Abstract
To understand genetic diversity in nutritional properties, 157 accessions of Korean-bred rice varieties were cultivated in 3 separate fields and harvested brown rice were used for determination of tocopherols (T), tocotrienols (T3), squalene (SQ), campesterol (CA), sitosterol (SI), and stigmasterol (ST) contents as well as fatty acid compositions. The average contents of α-T, γ-T, α-T3, γ-T3, SQ, CA, SI, and ST were 11.9, 1.6, 10.0, 13.9, 35.2, 42.1, 163.5, and 20.0 μg/g, respectively, and total tocols, SQ, and total phytosterols contents ranged 26.8-54.9, 7.9-78.4, and 162.9-320.2 μg/g, respectively. Ecotype of rice significantly affected phytonutrient contents in that japonica-type showed significantly higher α-T and α-T3 contents and α-T/γ-T, α-T3/γ-T3, and T/T3 ratios compared to indica-type varieties. Total T, SQ and ST contents were also higher in japonica-types, while total T3, CA and SI contents were not affected by ecotypes. Linoleic, oleic, and stearic acids were the 3 major fatty acids consisting 36.5, 35.8, and 22.9% of total fatty acids, respectively. Positive correlationships were observed among 3 phytosterols, while oleic acid showed negative correlation with palmitic (r = ‒0.662**) and linoleic (r = ‒0.810**) acids. Partial least squares discriminant analysis (PLS-DA) revealed that phytonutrient profiles may effectively differentiate rice ecotypes, and α-T3/γ-T3, γ-T and α-T3 contents were the key components with highest variable im-portance in projection (VIP) scores. All these results showed diverse genetic and ecotype-dependent variations in phytonutrients in Korean-bred rice varieties, which can be further used for developing a superior rice variety with higher nutritional value.
-
Key words: Phytochemical, Germplasm, Breeding, Ecotype, Nutrient
INTRODUCTION
Rice used to be a major staple food crop of the world, especially in Asian countries including Korea. The history of rice introduction to Korean peninsula is estimated as the early Bronze age (
Ahn 2010) resulting in diverse landraces in Korea (
Lee et al. 2018). Modernized rice researches in Korea started in 1906 as the establishment of Kwonup Mobeomjang and since that time rice varieties of diverse properties have been bred, released and cultivated in Korea (
Cho et al. 2020). High-yield, stable production, compatibility to developed cultivation practices, and disease-, insect-, and environmental stress-resistancy as well as specific food processing purpose used to be important phenotypes for rice breeders, although major target phe-notype for breeding programs have been changed with close association to social demands at each era. From late 1960’s high yield rice variety which can resist high fertilizer application and subsequent lodging was the major goal of breeding, and historic rice variety ‘Tongil’ with 4.5-5.7 MT/ha yield capacity developed in 1971 enabled rice self-sufficiency in South Korea (
Cho et al. 2020). Ironically, however, since rice self-sufficiency in late 1970’s, per capita rice consumption is rapidly decreasing and consequently recent breeding focuses are shifting from yield-related quantitative traits to qualitative traits of rice such as palatability, grain color, special flavor, and health-beneficial effects.
Rice grains, especially unpolished brown rice carrying nutrient-rich rice bran are well-known to contain various phytonutrients: ferulic acid, phenolics, anthocyanins, phytic acid, γ-oryzanol, octacosanol, dietary fiber, tocopherols, tocotrienols, and phytosterols, etc. (
Zhou et al. 2002;
Sohail et al. 2017;
Gong et al. 2020). Gamma (γ)-oryzanol might be a phytonutrient most intensively studied in rice due to its exclusive presence in rice and distinct cholesterol-lowering, antioxidative, and anti-cancer effects (
Qureshi et al. 2002;
Tan and Norhaizan 2017). Variations in rice oryzanol contents as affected by both environmental and genetic factors also have been reported (
Bergman and Xu 2003;
Huang et al. 2015; Cho
et al. 2019). Vitamin E, consisting of 8 vitamers: α-, β-, γ-, δ-tocopherols (T) and α-, β-, γ-, δ-tocotrienols (T3), collectively called as tocols is a lipid-soluble vitamin rich in rice bran and well known for its antioxidative, anticancer, skin-protective and many other therapeutic potentials (
Wong and Radhakrishnan 2012). Squalene, a triterpene compound used as a precursor for cholesterol synthesis in the human body have been widely commercialized as extracted from shark liver oil. Diverse plant species including rice, however, also contain squalene and due to its chemical structure’s similarity to lipids in human skin tissue, squalene shows strong preventive effects on skin, colon, and lung tumorigenesis in addition to its effects on reducing free radical oxidative damage to skin (
Huang et al. 2009). In analogy to the presence of cholesterols in animal and ergosterols in fungi tissues, phytosterols such as campesterol, sitosterol, and stigmasterol are generally found in plant tissues. Studies on phytosterols have revealed their diverse health-beneficial effects; inhibition of cholesterol absorption, lowering low-density lipoprotein (LDL) in blood as well as carrying anticarcinogenic, antioxidative and anti-inflammatory effects (
Jung et al. 2014). Fatty acids are primary metabolites of rice essential for completion of normal life cycle as constituents for lipids. Although rice is not a kind of lipid-enriched seed, brown rice contains about 2.5 to 3.0% of lipids and relatively high composition of unsaturated fatty acids are reported (
Kitta et al. 2005). Fatty acid composition also shows strong relationship with palatability and subsequent market quality and competitiveness of rice via affecting pasting properties after cooking (
Yoon et al. 2008;
Yoon et al. 2012).
Considering the long cultivation history of rice in Korea as a major food crop, the presence of diverse Korean landraces is a proof of continuous efforts and achievements in traditional rice breeding. Since ‘Namseon-1-ho’, the first modernized rice variety developed in 1932, more than 300 rice varieties for table and special purposes have ever been developed in Korea (
Cho et al. 2020). During the process of registering a new variety, its novelty, distinctness, uniformity, and stability properties are clearly provided in addition to major agronomic traits. The phytonutrient properties of a newly developed variety, however, are rarely investigated and provided. In this study, we characterized major lipophilic phytonutrients: tocopherols, tocotrienols, squalene, phytosterols, together with fatty acid compositions in brown rice of 157 varieties bred and cultivated in Korea.
MATERIALS AND METHODS
Plant materials
A rice core collection consisting of 294 accessions were established by a Center of Crop Breeding on Omics and Artificial Intelligence, Kongju National University (
Choi et al. 2018), and out of which 157 accessions of Korean-bred varieties corresponding to accession numbers of RWG138 to RWG294 were selected for this experiment. Selected 157 accessions consisted of 136 japonica-type and 21 indica-type varieties. List of accessions including their variety name, ecotype, origin, and year of development are provided in
Supplementary Table S1. The rice seeds were endowed from Kongju National University in 2018 and cul-tivated in 3 separate locations in South Korea: Suwon (37°17'28"N/127°0'32"E), Yesan (36°44'28.0"N/126°49'02.8"E), and Busan (35°13'59.7"N/129°04'46.0"E) in order to neutralize environmental factor-induced variation and clarify genetic factor-dependent phytonutrient property. Rice samples harvested in each location were stored at ‒70℃ in unhusked form, and prior to analysis samples were thawed overnight to room temperature, dehusked into brown rice using a rice huller (BH-06, SY21 THC, Incheon, Korea), pulverized with a milling machine (TM-05, Satake, Japan), and then passed through a 1.4-mm sieve.
Lipophilic phytonutrients such as tocopherols, tocotrienols, squalene, and phytosterols were quantified according to
Mahmud et al. (2018) after slight modifications. Powdered brown rice (2.0 g) in a 50 mL centrifuge tube were mixed with 0.1 g of ascorbic acid and 10 mL ethanol and kept in a water bath at 80℃ for 10 minutes for extraction, after which 300 μL of saturated KOH were added and kept in a water bath (80℃) again for additional 18 minutes for saponification. After saponification, test tubes were cooled in an ice bucket and unsaponifiable fractions were collected by liquid-liquid extraction in that 10 mL of hexane and 10 mL of distilled water were added, vigorously vortexted, cen-trifuged at 1,500 rpm for 10 minutes at 4℃, and then upper hexane layers were collected. This liquid-liquid extraction process was performed two more times and pooled hexane layers were washed twice with 10 mL of distilled water, and passed through Na
2SO
4 for removal of water. After vacuum rotary evaporation at 45℃, samples were re-suspended in 2 mL isooctane containing 100 ppm racTocol (CAS 119-98-2) as an internal standard and injected into a gas chromatography (GC, Model GC-2010 Plus, Shimadzu, Japan) coupled with a flame ionization detector (FID). For simultaneous separation of α- and γ-tocopherol (T), α- and γ-tocotrienol (T3), squalene (SQ), campesterol (CA), sitosterol (SI), and stigmasterol (ST), a DB-5 MS capillary column (30 m × 0.25 mm, Agilent, USA) was used. GC conditions for analysis were as follows: oven temperature started at 250℃ and hold for 2 minutes, increased up to 281℃ by 4℃/minute and hold for 30 minutes, and then increased to 315℃ by 10℃/minute. The injector and FID detector temperatures were 290℃, and the nitrogen carrier (99.99% purity) maintained a column flow rate of 0.41 mL/minute with a split ratio of 1:20. Authentic standard for α-T, γ-T, α-T3 and γ-T3, and racTocol were purchased from abcam (U.K.), and standards for squalene, campesterol, and stigmasterol were purchased from Sigma (USA), while sitosterol standard was purchased from EPRS (European Pharma References Standard).
A one-step extraction/methylation protocol (
Cho et al. 2006) after slight modifications were applied for analysis of fatty acid composition. About 0.2 g of brown rice samples were weighed into a 1.5-mL vial, into which 680 μL of reaction solution consisting of MeOH:benzene:2,2-dimethoxypropane:H
2SO
4 = 39:20:5:2 and 400 μL of heptane were added. After vigorous vortexing, the vials were heated for 2 hours at 80oC for extraction and methylation. After cooling down to room temperature reaction solutions were transferred into an e-tube and after centrifugation at 12,000 rpm for 5 minutes, heptane layers containing fatty acid methyl esters (FAME) were transferred into a glass vial and injected into a gas chromatography (GC-2010 Plus, Shimadzu, Japan). FAME were separated through a SP-2560 FAME column (100 m × 0.25 mm, Supelco, USA) using the following temperature program: start at 150℃ and hold for 2 minutes, increase to 215℃ at 4℃/minute and hold for 35 minutes. The injector and FID detector temperatures were 240℃, and the nitrogen carrier (99.99% purity) maintained a column flow rate of 0.95 mL/minute with a split ratio of 1:20. For the identification of FAME, the retention time of each peak was compared against those of 37 authentic FAME standards (Supelco, USA), and composition of each fatty acid was expressed as a percentage value of corresponding fatty acid’s peak area relative to the total area of all identified fatty acid peaks.
Rice samples cultivated in 3 different locations were used as 3 biological replications for all phytonutrient analyses. Statistics software SPSS (version 25, IBM Corp., USA) were used to calculate basic statistics and correlation coefficients at
P < 0.05. For multivariate analyses such as partial least square discriminant analyses (PLS-DA), MetaboAnalysis (
http://www.metaboanalyst.ca/) was used after mean-centered scaling followed by dividing by the standard deviation of each phytonutrient data.
RESULTS
Vitamin E contents
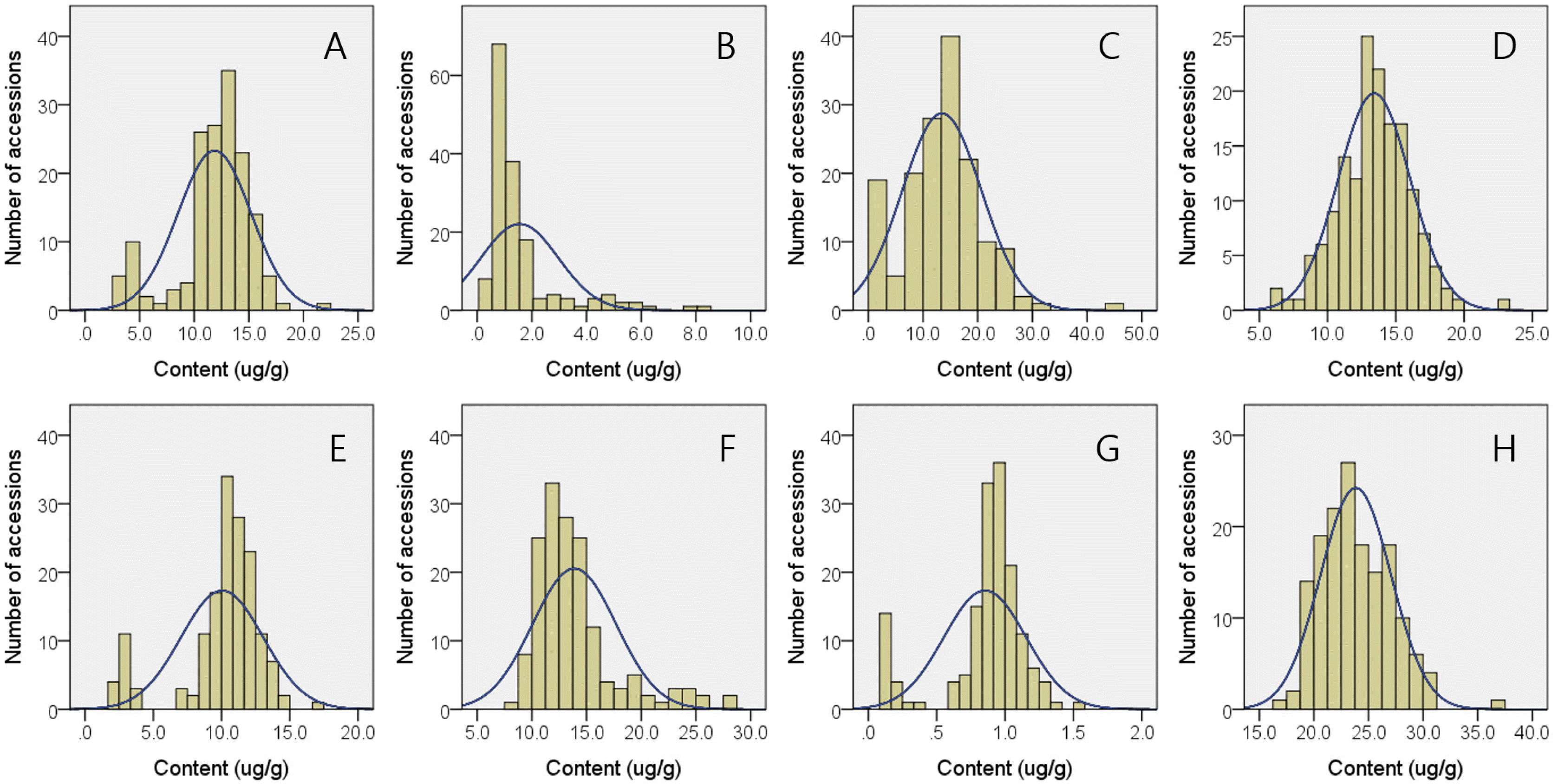
Among 8 vitamer forms of tocols, 4 major ones: α-tocopherol (T), γ-T, α-tocotrienol (T3), and γ-T3 were quantified for 157 Korean bred-rice varieties. The average contents of α-T, γ-T, α-T3, γ-T3, total T, total T3, and total tocols (vitamin E) were 11.9, 1.6, 10.0, 13.9, 13.4, 23.9, and 37.3 μg/g, respectively (
Table 1), and α-T, α-T3 and γ-T3 consisted 32%, 27%, and 37% of total tocols. The range of minimum and maximum contents for α-T, α-T3, and γ-T3 were 3.0-21.6, 1.8-17.1, and 8.2-27.7 μg/g so that the range of total T and total T3, and total tocols were 6.4-22.9, 17.4-36.8, and 26.8-54.9 μg/g, respectively. The accessions which showed the highest and lowest content for each tocol are as followings; α-T (RWG225, 21.6 μg/g and RWG204, 3.0 μg/g), α-T3 (RWG287, 17.1 μg/g and RWG238, 1.8 μg/g), γ-T (RWG165, 8.2 μg/g and RWG153, 0.3 μg/g), and g -T3 (RWG163, 27.7 μg/g and RWG216, 8.2 μg/g) (
Table 2). The tocols contents exhibited quite different compositional pattern between rice ecotypes. The average content of α-T in japonica-type varieties was 12.9 μg/g, which was about 2.5 times higher than that of indica-type ones. Similarly, α-T3 showed about 3 times higher contents in japonica-type rice (11.0 μg/g) compared to indica-type (3.8 μg/g), while γ-T and γ-T3 contents in japonica-type exhibited only 24% and 61% of indica-type varieties, respectively. These results are strongly related with ecotype-dependent differences in α-T/γ-T and α-T3/γ-T3 ratios in that japonica-type ones showed significantly higher α-T/γ-T (15.15) and α-T3/γ-T3 (0.95) ratio corresponding to 629% and 428% of indica-type ones, whose α-T/γ-T and α-T3/γ-T3 ratios were 2.41 and 0.22, respectively. These ecotype-dependent variations are responsible for bimodal distributions observed in histograms of α-T and α-T3 content as well as α-T/γ-T and α-T3/γ-T3 ratio (
Fig. 1). Regardless of ecotypes, total tocopherol contents were always lower than total tocotrienol contents so that the T/T3 ratios in 137 japonica-, 21 indica-types, and all 157 accessions were 0.60, 0.40, and 0.57, respectively. Although total tocotrienol contents showed no difference between ecotypes, japonica-type varieties showed statistically higher total tocopherol (14.0 μg/g) and total tocols (37.8 μg/g) contents compared to indica-type accessions.
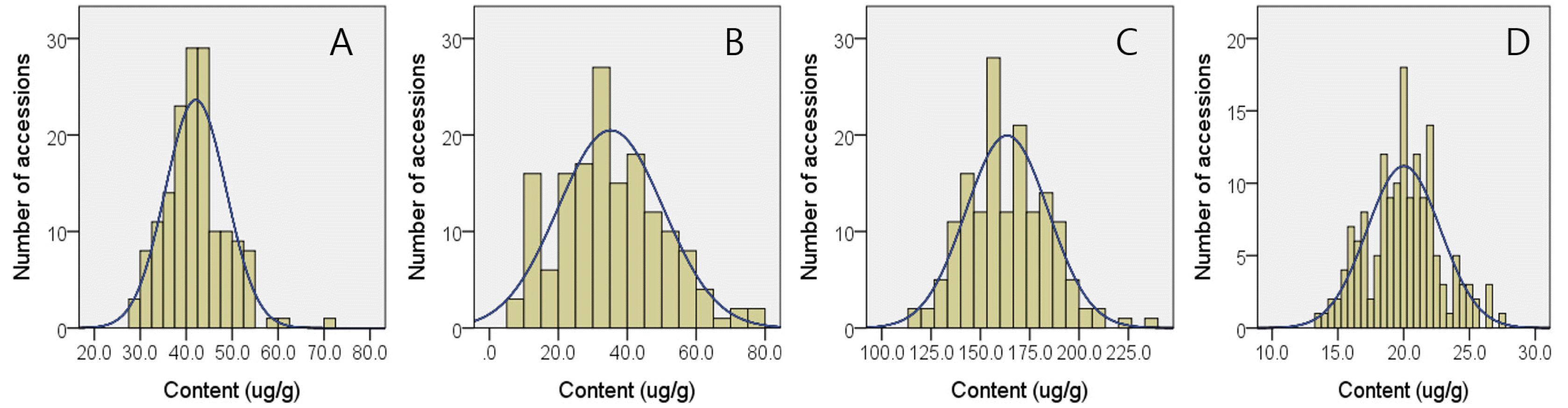
The squalene contents in 157 Korean-bred rice varieties ranged from 7.9 (RWG 258) to 78.4 μg/g (RWG225) with an average of 35.2 μg/g (
Tables 1,
2). Although both japonica-(7.9-78.4 μg/g) and indica-type (8.1-76.4 μg/g) varieties showed similar range of minimum and maximum squalene contents, 2.5 times higher average squalene contents were observed in japonica-type varieties (38.3 μg/g) compared to indica-type (15.2 μg/g) ones. Among 157 accession, highest and lowest brown rice campesterol contents were observed in RWG225 (71.4 μg/g) and RWG165 (27.8 μg/g), respectively and average CA content in 157 accessions was 42.1 μg/g. In the case of stigmasterol, RWG293 (27.7 μg/g) and RWG 190 (13.5 μg/g) recorded the highest and lowest content accessions, respectively and its average content was 20.0 μg/g. Among phytosterols, sitosterol was the major one consisting of over 70% of all phytosterols. Consequently, sitosterol showed relatively higher average content (163.5 μg/g) compared to CA and ST and its minimum and maximum contents were 114.8 μg/g and 235.3 μg/g as observed in RWG190 and RWG293, respectively. Total phytosterols content ranged from 162.9 μg/g (RWG185) to 320.2 μg/g (RWG225). Unlikely to the cases of tocols, both japonica- and indica-type varieties showed similar phytosterols content properties in that japonica-type varieties showed campesterol, sitosterol, stigmasterol, and total phytosterol contents ranging 28.2-71.4, 114.8-235.3, 13.5-27.7, and 162.9-320.2 μg/g, respectively, which were similar to 27.8-60.4, 141.1-198.6, 14.6-22.2, and 192.6-265.2 μg/g observed in indica-type varieties. Histograms of squalene and 3 phytosterols contents in 157 Korean-bred rice varieties are represented in
Fig. 2.
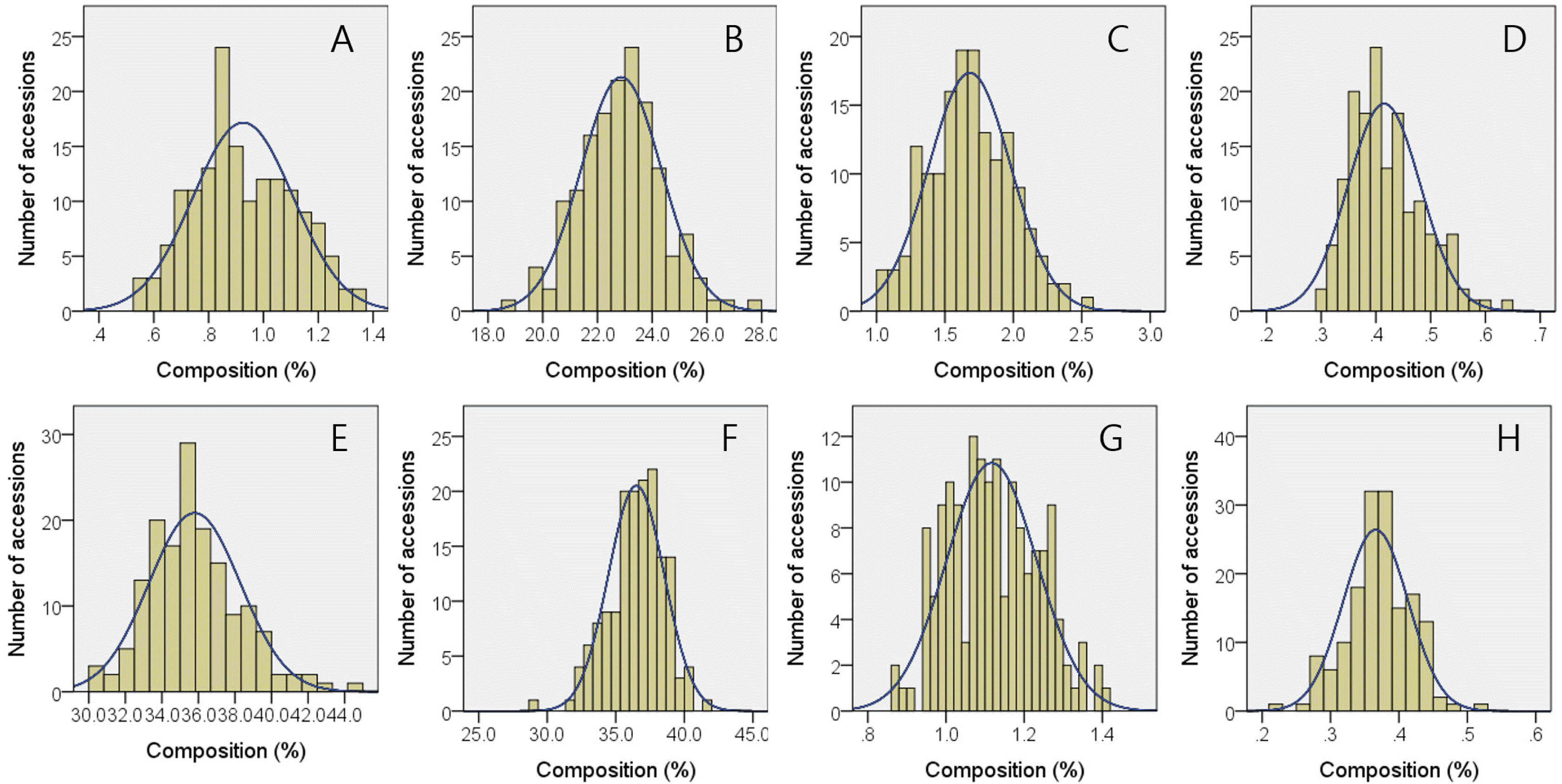
The results presented in
Table 3 revealed that major fatty acids in brown rice of 157 Korean-bred varieties were linoleic, oleic, palmitic acids consisting 36.5%, 35.8%, and 22.9% of 9 kinds of quantified fatty acids. Besides these major ones, stearic, linolenic, palmitic, arachidic, eicosenoic, and behenic acid showed average compositions of 1.68%, 1.12%, 0.93%, 0.41%, 0.37%, and 0.34%, respectively. These fatty acids compositions resulted in 26.25% of saturated, 36.16% of mono-unsaturated, and 37.61% of polyunsaturated fatty acid in brown rice of 157 Korean-bred rice varieties. Ecotype-dependent difference in fatty acid compositions were observed in that japonica-type 137 varieties showed higher average compositions in palmitic (22.57%), stearic (1.66%), oleic (36.10%), arachidic (0.41%), eicosenoic (0.38%), behenic (0.34%), and total saturated (25.91%) and mono-unsaturated (36.47%) fatty acids compared to indica-type varieties, while no statistical differences were observed in myristic, linoleic, linolenic, and polyunsaturated fatty acids. Histograms for brown rice fatty acid compositions are given in
Fig. 3.
All tested phytonutrients content and fatty acid composition data were used for correlationship (
Fig. 4,
Supplementary Table S2) and multivariate (
Fig. 5) analyses. Positive correlations could be observed between α-T and α-T3 (
r = 0.841**), γ-T and γ-T3 (
r = 0.703**), and α-T/γ-T and α-T3/γ-T3 (
r = 0.587**). Total tocols content showed relatively high positive correlations with α-T (
r = 0.603**) and α-T3 (
r = 0.559**) compared to γ-T or γ-T3. Quantified 3 kinds of phytosterols: campesterol, sitosterol and stigmasterol showed positive correlations between each other. No distinct relationship of squalene with other phytonutrients could be observed in our experiments. Among fatty acids, positive correlations could be observed between arachidic and stearic acids (
r = 0.684**), while negative correlations were found in palmitic and oleic acid (
r = ‒0.662**) and linoleic and oleic acids (
r = ‒0.810**). No distinct (
r > 0.6 or
r < 0.6) positive or negative relationships could be observed among fatty acid compositions and any of tested tocols, squalene, and phytosterol contents (
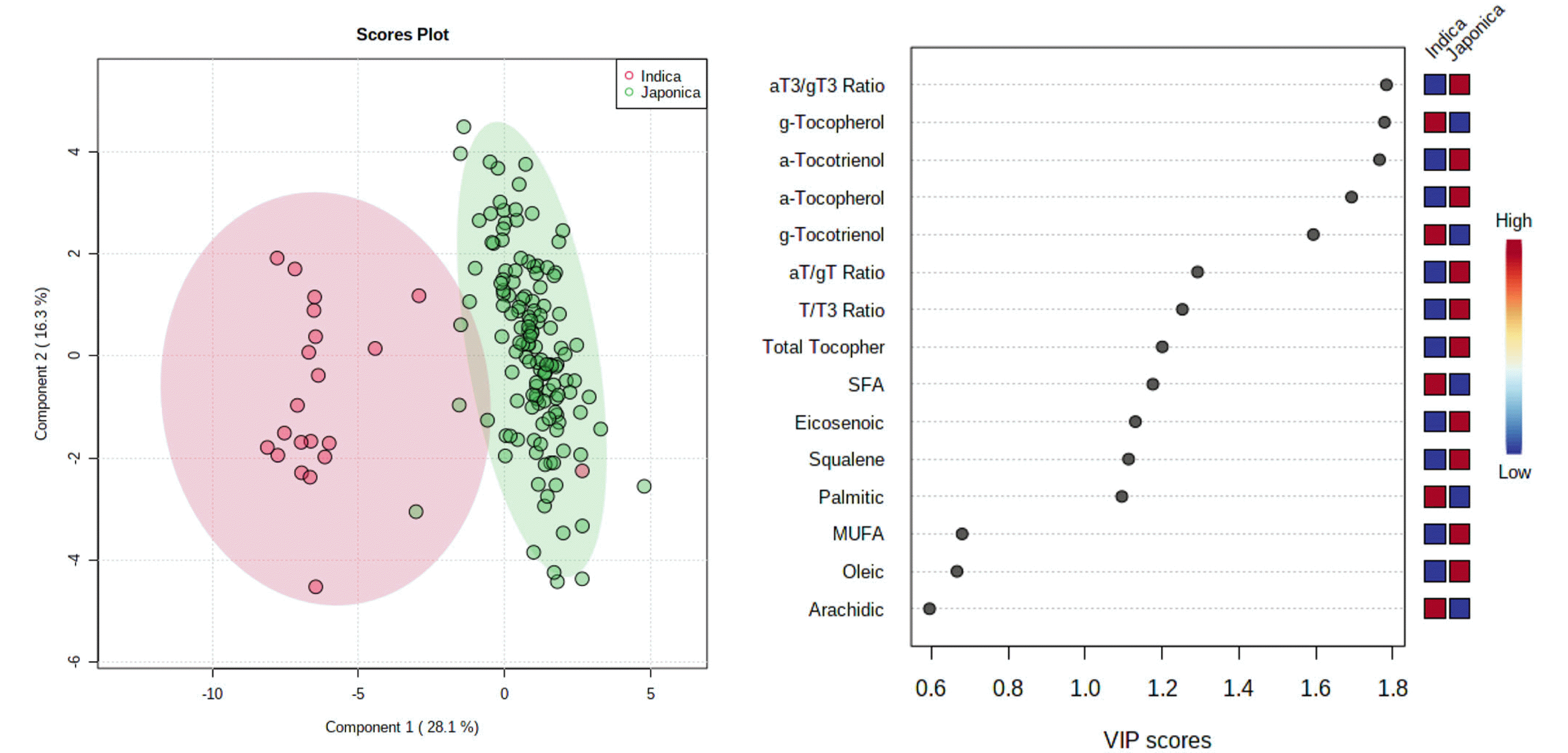
Supplementary Table S2). Phytonutrient contents and fatty acid composition-based differentiation of indica- and japonica-type varieties were tried by multivariate analysis, and partial least squares discriminant analysis (PLS-DA) successfully separated japonica- and indica-varieties (
Fig. 5A). Based upon variable importance projection (VIP) scores, α-T3/γ-T3 ratio (VIP score 1.78), γ-T (VIP score 1.78), and α-T3 (VIP score 1.77) followed by α-T (VIP score 1.69) and γ-T3 (VIP score 1.59) were proved as the major components which can be used for differentiating rice ecotypes (
Fig. 5B).
DISCUSSION
Long history of rice cultivation in Korean peninsula provided not only diverse wild and weedy rices but also various traditionally cultivated landraces. Modernized rice breeding programs in Korea started from 1936 have produced more than 600 Korean rice varieties (
Cho et al. 2020) until now. In order to counteract recently increasing customers’ demands on health-beneficial effects of rice, 136 japonica-type and 21 indica-type varieties that had been bred and cultivated in Korea were selected and their phytonutrient properties were evaluated.
In the present study, average contents of α-T, γ-T, α-T3, and γ-T3 were 11.9, 1.6, 10.0, and 13.9 μg/g, respectively. Although another form of tocols such as β-T, δ-T, β-T3 were also detected and quantified in our experiments, their average contents were as low as 0.69, 0.60, and 0.28 μg/g, respectively (data not shown) so that their data were provided in this report considering their relatively low content and subsequent low importance in health-related effects. These findings are quite similar to previous reports (
Huang et al. 2015;
Wang et al. 2015;
Yu et al. 2016;
Gong et al. 2020). In the case of phytonutrient studies conducted with white rice (
Kim et al. 2012), lower contents of tocols were observed, which may due to the lack of rice bran in white rice which contains most of lipophilic phytonutrients of rice. Ecotype-dependent differences in ratios of α-T/γ-T and α-T3/γ-T3 shown in our results (
Table 1) is a well-defined phenomenon, which is mainly due to the difference of
OsγTMT gene responsible for converting γ-T form of vitamin E into α-T form (
Wang et al. 2015). As described in
Fig. 4 and
Supplementary Table S2, positive correlation-ships between α-T and α-T3, γ-T and γ-T3, as well as α-T/γ-T ratios and α-T3/γ-T3 ratios also suggested unique and strong effects of
OsγTMT gene in determining compositional property of tocols.
Unlikely to the case tocopherols, studies on tocotrienol form of vitamin E are limited, and especially in the case biosynthetic pathway of tocotrienols it is assumed that same genes involved in tocopherol synthesis may also work for tocotrienol synthesis (
Lu et al. 2013) considering structural similarities between tocopherols and tocotrienols. In this study, the average α-T/γ-T ratio in japonica-type varieties was 15.2, which was about 6.3 times higher than average α-T/γ-T ratio observed in indica-type varieties. In the case of α-T3/γ-T3 ratio, however, japonica-type varieties showed value of 0.95, corresponding to only 4.3 times higher than that of indica-type varieties (
Table 1). These results showing inconsistency between α-T/γ-T and α-T3/γ-T3 ratios suggest possible presence of another enzyme transferring γ-T3 into α-T3 and their activity differences between japonica- and indica-type rice varieties. The compositional ratio of total tocopherols to total tocotrienols contents also showed significant differences between ecotypes in that japonica-type rice showed average value of 0.60, which was 1.5 times higher than 0.40 observed in indica-type varieties. These results suggest the ecotype-dependent differences in homogentisate geranyl-geranyl transferase (
HGGT) gene which produces 2-methyl-6-genranylgeranlybenzoquinol by condensing geranylgeranyl diphosphate (GGDP) and homogentisic acid (
Cahoon et al. 2003). Further genetic studies dissecting differences between indica- and japonica-type rice related
HGGT may elucidate pathways and accumulation of tocotrienols form of vitamin E, which shows stronger anticancer capacity than tocopherol form of vitamin E (
Jiang 2019).
Among tested 157 varieties, squalene content ranged from 7.9 to 78.4 μg/g with an average of 35.2 μg/g, which was similar to previous report of
Wang et al. (2019) who reported brown rice squalene contents ranging 4 to 50 μg/g. However,
Yoon et al. (2012) reported slightly lower level of 4.99 to 15.88 μg/g squalene content variations during 3 month storage of 7 rice varieties. Regarding ecotype-tendency of squalene, as shown in
Table 1, japonica-type varieties showed significantly higher than squalene (38.3 μg/g) contents compared to average contents of indica-types (15.2 μg/g). Such ecotype-dependent variation in squalene contents has already been reported as being affected by the gene locus Os09g0319800 (
Wang et al. 2019).
The 157 Korean-bred varieties used in this experiment consisted of 13 varieties developed from 1930’s to 1969, and 15, 24, 41, 50, and 14 varieties developed in 1970’s, 1980’s, 1990’s, 2000’s and 2010’s, respectively. When any trends in variety development year-dependent changing patterns in phytonutrient contents were evaluated, it was found that most of tested phytonutrients were rarely affected by development decades (
Supplementary Table S3). Exceptional cases were the rice varieties developed in 1970’s, which showed relatively lower contents of α-T (8.7 μg/g) and α-T3 (6.8 μg/g) but relatively high γ-T (3.1 μg/g) and γ-T3 (18.0 μg/g) compared to varieties developed in an-other decades. These findings, however, is deeply related with the fact that 1970’s was the decade when indica-type high yield rice variety were highly developed and released. As already described, indica-type rice contains lower α-T and α-T3 but higher γ-T and γ-T3 (
Table 1). Consequently, abovementioned variety development decade-dependent changes in phytonutrients are deeply related with ecotypes of varieties mainly developed at each decade.
Among 3 major fatty acids, palmitic and stearic acids and consequently saturated and mono-unsaturated fatty acid compositions were lower in japonica-type varieties compared to indica-type ones (
Table 3) and these results were consistent previous report (
Goffman et al. 2003). Polyunsaturated fatty acids such as linoleic and linolenic acids showed no ecotype-dependent differences. Although rice is not a lipid-enriched oil seed, lipid composition could be directly related with palatability and subsequent market quality of rice (
Yoon et al. 2012). As an example, storage conditions are known to affect lipids and fatty acids in brown rice (
Tsuzuki et al. 2014), and oxidations or degradations of lipids may readily produce various smaller molecular-weight lipid compounds, aldehydes, and ketones which may in turn cause off-flavors (
Jelen 2006), especially in long-term stored rice.
Application of multivariate analysis such as PLS-DA revealed that profiles of phytonutrients can be used for differentiation of rice ecotypes (
Fig. 5), and α-T3/γ-T3, α-T, α-T3, α-T, and γ-T were the major components in determining ecotype of rice based upon their high VIP scores (
Fig. 5B). Exceptional two cases were RWG164 and RWG262, which exhibited phytonutrient properties more similar to japonica- and indica-type, even though whose ecotypes are indica- and japonica-, respectively. Further studies on why RWG164, an indica-type accession could have high α-T content as well as high α-T/γ-T and α-T3/γ-T3 ratios similar to the levels of japonica-type varieties seems to be required.
In this report, genetic variations in tocols, squalene, phytosterols contents and fatty acid compositions in brown rice of Korean-bred 157 rice varieties were presented together with a list of varieties containing exceptionally high or low quantity of each phytonutrient. These findings may serve as genetic resource information for breeding a superior rice variety with higher nutritional value.
SUPPLEMENTARY MATERIALS
ACKNOWLEDGEMENTS
This work was supported by a grant from the Next-Generation BioGreen 21 Program (Plant Molecular Breeding Center No. PJ013280), Rural Development Administration, Republic of Korea and the Research Fund of Soonchunhyang University, Asan, Republic of Korea.
Fig. 1Histograms with expected normal distribution curves for phytonutrients in brown rice of 157 cultivars bred in Korea. (A) α-tocopherol, (B) γ-tocopherol, (C) ratio of α- to γ-tocopherol, (D) total tocopherol, (E) α-tocotrienol, (F) γ-tocotrienol, (G) ratio of α- to γ-tocotrienol, (H) total tocotrienol.
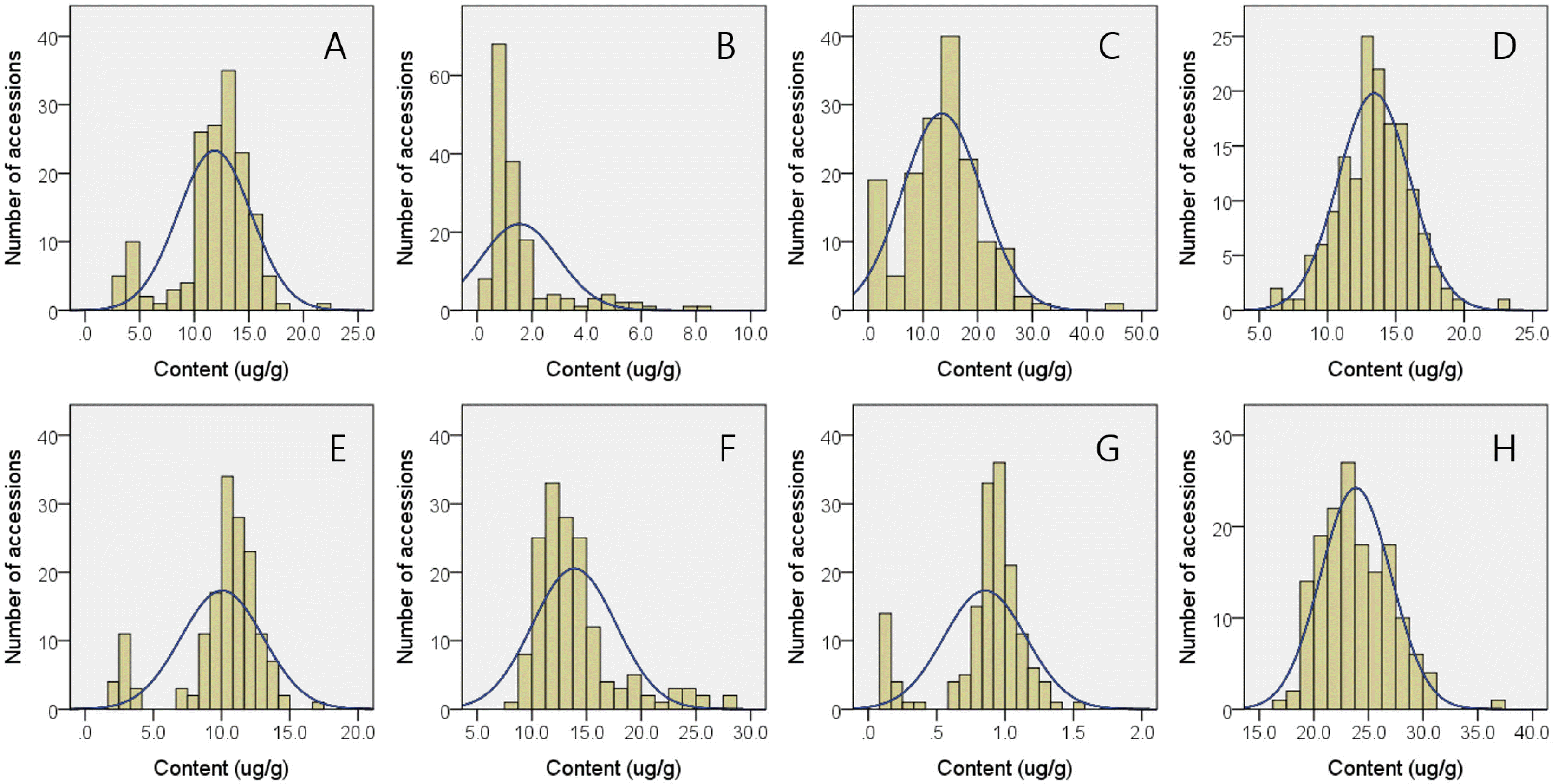
Fig. 2Histograms with expected normal distribution curves for squalene (A), campesterol (B), sitosterol (C), and stigmasterol (D) contents in brown rice of 157 cultivars bred in Korea.
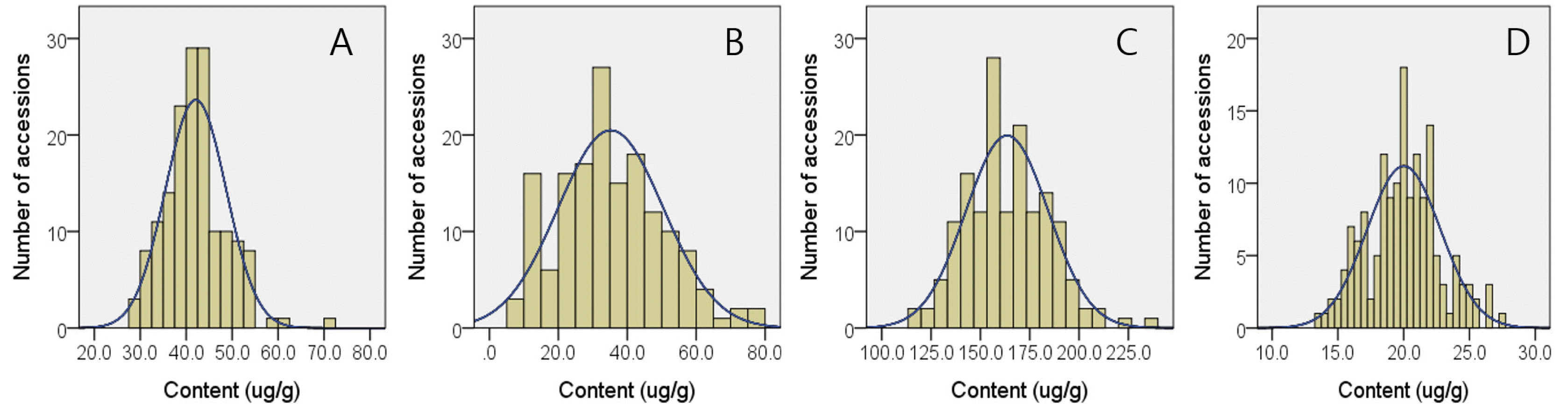
Fig. 3Histograms with expected normal distribution curves for fatty acid composition (%) in brown rice of 157 cultivars bred in Korea. (A) Myristic acid, (B) palmitic acid, (C) stearic acid, (D) behenic acid, (E) oleic acid, (F) linoleic acid, (G) linolenic acid, (H) eicosenoic acid.
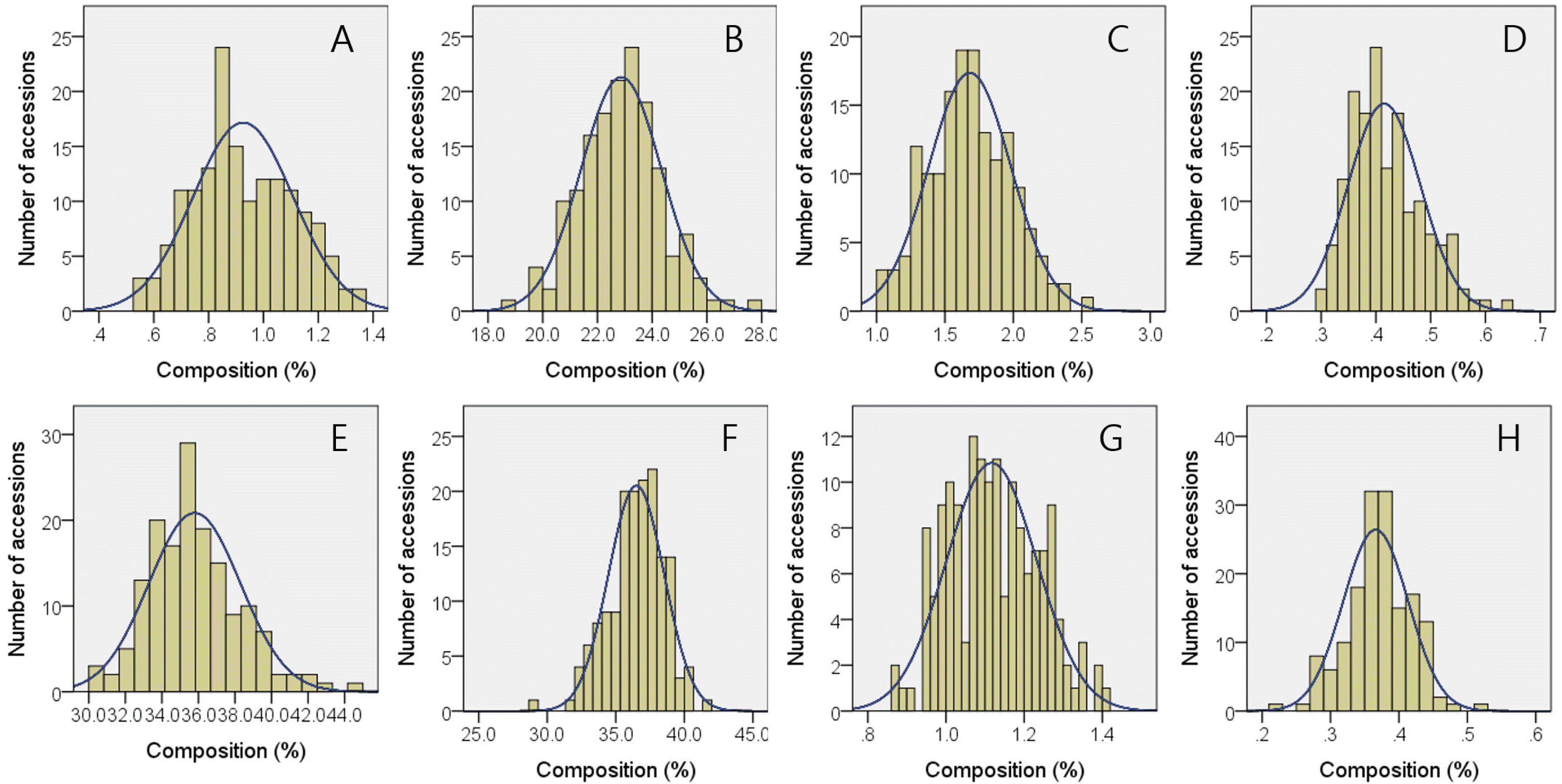
Fig. 4Correlations among phytonutrients and fatty acids in brown rice of 157 Korean-bred varieties. Both japonica and indica type rice data were pooled and used for statistical analyses.
Fig. 5Score plots of partial least squares discriminant analysis (PLS-DA) (A) and variable importance in projection (VIP) score plot (B) of phytonutrient contents and fatty acid compositions in brown rice of 157 varieties bred in Korea consisting of 136 japonica- and 21 indica-type varieties.
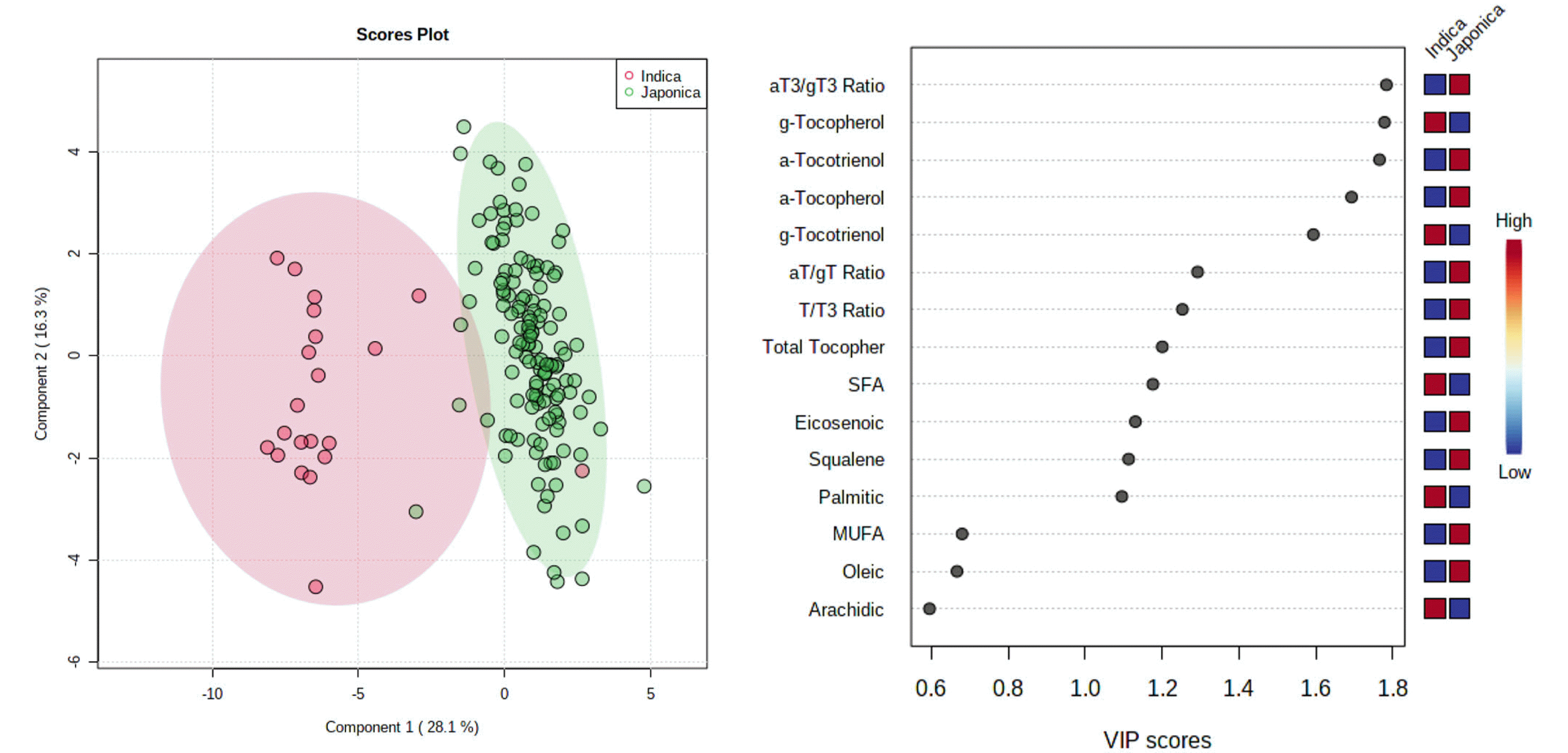
Table 1Statistics for tocopherol (T), tocotrienol (T3), squalene (SQ), campesterol (CA), sitosterol (SI), and stigmasterol (ST) contents (μg/g) in brown rice of 157 varieties bred in Korea.
Table 1
|
Ecotype |
Statistics |
α-T |
γ-T |
α-T3 |
γ-T3 |
SQ |
CA |
SI |
ST |
αT/gT ratio |
αT3/gT3 ratio |
T/T3 ratio |
Total T |
Total T3 |
Total tocols |
Total phytosterols |
|
Indica |
Case |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
|
Mean |
5.2 |
4.5 |
3.8 |
21.0 |
15.2 |
42.8 |
169.2 |
18.1 |
2.41 |
0.22 |
0.40 |
24.8 |
34.5 |
34.5 |
230.0 |
|
SDz)
|
2.5 |
1.9 |
2.6 |
4.2 |
14.3 |
9.3 |
18.2 |
2.4 |
4.40 |
0.24 |
0.11 |
3.7 |
4.5 |
4.5 |
23.2 |
|
Median |
4.4 |
4.6 |
3.0 |
20.7 |
12.0 |
40.7 |
169.3 |
17.6 |
0.95 |
0.15 |
0.39 |
26.3 |
35.8 |
35.8 |
233.4 |
|
Minimum |
3.0 |
0.8 |
1.8 |
13.4 |
8.1 |
27.8 |
141.1 |
14.6 |
0.59 |
0.10 |
0.24 |
19.3 |
26.8 |
26.8 |
192.6 |
|
Maximum |
12.4 |
8.2 |
13.6 |
27.7 |
76.4 |
60.4 |
198.6 |
22.2 |
19.22 |
1.14 |
0.75 |
30.7 |
40.4 |
40.4 |
265.2 |
|
RSDy) (%) |
49% |
41% |
70% |
20% |
94% |
22% |
11% |
13% |
183% |
107% |
28% |
15% |
13% |
13% |
10% |
|
Japonica |
Case |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
|
Mean |
12.9 |
1.1 |
11.0 |
12.8 |
38.3 |
42.0 |
162.6 |
20.3 |
15.15 |
0.95 |
0.60 |
23.7 |
37.8 |
37.8 |
224.9 |
|
SD |
2.0 |
0.5 |
1.6 |
2.2 |
13.0 |
6.1 |
21.2 |
2.7 |
6.01 |
0.16 |
0.09 |
3.1 |
4.6 |
4.6 |
27.7 |
|
Median |
12.8 |
1.0 |
11.0 |
12.5 |
36.2 |
41.6 |
159.7 |
20.2 |
14.30 |
0.94 |
0.59 |
23.3 |
37.9 |
37.9 |
224.4 |
|
Minimum |
8.6 |
0.3 |
6.9 |
8.2 |
7.9 |
28.2 |
114.8 |
13.5 |
3.81 |
0.31 |
0.38 |
17.4 |
28.3 |
28.3 |
162.9 |
|
Maximum |
21.6 |
2.9 |
17.1 |
22.5 |
78.4 |
71.4 |
235.3 |
27.7 |
46.57 |
1.56 |
0.95 |
36.8 |
54.9 |
54.9 |
320.2 |
|
RSD (%) |
16% |
44% |
14% |
18% |
34% |
15% |
13% |
14% |
40% |
17% |
15% |
13% |
12% |
12% |
12% |
|
Total |
Mean |
11.9* |
1.6* |
10.0* |
13.9* |
35.2* |
42.1x)
|
163.5x)
|
20.0* |
13.45* |
0.85* |
0.57* |
23.8* |
37.3x)
|
37.3* |
225.6x)
|
|
SD |
3.4 |
1.4 |
3.0 |
3.8 |
15.3 |
6.6 |
20.9 |
2.8 |
7.26 |
0.30 |
0.12 |
3.2 |
4.7 |
4.7 |
27.1 |
|
RSD (%) |
28% |
91% |
30% |
27% |
43% |
16% |
13% |
14% |
54% |
35% |
20% |
20% |
14% |
13% |
12% |
Table 2List of varieties which showed highest and lowest level of phytonutrients. The RWG accession numbers followed by its phytonutrient content are provided. Information on details of each RWG accession is provided in Supplementary Table S1.
Table 2
|
Phytonutrient |
Description |
RWG accession number (phytonutrient content, in μg/g) |
|
α-Tocopherol |
Highest |
225 (21.6) |
179 (17.6) |
143 (16.8) |
198 (16.7) |
158 (16.6) |
291 (16.5) |
224 (16.5) |
221 (16.2) |
|
Lowest |
204 (3.0) |
205 (3.0) |
208 (3.2) |
209 (3.3) |
272 (3.4) |
210 (3.8) |
|
|
|
α-Tocotrienol |
Highest |
287 (17.1) |
148 (14.3) |
155 (14.3) |
291 (14.0) |
150 (14.0) |
|
|
|
|
Lowest |
238 (1.8) |
165 (2.1) |
166 (2.1) |
208 (2.3) |
211 (2.5) |
|
|
|
|
γ-Tocopherol |
Highest |
165 (8.2) |
166 (8.0) |
206 (6.2) |
|
|
|
|
|
|
Lowest |
153 (0.3) |
215 (0.4) |
240 (0.4) |
181 (0.4) |
|
|
|
|
|
γ-Tocotrienol |
Highest |
163 (27.7) |
276 (27.6) |
211 (25.2) |
292 (25.0) |
166 (24.7) |
|
|
|
|
Lowest |
216 (8.2) |
275 (8.8) |
280 (8.9) |
196 (9.2) |
264 (9.2) |
195 (9.4) |
|
|
|
Squalene |
Highest |
225 (78.4) |
164 (76.4) |
222 (73.7) |
219 (72.5) |
|
|
|
|
|
Lowest |
258 (7.9) |
209 (8.1) |
204 (8.6) |
|
|
|
|
|
|
Campesterol |
Highest |
225 (71.4) |
292 (60.4) |
143 (57.8) |
|
|
|
|
|
|
Lowest |
165 (27.8) |
268 (28.2) |
164 (28.5) |
|
|
|
|
|
|
Sitosterol |
Highest |
187 (235.3) |
225 (224.8) |
290 (208.4) |
287 (208.4) |
|
|
|
|
|
Lowest |
190 (114.8) |
185 (116.8) |
212 (121.4) |
199 (125.0) |
157 (126.9) |
193 (127.7) |
|
|
|
Stigmasterol |
Highest |
293 (27.7) |
240 (26.7) |
139 (26.5) |
187 (26.4) |
|
|
|
|
|
Lowest |
190 (13.5) |
185 (14.2) |
157 (14.4) |
209 (14.6) |
199 (14.8) |
|
|
|
|
Total tocopherol |
Highest |
225 (22.9) |
179 (19.2) |
224 (19.0) |
198 (18.6) |
158 (18.3) |
143 (18.2) |
|
|
|
Lowest |
205 (6.4) |
208 (6.5) |
292 (6.9) |
|
|
|
|
|
|
Total tocotrienol |
Highest |
287 (36.8) |
236 (31.0) |
276 (30.7) |
163 (30.4) |
201 (30.1) |
|
|
|
|
Lowest |
216 (17.4) |
196 (17.7) |
185 (18.7) |
268 (18.9) |
|
|
|
|
|
Total tocols |
Highest |
287 (54.9) |
179 (48.7) |
291 (47.5) |
225 (46.9) |
151 (46.1) |
156 (45.9) |
198 (45.9) |
|
|
Lowest |
204 (26.8) |
208 (27.6) |
205 (28.1) |
209 (28.1) |
268 (28.3) |
|
|
|
|
Total phytosterols |
Highest |
225 (320.2) |
187 (315.2) |
287 (285.1) |
290 (283.9) |
293 (281.9) |
|
|
|
|
Lowest |
185 (162.9) |
190 (163.9) |
212 (169.9) |
199 (171.3) |
157 (172.8) |
|
|
|
Table 3Statistics for brown rice fatty acid composition (%) of 136 japonica- and 21 indica-type cultivars bred in South Korea.
Table 3
|
Ecotype |
Statistics |
Myristic (C14:0) |
Palmitic (C16:0) |
Stearic (C18:0) |
Oleic
(C18:1) |
Linoleic (C18:2) |
Linolenic (C18:3) |
Arachidic (C20:0) |
Eicosenoic (C20:1) |
Behenic (C22:0) |
Saturated fatty acid |
Mono-unsaturated fatty acid |
Poly-unsaturated fatty acid |
|
Indica |
Case |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
21 |
|
Mean |
0.94 |
24.76 |
1.80 |
33.83 |
36.43 |
1.15 |
0.46 |
0.30 |
0.32 |
28.29 |
34.13 |
37.58 |
|
SDz)
|
0.22 |
1.58 |
0.24 |
1.94 |
1.61 |
0.14 |
0.06 |
0.03 |
0.04 |
1.62 |
1.95 |
1.66 |
|
Median |
0.97 |
24.91 |
1.84 |
33.53 |
36.29 |
1.19 |
0.44 |
0.31 |
0.33 |
28.05 |
33.88 |
37.40 |
|
Mode |
0.97 |
19.64 |
1.97 |
30.40 |
38.22 |
1.24 |
0.43 |
0.29 |
0.29 |
27.71 |
30.69 |
34.32 |
|
Minimum |
0.55 |
19.64 |
1.40 |
30.40 |
33.34 |
0.91 |
0.32 |
0.22 |
0.24 |
23.25 |
30.69 |
34.32 |
|
Maximum |
1.37 |
27.90 |
2.41 |
39.32 |
40.12 |
1.34 |
0.59 |
0.36 |
0.37 |
31.52 |
39.63 |
41.46 |
|
Range |
0.82 |
8.26 |
1.01 |
8.92 |
6.78 |
0.43 |
0.27 |
0.14 |
0.13 |
8.27 |
8.94 |
7.14 |
|
RSDy) (%) |
23.3% |
6.4% |
13.5% |
5.7% |
4.4% |
11.8% |
13.4% |
11.5% |
11.3% |
5.7% |
5.7% |
4.4% |
|
Japonica |
Case |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
136 |
|
Mean |
0.92 |
22.57 |
1.66 |
36.10 |
36.51 |
1.11 |
0.41 |
0.38 |
0.34 |
25.91 |
36.47 |
37.62 |
|
SD |
0.18 |
1.22 |
0.31 |
2.45 |
2.10 |
0.11 |
0.06 |
0.04 |
0.04 |
1.19 |
2.47 |
2.14 |
|
Median |
0.90 |
22.66 |
1.66 |
35.79 |
36.70 |
1.10 |
0.39 |
0.37 |
0.34 |
25.99 |
36.16 |
37.84 |
|
Mode |
0.84 |
21.92 |
1.70 |
33.52 |
36.00 |
1.07 |
0.39 |
0.37 |
0.32 |
25.26 |
34.66 |
37.02 |
|
Minimum |
0.56 |
18.99 |
1.04 |
30.49 |
28.85 |
0.87 |
0.30 |
0.27 |
0.26 |
23.05 |
30.80 |
29.93 |
|
Maximum |
1.34 |
25.23 |
2.55 |
44.26 |
41.59 |
1.40 |
0.64 |
0.52 |
0.46 |
28.57 |
44.70 |
42.83 |
|
Range |
0.78 |
6.24 |
1.51 |
13.77 |
12.74 |
0.53 |
0.34 |
0.25 |
0.20 |
5.52 |
13.90 |
12.90 |
|
RSD (%) |
19.2% |
5.4% |
18.3% |
6.8% |
5.7% |
10.1% |
15.8% |
11.0% |
12.5% |
4.6% |
6.8% |
5.7% |
|
Total |
Mean |
0.93x)
|
22.87* |
1.68* |
35.79* |
36.50x)
|
1.12x)
|
0.41* |
0.37* |
0.34* |
26.23* |
36.16* |
37.61x)
|
|
SD |
0.18 |
1.47 |
0.30 |
2.50 |
2.04 |
0.12 |
0.07 |
0.05 |
0.04 |
1.49 |
2.53 |
2.08 |
|
RSD (%) |
19.7% |
6.4% |
17.9% |
7.0% |
5.6% |
10.3% |
16.0% |
13.0% |
12.5% |
5.7% |
7.0% |
5.5% |
References
- Ahn SM. 2010. The emergence of rice agriculture in Korea: archaeobotanical perspectives. Archaeol. Anthropol. Sci.. 2: 89-98.
- Bergman CJ, Xu Z. 2003. Genotype and environment effects on tocopherol, tocotrienol and gamma-oryzanol contents of Southern US rice. Cereal Chem.. 80: 446-449.
- Cahoon EB, Hall SE, Ripp KG, Ganzke TS, Hitz WD, Coughlan SJ. 2003. Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content. Nat. Biotechnol.. 21: 1082-1087.
- Cho KS, Kim HJ, Moon SM, Kang JH, Lee YS. 2006. Optimization of one-step extraction/methylation method for analysis of fatty acid composition in brown rice. Korean J. Crop Sci.. 51: 89-94.
- Cho YC, Baek MK, Park HS, Cho JH, Ahn EK, Suh JP, et al. 2020. History and results of rice breeding in Korea. Korean J. Breed. Sci.. 52: 58-72.
- Cho YH, Lim SY, Rehman A, Farooq M, Lee DJ. 2019. Characterization and quantification of γ-oryzanol in Korean rice landraces. J. Cereal Sci.. 88: 150-156.
- Choi B, Yoo JM, Lee SB, Kim GJ, Kim KW, Yoo JH, et al. 2018. Genome wide association study of rice core set related to grain weight. Plant Breed. Biotech.. 6: 293-304.
- Goffman FD, Pinson S, Bergman C. 2003. Genetic diversity for lipid content and fatty acid profile in rice bran. J. Am. Oil Chem. Soc.. 80: 485-490.
- Gong ES, Liu C, Li B, Zhou W, Chen H, Li T, et al. 2020. Phytochemical profiles of rice and their cellular antioxidant activity against ABAP induced oxidative stress in human hepatocellular carcinoma HepG2 cells. Food Chem.. 318: 126484
- Huang SH, Tung YH, Ng LT. 2015. Influences of environmental conditions on tocopherols tocotrienols and γ-oryzanol contents in rice grains. Taiwanese Journal of Agricultural Chemistry and Food science. 53: 216-224.
- Huang ZR, Lin YK, Fang JY. 2009. Biological and pharma-cological activities of squalene and related compounds: potential uses in cosmetic dermatology. Molecules. 14: 540-554.
- Jelen HH. 2006. Solid-phase microextraction in the analysis of food taints and off-flavors. J. Chromatogr. Sci.. 44: 399-415.
- Jiang Q. 2019. Natural forms of vitamin E and metabolites regulation of cancer cell death and underlying mechanisms. IUBMB Life. 71: 495-506.
- Jung YJ, Park JH, Shrestha S, Song MC, Cho S, Lee CH, et al. 2014. Phytosterols from the rice (Oryza sativa) bran. J. Appl. Biol. Chem.. 57: 175-178.
- Kim JK, Ha SH, Park SY, Lee SM, Kim HJ, Lim SH, et al. 2012. Determination of lipophilic compounds in genetically modified rice using gas chromatographytime-of-flight mass spectrometry. J. Food Compos. Anal.. 25: 31-38.
- Kitta K, Ebihara M, Iizuka T, Yoshikawa R, Isshiki K, Kawamoto S. 2005. Variations in lipid content and fatty acid composition of major non-glutinous rice cultivars in Japan. J. Food Compos. Anal.. 18: 269-278.
- Lee KJ, Lee JR, Shin MJ, Cho GT, Ma KH, Lee GA, et al. 2018. Genetic diversity and population structure of a Korean rice germplasm based on DNA profiles. Korean J. Crop Sci.. 63: 1-7.
- Lu Y, Rijzaani H, Karcher D, Ruf S, Bock R. 2013. Efficient metabolic pathway engineering in transgenic tobacco and tomato plastids with synthetic multigene operons. Proc. Natl. Acad. Sci. U. S. A.. 110: E623-632.
- Mahmud MMC, Oh Y, Kim TH, Cho YH, Lee YS. 2018. Effects of milling on aromatics lipophilic phytonutrients and fatty acids in unprocessed white rice of scented rice 'Cheonjihyang-1-se'. Food Sci. Biotechnol.. 27: 383-392.
- Qureshi AA, Sami SA, Khan FA. 2002. Effects of stabilized rice bran, its soluble soluble and fiber fractions on blood glucose levels and serum lipid parameters in humans with diabetes mellitus Types I and II. J. Nutr. Biochem.. 13: 175-187.
- Sohail M, Rakha A, Butt MS, Iqbal MJ, Rashid S. 2017. Rice bran nutraceutics: A comprehensive review. Crit. Rev. Food Sci. Nutr.. 57: 3771-3780.
- Tan BL, Norhaizan ME. 2017. Scientific evidence of rice byproducts for cancer prevention: Chemopreventive properties of waste products from rice milling on carcinogenesis in vitro and in vivo. Biomed. Res. Int.. 2017: 1-18.
- Tsuzuki W, Suzuki Y, Yamada S, Kano S, Ohnishi H, Fujimoto T, et al. 2014. Effect of oxygen absorber on accumulation of free fatty acids in brown rice and whole grain wheat during storage. LWT. 58: 222-229.
- Wang XQ, Kim KW, Chu SH, Phitaktansakul R, Park SW, Chung IM, et al. 2019. Genome wide association study for squalene contents and functional haplotype analysis in rice. ACS Omega. 4: 19358-19365.
- Wang XQ, Yoon MY, He Q, Kim TS, Tong W, Choi BW, et al. 2015. Natural variations in OsγTMT contribute to diversity of the α-tocopherol content in rice. Mol. Genet. Genomics. 290: 2121-2135.
- Wong RSY, Radhakrishnan AK. 2012. Tocotrienol research: past into present. Nutr. Rev.. 70: 483-490.
- Yoon MR, Koh HJ, Kang MY. 2008. Variation of properties of lipid components in rice endosperm affected on palatability. J. Appl. Biol. Chem.. 51: 207-211.
- Yoon MR, Rico CW, Koh HJ, Kang MY. 2012. A study on the lipid components of rice in relation to palatability and storage. Korean Soc. Appl. Biol. Chem.. 55: 515-521.
- Yu L, Li G, Li M, Xu F, Beta T, Bao J. 2016. Genotypic variation in phenolic acids vitamin E and fatty acids in whole grain rice. Food Chem.. 197: 776-782.
- Zhou Z, Robards K, Helliwell S, Blanchard C. 2002. Com-position and functional properties of rice. Int. J. Food Sci. Technol.. 37: 849-868.