Abstract
Genetic analysis of genes that regulate the color pigmentation of sterile lemma and apiculus has been conducted. “Josaengjado” has small and round grains with purple leaf, sterile lemma and apiculus. In the F2 population from a cross between Josaengjado and Daeribbyeo 1, 246 and 182 plants exhibited purple and straw-white sterile lemma, respectively. It fitted a 9:7 segregation ratio indicating that two complementary genes control the pigmentation in sterile lemma and apiculus. Genetic analysis was performed using the F2:3 and KASP (Kompetitive Allele-Specific PCR) markers. Genes for the coloration of leaf sheath, ligule, sterile lemma, and apiculus were detected on chromosomes 1 and 6. Sequence comparison showed a single nucleotide substitution C (Josaengjado) to A (Daeribbyeo 1) in the second exon of the Rd gene on chromosome 1 leading to a premature stop in Daeribbyeo 1. In C1, a 3-bp deletion in the second exon was detected in Daeribbyeo 1. Haplotype analysis was performed in the Rd and C1 genes of the 78 rice accessions. 78 accessions were divided into 14 groups. A total of 11 and 1 mutation sites were detected in OsC1 and Rd, respectively. The haplotype analysis also confirmed that two complementary genes, Rd and OsC1 are necessary to express anthocyanin pigmentation in sterile lemma and apiculus. To our knowledge, this is the first report to identify genes for the coloration of sterile lemma in rice.
-
Key words: Sterile lemma color, Apiculus color, KASP marker, Rice
INTRODUCTION
Pigmentation in plant organs is prevalent in wild rice, but the phenotype shows variation in cultivated rice (
Li and Chen 1993;
Saitoh et al. 2004). Various colors such as black hull, red pericarp, purple awn, purple apiculus and purple leaf margin are observed in wild rice, while most cultivars have already lost color in plant organs in the domestication process (
Sun et al. 2018). Flavonoid derivatives including anthocyanins, are responsible for a wide range of biological functions including pigmentation pat-terns, protection against UV radiation, signal molecules in plant-microbial interactions, and plant defense reactions (
Saitoh et al. 2004;
Lin-Wang et al. 2010).
Studies of anthocyanin pigmentation have been extensively carried out over the last half-century. In rice, previous genetic studies revealed that two genes,
C and
A are fundamental in regulating anthocyanin pigments (
Takahashi 1957). Structural and functional genes associated with anthocyanin biosynthesis in rice have been well characterized, but several regulatory genes are also involved in the biosynthetic pathway (
Furukawa et al. 2007;
Kim et al. 2011;
Shao et al. 2013;
Oikawa et al. 2015).
Pigmentation in the apiculus and sterile lemma was controlled by one basic gene and three complementary genes, respectively (
Siddiq et al. 1986).
C, the essential gene for anthocyanin biosynthesis located on rice chromosome 6 regulates red and purple of apiculus (
Reddy 1996;
Fan et al. 2008). The segregation ratio of purple and green fitted 3:1 in the 430 F
2 plants of the “SSSL W23-07-6-02-14” with purple apiculus and “HJX74” with green apiculus. The
Pa-6 gene was detected through fine mapping (
Liu et al. 2012). Entire
OsC gene and its full-length cDNA cloned from
Japonica landrace “Lijiangxintuanheigu” (LTH) with red apiculus and purple stigma were transformed into
japonica cultivar Kitaake with colorless apiculus and stigmas, all transformants had red apiculus but non-colored stigmas indicating that
OsC alone modulates anthocyanin pigmentation in the apiculus and represented a functional
C gene (
Zhao et al. 2016). The apiculus, the remnant of the awn maintains its color in many cultivars and seems not to have undergone artificial selection throughout domestication (
Choudhury et al. 2014). Thus, evaluating the sequence variation of related genes that underlie apiculus staining can be a good approach to the study of plant pigmentation.
The pigment characteristics of rice seeds are responsible for the accumulation of anthocyanins and proanthocyanins (PAs) synthesis, the secondary metabolites belonging to the kind of flavonoids that are derived from the common phenylpropanoid pathway (
Koes et al. 2005;
Grotewold 2006;
Lim and Ha 2013). In PAs synthesis of rice, the
Rc gene has been confirmed as a regulatory gene which encodes a bHLH DNA-binding protein. Mutations in the
Rc gene have been demonstrated in common rice strains with a white-colored pericarp (
Sweeney et al. 2006;
Furukawa et al. 2007). The activity of the
Rc from the red rice-type can be possibly disrupted by 14-bp deletion (
Lim and Ha 2013). Two single nucleotide substitutions at two sites of the 1
st and 2
nd exons of the
Rd gene had been observed in white rice (
Nakai et al. 1998). A single nucleotide substitution (C→A) of the 2
nd exon in the
Rd gene sequence generates a premature stop codon in white rice compared to the pigmented rice cultivars (
Lim and Ha 2013). In addition, 2-bp insertion was observed in the
OsB1 gene in the plant showing white and red pericarp. This mutation causes frameshift in the C-terminus, leading to early termination and regulating protein function (
Wang and Shu 2007;
Lim and Ha 2013).
According to the C–S–A gene system, rice hull pigmentation was controlled by the interaction of genes rather than by a single gene.
C1 and
A1 collectively determine the color variation, whereas
S1 diversifies the pigmentation at tissues. The
C1 operates as a switch in controlling color production when
C1 is expressed alone; it produces brown color in the apiculus, but in combination with
A1, produces purple color in apiculus. When
C1 and
S1 are expressed, brown is observed on the hull, and purple is observed on the hull if
C1,
A1 and
S1 are expressed. Several genes have been identified to color the various organs of plants, including apiculus, pericarp and hull (
Sun et al. 2018).
The present study was performed to detect and characterize genes that regulate pigmentation of sterile lemma and apiculus. Genetic analysis was performed using F2:3 populations from a cross between “Daeribbyeo 1” and “Josaengjado”, and the genetic map was constructed using Kompetitive Allele-Specific PCR (KASP) markers. Haplo-type analysis was performed using rice accession to confirm the genes to regulate color of sterile lemma and apiculus.
MATERIALS AND METHODS
Plant materials
Two temperate
japonica cultivars, ‘Josaengjado’ (a fe-male parent) with purple leaf, leaf sheath, sterile lemma, and apiculus and ‘Daeribbyeo 1’ (a male parent) with green leaf, leaf sheath, sterile lemma, and apiculus were crossed to develop F
2 and F
3 progenies for genetic analysis (
Fig. 1). The F
2 population (428 F
2 plants) was grown at the experiment station of Chungnam National University, Daejeon, Korea during the summer of 2018, and 120 F
3 lines were randomly selected and grown in 2019. The germinated seeds were sown on the 12
th of April and 30-day-old seedlings were transplanted with 15 × 30 cm intervals.
For haplotype analysis, the sequences of 78 Asian cul-tivated rice accessions (
Oryza sativa L.) from the KRICE_CORE at Kongju National University were compared (
Kim et al. 2016); the accessions included 31 temperate
japonica, 13 tropical
japonica, 28
indica, 3 aus, 1 admixture, and 2 aromatic (
Supplementary Table S1). The KRICE_CORE set plants were grown in the experimental field at Chung-cheongnam-do Agricultural Research and Extension Ser-vices (CNARES) in 2018.
In 2018 and 2019, the color of each plant in the F2 and F3 for the sterile lemma color (SC) and apiculus color (AC) was measured at 5-10 days after heading. The color was distinguished by representing purple and straw-white. In the F3 population, 10-15 plants per line were evaluated to check the segregation of the phenotype.
DNA extraction and PCR analysis
Genomic DNA was extracted from the fresh leaves of parents and F2 plants derived from Daeribbyeo 1 × Josaengjado cross. A total volume of 15 uL reaction mixture was composed of the 1 uL (10 ng/μL) of template DNA, 1.5 μL dNTP (2.5 μM each), 1.5 μL 10 × PCR buffer (10 μM Tris-HCl pH 8.3, 50 μM KCl, 1.5 μM MgCl2, 0.1% Gelatin), Forward + Reverse primer 1 μL (10 pmol each), Taq polymerase 0.25 μL (5 Unit/μL) and 8.75 μL triple distilled water. The PCR conditions were as follows: de-naturing at 94℃ for 2 minutes, followed by 34 cycles of 94℃ for 20 seconds, 58℃ for 15 seconds, and 72℃ for 30 seconds.
For substitution mapping to define the location of the genes, we used SSR markers located within the target region from the Gramene database (
www.gramene.org,
McCouch et al. 2002). A cleaved amplified polymorphic sequence (CAPS) marker was used to identifying the
Rd gene. CAPS marker for the
Rd gene was used to amplify DNA fragments, and these PCR products were digested with the
TaqI restriction enzyme (
Lim and Ha 2013).
Genetic map was developed using the Kompetitive Allele-Specific PCR (KASP) markers. KASP amplifications and allelic discriminations were performed using the Nexar system (LGC Douglas Scientific, Alexandria, USA) at the Seed Industry Promotion Center of the Foundation of Agricultural Technology Commercialization and Transfer at Gimje, Korea (
Cheon et al. 2018). Of the 506 markers, 127 polymorphic markers (25%) showed polymorphism and 89 markers were selected to construct the map. 120 F
2 plants were genotyped using 89 markers.
Linkage analysis was conducted using the Kosambi function of Mapmaker/EXP 3.0 software (
Lander et al. 1987). QTL analysis was performed by composite interval mapping (CIM) using the QTL Cartographer (Wang
et al. 2007). CIM analysis was conducted with a forward and backward method using model 6 with a 10 cM window size (
Lee et al. 2012). The log-likelihood (LOD) threshold significance level (
P < 0.05) about agronomic traits was calculated by computing 1,000 permutations and set to 3 as the LOD threshold significant level (
P < 0.05) about color traits. Gene sequencing reactions were performed by an external company (
www.solgent.com).
Rd gene was amplified into three sections using the primers Rd-1F (5ʹ-GTTAGGCAGTACAAGTGTGTGTAG-3ʹ), Rd-1R (5ʹ-TAGGAGCACGTGTAAAGGTAAGAT-3ʹ
Chin et al. 2016), Rd-2F (5ʹ-TAGGAACAACGATCCTC CACGTA-3ʹ), Rd-2R (5ʹ-GAGCTTCCACGACGAGAA GTG-3ʹ), Rd-3F (5ʹ-GCAAGTGATAATTGTGGTGGCA-3ʹ) and Rd-3R (5ʹ-TCAACACTCATTTGACCAACGC-3ʹ).
C1 gene was also amplified into three sections using the primers C1-1F (5ʹ-AAGTACAGCGCAAAAGTGGTAGA-3ʹ), C1-1R (5ʹ-AGCGTTAGCCAGCTTCAAAT-3ʹ), C1-2F (5ʹ-ATTTGGAGCTATTTGGTACTGTCG-3ʹ), C1-2R (5ʹ-TTCACGGTCGTGGAAGAAGAA-3ʹ), C1-3F (5ʹ-GGG CCGAACAGACAATGAAATC-3ʹ) and C1-3R (5ʹ-TTA TATACGGAAACCCGCAACTG-3ʹ).
RESULTS
Phenotypic variation of the parents and 120 F2 plants
The phenotypic differences between Daeribbyeo 1 and Josaengjado are shown (
Fig. 1). Two parents also showed significant differences including plant height. The color was observed on the leaf, leaf sheath, ligule, sterile lemma, and apiculus in Josaengjado, not Daeribbyeo 1. In 2018, 246 and 182 individuals exhibited purple sterile lemma and straw-white sterile lemma, respectively. Also, 220 and 208 individuals showed purple apiculus and straw-white api-culus, respectively (
Table 1). They fit a 9:7 segregation ratio indicating the action of two complementary genes. In 2019, the same segregation ratio was observed confirming that two genes regulate the color of the sterile lemma and the apiculus.
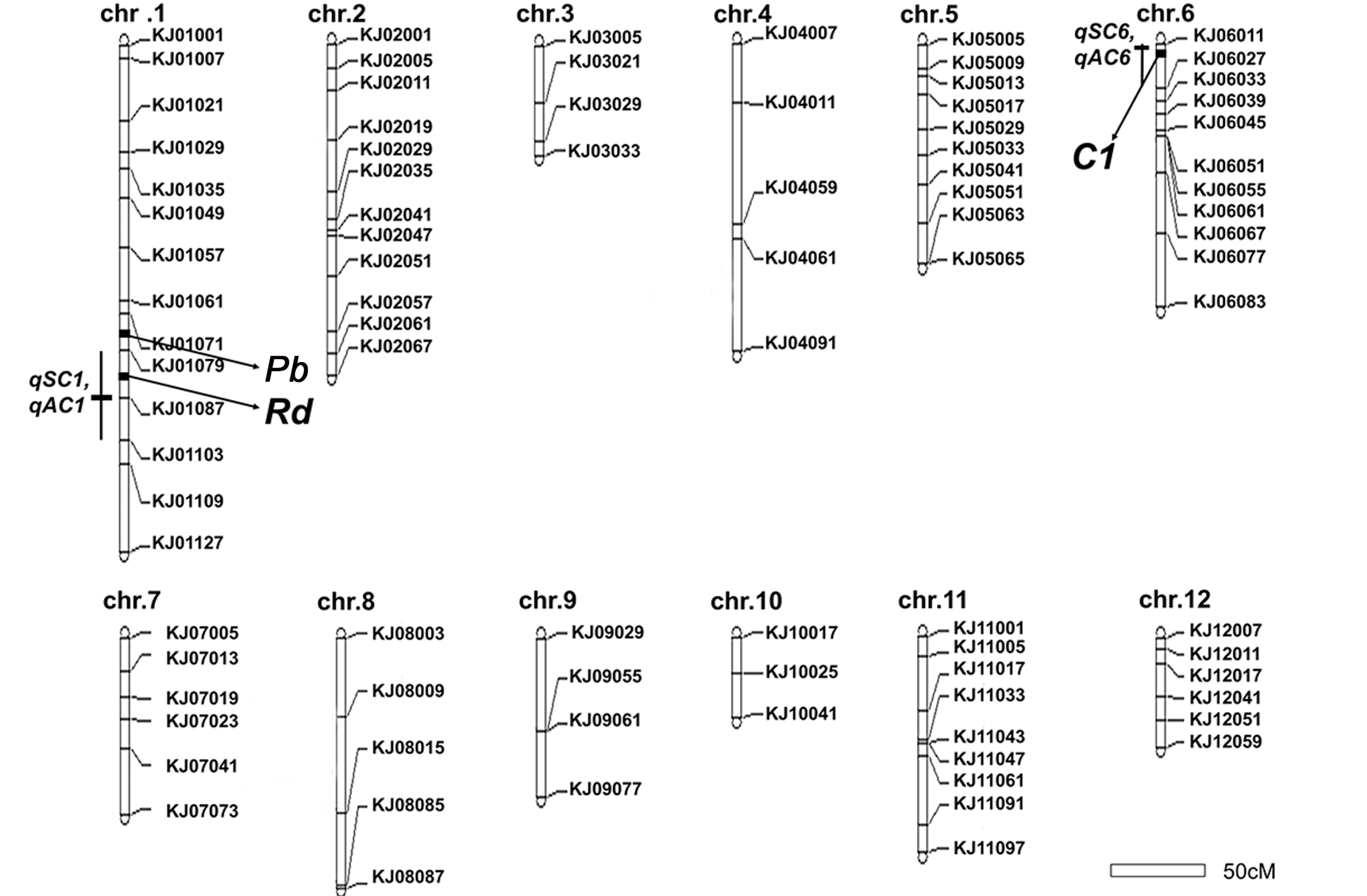
A genetic linkage map was constructed using 120 F
2 plants and 89 polymorphic KASP markers distributed over 12 chromosomes covering a total length of 1458.3 cM with an average distance of 16.3 cM (
Fig. 2). For sterile lemma color, two QTLs were detected on chromosomes 1 and 6.
qSC1 for sterile lemma color was located between KJ01079 and KJ01103 with
qSC6 between KJ06011 and KJ06027. The LOD score ranged from 6.77 to 9.76 (
Table 2). The
qSC1 accounted for 39.93% and
qSC6 accounted for 65.38% of the phenotypic variance. ‘Josaengjado’ alleles underlying all these QTLs showed positive additive effects contributing to the coloration of the sterile lemma. For apiculus color, two QTLs were detected on chromosomes 1 and 6.
qAC1 was located between KJ01079 and KJ01103, and
qAC6 was located between KJ06011 and KJ06027. The
qSC1 and
qAC1, and
qSC6 and
qAC6 were detected in the same region on chromosome 1 and 6, respectively. The LOD score ranged from 6.83 to 11.02 (
Table 2). Like
qSC, Josaengjado alleles underlying all these QTLs showed positive additive effects.
These results imply that qSC1 and qAC1 might be allelic to Rd gene controlling pericarp color with qSC6 and qAC6 allelic to C1 gene regulating apiculus color.
Substitution mapping of qSC and qAC
For the substitution mapping of
qSC1,
qAC1,
qSC6 and
qAC6, 16 F
3 lines with different chromosome segments within the region on chromosomes 1 and 6 were selected (
Fig. 3). Four F
3 lines (CN11, CN72, CN114, CN29) were homozygous for Josaengjado in the
qSC1 and
qAC1 region flanked by KASP marker KJ01079 and SSR marker RM11390, and homozygous for Daeribbyeo 1 in the
qSC6 and
qAC6 region defined by KASP marker KJ06011 and SSR marker RM19552. Both sterile lemma and apiculus color were not observed in both F
2 and F
3 lines. Furthermore, 4 F
3 lines (CN9, CN82, CN56, and CN2) were homozygous for Daerribyeo 1 in the
qSC1 and
qAC1 region defined by KASP marker KJ01079 and SSR marker RM11390, and homozygous for Josaengjado across the
qSC6 and
qAC6 region defined by KASP marker KJ06011 and SSR marker RM19552. Color phenotypes in these lines were not observed in both sterile lemma and apiculus in both F
2 and F
3 lines. On the other hand, two F
3 lines (CN74, CN59) were homozygous for Josaengjado in the region between KASP marker KJ01079 and SSR marker RM11390, homozygous for Daeribbyeo 1 in the region defined between KASP marker KJ06011 and SSR marker RM19552. These two lines showed both purple sterile lemma and apiculus observed in both F
2 and F
3. Another two F
3 lines (CN38 and CN30) were heterozygous in the
qSC1 and
qAC1 region, and homozygous for Josaengjado in the
qSC6 and
qAC6 region. These lines showed segregation for color and colorless sterile lemma and apiculus at F
3 (
Supplementary Fig. S1).
These results confirmed that two complementary genes regulate sterile lemma color and apiculus color.
Rd gene was located in
qSC1 and
qAC1 region with
OsC1 in
qSC6 and
qAC6 (
Saitoh et al. 2004;
Furukawa et al. 2007).
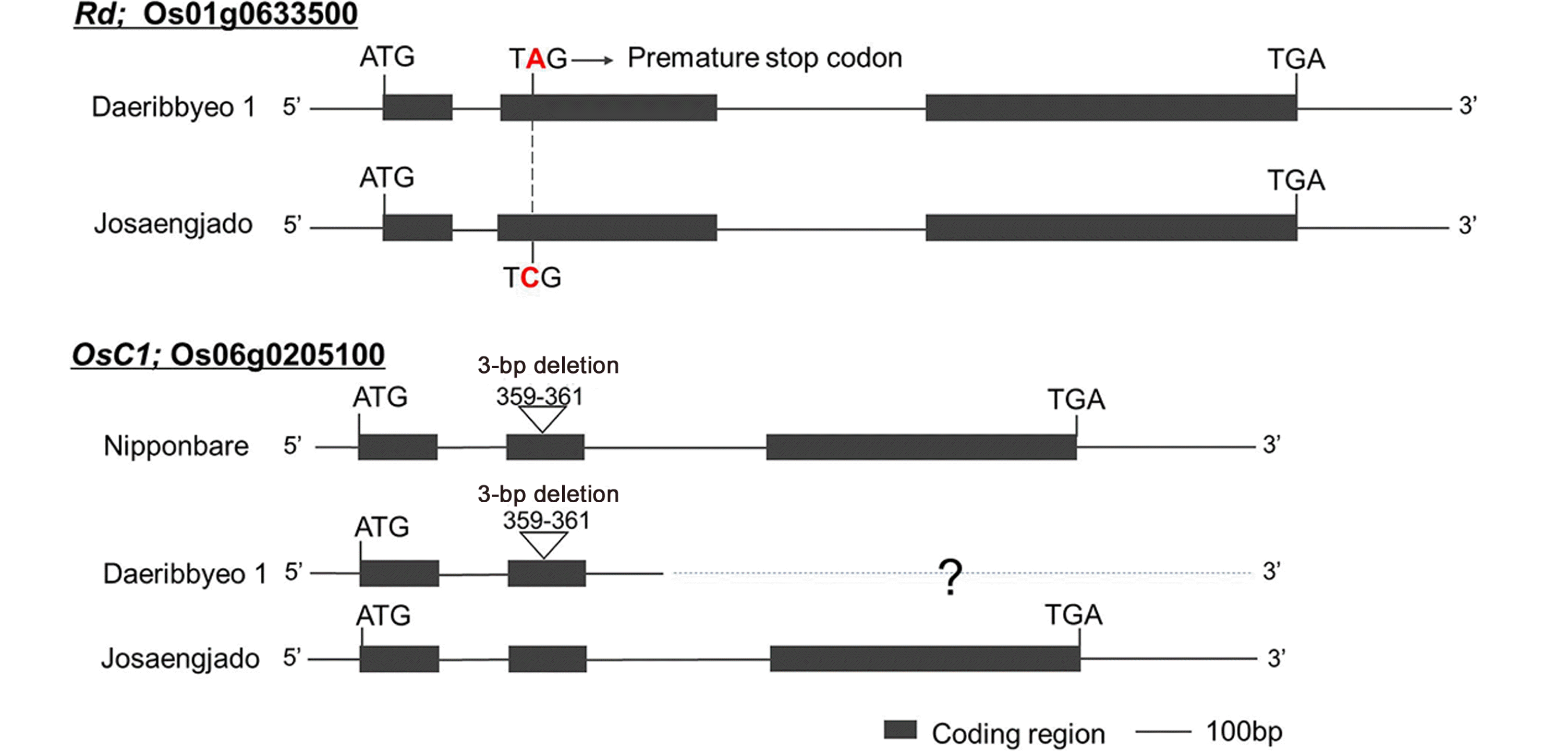
Comparative analysis of the
Rd and
OsC1 sequences of Daeribbyeo 1 and Josaengjado was performed. As a result, a 1-bp substitution (C→A) was found in the second exon of
Rd of Daeribbyeo 1, which was confirmed by the same mutation (
Fig. 4). As with previous studies, a 1-bp substitution appears to prematurely terminate the translation of amino acids and consequently the function of the
Rd gene (
Lim and Ha 2013). In
OsC1 gene of Daeribbyeo 1, a 3-bp deletion (GAG) was detected in Daeribbyeo 1 sequence; position 359-361 compared to Josaengjado. The gene en-coding an
R2R3-MYB transcription factor had been re-ported as
OsC1 and predicted to be a regulator in the anthocyanin biosynthesis pathway (
Saitoh et al. 2004). A 3-bp deletion in the second exon in Daeribbyeo 1 caused the deletion of loss of glutamic acid.
Haplotype analysis was performed to compare the sequence diversity of rice accessions for
Rd and
OsC1. SNPs and InDels data of
Rd and
OsC1 of 78 rice accessions were provided by Kongju University (
Kim et al. 2016). Based on the genomic sequence of
Rd and
OsC1, 78 accessions were divided into 14 types (
Table 3). 78 accessions showed two types at
Rd with nucleotides A and C. In
OsC1, 11 SNPs and deletions were detected that resulted in the change of amino acid. These included at least 11 mutations (H4 to H14) including three deletion regions and 12 SNP regions.
H1 type included Josaengjado and most of the lines had purple sterile lemma and apiculus. The color appeared on the sterile lemma and apiculus of the 12
japonica and 5
indica accessions with an exception of 8
japonica accessions. H2 and H3 types showed SNPs in
OsC1, and these SNPs did not result in the change of coloration in sterile lemma color and apiculus color. The substitution in H3 was considered not to affect the function. This SNP was also reported in Purpleputtu, which has a colored apiculus (
Reddy 1996). H3 was functional without the change in the phenotype in
OsC1 (
Zheng et al. 2019). H4 type had 2-bp deletion in
OsC1 and failed to show coloration. These deletions occurred in the positions of putative base-contacting residues and caused a frameshift (
Martin and Paz-Ares 1997). H5, H6, H8, H9, and H11 types had SNPs in
OsC1 resulting in the loss of color. Also, H7 type showed straw-shite sterile lemma and apiculus exhibiting the 10-bp deletion in the third exon. Out of 25 accessions, 20 were
indica with purple sterile lemma whereas two accessions had straw-colored sterile lemma. H10 had the 3-bp deletion in exon 2, with the straw-white sterile lemma and apiculus. H12 had mutantion in
Rd with the wild type allele in
OsC1, and no color appeared in sterile lemma and apiculus. In H13 and H14 types, an SNP was observed in
Rd with SNP and InDel in
OsC1. These results indicate that two genes are complementary for coloration in sterile lemma and apiculus. Although H1 type accessions had purple sterile lemma and apiculus, eight
japonica accessions in H1 did not have colored sterile lemma and apiculus. This might be due to the existence of additional mutations in the
Rd gene in the eight
japonica accessions, and additional study is needed to clarify these results (
Nakai et al. 1998).
DISCUSSION
Pigmentation is one of the various phenotypes that are common in wild rice but varied in cultivated rice (
Saitoh et al. 2004). Several genes have been reported to regulate anthocyanin pigmentation in leaves, leafsheath, pericarp and hull. However, the study on the color of sterile lemma is rare. In the previous report, the segregation ratio of 2.9:1 (Purple leafsheath:Green leafsheath) indicated that
OsC1 was responsible for purple leaf sheath (
Chin et al. 2016). In this study, we confirmed that the purple and straw-white apiculus and purple and straw-white sterile lemma segre-gated with a 9:7 ratio, respectively, indicating the involvement of two complementary genes. QTLs analysis and substitution mapping confirmed that one gene detected on chromosome 1 was located between KJ01079 and KJ01103 and the other detected on chromosome 6 was located between KJ06011 and KJ06027. The
Rd gene was colocalized with
qSC1 and and the
OsC1 gene was located in
qSC6 and
qAC6 sug-gesting that
qSC1 is allelic to
Rd and
qSC6 is allelic to
OsC1 (
Fig. 2).
In the previous study, two single nucleotide substitutions at two positions of the 1
st and 2
nd exons of the
Rd gene have been previously observed in white rice (
Nakai et al. 1998). In this study, only one single nucleotide mutation (C→A) in the 2
nd exon of
Rd was detected (
Fig. 4). Mutations cause premature stop resulting in white rice seed phenotype due to lack of anthocyanin and PA biosynthesis (
Lim and Ha 2013). A 3-bp deletion in the second exon at
OsC1 caused loss of gene function resulting in glutamic acid (E) deletion (
Nakai et al. 1998;
Sun et al. 2018).
Haplotype analysis was performed using KRICE_CORE accessions to confirm whether the two complementary genes regulate the sterile lemma and apiculus color in other rice accessions. 78 accessions could be divided into 14 types. Rd has an SNP that leads to a premature stop. In C1, a total of 11 mutation events were identified, including 5 non-synonymous, 2 synonymous, 1 premature stop, and 3 deletions.
Almost all
indica had the 10-bp (ACTGGAACAG) deletion and 2-bp (TC) and 3-bp (GAG) deletions were mostly detected in
japonica accessions. These seem to indicate that mutations in
OsC1 occur independently in
indica and
japonica. Accessions with wild type alleles at both
Rd and
C1 have purple sterile lemma and apiculus. It can endorse that pigmentation in the apiculus is observed when
Rd and
OsC1 are expressed (
Sun et al. 2018). Additionally, the results support the conclusion that deletions or substitutions are partially associated with phenotypic changes observed in rice accessions with different
C1 alleles (
Saitoh et al. 2004;
Zheng et al. 2019).
SUPPLEMENTARY MATERIALS
ACKNOWLEDGEMENTS
This study was financially supported by research fund of Chungnam National University in 2018.
Fig. 1Morphological characteristics of Daeribbyeo 1 (D), Josaengjado (J), and F2 plants. Josaengjado shows abnormal gross morphologies, such as dwarfism, shortened internodes, small-rounded seeds, and erect leaves compared with Daeribbyeo 1.

Fig. 2Construction of a genetic map using 89 KASP markers with 120 F2 plants from a cross between Daeribbyeo 1 and Josaengjado. Right sides of each chromosome indicate the name of each marker, the left sides of each chromosome indicate the region of QTLs, and black box means the nearest regions of QTLs, respectively. The genetic distance, measured in centimorgans (cMs). Linkage analysis was performed using Mapmaker/EXP 3.0 software. QTL analysis was performed by simple internal mapping using QTL Cartographer software.
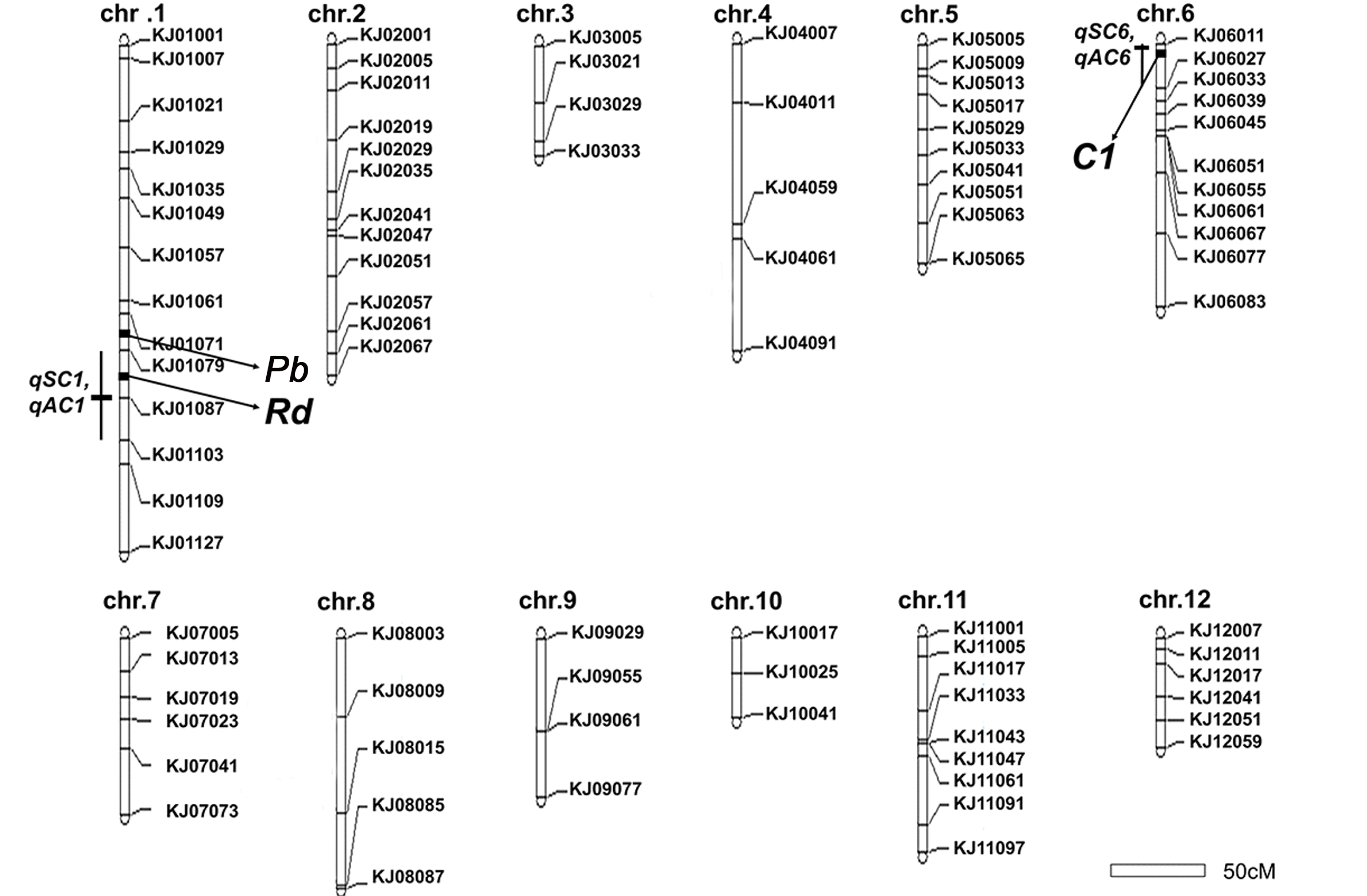
Fig. 3Substitution mapping of the qSC1, qAC1, qSC6, and qAC6 on the chromosome 1 and 6 based on 16 F3 lines and two parents, Daeribbyeo 1 and Josaengjado. White, black, and gray bars indicate homozygous regions for Daeribbyeo 1, Josaengjado alleles and heterozygous, and slashed areas indicate crossing-over regions. The right table indicates phenotypes of each line. Parenthesis means a number of individuals.

Fig. 4Comparison of nucleotide sequences of Daeribbyeo 1 and Josaengjado at Rd and OsC1 (Nipponbare sequence at OsC1 is included). The black boxes and lines between black boxes indicate exons and introns, respectively. Letters above and below the boxes are sequences of Hwaseong and O. rufipogon, respectively. Non-synonymous nucleotide substitution is indicated in red letter in Daeribbyeo 1 at the Rd gene. A 3-bp deletion in OsC1 was observed in Daeribbyeo 1 and Nipponbare compared to Josaengjado. The sequence of the slashed box in the OsC1 gene of Daeribbyeo 1 is not available because we failed to clone. including the 3rd exon (shown in slashed box).
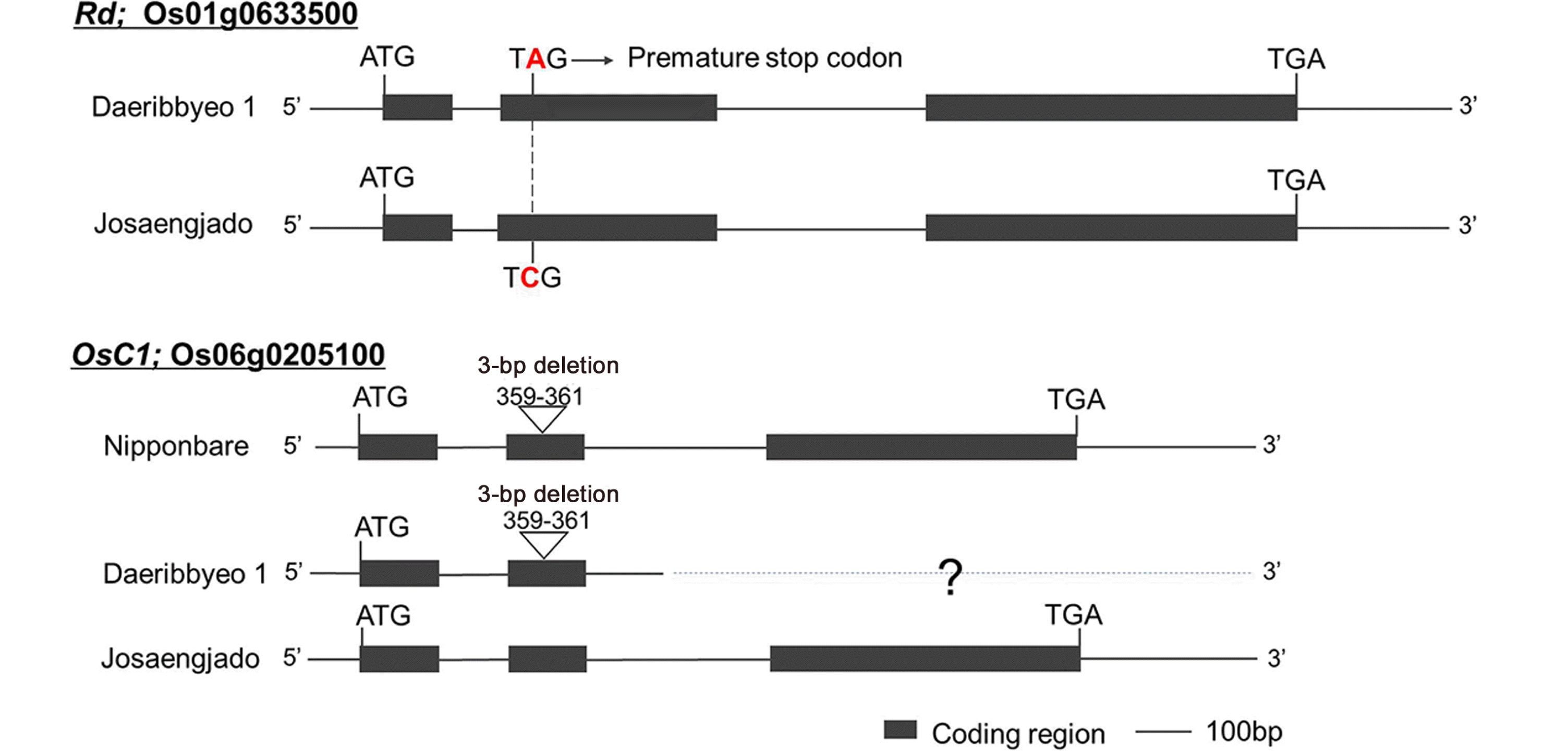
Table 1Segregations of two morphological traits in the F2 population.
Table 1
|
Trait |
|
Purple |
Straw-white |
Expected ratio |
Chi-square |
P
|
|
Sterile lemma color |
2018 |
246 |
182 |
9:7 |
0.01 |
0.92 |
|
2019 |
139 |
101 |
|
0.02 |
0.90 |
|
Apiculus color |
2018 |
220 |
208 |
9:7 |
0.15 |
0.70 |
|
2019 |
139 |
101 |
|
0.02 |
0.90 |
Table 2Characteristics of QTLs for morphological traits in the F2 population.
Table 2
|
Trait |
QTL |
Chr. |
Marker intervalz)
|
LOD |
R2 (%)y)
|
Additive effectx)
|
Candidate genew)
|
|
Sterile lemma color |
qSC1
|
1 |
KJ01079/KJ01087/KJ01103 |
6.77 |
39.93 |
0.95 |
Rd
|
|
qSC6
|
6 |
KJ06011/KJ06027 |
9.76 |
65.38 |
1.27 |
C1
|
|
Apiculus color |
qAC1
|
1 |
KJ01079/KJ01087/KJ01103 |
6.83 |
41.05 |
0.94 |
Rd
|
|
qAC6
|
6 |
KJ06011/KJ06027 |
11.02 |
67.45 |
1.30 |
C1
|
Table 3Haplotype analysis of Rd and C1 using the Korean rice core set (KRICE_CORE).
Table 3
|
Haplotypez) (phenotype) |
Rd
|
|
C1
|
Total |
Sterile lemma and apiculus color |
|
276y)
|
4 |
51 |
122 |
277 |
360 |
786 |
795 |
844 |
918 |
1010 |
1012 |
Jap |
Ind |
Aus |
Aro |
Adm |
|
H1 (3) |
C |
|
G |
G |
C |
G |
GAG |
TC |
ACTGG
AACAG |
C |
C |
G |
G |
25 |
12 (8)v)
|
5 |
|
|
|
|
H2 (3) |
- |
|
- |
- |
- |
- |
- |
- |
- |
T |
- |
- |
- |
1 |
|
1 |
|
|
|
|
H3 (3) |
- |
|
- |
- |
- |
- |
- |
- |
- |
- |
A
(P→Q) |
- |
- |
1 |
1 |
|
|
|
|
|
H4 (1) |
- |
|
- |
- |
- |
- |
- |
‒2 bp |
- |
- |
- |
- |
- |
2 |
2 |
|
|
|
|
|
H5 (1) |
- |
|
T (Gx)→W) |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
1 |
|
|
|
|
|
H6 (1) |
- |
|
- |
A
(Stop) |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
1 |
|
|
|
|
|
H7 (1) |
- |
|
- |
- |
- |
- |
- |
- |
‒10 bp |
- |
- |
- |
- |
25 |
2 |
20 (2) |
|
|
1 |
|
H8 (1) |
- |
|
- |
- |
G
(P→R) |
- |
- |
- |
- |
- |
- |
- |
- |
3 |
|
|
2 (1) |
|
|
|
H9 (1) |
- |
|
- |
- |
- |
A
(R→Q) |
- |
- |
- |
- |
- |
- |
- |
3 |
3 |
|
|
|
|
|
H10 (1) |
- |
|
- |
- |
- |
-
|
‒3 bp |
- |
- |
- |
- |
- |
- |
3 |
3 |
|
|
|
|
|
H11 (1) |
- |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
C
(A→P) |
T |
3 |
3 |
|
|
|
|
|
H12 (1) |
A
(Stop) |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
4 |
2 |
|
|
2 |
|
|
H13 (1) |
A
(Stop) |
|
- |
- |
- |
A
(R→Q) |
- |
- |
- |
- |
- |
- |
- |
1 |
1 |
|
|
|
|
|
H14 (1) |
A
(Stop) |
|
- |
- |
- |
- |
‒3 bp |
- |
- |
- |
- |
- |
- |
5 |
5 |
|
|
|
|
|
Amino acidw)
|
Ps |
|
Ns |
Ps |
Ns |
Ns |
Del |
Del |
Del |
S |
Ns |
Ns |
S |
|
|
|
|
|
|
References
- Cheon KS, Baek J, Cho YI, Jeong YM, Lee YY, Oh J, et al. 2018. Single nucleotide polymorphism (SNP) discovery and Kompetitive allele-specific PCR (KASP) marker development with Korean Japonica rice varieties. Plant Breed. Biotech.. 6: 391-403.
- Chin HS, Wu YP, Hour AL, Hong CY, Lin YR. 2016. Genetic and evolutionary analysis of purple leaf sheath in rice. Rice (N. Y.). 9: 8
- Choudhury BI, Khan ML, Dayanandan S. 2014. Patterns of nucleotide diversity and phenotypes of two domestication related genes (OsC1 and Wx) in indigenous rice varieties in Northeast India. BMC Genet.. 15: 71
- Fan FJ, Fan YY, Du JH, Zhuang JY. 2008. Fine mapping of C (Chromogen for anthocyanin) gene in rice. Rice Sci.. 15: 1-6.
- Furukawa T, Maekawa M, Oki T, Suda I, Iida S, Shimada H, et al. 2007. The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp. Plant J.. 49: 91-102.
- Gao X, Liang W, Yin C, Ji S, Wang H, Su X, et al. 2010. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol.. 153: 728-740.
- Grotewold E. 2006. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol.. 57: 761-780.
- Hong L, Qian Q, Zhu K, Tang D, Huang Z, Gao L, et al. 2010. ELE restrains empty glumes from developing into lemmas. J. Genet. Genomics. 37: 101-115.
- Ikeda K, Ito M, Nagasawa N, Kyozuka J, Nagato Y. 2007. Rice ABERRANT PANICLE ORGANIZATION 1, encoding an F-box protein, regulates meristem fate. Plant J.. 51: 1030-1040.
- Kim CK, Cho MA, Choi YH, Kim JA, Kim YH, Kim YK, et al. 2011. Identification and characterization of seed-specific transcription factors regulating anthocyanin biosynthesis in black rice. J. Appl. Genet.. 52: 161-169.
- Kim TS, He Q, Kim KW, Yoon MY, Ra WH, Li FP, et al. 2016. Genome-wide resequencing of KRICE_CORE reveals their potential for future breeding, as well as functional and evolutionary studies in the post-genomic era. BMC Genomics. 17: 408
- Kobayashi K, Maekawa M, Miyao A, Hirochika H, Kyozuka J. 2010. PANICLE PHYTOMER2 (PAP2), encoding a SEPALLATA subfamily MADS-box protein, positively controls spikelet meristem identity in rice. Plant Cell Physiol.. 51: 47-57.
- Koes R, Verweij W, Quattrocchio F. 2005. Flavonoids: a colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci.. 10: 236-242.
- Lander ES, Green P, Abrahamson J, Barlow A, Daly MJ, Lincoln SE, et al. 1987. MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics. 1: 174-181.
- Lee HS, Sasaki K, Higashitani A, Ahn SN, Sato T. 2012. Mapping and characterization of quantitative trait loci for mesocotyl elongation in rice (Oryza sativa L.). Rice (N. Y.). 5: 13
- Li D, Chen C. 1993. The characteristics of two ecotypes of O. rufipogon in China and ecological investigation. J. Southern Agric.. 1: 6-11.
- Lim SH, Ha SH. 2013. Marker development for the identification of rice seed color. Plant Biotechnol. Rep.. 7: 391-398.
- Lin-Wang K, Bolitho K, Grafton K, Kortstee A, Karunairetnam S, McGhie TK, et al. 2010. An R2R3 MYB transcription factor associated with regulation of the anthocyanin biosynthetic pathway in Rosaceae. BMC Plant Biol.. 10: 50
- Liu X, Sun X, Wang W, Ding H, Liu W, Li G, et al. 2012. Fine mapping of Pa-6 gene for purple apiculus in rice. J. Plant Biol.. 55: 218-225.
- Martin C, Paz-Ares J. 1997. MYB transcription factors in plants. Trends Genet.. 13: 67-73.
- McCouch SR, Teytelman L, Xu Y, Lobos KB, Clare K, Walton M, et al. 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res.. 9: 199-207.
- Nakai K, Inagaki Y, Nagata H, Miyazaki C, Iida S. 1998. Mole-cular characterization of the gene for dihydroflavonol 4-reductase of japonica rice varieties. Plant Biotechnol.. 15: 221-225.
- Oikawa T, Maeda H, Oguchi T, Yamaguchi T, Tanabe N, Ebana K, et al. 2015. The birth of a black rice gene and its local spread by introgression. Plant Cell. 27: 2401-2414.
- Reddy AR. Khush GS., 1996. Genetic and molecular analysis of the anthocyanin pigmentation pathway in rice. editor. Rice Genetics III. International rice Research Institute. Manila, Philippines: pp. 341-352.
- Ren D, Li Y, Zhao F, Sang X, Shi J, Wang N, et al. 2013. MULTI-FLORET SPIKELET1, which encodes an AP2/ERF protein, determines spikelet meristem fate and sterile lemma identity in rice. Plant Physiol.. 162: 872-884.
- Saitoh K, Onishi K, Mikami I, Thidar K, Sano Y. 2004. Allelic diversification at the C (OsC1) locus of wild and cultivated rice: nucleotide changes associated with pheno-types. Genetics. 168: 997-1007.
- Shao Y, Xu F, Tang F, Bao J. 2013. The temporal and spatial expression pattern of anthocyanin related genes in rice (Oryza sativa L.). J. Nuclear Agr. Sci.. 27: 9.
- Siddiq EA, Sadananda AR, Zaman FU. 1986. Use of primary trisomic of rice in genetic analysis. Rice Genetics, Proceedings of the International Rice Genetics Symposium. International Rice Research Institute. Manila, Philippines: pp. 185-197.
- Sun X, Zhang Z, Chen C, Wu W, Ren N, Jiang C, et al. 2018. The C-S-A gene system regulates hull pigmentation and reveals evolution of anthocyanin biosynthesis pathway in rice. J. Exp. Bot.. 69: 1485-1498.
- Sweeney MT, Thomson MJ, Pfeil BE, McCouch S. 2006. Caught red-handed: Rc encodes a basic helix-loop-helix protein conditioning red pericarp in rice. Plant Cell. 18: 283-294.
- Takahashi ME. 1957. Analysis on apiculus color genes essential to anthocyanin coloration in rice. J. Fac. Agr. Hokkaido Univ.. 50: 266-362.
- Wang C, Shu Q. 2007. Fine mapping and candidate gene analysis of purple pericarp gene Pb in rice (Oryza sativa L.). Chinese Sci. Bull.. 52: 3097-3104.
- Yoshida A, Suzaki T, Tanaka W, Hirano HY. 2009. The homeotic gene long sterile lemma (G1) specifies sterile lemma identity in the rice spikelet. Proc. Natl. Acad. Sci. U. S. A.. 106: 20103-20108.
- Zhao S, Wang C, Ma J, Wang S, Tian P, Wang J, et al. 2016. Map based cloning and functional analysis of the chromogen gene C in rice (Oryza sativa L.). J. Plant Biol.. 59: 496-505.
- Zheng J, Wu H, Zhu H, Huang C, Liu C, Chang Y, et al. 2019. Determining factors, regulation system, and domestication of anthocyanin biosynthesis in rice leaves. New Phytol.. 223: 705-721.