Abstract
Molecular mapping and application of quantitative trait loci (QTL) associated with a higher level of grain Zinc is a viable option to enhance zinc content in rice through breeding. An F2 population derived from a cross between a high yielding variety, BRRI dhan28, and a locally adapted Zn enriched cultivar, Kalobokri was used to map QTLs associated with higher levels of Zn in rice grain. The F2:3 progenies varied significantly (P < 0.0001) in Zinc contents with a mean value remarkably higher than those in the superior parent. Through marker by trait analysis using IciMapping, we detected a large-effect QTL, qGZn3 on chromosome 3 between RM5419 and RM1164 spanning 1.83 Mb interval at the 0.05 level of significance with a threshold LOD of 10.61. This QTL showed a 21.1% (R2 value) contribution to the total phenotypic variation for zinc content in the unpolished rice grains with 4.68 μg/g additive effect of Kalobokri alleles. We also detected 11 metal homeostasis related genes within the interval of qGZn3. In-silico analysis showed that four expressed sequence tags of one candidate gene (LOC_Os03g22810) encoding Cu/Zn superoxide dismutase, a metal-binding protein, are highly active in the endosperm as well as in the embryonic tissue of rice grain. Taken together, our results suggest that qGZn3 is a major QTL associated with high grain Zn content in the F2 progenies of rice. Our findings offer valuable genetic resources to facilitate breeding for high yielding and Zinc-enriched rice variety.
-
Key words: Biofortification, Quantitative trait loci, Selective genotyping, Simple sequence repeat, Zinc homeostasis related genes
INTRODUCTION
Malnutrition associated with micronutrient deficiency is recognized as a global public health concern mainly in developing countries. Among the major micronutrient malnutrition, Zinc (Zn) deficiency is mostly predominant in rice-consuming countries like Bangladesh (
Kennedy et al. 2002;
Nakandalage et al. 2016). It has been estimated that about one-third of the world’s population is under threat of Zn deficiency resulting in more than 400,000 children’s (age below 5 years) death every year (
Myers et al. 2015;
Harvest plus 2016). In Asia, the present scenario is worsening gradually, accounting for more than one million people, particularly children and women are extremely suffered from Zn deficiency-related health problems (
Swamy et al. 2016;
Swamy et al. 2018). Therefore, adequate Zn improved diet needs to be ensured for maintaining balanced growth and development, immunologic and reproductive functions to lead a healthy and productive life.
Over the last decade, a good number of attempts have been made to biofortify food crops, especially cereal crops with enhanced levels of micronutrients, which led to a meaningful understanding of the physiological, genetic and molecular basis of high Zn bioaccumulation capacity in grains (
Impa and Johnson-Beebout 2012;
Shahzad et al. 2014;
Nakandalage et al. 2016). In the past, slow progress in the development of improved cultivars was mainly due to poor understanding of genetic factors that impart Zn uptake, translocation, and uploading into the grains (
Karthika et al. 2018). Currently, efforts are continuing to ameliorate the efficacy of breeding programs by employing linkage mapping and marker-assisted selection for several complex polygenic traits in rice (
Kumar et al. 2014). Several QTLs on chromosomes 3, 5, 7, 8 and 12 of rice conferring grain Zn content have been reported (
Stangoulis et al. 2007;
Lu et al. 2008;
Garcia-Oliviera et al 2009;
Norton et al. 2010;
Anuradha et al. 2012;
Zhang et al. 2014;
Nawaz et al. 2015;
Swamy et al. 2016;
Swamy et al. 2018;
Jeong et al. 2020;
Lee et al. 2020). These findings suggest that grain Zn in rice is quantitatively inherited and governed by many genes located on different regions of the chromosome with different levels of phenotypic effects (
Lu et al. 2008;
Garcia-Oliveira et al 2009;
Zhang et al. 2011). Although several reports of QTLs for grain Zn content of rice are reported but only a few of them were found useful in breeding programs, others showed inconsistency across the environments and genetic backgrounds (
Swamy et al. 2016). More importantly, most of the high yielding rice varieties (HYV) are poor in grain Zn content (
Graham et al. 1999;
Gregorio et al. 1999;
Lu et al. 2008). On the contrary, several germplasms showed stable and high grain Zn density compared with the cultivated HYV rice (
Hossain et al. 2014), which reflected the existence of an extensive variation in grain Zn content among the rice germplasm. Therefore, it has a great possibility to breed high Zn rice by exploiting locally adapted high grain Zn-donor germplasms, and understanding the genetic architecture of this trait is essential for Zn biofortification of rice.
Rice is the staple food for more than 160 million people of Bangladesh and a primary or secondary major food of about 50% of the world’s total population (
Akter and Basher 2014;
Gross and Zhao 2014). As a matter of fact, rice is a poor source of essential micronutrients (
Cakmak and Kutman 2018) such as Zn in comparison to other staple cereal crops like wheat and maize (
Kennedy et al. 2002;
Sharma et al. 2013;
Kumar et al. 2014). Hence, biofortification of rice is regarded as one of the cost-effective and sustainable strategies to develop new cultivars of rice with improved levels of Zn concentration which might be a possible solution to support in combating Zn malnutrition (
Shahzad et al. 2014;
Swamy et al. 2016). Although recently a few Zn-enriched rice varieties have been released in some South and Southeast Asian countries using conventional breeding techniques (
Harvest Plus 2016), successful application of QTLs for increasing Zn concentration in rice has yet been reported, thereby necessitating further investigation to get in depth insights underlying the genetic control of high Zn content in rice grain.
In the present study, we performed a field trial of an F2 population derived from a cross between BRRI dhan28 (high yielding) and Kalobokri (Zn-rich) varieties of Oryza sativa L. for assessing the variation of Zn content in the grains. The F2 population was used for linkage mapping of QTLs responsible for accumulating high grain Zn content using polymorphic simple sequence repeat (SSR) markers. Our findings provide an opportunity to accelerate the development and deployment of cultivars of rice with high grain Zn content.
MATERIALS AND METHODS
Plant materials
A locally adapted rice cultivar
Kalobokri and a high yielding mega-variety BRRI dhan28 were used in this study.
Kalobokri is a local Aus rice variety having high grain Zn content compared to BRRI dhan28, which is an early maturing
Boro (November to May) variety (
Hossain et al. 2014). The grain of BRRI dhan28 is straw-yellow colored and long slender, while the grain of
Kalobokri is black and medium bold with long awn.
A standing crop of the F1 generation of a cross between BRRI dhan28 and Kalobokri was available in the field laboratory of the Plant Breeding Division of Bangladesh Rice Research Institute (BRRI). At the maturity stage, the F1 plants were confirmed considering grain types, color, presence/absence of awn compared to their parents. Seeds of all the selected F1 plants were harvested in bulk, dried, and used for raising the F2 mapping population.
Evaluation of parents and mapping population for grain Zn content
During Boro season, a total of 806 F2 progenies along with their parents were grown in the field with a planting density of 20 cm × 20 cm using single seedling per hill. The parents were grown in three replications. At maturity of the crop, panicles from the individual plants were collected in brown paper bags and dried in an oven at 40℃ for 3 days to reduce the moisture content to about 12-14%. After drying, panicles were carefully threshed and cleaned manually and weighed. Seeds from 396 plants out of 806, which produced at least ca.20 g dried and cleaned seeds were considered for Zn analysis taking into account that this amount would be good enough to produce at least 12-14 g brown rice after dehulling. The seeds were dehulled in a bench-top mini-dehuller fabricated with Zn and Fe contamination-free polyurethane roller. The brown rice samples were then read in XRF (X-ray Fluorescence) machine (X-supreme 8000, Oxford Instruments) for grain Zn content. The brown rice sample of each parent and F2 plant were divided into three sub-samples (ca.6 g – 10 g) to read as three botanical replications. The samples, which were not sufficient for three replications, were divided into two sub-samples for zinc analysis. Rice grain samples of IR68144 were read as standard reference at every after nine samples analyzed to monitor the precision of the machine to detect grain Zn content.
DNA extraction and polymerase chain reaction (PCR)
Green leaf samples were collected for genomic DNA extraction from all individual F
2 plants and the parents at four weeks after transplanting. Genomic DNA was extracted following the mini prep CTAB method as described in
Virk et al. (1995). SSR genotyping was performed following the protocol described in
Syed et al. (2016) with minor modifications. Briefly, PCR was performed in 10 μL reactions containing 2 μL DNA template (around 50 ng), 1 μL of 10X TB buffer (containing 0.2 μL dd water; 0.5 μL of 1 M Tris-HCl, pH 8.4; 0.2 μL of 5 M KCl; 0.1 μL of 15 μM MgCl2; 0.1×10
‒7 g Gelatin), 0.5 μL of 1 μM dNTPs, 0.5 μL each of 10 μM forward and reverse primers, 5.3 μL nano pure ddH2O and 0.2 μL Taq DNA polymerase (5 U/μL) using a thermal cycler. After initial denaturation for 2 minutes at 94℃, each cycle comprises 30 seconds denaturation at 94℃, 30 seconds annealing at 55℃, and 30 seconds extension at 72℃ with a final extension for 5 minutes at 72℃ at the end of 32 cycles. The PCR product was preserved at 4℃ temperature in the thermal cycler.
For the genotyping of F
2 populations, selective genotyping technique was used in this study according to the previously described method by Lander and
Botstein (1989). A total of 94 F
2 plants comprising 47 from the lower tail and 47 from the upper tail of the frequency distribution of Zn contents of 396 F
2:3 progenies were used in genotyping. Six hundred and twenty SSR markers distributed over 12 chromosomes were analyzed to identify polymorphic markers between BRRI dhan28 and
Kalobokri. SSR analysis was performed following the protocol described above. The amplified PCR products were separated in 6% polyacrylamide gel at 100 V for 1.5 to 2.5 hours against a known DNA ladder. DNA bands were visualized with the exposure to ultra-violet light in the gel documentation system and saved as jpeg format. Allele scoring was performed considering the relative position of the bands in the gel images compared to the position of parental bands.
Linkage map was constructed based on Kosambi function using QTL IciMapping version 4.0 (
www.isbreeding.net/software/?type =detail&id=14). A total of 106 polymorphic SSRs distributed over 12 chromosomes of rice were used to construct a linkage map. Linkage groups were determined using a minimum likelihood ratio (LOD) score of 3.0. The order of the markers in the groups was determined with the ‘nnTwoOpt’ algorithm. QTL analysis was carried out using the composite interval mapping (CIM) option in QTL IciMapping v4.0 (hereafter IciM). Significant QTLs were detected at LOD of 3.45 with a 0.05 level of significance with 1000 iterations in the permutation test. The QTLs were confirmed by further analysis with composite interval mapping option in QGene v3.0.2 (
Nelson 1997) and QTLNetworkv2.0 (
Yang et al. 2008). QTLs were named following the nomenclature system proposed by
McCouch et al (1997), and
McCouch and CGSNL (2008).
The genomic region encompassing
qGZn3 was analyzed
in silico for the presence of putative candidate genes. The physical length and position of
qGZn3 was determined based on the physical position of the flanking markers using Gramene annotated sequence map set (
www.gramene.org) and NCBI (
www.ncbi.nlm.nih.gov) BLAST. The candidate genes underlying
qGZn3 were retrieved and annotated for gene function using Rice Annotation Project Database (RAP-DB) genome browser.
The putative candidate genes underlying the
qGZn3 were analyzed using RiceXPro 3.0 (
Sato et al. 2013), a web-based bioinformatics tool (available at
https://ricexpro.dna.affrc.go.jp/) to navigate the tissue-specific expression pattern. MSU IDs of the candidate genes were used as search query in the keyword search box of ‘ExProFlip’ tab of RiceXPro against the inbuilt embryo and endosperm specific gene expression profile dataset to locate the expression pattern and determine the underlying expressed sequence tags. A heatmap was generated based on clustering of correlation distance and complete linkage of each gene using inbuilt ‘gplots’ package of R with RiceXPro.
One way analysis of variance (ANOVA) procedures in R (
R Development Core Team, 2008) was used to analyze variance and other descriptive statistics for grain Zn content among F
2:3 progenies. Heritability (in broad sense) was estimated from the variance components obtained from the ANOVA. The average grain Zn contents of the parents were compared by the Student’s
t-test at 0.05 and 0.01 levels of significance.
RESULTS
Zn content in unpolished grain samples of the parents and their F2:3 progenies
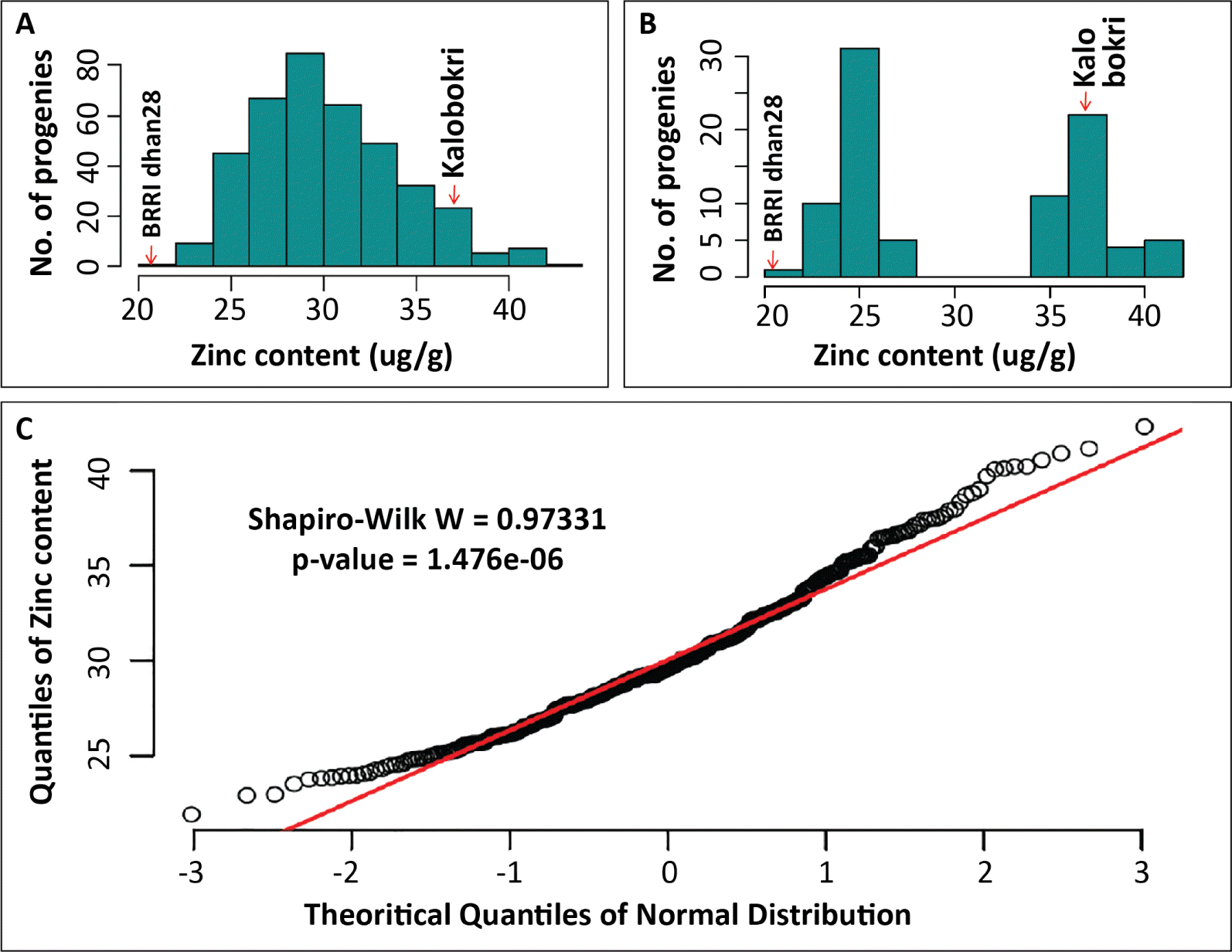
Zn content in the unpolished grains of
Kalobokri (37.5 μg/g) was significantly higher than in BRRI dhan28 (20.2 μg/g). The F
2:3 progenies also showed a wide and significant (
P < 0.0001) variation in Zn contents (
Table 1). The concentration of Zn in the unpolished grains of F
2:3 progenies varied from 21.9 μg/g to 41.3 μg/g with an average of 30.2 μg/g. A high heritability estimate (
Table 1), and a continuous distribution and transgressive segregation in grain Zn content was found among the F
2:3 progenies (
Fig. 1A), which indicated that the trait is controlled quantitatively by many genes. Moreover, the normality test based on Shapiro-Wilk Coefficient showed almost a normal distribution in the Zn content in the 396 F
2:3 genotypes at 0.0001% level of significance (
Fig. 1C). Out of 396 F
2:3 progenies, 47 plants from the lower tail and 47 from the upper tail of the frequency distribution were selected for genotyping. The Zn contents in the upper tail ranged from 34.5 μg/g to 44.0 μg/g, while the lower tailed genotypes had Zn content ranging from 21.9 μg/g to 26.9 μg/g (
Fig. 1B). The progenies at the extreme tails of the frequency distribution of grain Zn content were used in genotyping.
Primer survey is an important prerequisite before starting any QTL mapping work. In this study, we used 620 microsatellite markers distributed over 12 chromosomes of rice to find out polymorphic markers between the two parents. The chromosomal positions of the SSR markers were obtained from the Gramene database (
www.gramene.org). Out of 620 SSR analyzed, we found that 230 and 314 makers were monomorphic and polymorphic, respectively between BRRI dhan28 and
Kalobokri (
Table 2), and the remaining 76 markers were not amplified. The average rate of polymorphism was 50.6%. Among the polymorphic markers, there were many markers very close to each other and in some cases the intervals between the adjacent markers were comparatively higher, which was up to 32.6 cM. For linkage map construction, the markers that showed clear and distinguishable polymorphism and not located very close to each other were considered with the exception in few cases. Segregation analysis was performed in QTL IciMapping v4.0 and the distorted markers with
P-value < 0.05 were discarded. The linkage map was constructed based on 106 polymorphic SSR markers distributed over 12 chromosomes (
Table 2,
Supplementary Fig. S1).
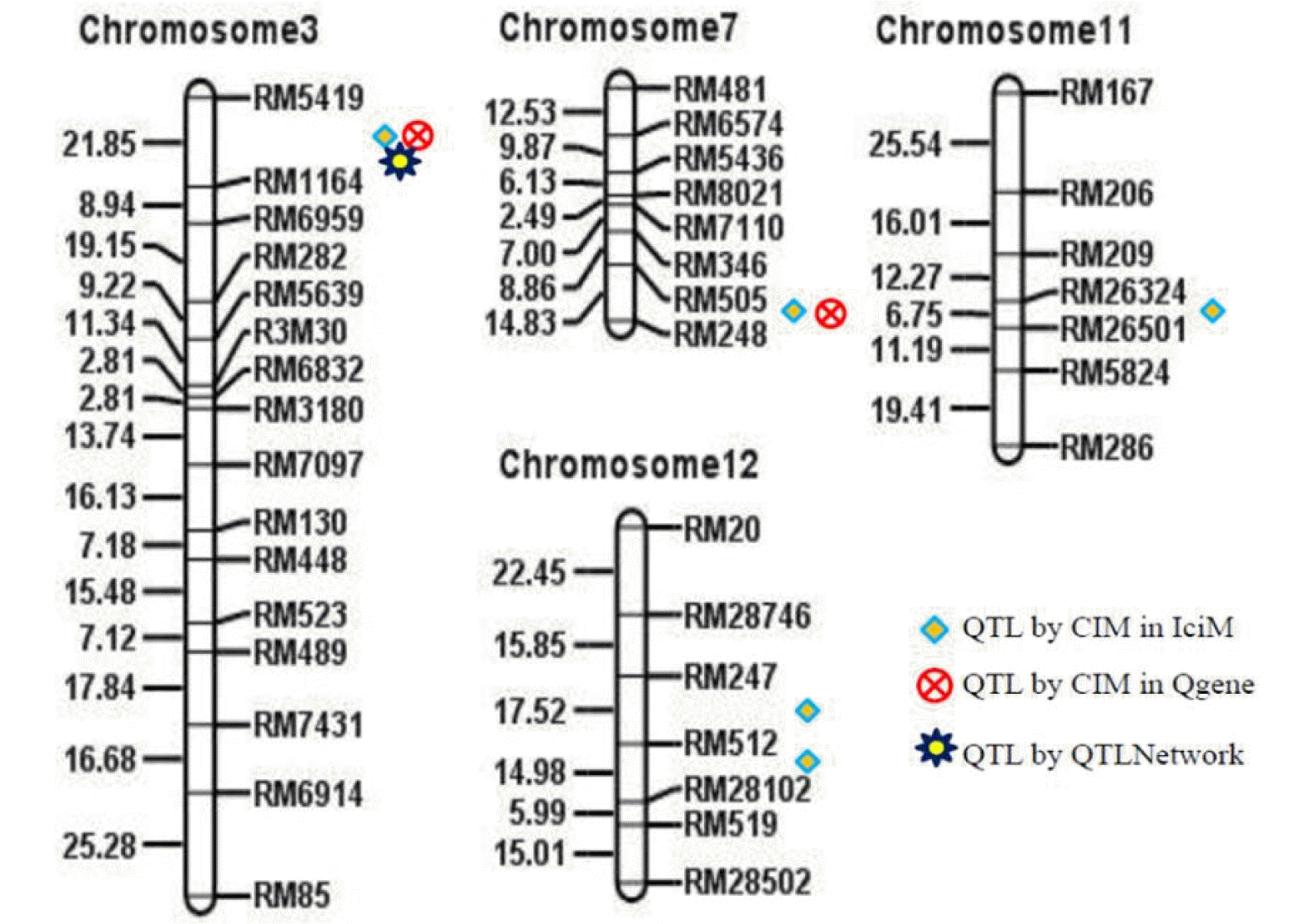
A total of 5 QTLs were detected for grain Zn content on chromosome 3, 7, 11 and 12 through Inclusive Composite Interval Mapping (ICIM) analysis using IciMapping QTL mapping tool (
Table 3,
Fig. 2) with a threshold LOD value of 3.45 at the 0.05 level of significance after permutation test with 1000 iterations. These QTLs and their confidence intervals, linked makers, additive effects, and contributions in explaining phenotypic variations are presented in
Tables 3 and
4. Among the five QTLs, one QTL was detected on each of the chromosomes 3, 7 and 11, and two QTLs on chromosome 12 (
Fig. 2).
Out of five QTLs,
qGZn3 on chromosome 3 showed the highest LOD value with the highest phenotypic contribution (
Table 3). This QTL was detected between RM5419 and RM1164 (
Table 3,
Fig. 2) with an estimated physical length of 1.83 Mb spanning from 13.03 Mb to 14.86 Mb on rice chromosome 3 (
Table 4). On the other hand,
qGZn7, which was detected between RM505 and RM248 on chromosome 7 showed the second-highest LOD and phenotypic contribution with a positive additive effect. The cumulative contribution of these two QTLs was 40.9% with individual additive effects of 4.68 and 2.06 μg/g (
Table 3). When the same set of genotyping and phenotyping data were analyzed using Qgene v4.0.2, only
qGZn3 and
qGZn7 were found significant at
P > 0.05 after the permutation test with 1000 iterations (
Table 3). In this approach, the cumulative contribution of these two QTLs was estimated as 54.3% with individual effects of 34.2% and 20.1%, respectively and their individual additive effects were 3.74 μg/g and 2.42 μg/g. The
qGZn3 was further confirmed on chromosome 3 in the same marker interval by the CIM analysis using QTLNetwork v2.0 with phenotypic contribution of 35.8%.
Epistasis for grain zinc content was analyzed using both ICiMapping and QTLNetwork. No epistatic interaction between the identified QTLs was noticed in any of the cases.
Candidate gene analysis
A total of 306 genes were detected within the 1.83 Mb length of
qGZn3 spanning from 13,025,502 bp to 14,860,443 bp on chromosome 3 between the flanking markers, RM5419 and RM1164 (
Supplementary Table S1) and annotated for gene functions using Rice Annotation Project Database (RAP-DB) genome browser. Among several functional categories, the genes belonging to metal ion transporter activity and transcription factor regulation activity were short-listed and thereby a total of 11 zinc homeostasis-related genes were found underlying the
qGZn3 (
Table 5).
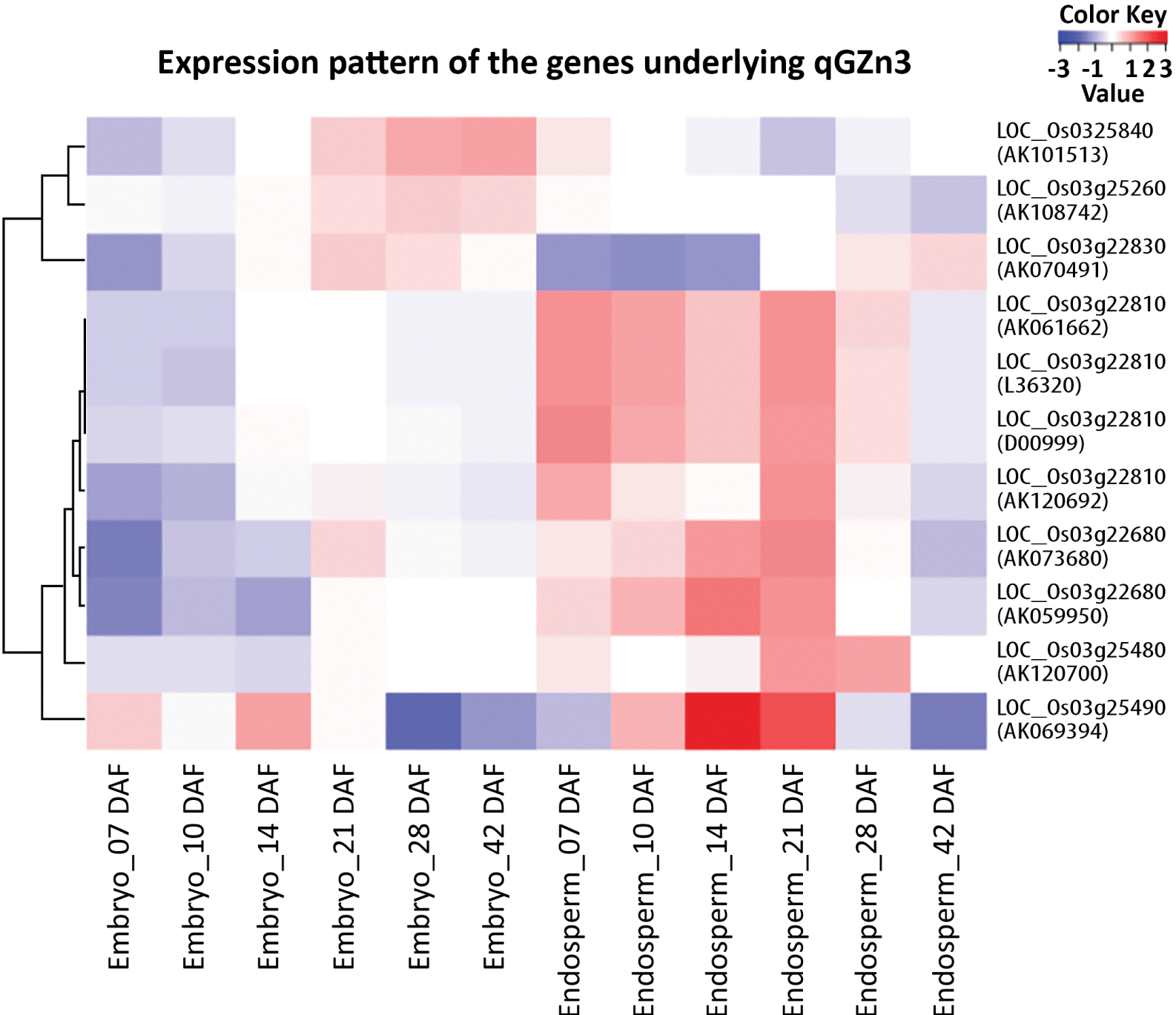
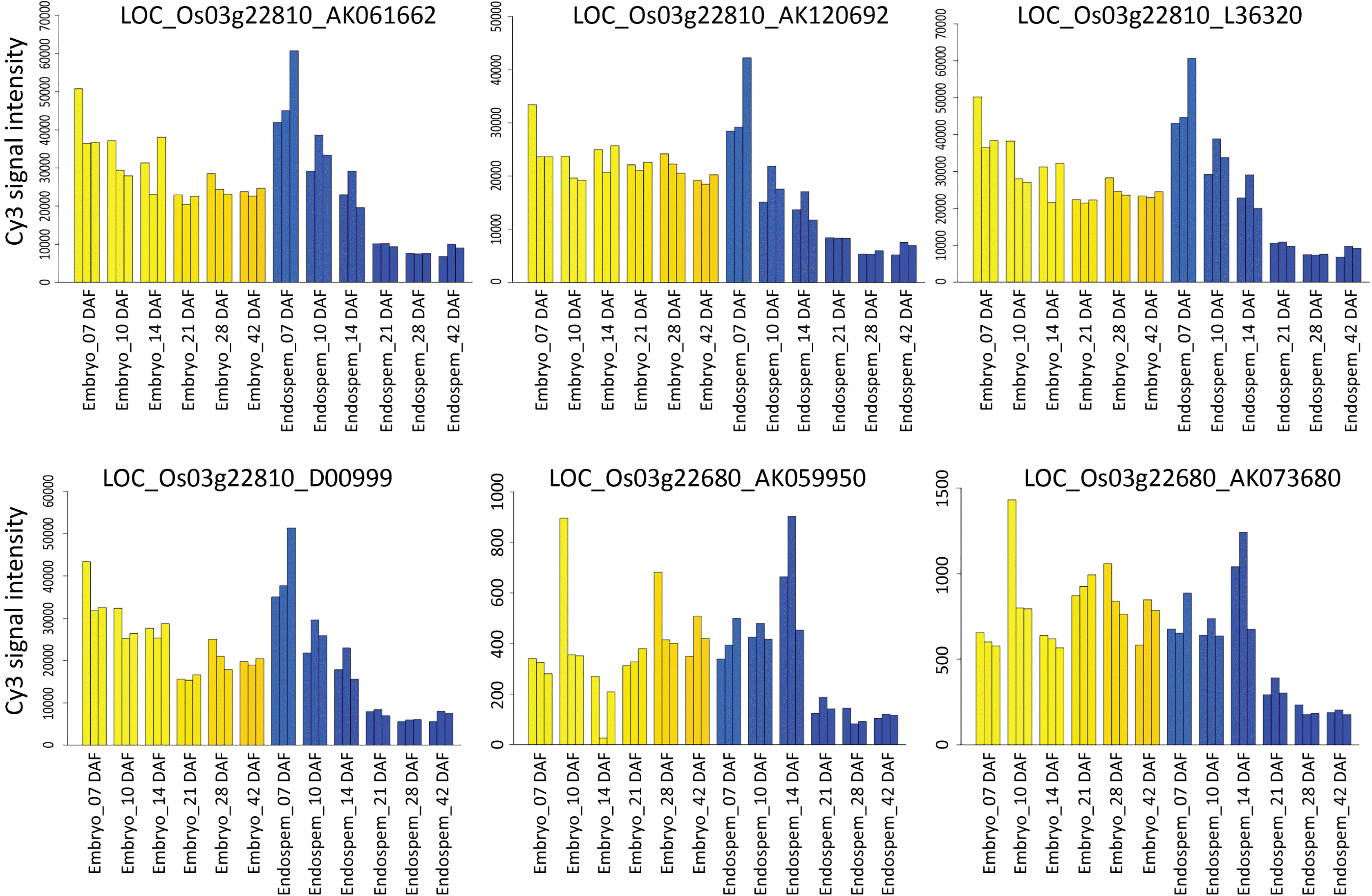
Eleven accessions of expressed sequence under six putative candidate genes out of 11 were found to match with the RiceXPro transcriptome data set for embryo and endosperm specific expression in rice (
Fig. 3). Four accessions D00999, L36320, AK061662 and AK120692 under LOC_Os03g22810 were found to show the highest Cy3 signal intensity either in the embryonic tissue or in the endosperm at all intervals of DAF starting from 7 DAF to 42 DAF among all the expressed accessions of other gene loci. It indicated that these accessions were highly expressed in both embryonic and endosperm tissues at various stages of development. However, in all these four accessions, expression levels were maximum at 7 DAF for both embryo and endosperm and then it gradually decreased in embryonic tissue, but the decreasing rate was sharp in the case endosperm tissue (
Fig. 4). On clustering of the expression profiles based on the correlation distance and complete linkage of each gene into a heat map, the accessions AK061662, L36320 and D00999 of LOC_Os03g22810 were found highly expressed in the endosperm at 7 DAF and 21 DAF, while AK120692 expressed comparatively higher in 21 days-old endosperm. On the contrary, AK069394 of LOC_Os03g25490 showed the highest expression followed by AK059950 and AK073680 of LOC_Os03g22680 in the endosperm tissue at 14 DAF (
Fig. 3), although Cy3 signal intensity for these expressed accessions was poor compared to other expressed accessions (data not shown).
DISCUSSION
Grain Zn content and yield of rice is generally negatively correlated (
Sato et al. 2013) but careful selection, particularly at early generation for grain Zn content, can identify high yielding and zinc enriched genotypes (
Swamy et al. 2016). Understanding the underlying genetic mechanism associated with higher bioavailable Zn in rice endosperm is a prerequisite for the development of high yielding zinc enriched rice. The linkage mapping for identification of genes/major effect QTLs have been shown promise in the molecular basis of increasing grain Zn content in rice (
Swamy et al. 2016;
Swamy et al. 2018). For genetic linkage mapping, the existence of significant and sharp variations between the parents and the progenies is crucial. In our present study, we observed a wide variation in Zn contents (21.9-41.3 μg/g) in unpolished rice grain of segregating F
2:3 progenies (
Table 1), which could be attributed due to the diverse combination of alleles from the two contrasting parents for Zn content (
Kumar et al. 2014). We observed that the 396 F
2:3 plants had significantly variable (
P < 0.0001) Zn contents in rice grains up to 41.3 μg/g with an average of 30.2 μg/g, which was remarkably higher compared to the superior parent (
Table 1). Our findings indicate the presence of transgressive variation among the progenies (
Fig. 1). The results of our research are in agreement with the findings of earlier investigations (
Zhang et al. 2008;
Syed et al. 2016;
Crespo-Herrera et al. 2017). However, a further precise QTL mapping and identification of gene(s) involved in the mineral uptake and transport mechanism using such a promising F
2 population are warranted.
The selective genotyping approach (
Lander and Botstein 1989) is a cost-effective genotyping technique. This technique not only saves cost by reducing the size of F
2 population into a judicious size with no significant sacrifice in phenotypic variation but it also enriches frequencies of contrasting alleles (
Collard et al. 2005), although this technique is biased towards the detection of major effect QTLs (
Navabi et al. 2009). Since our intention was also to detect main effect QTLs through a cost-effect approach, we used selective genotyping technique for this study. Approximately 12% of 396 F
2:3 progenies from each of the extreme tails of the frequency distribution were used for genotyping following the principle of selective genotyping technique (
Fig. 1B). The truncated chunk of the progenies had a contrasting amount of Zn content of 34.5 μg/g to 40.9 μg/g at the upper tail and 21.9 μg/g to 26.9 μg/gin the lower tail, which in fact, was dispersed around the parental mean values for Zn contents (
Table 1). Our findings argue the immense contributions of
Kalobokri rather than the BRRI dhan28 allele to raise the Zn content in rice grain. Another important finding of the current study is the grain Zn content in F
2 progenies is a polygenic trait. The high heritability estimate and significant Shapiro-Wilkcoefficient (at 0.0001% level of significance) clearly indicated the polygenic control of grain Zn content in the mapping population of rice derived from BRRI dhan28 ×
Kalobokri (
Fig. 1C). The polygenic control of grain Zn content of rice has been reported by several researchers (
Lu et al. 2008;
Garcia-Oliveira et al 2009;
Ishikawa et al. 2010;
Anuradha et al. 2012;
Zhang et al. 2014;
Swamy et al. 2016).
The QTLNetwork (
Yang et al. 2008), a QTL mapping tool which uses mixed linear model framework with Henderson method III (
Searle et al. 2009) is efficient to detect main effect QTLs and epistatic interactions among them. In our study, we did not find any epistatic interaction among the identified QTLs rather,
qGZn3 appeared as a main effect QTL with phenotypic contribution up to 35.8% to the total variations present in the mapping population for grain zinc content. This implies the conservation of few alleles for Zn contents in between BRRI dhan28 and
Kalobokri. The physical coordinates obtained from IRGSP map set (IRGSP and Sasaki 2005) demonstrated that the estimated physical length of
qGZn3 is 1.83 Mb, while it is 4.81 Mb for
qGZn7 (
Table 4). Moreover, we detected another three QTLs (
qGZn11,
qGZn12.1, and
qGZn12.2) within the marker intervals of RM26324 - RM26501 on chromosome 11 and RM247 - RM512 and RM512 - RM28102 on chromosome 12, although they were found not significant in CIM analysis with Qgene. The QTL,
qGZn3 showing a very high phenotypic contribution of 21.19%, 34.2%, and 35.8% in the analysis with IciMapping, Qgene, and QTLNetwork, respectively indicated to be the major effect QTL (
Table 3). Notably that the
qGZn3 was located outside the previously reported QTLs by other investigators (
Table 4) except
qGZn3.1 mapped by
Swamy et al. (2018), which spanned 16.96 Mb on chromosome 3. We mapped
qGZn3 with a relatively smaller physical length (1.83 Mb) within the
qGZn3.1. Furthermore, we summarized metal homeostasis-related genes underlying
qGZn3 for grain zinc contents using RAP-DB. A total of 11 metal homeostasis related genes were identified in the vicinity of
qGZn3 (
Table 5). Different types of zinc finger protein domains (RING, TRAF, SWIM, HIT), which show overexpression under both zinc deficiency and toxicity conditions and take part in metal ion binding homeostasis were found underlying
qGZn3. Another gene, copper/zinc superoxide dismutase (
LOC_Os03g22810), which regulates metal ion transport in plants (
van de Mortel et al 2006), was also found underlying the
qGZn3. Besides, MYB transcription factors were found to express in the vicinity of
qGZn3. MYB transcription factors show overexpression in both zinc deficiency and toxicity conditions and enhance expression of OsNAS4, which is essential for plant survival under Fe and Zn deficiency condition (
Palmer et al. 2013). Cytochrome P450 family protein (CYP) domains play an important role in electron transport or energy pathways. CYPs also show overexpression under zinc deficiency and toxicity conditions. CYP family protein domains were identified within the
qGZn3. The co-location of metal homeostasis genes with Zn QTLs provides positive evidence for the accuracy of the mapping and usefulness of these QTLs in marker-assisted breeding for biofortification of Zn in rice.
Expressed sequence tag, a short sub-sequence of cDNA sequence of a gene of interest, is used to identify the gene transcripts. EST analysis has been widely used in gene cloning and expression studies because of it high reliability (
He et al. 2012).
In-silico analysis of the putative candidate genes of
qGZn3 showed that out of 11, six candidate genes matched with the expressed sequences with varying degree of expression in the embryonic and endosperm tissues at different developmental stages of ripening rice grain. Our results show that four expressed sequences of LOC_Os03g22810, which encode Cu/Zn superoxide dismutase, a metal-binding protein, are highly active in the endosperm as well as in the embryonic tissue, which strongly suggests LOC_Os03g22810 as the potential candidate for high zinc content in rice grains. However, we need to validate this result using wet-lab based RT-PCR data before to use in marker-assisted selection for higher grain Zn rice lines.
Overall, the present study successfully identified one major effect QTLs located on chromosome 3 of rice responsible for maintaining high levels of Zn in grain and explored primary insight of linkage mapping for mineral density in the grains. Our results suggest that qGZn3 could be used as the new gene resources for improving grain Zn concentration that might be useful for marker assisted-selection breeding programs for the development of Zn-enriched rice varieties. Furthermore, we suppose our findings lay a basis for cloning of the major QTL for grain Zn that has also potential applications for rice breeding.
SUPPLEMENTARY MATERIALS
ACKNOWLEDGEMENTS
The authors are thankful to the Head, Plant Breeding Division, Bangladesh Rice Research Institute (BRRI) for donating seeds of parental materials and F2 population, and for allowing utilizing field and laboratory facilities to conduct this study. The authors express their immense sense of gratitude to BRRI authority for financial support for this study.
-
AUTHOR CONTRIBUTIONS
All authors contributed extensively to this study. M.A. Haque, M.T. Islam, M.M. Hauqe and P.S. Biswas designed and conceived the research theme. S.J. Mohiuddin performed research under direct guidance of P.S. Biswas. S.J. Mohiuddin and P.S. Biswas analyzed the data and wrote the manuscript. P.S. Biswas and M.T. Islam contributed to the paper revision.
-
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Fig. 1Frequency distribution of 396 F2:3 progenies derived from a cross BRRI dhan28 × Kalobokri showing normal distribution for Zn content in the unpolished grain. Plate A shows distribution grain zinc content of all 396 progenies. Plate B shows truncated distribution of 47 plants from the lower tail and 47 from the upper tail of the frequency distribution of 396 F2:3 progenies used in selective genotyping. The Zn contents in the upper tail ranged from 34.5 μg/g to 44.0 μg/g, while the lower tailed genotypes had Zn content ranging from 21.9 μg/g to 26.9 μg/g. Plate C represents a Q-Q plot with Shapiro-Wilk Coefficient showing normal distribution of the grain Zn content of 396 F2:3 progenies.
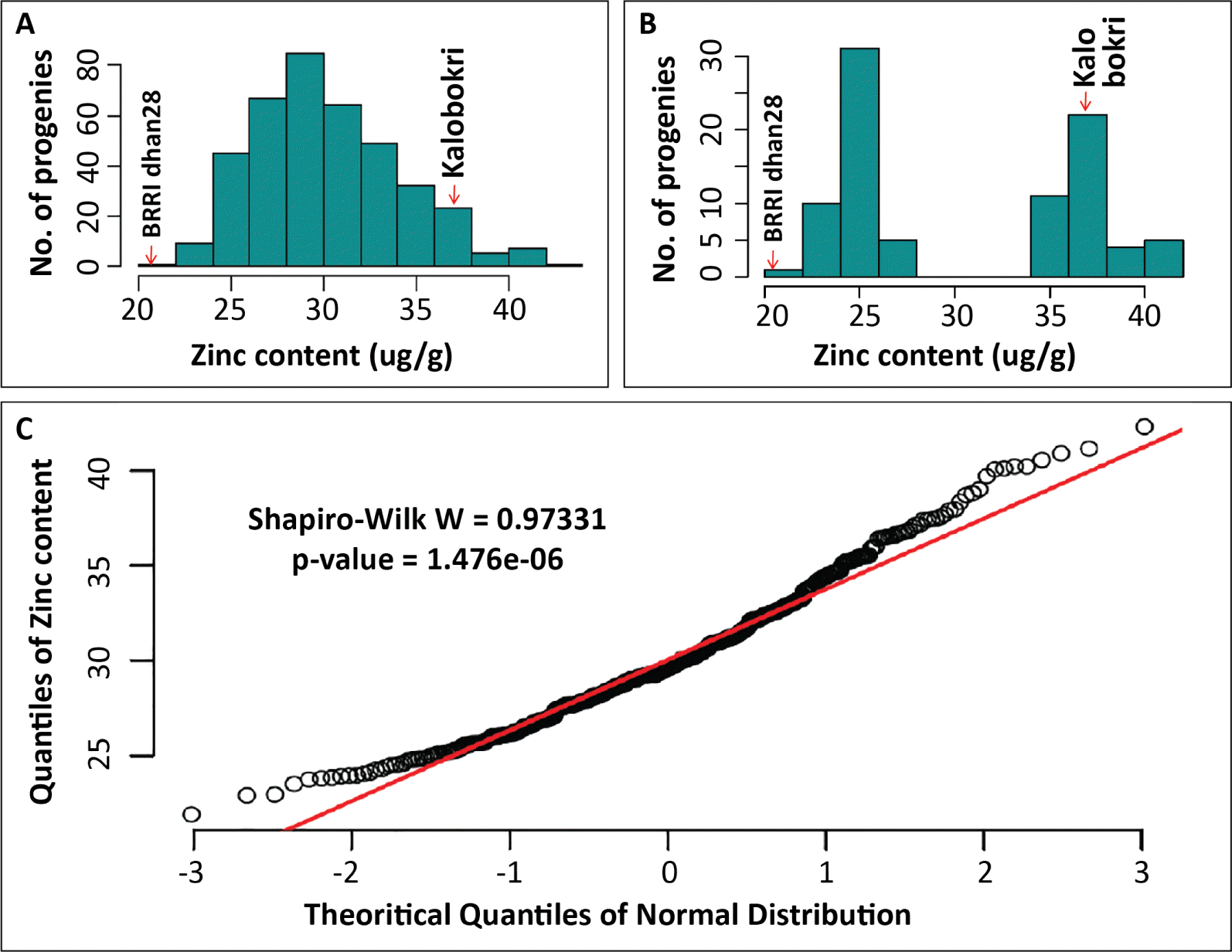
Fig. 2The QTLs identified through three different QTL mapping software for grain Zn content from BRRI dhan28 × Kalobokri F2:3 population showing overlaid locations on respective linkage groups. Five QTLs were detected on chromosome 3, 7, 11 and 12 through IciM, two QTLs on chromosome 3 and 7 through Qgene, and one QTL on chromosome 3 was detected through QTLNetwork.
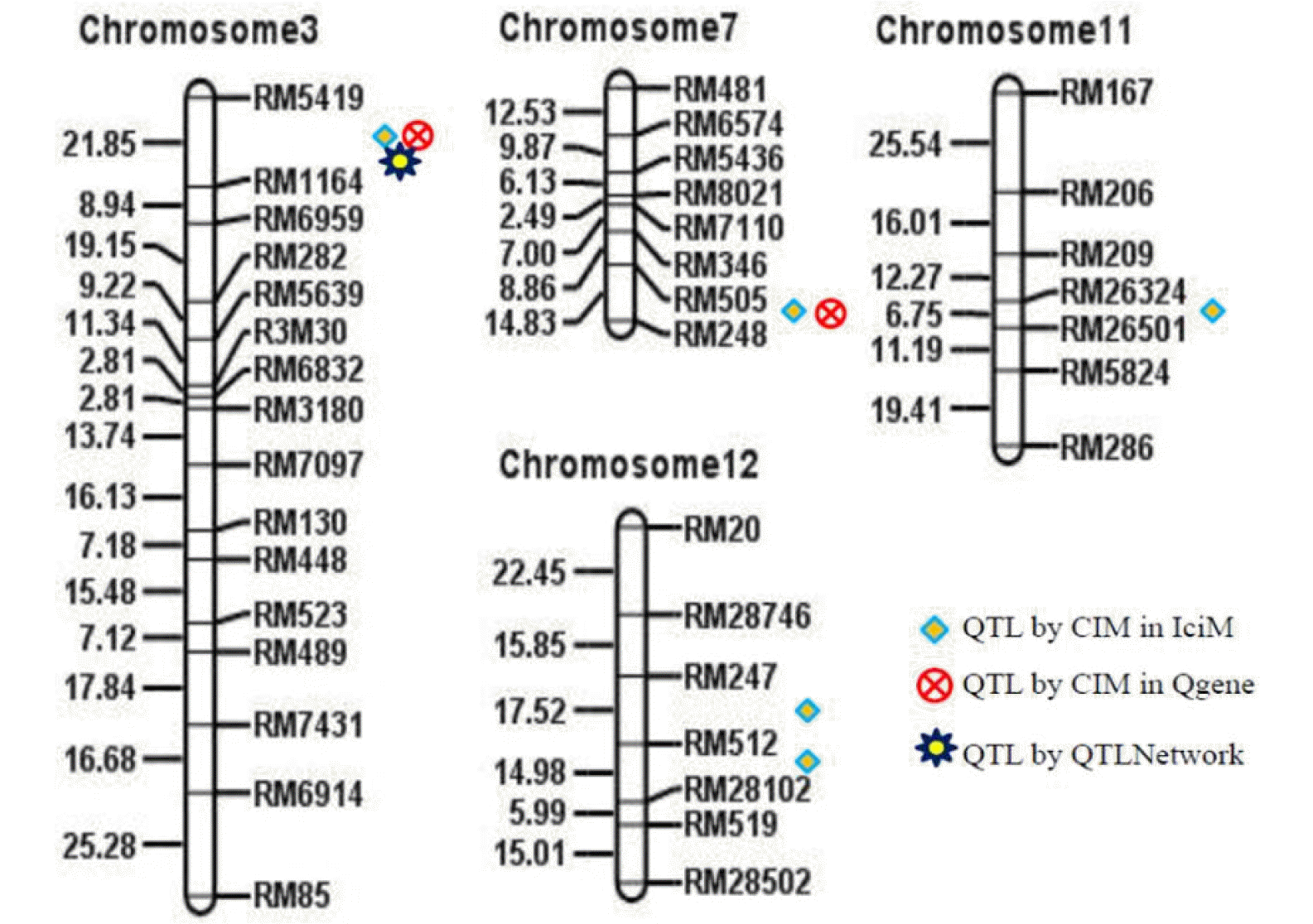
Fig. 3Expression domains of Expression Sequence Tags of six putative candidate genes underlying qGZn3 for higher grain zinc content in rice. The expressed sequences were localized through query search for putative candidate genes in RiceXPro 3.0 using RXP_0012 data set for embryo and endosperm specific expression (
https://ricexpro.dna.affrc.go.jp). A total 11expressed sequences were found to express in the embryo and endosperm tissue of rice. A heatmap was generated based clustering of correlation distance and complete linkage of each gene using inbuilt ‘gplots’ package of R with RiceXPro.
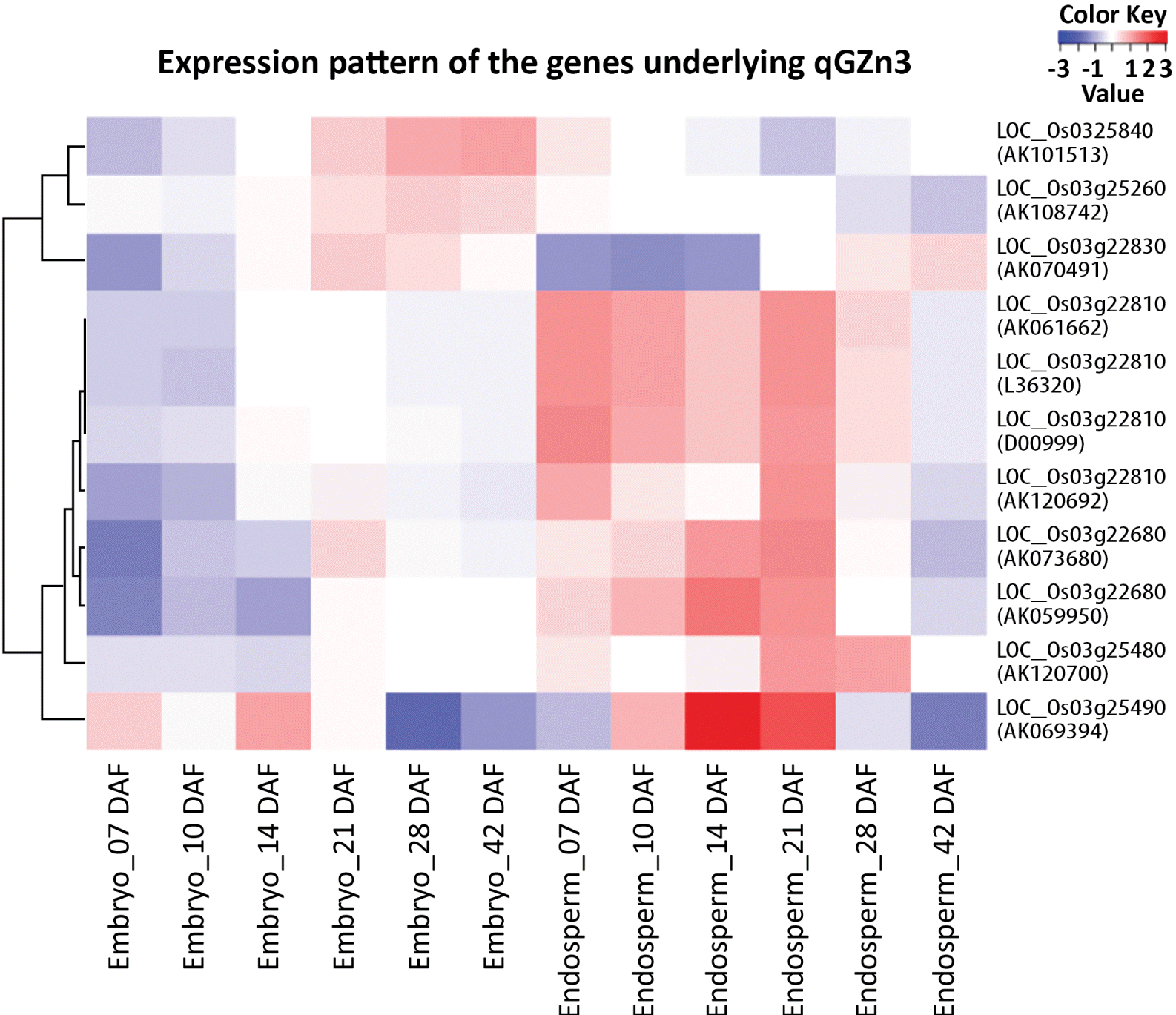
Fig. 4Expression profile of cDNA accessions underlying putative candidate genes of qGZn3 developed through query search in RiceXPro 3.0 using RXP_0012 data set for embryo and endosperm specific expression. Blue color coded bars denotes the expression profile in the endosperm tissue, while yellow denotes expression in embryonic tissue. The title of each plate denotes the gene locus with the underlying expressed cDNA accession.
Table 1Descriptive statistics of Zinc content in unpolished grain of 396 BRRI dhan28 × Kalobokri F2:3 progenies.
Table 1
|
Descriptive statistics |
Zinc content (ug/g) |
|
F2:3 families (Mean ± SD) |
30.2 ± 0.71 |
|
F2:3 families (CV) |
2.37 |
|
F2:3 families (Ranges) |
21.9-41.3 |
|
BRRI dhan28 (Low Zinc parent) |
20.2 ± 0.3 |
|
Kalobokri (High Zinc parent) |
37.5 ± 0.6** |
|
P-value |
< 0.0001 |
|
h2b (%) |
95.9 |
Table 2Results of parental survey and chromosome-wise SSR marker density in the linkage map used in genotyping of F2:3 mapping population.
Table 2
|
Chromosome |
Total number of markers used in parental survey |
No. of polymorphic markers |
% polymorphism |
No. of polymorphic marker used in genotyping |
Range of genetic distance (cM) |
|
1 |
95 |
48 |
50.5 |
14 |
7.2-17. 9 |
|
2 |
59 |
29 |
49.2 |
11 |
9.6-18.2 |
|
3 |
55 |
33 |
60.0 |
11 |
9.9-18.1 |
|
4 |
42 |
21 |
50.0 |
9 |
9.8-19.4 |
|
5 |
46 |
24 |
52.2 |
9 |
7.1-22.9 |
|
6 |
52 |
24 |
46.2 |
7 |
10.5-32.6 |
|
7 |
52 |
26 |
50.0 |
8 |
7.2-19.3 |
|
8 |
43 |
28 |
65.1 |
10 |
6.8-17.8 |
|
9 |
47 |
19 |
40.4 |
5 |
18.2-26.9 |
|
10 |
33 |
16 |
48.5 |
6 |
7.8-31.7 |
|
11 |
54 |
23 |
42.6 |
7 |
13.3-16.8 |
|
12 |
42 |
23 |
54.8 |
9 |
5.8-31.4 |
|
Total |
620 |
314 |
50.6 |
106 |
5.8-32.6 |
Table 3Linked makers and salient features of the identified QTLs from F2:3 population of BRRI dhan28 × Kalobokri for high grain Zinc content through different mapping tools.
Table 3
QTL
namez)
|
Linked marker |
CIM in IciMapping |
|
CIM in Qgene |
|
QTLNetwork |
|
Peak LODy)
|
R2 (%)x)
|
Additive effect |
Peak
LODy)
|
R2 (%)x)
|
Additive effect |
Peak
LODy)
|
R2 (%)x)
|
Additive effect |
|
qGZn3
|
RM5419 - RM1164 |
10.61 |
21.1 |
4.68 |
|
10.65 |
34.2 |
3.74 |
|
15.5 |
35.8 |
8.81 |
|
qGZn7
|
RM505 - RM248 |
9.08 |
19.8 |
2.05 |
|
4.39 |
20.1 |
2.42 |
|
- |
- |
- |
|
qGZn11
|
RM26324 - RM26501 |
4.43 |
11.5 |
‒0.54 |
|
- |
- |
- |
|
- |
- |
- |
|
qGZn12.1
|
RM247 - RM512 |
3.69 |
10.8 |
‒2.28 |
|
- |
- |
- |
|
- |
- |
- |
|
qGZn12.2
|
RM512 - RM28102 |
3.45 |
10.6 |
‒2.72 |
|
- |
- |
- |
|
- |
- |
- |
Table 4The map location and physical length of the reference QTLs at the vicinity of the newly detected from F2:3 population of BRRI dhan28 × Kalobokri for grain zinc content.
Table 4
|
QTL name |
Chromosome |
Interval between
linked markers (bp) |
Estimated
length (Mb) |
Reference QTL at the vicinity |
|
QTL Name |
Flanking markers |
Interval between linked markers (bp) |
Authors |
|
qGZn3
|
3 |
13,025,502-14,860,443 |
1.83 |
qZn3.1
|
RM517 - RM16 |
6,165,992-23,126,231 |
, Swamy et al. (2018)
|
|
qZn3.2
|
RM55 - RM520 |
29,052,318-30,912,804 |
, Swamy et al. (2018)
|
|
qZn3.1
|
RM7 - RM517 |
9,828,364-6,165,992 |
, Anuradha et al. (2012)
|
|
qZn3
|
RM3562 |
11,683,187 |
, Nawaz et al (2015)
|
|
|
|
|
qZn3.1
|
ad03013905 - ad03014175 |
25,900,000-27,100,00 |
, Lee et al (2020)
|
|
qGZn7
|
7 |
24,526,356-29,339,845 |
4.81 |
qZn7
|
RM234 - R1789 |
25,472,688-26,527,102 |
, Lu et al (2008)
|
|
qZn7.1
|
RM234 - RM248 |
25,472,688-29,339,845 |
, Anuradha et al (2012)
|
|
qZn7.2
|
RM248 - RM8007 |
29,339,845-7,710,329 |
, Anuradha et al. (2012)
|
|
qZn7.3
|
RM501 - OsZip2 |
6,755,439-17,010,137 |
, Anuradha et al. (2012)
|
|
qZn7
|
RM248 |
29,339,845 |
, Zhang et al (2014)
|
|
qZn7
|
R1440 |
16,872,010 |
, Norton et al (2010)
|
|
|
|
|
qZn7
|
RM10 - RM1973 |
22,848,239-20,164,121 |
, Jeong et al (2020)
|
|
qGZn11
|
11 |
7,407,655-11,103,286 |
3.7 |
qZn11
|
C794 - RG118 |
3,033,385-4,413,928 |
, Lu et al (2008)
|
|
|
|
|
qZn11.1
|
RM1812 - RM332 |
2,405,106-2,840,211 |
, Jeong et al (2020)
|
|
|
|
|
qZn11.2
|
RM332 - RM552 |
2,840,211-5,104,502 |
, Jeong et al (2020)
|
|
|
|
|
qZn11.3
|
RM229 - RM21 |
18,407,879-19,639,406 |
, Jeong et al (2020)
|
|
qGZn12.1
|
12 |
3,185,384-5,104,270 |
1.9 |
qZn12.1
|
RM235 - RM17 |
26,107,709-26,954,657 |
, Stangoulis et al (2007)
|
|
qGZn12.2
|
12 |
5,104,270-15,907,723 |
10.8 |
qZn12.1
|
RM17 - RM260 |
26,954,657-1,083,931 |
, Anuradha et al. (2012)
|
|
|
|
|
qZn12.2
|
RM260 - RM7102 |
10833,931-13,258,404 |
, Anuradha et al. (2012)
|
|
|
|
|
qZn12
|
R1709 - C1069 |
23,517,144-25,047,073 |
, Ishikawa et al (2010)
|
|
|
|
|
qZn12.1
|
RM3331 |
23,460,827 |
, Garcia-Oliviera et al (2009)
|
|
|
|
|
qZn12.1
|
RM415 - RM19 |
425,723-2,432,426 |
, Swamy et al. (2018)
|
|
|
|
|
qZn12
|
RM8216 - RM247 |
14,49,696-31,85,384 |
, Jeong et al (2020)
|
|
|
|
|
qZn12.1
|
CrP4887439-12172332 |
4,880,000-5,370,000 |
, Calayugan
|
Table 5Chromosomal position, putative function, and annotation of Zinc homeostasis related genes underlying qGZn3.
Table 5
|
Locus ID |
Map location (bp) |
Putative function |
GO annotation |
|
LOC_Os03g22680
|
Chr3:13098285 - 13095798 |
RING finger and CHY zinc finger domain-containing protein 1, putative, expressed |
Protein metabolism |
|
LOC_Os03g22810
|
Chr3:13179170 - 13184591 |
copper/zinc superoxide dismutase, putative, expressed |
Metal ion transport |
|
LOC_Os03g22830
|
Chr3:13201307 - 13198991 |
zinc finger, C3HC4 type domain containing protein, expressed |
Zinc ion binding |
|
LOC_Os03g24184
|
Chr3:13748160 - 13743553 |
TRAF-type zinc finger domain-containing protein 1, putative, expressed |
Zinc ion binding |
|
LOC_Os03g24500
|
Chr3:13966686 - 13967778 |
zinc finger, RING-type, putative, expressed |
Zinc ion binding |
|
LOC_Os03g24970
|
Chr3:14254884 - 14253793 |
SWIM zinc finger family protein, putative |
Zinc ion binding |
|
LOC_Os03g25260
|
Chr3:14424629 - 14427869 |
HIT zinc finger domain containing protein, expressed |
|
|
LOC_Os03g25304
|
Chr3:14447137 - 14447670 |
myb-like DNA-binding domain containing protein, expressed |
Transcription |
|
LOC_Os03g25480
|
Chr3:14553102 - 14556229 |
cytochrome P450, putative, expressed |
Electron transport |
|
LOC_Os03g25490
|
Chr3:14559322 - 14561845 |
cytochrome P450 72A1, putative, expressed |
Electron transport |
|
LOC_Os03g25550
|
Chr3:14599572 - 14598168 |
myb-like DNA-binding domain containing protein, putative, expressed |
Transcription |
References
- Akter S, Basher SA. 2014. The impacts of food price and income shocks on household food security and economic well-being: evidence from rural Bangladesh. Glob. Environ. Change. 25: 150-162.
- Anuradha K, Agarwal S, Rao YV Rao KV, Viraktamath BC, Sarla N. 2012. Mapping QTLs and candidate genes for iron and zinc concentrations in unpolished rice of Madhukar × Swarna RILs. Gene. 508: 233-240.
- Calayugan MI, Formantes AK, Amparado A, Descalsota-Empleo GI, Nha CT, Inabangan-Asilo MA, et al. 2020. Genetic analysis of agronomic traits and grain iron and zinc concentrations in double haploid population of rice (Oryza sativa L). Sci. Rep.. 10: 1-14.
- Cakmak I, Kutman UB. 2018. Agronomic biofortifcation of cereals with zinc: a review. Eur. J. Soil Sci.. 69: 172-180.
- Collard BC, Jahufer MZZ, Brouwer JB, Pang ECK. 2005. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: the basic concepts. Euphytica. 142: 169-196.
- Crespo-Herrera LA, Govindan V, Stangoulis J, Hao Y, Singh RP. 2017. QTL Mapping of grain Zn and Fe concen-trations in two hexaploid wheat RIL populations with ample transgressive segregation. Front. Plant Sci.. 8: 1800
- Dixit S, Singh UM, Abbai R, Ram T, Singh VK, Paul A, et al. 2019. Identification of genomic region(s) responsible for high iron and zinc content in rice. Sci. Rep.. 9: 1-8.
- Garcia-Oliveira AL, Tan L, Fu Y, Sun C. 2009. Genetic identification of quantitative trait loci for contents of mineral nutrients in rice grain. J. Integr. Plant Biol.. 51: 84-92.
- Graham R, Senadhira D, Beebe S, Iglesias C, Monasterio I. 1999. Breeding for micronutrient density in edible portions of staple food crops: conventional approaches. Field Crops Res.. 60: 57-80.
- Gregorio GB, Senadhira D, Graham RD, Htut T. 1999. Improving iron and zinc value of rice for human nutrition. Agric. & Develop.. 23: 77-81.
- Gross BL, Zhao Z. 2014. Archaeological and genetic insights into the origins of domesticated rice. Proc. Natl. Acad. Sci. U.S.A.. 111: 6190-6197.
- HarvestPlus2016. Reaching millions: annual report 2015 HavestPlus, IFPRI. Washington D.C., U.S.A..
- He WY, Rao ZC, Zhou DH, Zheng SC, Xu WH, Feng QL. 2012. Analysis of expressed sequence tags and characterization of a novel gene, Slmg7, in the midgut of the common cutworm, Spodoptera litura. PLoS One. 7: e33621
- Hossain MA, Islam SA, Zaman MK. Saleque MA., Kashem MA., Ali MA., Kabir SK., 2014. Screening of BRRI germplasm for high zinc and iron enriched cultivar. editors. Bangladesh Rice Research Abstracts. Bangladesh Rice Research Institute. Gazipur, Bangladesh: pp. 47.
- Impa SM, Johnson-Beebout SE. 2012. Mitigating zinc deficiency and achieving high grain Zn in rice through integration of soil chemistry and plant physiology research. Plant Soil. 361: 3-41.
- Sasaki T. IRGSP (International Rice Genome Sequencing Project)2005. The map-based sequence of the rice genome. Nature. 436: 793-800.
- Ishikawa S, Abe T, Kuramata M, Yamaguchi M, Ando T, Yamamoto T, et al. 2010. A quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on short arm of chromosome 7. J. Exp. Bot.. 61: 923-934.
- Jeong OY, Lee JH, Jeong EG, Chun A, Bombay M, Banzon Ancheta M, et al. 2020. Analysis of QTL responsible for grain iron and zinc content in doubled haploid lines of rice (Oryza sativa L.) derived from an intra-japonica cross. Plant Breed.. 139: 344-355.
- Karthika KS, Rashmi I, Parvathi MS. Hasanuzzaman M., Fujita M., Oku H., Nahar K., Hawrylak-Nowak B., 2018. Biological functions, uptake and transport of essential nutrients in relation to plant growth. editors. Biological functions, uptake and transport of essential nutrients in relation to plant growth, p. 1-49. Plant nutrients and abiotic stress tolerance. Springer. Singapore:
- Kennedy G, Burlingame B, Nguyen VN. 2002. Nutritional contribution of rice and impact of biotechnology and biodiversity in rice-consuming countries. In: 20th International Rice Commission Bangkok Thailand; 23-26 July, 2002.
- Kumar J, Jain S, Jain RK. 2014. Linkage mapping for grain iron and zinc content in F2 population derived from the cross between PAU201 and Palman 579 in rice (Oryza sativa L.). Cereal Res. Commun.. 42: 389-400.
- Lander ES, Botstein D. 1989. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics. 121: 185-199.
- Lee SM, Kang JW, Lee JY, Seo J, Shin D, Cho JH, et al. 2020. QTL analysis for Fe and Zn concentrations in rice grains using a double haploid population derived from a cross between rice (Oryza sativa) cultivar 93-11 and Milyang 352. Plant Breed. Biotech.. 8: 69-76.
- Lu K, Li L, Zheng X, Zhang Z, Mou T, Hu Z. 2008. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains. J. Genet.. 87: 305-310.
- McCouch SR. CGSNL (Committee on Gene Symbolization, Nomenclature and Linkage, Rice Genetics Cooperative)2008. Gene nomenclature system for rice. Rice (N. Y.). 1: 72-84.
- McCouch SR, Chen X, Panaud O, Temnykh S, Xu Y, Cho YG, et al. Sasaki T., Moore G., 1997. Microsatellite marker development, mapping and applications in rice genetics and breeding. editors. Oryza: From molecule to plant. Springer. Dordrecht, Netherlands: pp. 88-99.
- Myers SS, Wessells KR, Kloog I, Zanobetti A, Schwartz J. 2015. Effect of increased concentrations of atmospheric carbon dioxide on the global threat of zinc deficiency: a modelling study. Lancet Glob. Health. 3: e639-e645.
- Nakandalage N, Nicolas M, Norton RM, Hirotsu N, Milham PJ, Seneweera S. 2016. Improving rice zinc biofortification success rates through genetic and crop management approaches in a changing environment. Front. Plant Sci.. 7: 764
- Navabi A, Mather DE, Bernier J, Spaner DM, Atlin GN. 2009. QTL detection with bidirectional and unidirectional selective genotyping: marker-based and trait-based analyses. Theor. Appl. Genet.. 118: 347-358.
- Nawaz Z, Kakar KU, Li X, Zhang B, Shou H, Shu Q. 2015. Genome wide association mapping of QTLs for contents of eight elements in brown rice (Oryza sativa L). J. Agric. Food Chem.. 63: 8008-8016.
- Nelson JC. 1997. QGENE: software for marker-based genomic analysis and breeding. Mol. Breed.. 3: 239-245.
- Norton GJ, Deacon CM, Xiong L, Huang S, Meharg AA, Price AH. 2010. Genetic mapping of the rice ionome in leaves and grain: identification of QTLs for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil. 329: 139-153.
- Palmer CM, Hindt MN, Schmidt H, Clemens S, Guerinot ML. 2013. MYB10 and MYB72 are required for growth under Fe-limiting conditions. PLoS Genet.. 9: e1003953
- R Development Core Team.2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria..
- Sato Y, Takehisa H, Kamatsuki K, Minami H, Namiki N, Ikawa H, et al. 2013. RiceXPro version 3.0: expanding the informatics resource for rice transcriptome. Nucleic Acids Res.. 41: D1206-D1213.
- Searle SR, Casella G, McCulloch CE. 2009. Variance components. John Wiley & Sons. New York, U.S.A..
- Shahzad Z, Rouached H, Rakha A. 2014. Combating mineral malnutrition through iron and zinc biofortification of cereals. Compr. Rev. Food Sci. Food Saf.. 13: 329-346.
- Sharma A, Patni B, Shankhdhar D, Shankhdhar SC. 2013. Zinc-an indispensable micronutrient. Physiol. Mol. Biol. Plants. 19: 11-20.
- Stangoulis JC, Huynh BL, Welch RM, Choi EY, Graham RD. 2007. Quantitative trait loci for phytate in rice grain and their relationship with grain micronutrient content. Euphytica. 154: 289-294.
- Swamy BPM, Kaladhar K, Anuradha K, Batchu AK, Longvah T, Sarla N. 2018. QTL analysis for grain zinc concentrations in two O. nivara derived backcross populations. Rice Sci.. 25: 197-207.
- Swamy BPM, Rahman MA, Inanbagan-Asio MA, Amparado A, Manito C, Chadha-Mohanty P, et al. 2016. Advances in breeding for high grain zinc in rice. Rice (N. Y.). 9: 49
- Syed MA, Iftekharuddaula KM, Mian MAK, Rasul MG, Rahmam GKMM, Panaullah GM, et al. 2016. Main effect QTLs associated with arsenic phyto-toxicity tolerance at seedling stage in rice (Oryza sativa L.). Euphytica. 209: 805-814.
- van de Mortel JE, Villanueva LA, Schat H, Kwekkeboom J, Coughlan S, Moerland PD, et al. 2006. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol.. 142: 1127-1147.
- Virk PS, Ford-Lloyd VB, Jackson MT, Newbury HJ. 1995. Use of RAPD for the study of diversity within plant germplasm collections. Heredity. 74: 170-179.
- Yang J, Hu C, Hu H, Yu R, Xia Z, Ye X, et al. 2008. QTLNetwork: mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics. 24: 721-723.
- Zhang J, Wu LH, Wang MY. 2008. Iron and zinc biofortification in polished rice and accumulation in rice plant (Oryza sativa L.) as affected by nitrogen fertilization. Acta Agric. Scand. B. Soil Plant Sci.. 58: 267-272.
- Zhang X, Zhang G, Guo L, Wang H, Zeng D, Dong G, et al. 2011. Identification of quantitative trait loci for Cd and Zn concentrations of brown rice grown in Cd-polluted soils. Euphytica. 180: 173-179.
- Zhang M, Pinson SRM, Tarpley L, Huang XY, Lahner B, Yakubova E, et al. 2014. Mapping and validation of quantitative trait loci associated with concentration of 16 elements in unmilled rice grain. Theor. Appl. Genet.. 127: 137-165.