Abstract
Chloroplasts are essential plant organelles that play important roles in photosynthesis and are involved in many fundamental metabolic pathways. Plastid ribosomal proteins are essential components of protein synthesis machinery and have diverse roles in plant growth and development during chloroplast differentiation. In this study, we isolated and characterized T-DNA-tagged rice mutant (prpl13), which exhibited albino seedling lethality. PRPL13 is a nuclear gene encoding the 50S ribosomal protein L13, which is localized in chloroplasts. Transmission electronic microscopy analysis showed abnormal plastid development in the thylakoids of the chloroplasts of the prpl13 mutant seedlings. Chlorophyll and carotenoid contents were also significantly reduced in the leaves of the mutants. Quantitative reverse transcription-PCR analysis revealed that the prpl13 mutations altered the expression levels of genes involved in photosynthesis and chloroplast development. Thus, our data indicate that nuclear-encoded PRPL13 plays an important role in chloroplast development in rice.
-
Key words:
Oryza sativa
,
PRPL13
, WLP1, Albino, Chloroplast development, Rice
INTRODUCTION
Chloroplasts (plastids) are DNA-containing organelles that evolved from a photosynthetically active cyanobacterium (
Gray 1993). Plastid-encoded genes are precisely transcribed by two RNA polymerases, the plastid-encoded RNA polymerase (PEP) and the nucleus-encoded RNA polymerase (NEP) (
Hess and Borner 1999;
Shiina et al. 2005). During chloroplast development, NEP and PRP direct the transcription of housekeeping genes and chloroplast genes, respectively, that encode proteins involved in photosynthesis (
Hajdukiewicz et al. 1997).
Ribosomes are essential for protein synthesis in the cytoplasm, plastids, and mitochondria. Plastid ribosomal proteins (PRPs) play important roles in the build-up step of chloroplast differentiation. Plastid proteins are translated by bacterial-type 70S ribosomes comprising one small (30S) and one large (50S) ribosomal subunit (Schippers and Mueller-Roeber 2010;
Tiller and Bock 2014;
Bieri et al. 2017). The 30S small subunit comprises 16S ribosomal RNA (rRNA) and 25 proteins (Yamaguchi
et al. 2000), while the 50S large subunit is made up of 23S rRNA and 33 proteins, 25 of which are nuclear-encoded (
Yamaguchi and Subramanian 2003). Although much is known about the composition of chloroplast ribosomes, only a few PRP mutants have been functionally analyzed.
In Arabidopsis, many plastid ribosomal proteins (
RPS5,
9,
13, and
20 and
RPL1,
4,
6,
10,
13,
18,
21,
27,
31, and
35) were reported to be essential for embryogenesis (
Pesaresi et al. 2001; Morita-Yamamuro
et al. 2004;
Hsu et al. 2010;
Bryant et al. 2011;
Romani et al. 2012;
Tiller et al. 2012;
Yin et al. 2012;
Zhang et al. 2016). The maize
high chlorophyll fluorescent 60 (
hcf60) mutant showed an unstable pale green seedling-lethal phenotype, and
HCF60 encodes chloroplast ribosomal small subunit protein 17 (
Schultes et al. 2000). Also, the maize
lethal embryo 1 (
lem1) mutant encoding plastid ribosomal protein S9 induces early embryo lethality (
Ma and Dooner 2004).
The rice PRP mutants
albino seedling lethality 1 (
asl1) and
asl2 exhibit albino and seedling-lethal phenotypes (
Gong et al. 2013;
Lin et al. 2015;
Zhao et al. 2016).
ASL1 encodes the plastid 30S ribosomal protein S20 (PRPS20), and
ASL2 encodes the chloroplast 50S ribosome protein L21 (PRPL21). The rice
white leaf and panicles 1 (
wlp1) mutant exhibits an albino phenotype at low temperatures (
Song et al. 2014).
WLP1 encodes the plastid 50S ribosomal protein L13. The
thermo-sensitive chlorophyll-deficient mutant 11 (
tcd11) mutant also exhibits albino phenotypes at low temperatures (
Wang et al. 2017).
TCD11 encodes the plastid ribosomal protein S6. The rice
albino lethal 1 (
al1) mutant exhibits an albino lethal phenotype at the seedling stage (
Zhao et al. 2016).
AL1 encodes the PRPL12 protein. Recently, the rice
white green leaf 2 (
wgl2) mutant was characterized. It displayed an albino phenotype at the three-leaf stage that gradually transitioned to green during subsequent developmental stages, and
WGL2 was shown to encode the plastid ribosomal protein S9 (
Qiu et al. 2018).
In this study, we isolated rice T-DNA mutants (prpl13) with an albino lethal phenotype at the seedling stage. The transcription levels of genes associated with chloroplast development and photosynthesis were severely altered and the chloroplast ultrastructure was defective in the mutants. Thus, we concluded that PRPL13 plays a crucial role in chloroplast development.
MATERIALS AND METHODS
Plant growth
The mutant lines (
prpl13-1 and
prpl13-2) used in this study were isolated from a T-DNA insertional mutant population (
Jeon et al. 2000;
Jeong et al. 2002). Surface-sterilized T2 seeds of the mutant and wild-type (WT)
Oryza sativa cv. Dongjin plants were germinated in half strength (1/2) Murashige and Skoog (MS) medium comprising 3% sucrose and 0.4% phytagel. The plants were grown in a growth chamber, and genotyping was performed using specific PCR primers, which are listed in
Table 1.
To investigate the subcellular localization of PRPL13, the full-length ORF was amplified without a stop codon (PCR primers in
Table 1) and introduced into the vector pGA3651 (
pUbi::GFP) (
Kim et al. 2009). The GFP empty vector (pGA3651) and
pUbi::PRPL13-GFP vector were transfected into the Arabidopsis protoplast cells using the PEG method (
Yoo et al. 2007). After 12 hours of incubation, the transformed protoplasts were observed in the GFP and RFP channel (chlorophyll autofluorescence) with a Nikon Eclipse Ti2 inverted fluorescent microscope (Nikon, JAPAN).
Total RNA was isolated from different organs including 10-day-old roots, shoots, mature flag leaves, and immature panicles using QIAzol lysis reagent. cDNA from the different organs was synthesized using 2 mg of total RNA following the procedure described in the manufacturer’s manual (Qiagen, Solgent, Korea). The synthesized cDNA was used as a template for reverse transcription PCR (RT-PCR) and quantitative RT-PCR (qRT-PCR). qRT-PCR was performed with the CFX connect Real-time System (BIO-RAD) using the TOYOBO THUNDERBIRD SYBR qPCR Mix (TOYOBO, JAPAN). Rice
actin2 and
UBQ5 were used as internal controls for RT-PCR and qRT-PCR, respectively. All of the primers used are listed in
Table 1. The data are expressed as the mean ± SD of three biological replicates. Relative expression levels were calculated by 2
‒ΔΔCT (
Livak and Schmittgen 2001).
The total chlorophyll and carotenoid contents of the 10-day-old
prpl13-1 and WT plants were determined according to a previously described method (
Lichtenthaler 1987).
A BLASTP search of the PRPL13 sequences using the National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/) website was conducted for the phylogenetic analysis. The ClustalW program (DDBJ, https://www.ddbj.nig.ac.jp/index-e.html) was used for the full-length sequence alignment, and a neighbor-joining tree was constructed using the MEGA version 7.0 software with the Poisson-correction method (
Kumar et al. 2016).
The leaf samples were fixed in 2.5% glutaraldehyde in 0.02 M phosphate buffer, pH 7.2, for 2 hours at 4℃. After several washes with phosphate buffer, the samples were post-fixed in 1% osmium tetroxide overnight at 4℃ and again washed with phosphate buffer. The samples were dehydrated in a graded ethanol series. The ethanol was then substituted with propylene oxide, and the samples were then embedded in Epon 812 for 48 hours at 60℃. Sections (100 nm) were cut on a Leica Ultracut UCT ultramicrotome using a diamond knife (Diatome). Ultrathin sections were transferred onto 200 mesh copper grids and stained with 2% uranyl acetate for 20 minutes and lead citrate (
Reynolds 1963) for an additional 10 minutes. The sections were examined with a TEM (Hitachi, H 7650) at 80 kV.
RESULTS
Isolation of albino mutants in the rice T-DNA-tagged pool
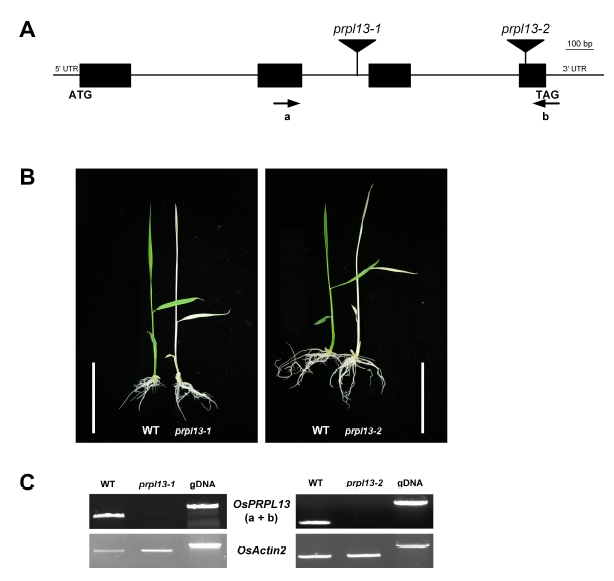
We isolated two independent T-DNA-tagged mutants exhibiting the albino phenotype at the seedling stage (
Jeon et al. 2000;
Jeong et al. 2002). Analyses of the flanking sequences determined by TAIL-PCR in the albino mutant lines demonstrated that T-DNA insertions were present in the second intron and fourth exon of
PRPL13 (
LOC_ Os01g54540) (http://signal.salk.edu/cgi-bin/RiceGE; PFG_ 3A-00683 and PFG_1C-10407) (
Fig. 1A). The
PRPL13 gene encodes the 50S plastid ribosomal protein L13 consisting of 233 amino acids. Genotyping analysis showed that homozygous plants displaying the albino phenotype cosegregated with each T-DNA insertion, indicating that these mutations were recessive (
Fig. 1B). RT-PCR analyses revealed that full-length transcripts were not found in the
prpl13-1 and
prpl13-2 mutants demonstrating that they were mRNA null mutants (
Fig. 1C). In the rice genome, five
PRPL13-like genes were present (
PRPL13L1 to
PRPL13L5) (
Fig. 2A). PRPL13 is closed to the PRPL13L1 protein in the phylogenetic tree, however, pairwise sequence alignment indicated that the PRPL13 protein had 30% similarity to the PRPL13L1 protein, implying that the
PRPL13 gene plays an important role in chloroplast development as a single genomic copy (
Fig. 2B).
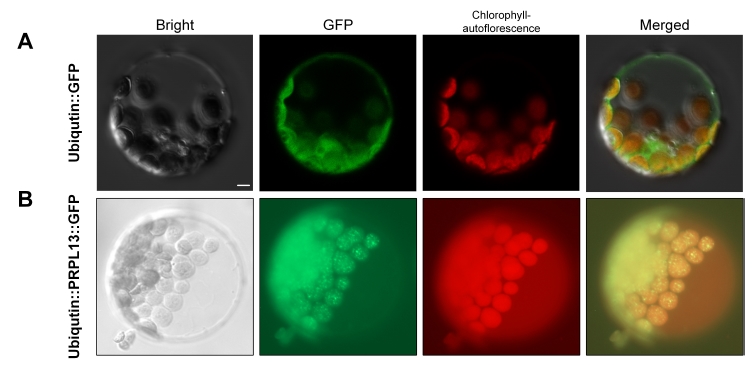
To investigate the actual subcellular localization of the PRPL13 proteins, transient expression of
pUbi::GFP (pGA3651) and
pUbi:PRPL13:GFP was performed in Arabidopsis protoplasts using the PEG method. The
pUbi::GFP vector was used as a negative control in which GFP signals were found in the cytosol and nucleus (
Fig. 3A). In contrast, the PRPL13-GFP fusion signals colocalized with the chlorophyll autofluorescence in the chloroplasts (
Fig. 3B). This result strongly suggested that the RPRL13 protein is localized in the chloroplast.
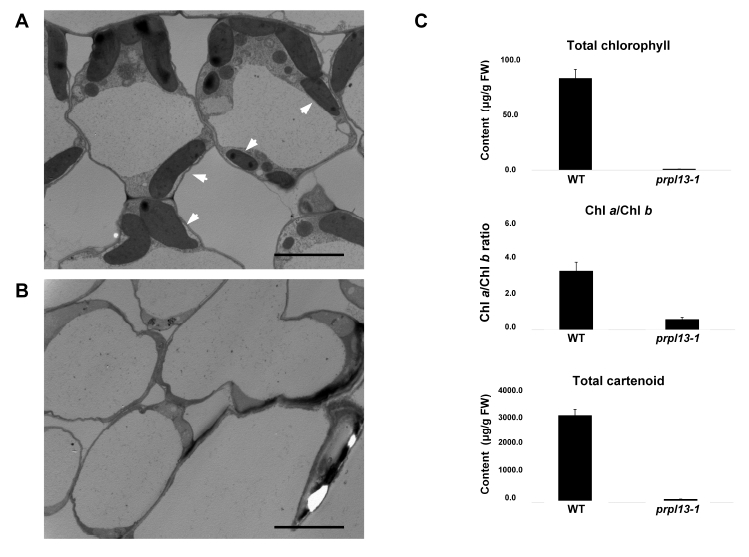
We used a TEM to investigate the morphology of the plastids in the leaves of the10-day-old albino and WT seedlings. The TEM revealed that chloroplast development was severely impaired in the mutant (
Fig. 4A, 4B). We also measured the chlorophyll and carotenoid contents of the
prpl13-1 and WT plants. The albino
prpl13-1 mutants exhibited lower chlorophyll and carotenoid contents, which was likely due to damaged chloroplast development (
Fig. 4C). These observations imply that the
PRPL13 gene plays a critical role in chloroplast development during early development.
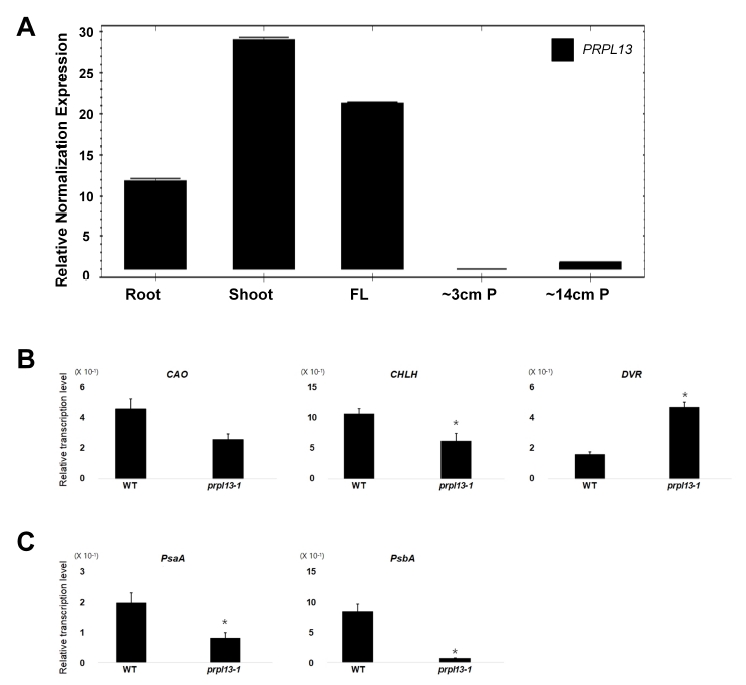
We examined the gene expression of
PRPL13 in the WT plants using qRT-PCR. Because plastids are also present in the roots and chloroplasts,
PRPL13 transcripts were detected in the 10-day-old young roots, young shoots, mature flag leaves, and immature panicles (∼3 and ∼14 cm panicles) at the flowering stage.
PRPL13 was highly expressed in the green leaf tissues such as the young shoots and flag leaves (
Fig. 5A).
We examined the expression levels of genes associated with chlorophyll biosynthesis and photosynthesis in the
prpl13 and WT plants. We examined the expression of the
chlorophyll a oxygenase1 (
CAO1),
Mg chelatase H subunit (
CHLH), and
divinyl reductase (
DVR) genes as they are chlorophyll biosynthesis-associated genes (
Wu et al. 2007). The expression levels of the
CAO1 and
CHLH genes were downregulated in the
prpl13 mutants. However, the transcript levels of
DVR increased in the
prpl13 mutants (
Fig. 5B). We also examined the expression levels of the photosynthesis-related transcripts of the plastid genes
photosystem I P700 chlorophyll a apoprotein A1 (
psaA) and
photosystem II protein D1 (
psbA), which encode components of photosystems I and II. Both genes,
psaA and
psbA, were suppressed in the
prpl13 mutant plants and not in the WT plants (
Fig. 5C).
DISCUSSION
Chloroplasts are important cellular organelles that conduct photosynthesis. Chloroplast development is regulated by the coordinated expression of both nuclear and plastid genes (
Kusumi and Iba 2014). PRPs are important components for chloroplast biogenesis, plastidial protein biosynthesis, and early chloroplast development (
Lin et al. 2015). Many PRP mutants have been identified in higher plants, and some PRPs have been shown to act in diverse biological processes in Arabidopsis (
Romani et al. 2012). More than 70 rice mutants exhibiting albino, chlorine, stripe, virescent, yellow-green, and zebra leaves have been reported (
Jung et al. 2003;
Kurata et al. 2005). However, few studies have reported the functions of PRPs in rice.
The
ASL1 gene encodes PRPS20, whose mutation shows an albino lethal phenotype at the seedling stage and alters the expression of plastid and nuclear genes associated with chloroplast development (
Gong et al. 2013). Mutation of the rice
ASL2 gene encoding PRPL21 causes defective chloroplast development and seedling death (
Lin et al. 2015). Additionally,
AL1 encodes
PRPL12, which causes defective plastid ribosomes and early seedling lethality (
Zhao et al. 2016). The rice
tcd11 mutant displays an albino phenotype at low temperatures, similar to the
wlp1 mutant.
TCD encodes
PRPS6, and
WLP1 encodes
PRPL13 (
Song et al. 2014;
Wang et al. 2017).
WGL2 encodes plastid ribosomal protein S9, and the mutant shows an albino phenotype at the three-leaf stage that gradually transitions to green during later developmental stages (
Qiu et al. 2018).
In this study, we isolated and characterized T-DNA insertion mutants exhibiting an albino seedling-lethal phenotype with disrupted
PRPL13 genes. The
prpl13 mutants exhibited altered pigment contents and defective chloroplast development. The
prpl13-1 and
prpl13-2 mutants showed a loss-of-function phenotype, and they were identified as new alleles of
wlp1, which exhibits the albino and seedling-lethal phenotypes (
Song et al. 2014). In contrast, the
wlp1 mutant displays the albino phenotype at low temperatures (23℃) and is not lethal. It even exhibits the same phenotype as that of the WT plants at 30℃. Also, the
wlp1 mutant containing a single-base substitution is not lethal, but it has a cold-sensitive phenotype, suggesting that the mutant WLP1 protein is partially functional (
Song et al. 2014). Interestingly,
prpl13-1 and
prpl13-2 showed albino and seedling-lethal phenotypes under normal growth conditions, indicating that they are stronger
wlp1 alleles and
PRPL13 loss-of-function causes abnormal development of the chloroplasts. The qRT-PCR results demonstrated a significant decrease in the expression of genes involved in chlorophyll biosynthesis (
CAO1 and
CHLH) and photosynthesis (
psaA,
psbA) in the mutants. Many mitochondria and chloroplast proteins are now encoded in the nucleus, which means the communication between nucleus and chloroplast (anterograde and retrograde signal) requires coordinated expression to produce organelle proteins and to maintain organelle function (
Hess et al. 1993, 1994;
Pesaresi et al. 2007;
Woodson and Chory 2008). However, the transcript of
DVR, nuclear-encoded gene, was upregulated in the
prpl13 mutant suggesting that a retrograde signaling may exist. Also, lower total chlorophyll contents and carotenoid contents were observed in the
prpl13 mutants. Taken together, PRPL13 plays a critical role in chloroplast development, as a key component of chloroplast ribosomal proteins.
ACKNOWLEDGEMENTS
The authors thank Junhyun Lim and Sang Dae Yun for taking care of the Arabidopsis plants used in the subcellular localization experiments. This work was supported by a grant from the Next-Generation BioGreen 21 program (Project No. PJ01369001), Rural Development Administration, Republic of Korea.
Fig. 1Phenotype of prpl13 mutant. (A) Genomic structure of PRPL13 and T-DNA insertion position. The black boxes represent exons, the intervening lines represent introns, and a and b primers were used for genotyping and RT-PCR analysis. (B) Phenotypes of the 7-day-old prpl13-1 and prpl13-2 plants. The albino phenotypes cosegregated with the T-DNA insertion. Scale bar = 2 cm. (C) RT-PCR analysis of the PRPL13 transcripts in the WT, prpl13-1, and prpl13-2 mutants. OsActin2 was used as a control.
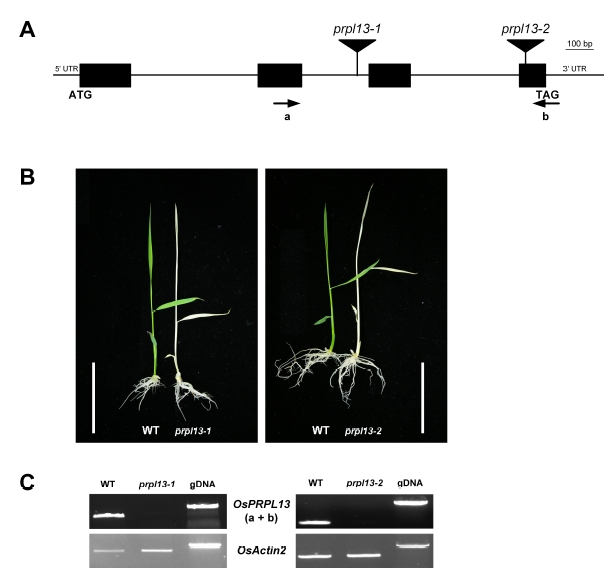
Fig. 2Multiple sequence alignment and phylogenetic tree of PRPL13 family. (A) Phylogenetic tree constructed using the neighbor-joining method with the MEGA 7 program (Kumar et al. 2016). Bootstrap values indicate the number of amino acid substitutions per site. (B) Sequence alignment of the rice PRPL13 and PRPL13L1 proteins. Black shading = identical residues and gray shading = similar residues. The sequence was aligned with OsPRPL13 (Os01g54540) and PRPL13L1 (Os03g54890). The numbers on the left indicate the residue positions in the proteins.

Fig. 3Subcellular localization of PRPL13 protein. (A) Transient expression of pUbi::GFP in the Arabidopsis protoplast. (B) Transient expression of pUbi::PRPL13::GFP in the Arabidopsis protoplast. The pUbi::GFP vector was used as a control. Scale bar = 5 µm.
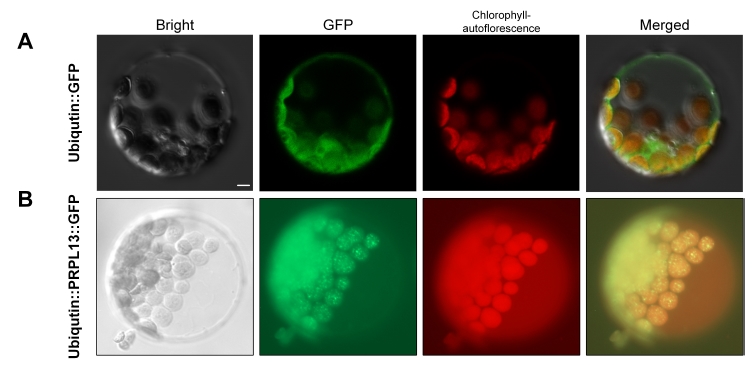
Fig. 4TEM analysis and chlorophyll contents in the WT and albino mutant. (A) Section of WT chloroplasts. (B) Section of prpl13-1. The arrowheadsindicate the chloroplasts. Scale bar = 5 µm. (C) The pigment contents of the WT and prpl13-1 mutant plants. Total chlorophyll and carotenoid concentrations were obtained from 10-day-old plants. All values are the means of three biological repeats. The error bars indicate standard deviation (SD).
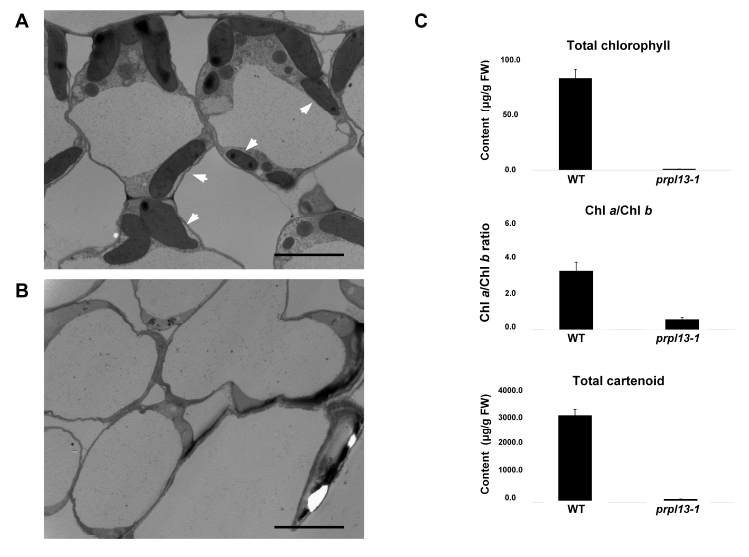
Fig. 5Expression pattern analysis of PRPL13 and of chlorophyll biosynthesis-and photosynthesis-related genes. (A) Relative expression levels of PRPL13 in various organs. RNA was extracted from 10-day-old roots, shoots, mature flag leaves (FL), young panicles of ∼3 cm (∼3 cm P), and young panicles of ∼14 cm (∼14 cm P). The error bars indicate standard deviation (SD). (B and C) Relative expression levels of chlorophyll biosynthesis-related genes and photosynthesis-related genes in the WT and prpl13-1 mutant plants. These experiments were repeated more than twice with similar results. Levels of significant difference are indicated by *(P < 0.05).
Table 1List of primers used for qRT-PCR, DNA constructs and genotyping in this study.
Table 1
|
Name |
Primer sequence (5ʹ→3ʹ) |
Remarks |
|
Primers used for qRT-PCR |
|
RPL13-qRT_F1
|
CTAAGGGCAGACTGGGAAGA |
|
RPL13-qRT_R1
|
TTGATAGGCAGTGGAACAGG |
|
CHLH-F1
|
TGACTCAGACCCGACAAAGC |
|
CHLH-R1
|
TCCCCTCGTACCACTTAGGG |
|
DVR-F1
|
CAGGTCGAGACCGTCAAGAAC |
|
DVR-R1
|
ATGACCTGGATCGGCACCTTG |
|
CAO1-F1
|
GACACCTTCATCTGGGCTTCAA |
|
CAO1-R1
|
CGAGAGACATCCGGTAGAGC |
|
PsaA-F1
|
GTTTTCGCGGAGGGCTAGAT |
|
PsaA-R1
|
TGACCTGCGATCAGGAAAAGA |
|
PsbA-F1
|
ACTAGCACCGAAAACCGTCTTT |
|
PsbA-R1
|
CAGCGATGAAGGCGATAATAAA |
|
UBQ5-F1
|
CTCGCCGACTACAACATCCA |
|
UBQ5-R1
|
TCTTGGGCTTGGTGTACGTCTT |
|
Primers used for genotyping |
|
3A-00683-F1 |
GTAGGCTTGCATCCACCATT |
|
3A-00683-R1 |
CCGTTTCGGCATTCAATTAG |
|
1C-10407-F1 |
CTAATTGAATGCCGAAACGG |
|
1C-10407-R1 |
ACACTCCATTCCCCAAACAG |
|
RB
|
TTGGGGTTTCTACAGGACGTAAC |
T-DNA right border |
|
Primers used for RT-PCR |
|
OsActin2-F1
|
AGGCTCCTCTCAACCCCAAGGCCAATCG |
|
OsActin2-R1
|
AGGTAATCAGTGAGATCACGCCCAGC |
|
PRPL13-RT-F1
|
GTAGGCTTGCATCCACCATT |
|
PRPL13-RT-R1
|
ACACTCCATTCCCCAAACAG |
|
Primers used to generate DNA constructs |
|
PRPL13_FL-F
|
GGATCCATGGCTACGGCCATCGCA |
Full length cDNA |
|
PRPL13_FL-R
|
ACTAGTCTTCTCAGACTTCTGTATTCTTTTATCC |
References
Reference
- Bieri P, Leibundgut M, Saurer M, Boehringer D, Ban N. 2017. The complete structure of the chloroplast 70S ribosome in complex with translation factor pY. EMBO J.. 36: 475-486.
- Bryant N, Lloyd J, Sweeney C, Myouga F, Meinke D. 2011. Identification of nuclear genes encoding chloroplast-localized proteins required for embryo development in Arabidopsis. Plant Physiol.. 155: 1678-1689.
- Gong X, Jiang Q, Xu J, Zhang J, Teng S, Lin D, et al. 2013. Disruption of the rice plastid ribosomal protein s20 leads to chloroplast developmental defects and seedling lethality. G3 (Bethesda). 3: 1769-1777.
- Gray MW. 1993. Origin and evolution of organelle genomes. Curr. Opin. Genet. Dev.. 3: 884-890.
- Hajdukiewicz PTJ, Allison LA, Maliga P. 1997. The two RNA polymerases encoded by the nuclear and the plastid compartments transcribe distinct groups of genes in tobacco plastids. EMBO J.. 16: 4041-4048.
- Hess WR, Börner T. 1999. Organellar RNA polymerases of higher plants. Int. Rev. Cytol.. 190: 1-59.
- Hess WR, Prombona A, Fieder B, Subramanian AR, Borner T. 1993. Chloroplast rps15 and the rpoB/C1/C2 gene cluster are strongly transcribed in ribosome-deficient plastids: evidence for a functioning non-chloroplast-encoded RNA polymerase. EMBO J.. 12: 563-571.
- Hess WR, Muller A, Nagy F, Borner T. 1994. Ribosome-deficient plastids affect transcription of light-induced nuclear genes: genetic evidence for a plastid-derived signal. Mol. Gen. Genet.. 242: 305-312.
- Hsu SC, Belmonte MF, Harada JJ, Inoue K. 2010. Indispensable roles of plastids in Arabidopsis thaliana embryogenesis. Curr. Genomics. 11: 338-349.
- Jeon JS, Lee S, Jung KH, Jun SH, Jeong DH, Lee J, et al. 2000. T-DNA insertional mutagenesis for functional genomics in rice. Plant J.. 22: 561-570.
- Jeong DH, An S, Kang HG, Moon S, Han JJ, Park S, et al. 2002. T-DNA insertional mutagenesis for activation tagging in rice. Plant Physiol.. 130: 1636-1644.
- Jung KH, Hur J, Ryu CH, Choi Y, Chung YY, Miyao A, et al. 2003. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol.. 44: 463-472.
- Kim SR, Lee DY, Yang JI, Moon S, An G. 2009. Cloning vectors for rice. J. Plant Biol.. 52: 73-78.
- Kumar S, Stecher G, Tamura K. 2016. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol.. 33: 1870-1874.
- Kurata N, Miyoshi K, Nonomura K, Yamazaki Y, Ito Y. 2005. Rice mutants and genes related to organ development, morphogenesis and physiological traits. Plant Cell Physiol.. 46: 48-62.
- Kusumi K, Iba K. 2014. Establishment of the chloroplast genetic system in rice during early leaf development and at low temperatures. Front. Plant Sci.. 5: 386
- Lichtenthaler HK. 1987. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol.. 148: 350-382.
- Lin D, Jiang Q, Zheng K, Chen S, Zhou H, Gong X, et al. 2015. Mutation of the rice ASL2 gene encoding plastid ribosomal protein L21 causes chloroplast developmental defects and seedling death. Plant Biol. (Stuttg). 17: 599-607.
- Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods. 25: 402-408.
- Ma ZR, Dooner HK. 2004. A mutation in the nuclear-encoded plastid ribosomal protein S9 leads to early embryo lethality in maize. Plant J.. 37: 92-103.
- Morita-Yamamuro C, Tsutsui T, Tanaka A, Yamaguchi J. 2004. Knock-out of the plastid ribosomal protein S21 causes impaired photosynthesis and sugar-response during germination and seedling development in Arabidopsis thaliana
. Plant Cell Physiol.. 45: 781-788.
- Pesaresi P, Varotto C, Meurer J, Jahns P, Salamini F, Leister D. 2001. Knock-out of the plastid ribosomal protein L11 in Arabidopsis: effects on mRNA translation and photosynthesis. Plant J.. 27: 179-189.
- Pesaresi P, Schneider A, Kleine T, Leister D. 2007. Interorganellar communication. Curr. Opin. Plant Biol.. 10: 600-606.
- Qiu Z, Chen D, He L, Zhang S, Yang Z, Zhang Y, et al. 2018. The rice white green leaf 2 gene causes defects in chloroplast development and affects the plastid ribosomal protein S9. Rice (N. Y.). 11: 39
- Reynolds ES. 1963. The use of lead citrate at high pH as an electron-opaque stain for electron microscopy. J. Cell Biol.. 17: 208-212.
- Romani I, Tadini L, Rossi F, Masiero S, Pribil M, Jahns P, et al. 2012. Versatile roles of Arabidopsis plastid ribosomal proteins in plant growth and development. Plant J.. 72: 922-934.
- Schippers JH, Mueller-Roeber B. 2010. Ribosomal composition and control of leaf development. Plant Sci.. 179: 307-315.
- Schultes NP, Sawers RJH, Brutnell TP, Krueger RW. 2000. Maize high chlorophyll fluorescent 60 mutation is caused by an Ac disruption of the gene encoding the chloroplast ribosomal small subunit protein 17. Plant J.. 21: 317-327.
- Shiina T, Tsunoyama Y, Nakahira Y, Khan MS. 2005. Plastid RNA polymerases, promoters, and transcription regulators in higher plants. Int. Rev. Cytol.. 244: 1-68.
- Song J, Wei X, Shao G, Sheng Z, Chen D, Liu C, et al. 2014. The rice nuclear gene WLP1 encoding a chloroplast ribosome L13 protein is needed for chloroplast development in rice grown under low temperature conditions. Plant Mol. Biol.. 84: 301-314.
- Tiller N, Bock R. 2014. The translational apparatus of plastids and its role in plant development. Mol. Plant. 7: 1105-1120.
- Tiller N, Weingartner M, Thiele W, Maximova E, Schottler MA, Bock R. 2012. The plastid-specific ribosomal proteins of Arabidopsis thaliana can be divided into non-essential proteins and genuine ribosomal proteins. Plant J.. 69: 302-316.
- Wang WJ, Zheng KL, Gong XD, Xu JL, Huang JR, Lin DZ, et al. 2017. The rice TCD11 encoding plastid ribosomal protein S6 is essential for chloroplast development at low temperature. Plant Sci.. 259: 1-11.
- Woodson JD, Chory J. 2008. Coordination of gene expression between organellar and nuclear genomes. Nat. Rev. Genet.. 9: 383-395.
- Wu Z, Zhang X, He B, Diao L, Sheng S, Wang J, et al. 2007. A chlorophyll-deficient rice mutant with impaired chlorophyllide sterification in chlorophyll biosynthesis. Plant Physiol.. 145: 29-40.
- Yamaguchi K, Subramanian AR. 2003. Proteomic identification of all plastid specific ribosomal proteins in higher plant chloroplast 30S ribosomal subunit -PSRP-2 (U1A-type domains), PSRP-3 alpha/beta (ycf65 homologue) and PSRP-4 (thx homologue). Eur. J. Biochem.. 270: 190-205.
- Yin T, Pan G, Liu H, Wu J, Li Y, Zhao Z, et al. 2012. The chloroplast ribosomal protein L21 gene is essential for plastid development and embryogenesis in Arabidopsis. Planta. 235: 907-921.
- Yoo SD, Cho YH, Sheen J. 2007. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc.. 2: 1565-1572.
- Zhang J, Yuan H, Yang Y, Fish T, Lyi SM, Thannhauser TW, et al. 2016. Plastid ribosomal protein S5 is involved in photosynthesis, plant development, and cold stress tolerance in Arabidopsis. J. Exp. Bot.. 67: 2731-2744.
- Zhao DS, Zhang CQ, Li QF, Yang QQ, Gu MH, Liu QQ. 2016. A residue substitution in the plastid ribosomal protein L12/AL1 produces defective plastid ribosome and causes early seedling lethality in rice. Plant Mol. Biol.. 91: 161-177.