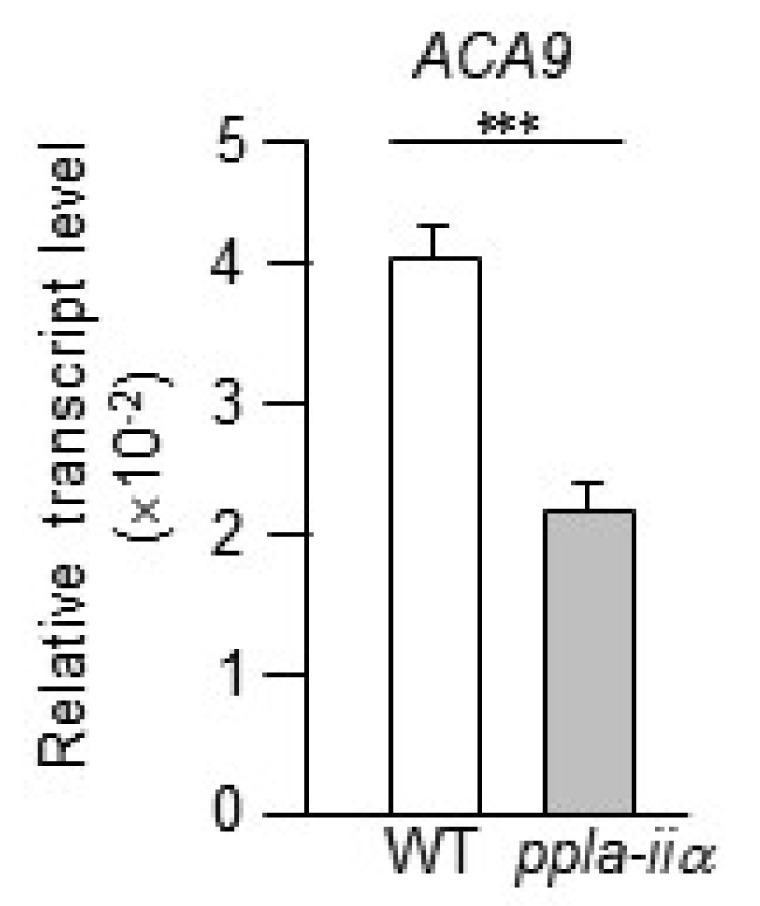
Fig. 7Evaluation of the effect of pPLA-IIa on the expression of hormone-related genes. Total RNA was isolated from the flowers of WT and ppla-iia mutant plants, and expression levels of COI1 and BRI1 were examined by real-time qRT-PCR using gene-specific primers. Data shown are means ± SE from three biological replications and three technical repeats in each. Asterisks indicate statistically significant differences in grain weight (**P < 0.01, ***P < 0.001; Student’s t-test) between WT and ppla-iia mutant plants.
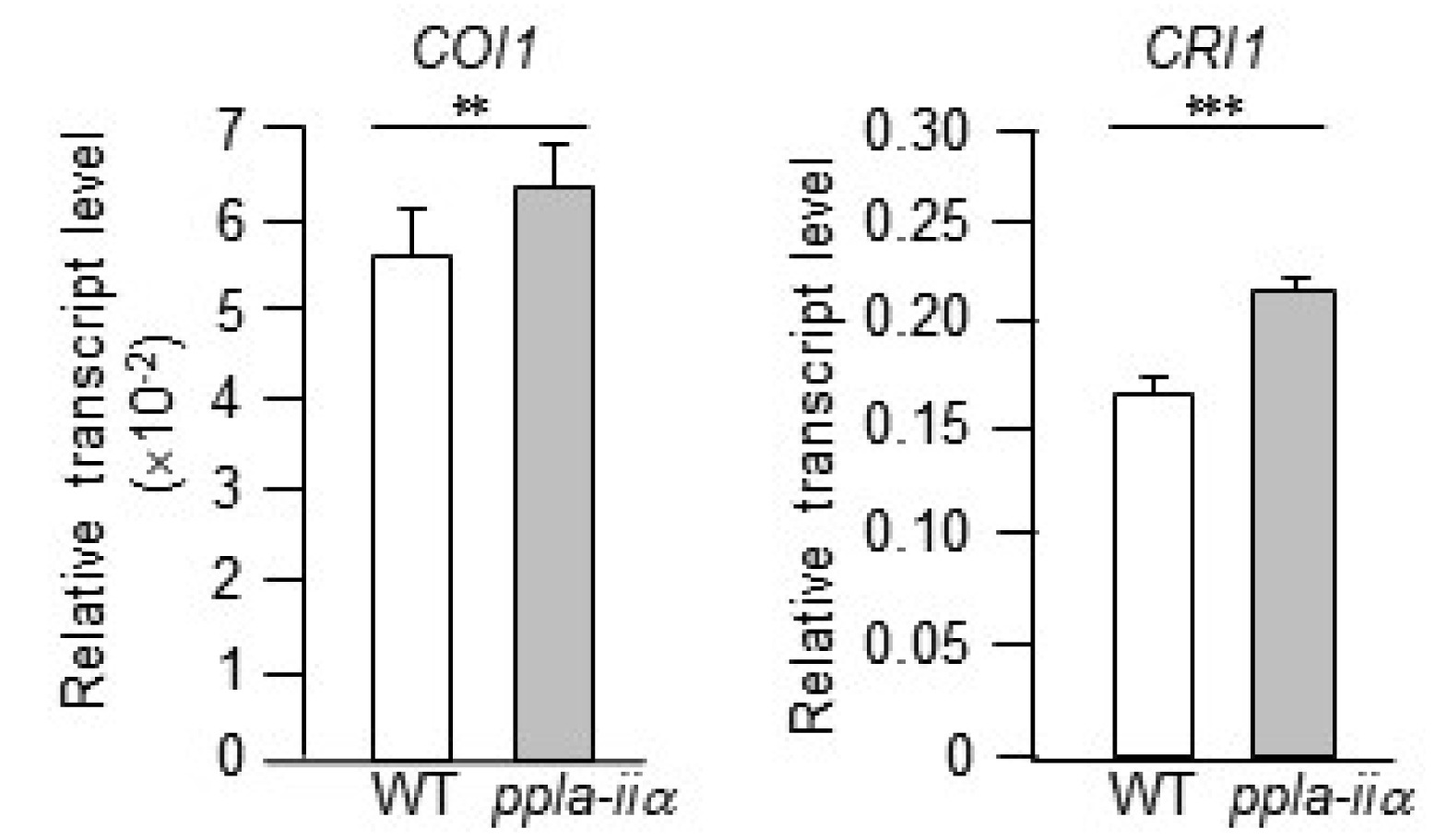
Table 1List of primers used in this study.
Table 1
|
Category |
Gene |
Forward primer |
Reverse primer |
PCR product (bp) |
|
Ion pump |
ACA9 |
5’-GCAGGCTTAAGCATTCTTGG-3’ |
5’-TTCTTGCGTTGAACTCGTTG-3’ |
120 |
|
Receptor |
LIP1 |
5’-TGAAACAAACGCAACTGCTC-3’ |
5’-CGTGATCTCTACCGCTGTCA-3’ |
149 |
|
LIP2 |
5’-GAGCTTATCACGGGTCGAAG-3’ |
5’-GATGCTTCCTCTTGCACACA-3’ |
200 |
|
ANX1 |
5’-GAGGCGACTCTCACACATCA-3’ |
5’-CAATGACGTTGGAATCATCG-3’ |
192 |
|
ANX2 |
5’-GCAGGCTTAAGCATTCTTGG-3’ |
5’-GCAGGCTTAAGCATTCTTGG-3’ |
146 |
|
MDIS1 |
5’-ATGGGCGTCAAAGTATCTGG-3’ |
5’-GGTCTTTGGCTTTCGTCAAG-3’ |
138 |
|
PRK1 |
5’-GGATTTAGCGAGTTGGGTGA-3’ |
5’-TCTTCTCCACAGCCTGTCCT-3’ |
182 |
|
Attractant |
LURE1.1 |
5’-CCTTTCGATCCACGTTCACT-3’ |
5’-GCAACGACAAAGTTTTCTTTCAA-3’ |
135 |
|
RALF4 |
5’-CATTGGCTACGATGCTCTCA-3’ |
5’-CATCCACGCCTGTAAGGATT-3’ |
108 |
|
CLE45 |
5’-ACGCGTTCTCGAGGAAGTTA-3’ |
5’-GGCTGAGCTTTGTTGTGGAT-3’ |
141 |
|
Activator |
AUN1 |
5’-GAGGCCATCAGGTAGTGGAA-3’ |
5’-ACCGGCATTGTCAAACTCTC-3’ |
104 |
|
LRX10 |
5’-CGGACAAGGTGATTCATGTG-3’ |
5’-ACTGGACGGCTAATCACCAC-3’ |
129 |
|
MYB97 |
5’-GCTTCTTGACTGGGGAAGTG-3’ |
5’-TGGCAAGTTGTTGGTGTCAT-3’ |
139 |
|
ROP1 |
5’-TTCAGTGCCAATGTTGTGGT-3’ |
5’-TGCTCCACGGTAACTCAGTG-3’ |
108 |
|
Meiosis gene |
DUO1 |
5’-TGGTTTGGTTGAGGAAGAGG-3’ |
5’-AGCGAAAGGCTGATGCTCTA-3’ |
133 |
|
CENH3 |
5’-AGAGATTCGCCATTTCCAGA-3’ |
5’-AGCTTCAGCTGTCCAACGAT-3’ |
124 |
|
CENPC |
5’-TCTTACGCATGAGGGGAAAC-3’ |
5’-TCGAGCGGTCTCGACTTAAT-3’ |
111 |
|
CTF7 |
5’-TTTTAAGGGTTGGCAGAACG-3’ |
5’- GTTTCTGTGTGCAGGGGAAT -3’ |
100 |
|
OSD1 |
5’-TGGCCGAGTTATTGAGATCC-3’ |
5’-GCTTGAATCCCACAGATGGT-3’ |
119 |
|
Hormone related gene |
COI1 |
5’-CTTCCGCCTTGTCTTACTCG-3’ |
5’-GTTAAGCCGCCTTGTCTCAG-3’ |
132 |
|
BRI1 |
5’-ACGAGCGGCTTCTTGTGTAT-3’ |
5’-GCCGTGTGGACCAGTTTAGT-3’ |
102 |
References
- Bajguz A. 2007. Metabolism of brassinosteroids in plants. Plant Physiol. Biochem.. 45(2): 95-107.
- Baumberger N, Doesseger B, Guyot R, Diet A, Parsons RL, Clark MA, et al. 2003. Whole-genome comparison of leucine-rich repeat extensins in Arabidopsis and rice: a conserved family of cell wall proteins form a vegetative and a reproductive clade. Plant Physiol.. 131: 1313-1326.
- Boisson-Dernier A, Roy S, Kritsas K, Grobei MA, Jaciubek M, Schroeder JI, et al. 2009. Disruption of the pollenexpressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development. 136: 3279-3288.
- Bolaños-Villegas P, Yang X, Wang HJ, Juan CT, Chuang MH, Makaroff CA, et al. 2013. Arabidopsis CHROMOSOME TRANSMISSION FIDELITY 7 (AtCTF7/ECO1) is required for DNA repair, mitosis and meiosis. Plant J.. 75(6): 927-940.
- Borg M, Brownfield L, Khatab H, Sidorova A, Lingaya M, Twell D. 2011. The R2R3 MYB transcription factor DUO1 activates a male germline specific regulon essential for sperm cell differentiation in Arabidopsis. Plant Cell. 23: 534-549.
- Brownfield L, Hafidh S, Borg M, Sidorova A, Mori T, Twell D. 2009. A plant germline-specific integrator of sperm specification and cell cycle progression. PLoS Genet.. 5: e1000430
- Burgos-Rivera B, Dawe RK. 2012. An Arabidopsis Tissue-Specific RNAi Method for Studying Genes Essential to Mitosis. PLoS One. 7(12): e51388
- Cromer L, Heyman J, Touati S, Harashima H, Araou E, Girard C, et al. 2012. OSD1 promotes meiotic pro-gression via APC/C inhibition and forms a regulatory network with TDM and CYCA1; 2/TAM. PLoS Genet.. 8(7): e1002865
- Jolivet S, Froger N, Catrice O, Novatchkova M, Mercier R. d' Erfurth I2009. Turning meiosis into mitosis. PLoS Biol.. 7: e1000124
- Dawe RK, Reed LM, Yu HG, Muszynski MG, Hiatt EN. 1999. A maize homolog of mammalian CENPC is a constitutive component of the inner kinetochore. Plant Cell. 11: 1227-1238.
- Dessen A. 2000. Structure and mechanism of human cytosolic phospholipase A2. Biochim. Biophys. Acta. 1488: 40-7.
- Du Y, Topp CN, Dawe RK. 2010. DNA binding of Centromere Protein C (CENPC) is stabilized by single-stranded RNA. PLoS Genet.. 6(2): e1000835
- Durbarry A, Vizir I, Twell D. 2005. Male germ line development in Arabidopsis. duo pollen mutants reveal gametophytic regulators of generative cell cycle progression. Plant Physiol.. 137: 297-307.
- Dwivedi SL, Britt AB, Tripathi L, Sharma S, Upadhyaya HD, Ortiz R. 2015. Haploids: constraints and opportunities in plant breeding. Biotechnol. Adv.. 33: 812-829.
- Effendi Y, Radatz K, Labusch C, Rietz S, Wimalasekera R, Helizon H, et al. 2014. Mutants of phospholipase A (pPLA-I) have a red light and auxin phenotype. Plant Cell Environ.. 37: 1626-1640.
- Feys B, Benedetti C, Penfold C, Turner J. 1994. Arabidopsis mutants selected for resistance to the phytotoxin coronatine are male sterile, insensitive to methyl jasmonate, and resistent to a bacterial pathogen. Plant Cell. 6: 751-759.
- Franck CM, Westermann J, Bürssner S, Lentz R, Lituiev DS, Boisson-Dernier A. 2018. The protein phosphatases ATUNIS1 and ATUNIS2 regulate cell wall integrity in tip-growing cells. Plant Cell. 30(8): 1906-1923.
- Fu Y, Wu G, Yang Z. 2001. Rop GTPase-dependent dynamics of tip-localized F-actin controls tip growth in pollen tubes. J Cell Biol.. 152: 1019-1032.
- Ghosh M, Tucker DE, Burchett SA, Leslie CC. 2006. Properties of the Group IV phospholipase A2 family. Prog. Lipid Res.. 45: 487-510.
- Gilles LM, Khaled A, Laffaire JB, Chaignon S, Gendrot G, Laplaige J, et al. 2017. Loss of pollen-specific phospholipase NOT LIKE DAD triggers gynogenesis in maize. EMBO J.. 36: 707-717.
- Grove MD, Spencer GF, Rohwedder WK, Mandava N, Worley JF, David Warthen J, et al. 1979. Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen. Nature. 281(5728): 216-217.
- Gui CP, Dong X, Liu HK, Huang WJ, Zhang D, Wang SJ, et al. 2014. Overexpression of the tomato pollen receptor kinase LePRK1 rewires pollen tube growth to a blebbing mode. Plant Cell. 26: 3538-3555.
- Higashiyama T, Hamamura Y. 2008. Gametophytic pollen tube guidanc. Sex. Plant Reprod.. 21: 17-26.
- Higashiyama T, Yabe S, Sasaki N, Nishimura Y, Miyagishima S, Kuroiwa H, et al. 2001. Pollen tube attraction by the synergid cell. Science. 293: 1480-1483.
- Higashiyama T. 2010. Peptide signaling in pollen-pistil interactions. Plant Cell Physiol.. 51: 177-189.
- Holk A, Rietz S, Zahn M, Quader H, Scherer GF. 2002. Molecular identification of cytosolic, patatin related phospholipases A from Arabidopsis with potential functions in plant signal transduction. Plant Physiol.. 130: 90-101.
- Iwata E, Ikeda S, Matsunaga S, Kurata M, Yoshioka Y, Criqui MC, et al. 2011. GIGAS CELL1, a novel negative regulator of the anaphase-promoting complex/cyclosome, is required for proper mitotic progression and cell fate determination in Arabidopsis. Plant cell. 23: 4382-4393.
- Kelliher T, Starr D, Richbourg L, Chintamanani S, Delzer B, Nuccio ML, et al. 2017. MATRILINEAL, a sperm-specific phospholipase, triggers maize haploid induction. Nature. 542: 105-109.
- Kost B, Lemichez E, Spielhofer P, Hong Y, Tolias K, Carpenter C, et al. 1999. Rac homologues and compartmentalized phosphatidylinositol 4,5-bisphosphate act in a common pathway to regulate polar pollen tube growth. J. Cell Biol.. 145: 317-330.
- Kuppu S, Ron M, Marimuthu MPA, Li G, Huddleson A, Siddeek MH, et al. 2020. A variety of changes, including CRISPR/Cas9-mediated deletions, in CENH3 lead to haploid induction on outcrossing. Plant Biotechnol. J.. 18(10): 2068-2080.
- Kwak JS, Kwon DH, Song JT, Seo HS. 2021. A mutation in the pPLA-IIa gene encoding PATATIN-RELATED PHOSPHOLIPASE A causes late flowering in Arabidopsis. Biochem. Biophys. Res. Commun.. 582: 16-20.
- Labusch C, Shishova M, Effendi Y, Li M, Wang X, Scherer GFE. 2013. Patterns and timing in expression of early auxin-induced genes imply involvement of phospho-lipases A (pPLAs) in the regulation of auxin responses. Mol. Plant. 6: 1473-1486.
- Lee HS, Karunanandaa B, McCubbin A, Gilroy S, Kao TH. 1996. PRK1, a receptor-like kinase of Petunia inflata, is essential for postmeiotic development of pollen. Plant J.. 9: 613-624.
- Li H, Lin Y, Heath RM, Zhu MX, Yang Z. 1999. Control of pollen tube tip growth by a Rop GTPase-dependent pathway that leads to tip-localized calcium influx. Plant Cell. 11: 1731-1742.
- Li M, Bahn SC, Fan C, Li J, Phan T, Oritiz M, et al. 2013. Patatin-related phospholipase pPLAIII delta increases seed oil content with long-chain fatty acids in Arabidopsis. Plant Physiol.. 162: 39-51.
- Liang Y, Tan ZM, Zhu L, Niu QK, Zhou JJ, Li M, et al. 2013. MYB97, MYB101 and MYB120 function as male factors that control pollen tube-synergid interaction in Arabidopsis thaliana fertilization. PLoS Genet.. 9(11): e1003933
- Liu C, Li X, Meng D, Zhong Y, Chen C, Dong X, et al. 2017. A 4- bp insertion at ZmPLA1 encoding a putative phospholipase A generates haploid induction in maize. Mol. Plant. 10: 520-522.
- Liu J, Zhong S, Guo X, Hao L, Wei X, Huang Q. 2013. Membrane-bound RLCKs LIP1 and LIP2 are essential male factors controlling male-female attraction in Arabidopsis. Curr. Biol.. 23(11): 993-998.
- McConn M, Browse J. 1996. The critical requirement for linolenic acid is pollen development, not photosynthesis, in an Arabidopsis mutant. Plant Cell. 8: 403-416.
- Mecchia MA, Santos-Fernandez G, Duss NN, Somoza SC, Boisson-Dernier A, Gagliardini V, et al. 2017. RALF4/19 peptides interact with LRX proteins to control pollen tube growth in Arabidopsis. Science. 358(6370): 1600-1603.
- Meijer HJG, Munnik T. 2003. Phospholipid-based signaling in plants. Annu. Rev. Plant Biol.. 54: 265-306.
- Miyazaki S, Murata T, Sakurai-Ozato N, Kubo M, Demura T, Fukuda H, et al. 2009. ANXUR1 and 2, sister genes to FERONIA/SIRENE, are male factors for coordinated fertilization. Curr. Biol.. 19: 1327-1331.
- Cesiliato PH, Ribeiro B, Ortiz Morea FA, Franco Garcia AA, Silva-Filho MC, et al. Morato do Canto A2014. Biological activity of nine recombinant AtRALF peptides: implications for their perception and function in Arabidopsis. Plant Physiol. Biochem.. 75: 45-54.
- Mori T, Igawa T, Tamiya G, Miyagishima SY, Berger F. 2014. Gamete attachment requires GEX2 for successful fertilization in Arabidopsis. Curr. Biol.. 24: 170-175.
- Muschietti J, Eyal Y, McCormick S. 1998. Pollen tube localization implies a role in pollen-pistil interactions for the tomato receptor-like protein kinases LePRK1 and LePRK2. Plant Cell. 10: 319-330.
- Ogura Y, Shibata F, Sato H, Murata M. 2004. Characteri-zation of a CENP-C homolog in Arabidopsis thaliana. Genes Genet. Syst.. 79: 139-144.
- Okuda S, Tsutsui H, Shiina K, Sprunck S, Takeuchi H, Yui R, et al. 2009. Defensin-like polypeptide LUREs are pollen tube attractants secreted from synergid cells. Nature. 458: 357-361.
- Park BS, Song JT, Seo HS. 2011. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat. Commun.. 2: 400
- Punwani JA, Drews GN. 2008. Development and function of the synergid cell. Sex. Plant Reprod.. 21: 7-15.
- Ravi M, Chan SWL. 2010. Haploid plants produced by centromere-mediated genome elimination. Nature. 46: 615-618.
- Ravi M, Kwong PN, Menorca RMG, Valencia JT, Ramahi JS, Stewart JL, et al. 2010. The rapidly evolving centromere- specific histone has stringent functional requirements in Arabidopsis thaliana. Genetics. 186: 461-471.
- Rietz S, Dermendjiev G, Oppermann E, Tafesse FG, Effendi Y, Holk A, et al. 2010. Roles of Arabidopsis patatin-related phospholipases A in root development are related to auxin responses and phosphate deficiency. Mol. Plant. 3: 524-538.
- Rotman N, Durbarry A, Wardle A, Yang WC, Chaboud A, Faure JE, et al. 2005. A novel class of MYB factors controls sperm cell formation in plants. Curr. Biol.. 15: 244-248.
- Rydel TJ, Williams JM, Krieger E, Moshiri F, Stallings WC, Brown SM, et al. 2003. The crystal structure, mutagenesis, and activity studies re-veal that patatin is a lipid acyl hydrolase with a Ser-Asp catalytic dyad. Biochemistry. 42: 6696-6708.
- Ryu SB. 2004. Phospholipid-derived signaling mediated by phospholipase A in plants. Trends Plant Sci.. 9: 229-235.
- Salem T, Mazzella A, Barberini ML, Wengier D, Motillo V, Parisi G, et al. 2011. Mutations in two putative phosphorylation motifs in the tomato pollen receptor kinase LePRK2 show antagonistic effects on pollen tube length. J Biol. Chem.. 286: 4882-4891.
- Scherer GF, Pietrzyk P. 2014. Gravity-dependent differen-tiation and root coils in Arabidopsis thaliana wild type and phospholipase-A-I knockdown mutant grown on the International Space Station. Plant Biol. 16(Suppl 1). 1: 97-106.
- Scherer GF, Ryu SB, Wang X, Matos AR, Heitz T. 2010. Patatin-related phospholipaseA: nomenclature, subfamilies and functions in plants. Trends Plant Sci. 15: 693-700.
- Scherer GF. 1996. Phospholipid signalling and lipid-derived second messengers in plants. Plant Growth Regul.. 18: 125-133.
- Schiøtt M, Romanowsky SM, Baekgaard L, Jakobsen MK, Palmgren MG, Harper JF. 2004. A plant plasma membrane Ca2+ pump is required for normal pollen tube growth and fertilization. Proc. Natl. Acad. Sci. USA.. 101(25): 9502-9527.
- Sede AR, Borassi C, Wengier DL, Mecchia MA, Estevez JM, Muschietti JP. 2018. Arabidopsis pollen extensins LRX are required for cell wall integrity during pollen tube growth. FEBS Lett.. 592(2): 233-243.
- Senda K, Yoshioka H, Doke N, Kawakita K. 1996. A cytosolic phospholipase A2 from potato tissues appears to be patatin. Plant Cell Physiol.. 37: 347-353.
- Sheard LB, Tan X, Mao H, Withers J, Ben-Nissan G, Hinds TR, et al. 2010. Jasmonate perception by inositol- phosphate-potentiated COI1-JAZ co-receptor. Nature. 468: 400-405.
- Shewry PR. 2003. Tuber storage proteins. Ann. Bot.. 91: 755-769.
- Shimizu T, Ohto T, Kita Y. 2006. Cytosolic phospholipaseA2: Biochemical properties and physiological roles. IUBMB. Life. 58: 328-333.
- Shono N, Ohzeki J, Otake K, Martins NM, Nagase T, Kimura H, et al. 2015. CENP-C and CENP-I are key connecting factors for kinetochore and CENP-A assembly. J. Cell Sci.. 128: 4572-4587.
- Singh DK, Andreuzza S, Panoli AP, Siddiqi I. 2013. AtCTF7 is required for establishment of sister chromatid cohesion and association of cohesin with chromatin during meiosis in Arabidopsis. BMC Plant Biol.. 13: 117
- Six DA, Dennis EA. 2000. The expanding superfamily of phospholipase A 2 enzymes: classification and charac-terization. Biochim. Biophys. Acta-Mol. Cell Biol. Lipid. 1488: 1-19.
- Takeuchi H, Higashiyama T. 2012. A species-specific cluster of defensin-like genes encodes diffusible pollen tube attractants in Arabidopsis. PLoS Biol.. 10: e1001449
- Takeuchi H, Higashiyama T. 2016. Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature. 531: 245-248.
- Wang T, Liang L, Xue Y, Jia PF, Chen W, Zhang MX, et al. 2016. A receptor heteromer mediates the male perception of female attractants in plants. Nature. 531: 241-244.
- Wilhelmi LK, Preuss D. 1997. Blazing new trails (Pollen tube guidance in flowering plants). Plant Physiol.. 113: 307-312.
- Xie DX, Feys B, James S, Nieto-Rostro M, Turner J. 1998. COI1: An Arabidopsis gene required for jasmonate- regulated defense and fertility. Science. 280: 1091-1094.
- Yang W, Devaiah SP, Pan X, Isaac G, Welti R, Wang X. 2007. AtPLAI is an acyl hydrolase involved in basal jasmonic acid production and Arabidopsis resistance to Botrytis cinerea. J. Biol. Chem.. 282: 18116-18128.
- Yang WY, Zheng Y, Bahn SC, Pan XQ, Li MY, Vu HS, et al. 2012. The patatin-containing phospholipase ApPLAIIa modulates oxylipin formation and water loss in Arabidopsis thaliana. Mol. Plant. 5: 452-460.
- Yao L, Zhang Y, Liu C, Liu Y, Wang Y, Liang D, et al. 2018. OsMATL mutation induces haploid seed formation in indica rice. Nat. Plants. 4: 530-533.
- Ye Q, Zhu W, Li L, Zhang S, Yin Y, Ma H, Wang X. 2010. Brassinosteroids control male fertility by regulating the expression of key genes involved in Arabidopsis anther and pollen development. Proc. Natl. Acad. Sci. USA. 107 (13): 6100-6105..
- Zhang D, Wengier D, Shuai B, Gui CP, Muschietti J, McCormick S, et al. 2008. The pollen receptor kinase LePRK2 mediates growth-promoting signals and positively regulates pollen germination and tube growth. Plant Physiol.. 148: 1368-1379.
- Zhou J, Rumeau D, Showalter AM. 1992. Isolation and characterization of two wound-regulated tomato extensin genes. Plant Mol. Biol.. 20: 5-17.