Abstract
Buckwheat sprouts are used as a functional food in several countries owing to their soft and slightly crispy texture and attractive fragrance. They possess excellent anti-oxidant activity and are rich in amino acids, minerals, and flavonoids; however, sprout growth and flavonoid content may vary among buckwheat germplasms. The present study was performed to evaluate 87 common and 13 tartary buckwheat germplasms for sprout growth traits and rutin and quercetin content. We found that the common type buckwheat sprouts had considerably higher leaf length and width, fresh shoot weight, and extract weight than those of tartary buckwheat sprouts, but the rutin and quercetin content was significantly higher in tartary buckwheat sprouts. The rutin content ranged from 323.7 to 750.6 mg/ 100 g DW in common buckwheat sprouts and 2220 to 3185 mg/100 g DW in tartary buckwheat sprouts. Similarly, the quercetin content ranged from 0.27 to 1.82 and 4.76 to 30.41 mg/100 g DW in common and tartary buckwheat sprouts, respectively. Additionally, there was no significant correlation between the growth traits and rutin content in common buckwheat sprouts, whereas, the leaf length of tartary buckwheat sprouts showed a significant positive correlation with the rutin content. This information can be used by plant breeders and plant geneticists to study the genetic and genomic approaches for buckwheat breeding program.
-
Key words: Germplasm, Common buckwheat, Tartary buckwheat, Sprout, Rutin, Quercetin
INTRODUCTION
Buckwheat (
Fagopyrum species) is a short seasonal and one of the most important traditional and functional crops grown and consumed worldwide. Although its name includes the word “wheat,” this crop does not have any taxonomic relationship with common wheat and is considered a pseudo-cereal (
Joshi and Rana 1995). Buckwheat is a dicotyledonous plant belonging to the family Polygonaceae (
Chauhan et al. 2010;
Ahmed et al. 2013;
Koyama et al. 2013). The genus has more than 20 species, of which just two cultivated species, namely, common buckwheat (
Fagopyrum esculentum Moench, ES) and tartary buckwheat (
Fagopyrum tataricum Gaertner, TA), are of dietary importance for human consumption (
Jiang et al. 2007;
Nam et al. 2015;
Chen et al. 2018). Common buckwheat is usually consumed in Asia, Europe, North America, South Africa, and Australia, and it has the advantages of sweet taste and large seed size (
Li and Zhang 2001;
Ratan and Kothiyal 2011). Tartary buckwheat is rarely consumed owing to its bitter taste (
Liu et al. 2008) and small seed size (
Bonafaccia et al. 2003); however, it possesses several nutraceutical compounds and therefore tartary buckwheat is widely grown and utilized in western China (
Zielińska et al. 2012).
Buckwheat is a gluten-free grain crop. It has abundant levels of amino acids, proteins, lipids, saccharides, fiber, minerals, vitamins, and flavonoids (
Huang et al. 2014;
Nam et al. 2015). Recently, flavonoids have received considerable attention because of their anti-oxidant, anti-tumor, and anti-inflammatory effects. Flavonoids are the most common and widely distributed group of plant phenolic compounds but cannot be synthesized by humans and animals (
Cook and Samman 1996).
Hertog et al. (1995),
Chao et al. (2002), and
Li et al. (2016), based on their epidemiological studies, have suggested a protective role of buckwheat seed flavonoids in preventing diabetic, cancer and heart diseases. The content and composition of flavonoids in common buckwheat and tartary buckwheat seeds vary widely. The tartary buckwheat seeds contain higher flavonoid content than that of common buckwheat seeds (
Li and Zhang 2001). Moreover, the sprouts of buckwheat contain a high amount of rutin in the stem and leaf than that in the buckwheat seeds (
Wiczkowski et al. 2014).
Seed germination is a complex biological process in which the seed absorbs water and starts to elongate its embryonic axis (
Welbaum et al. 1998). In this process, several dormant enzymes are activated to initiate various biochemical processes that change the biochemical composition of germinating seeds (
Nonogaki 2006). The germinated seeds require high amount of energy for their growth and development, resulting in elevated levels of polyphenols (
Koyama et al. 2013), anti-oxidants (
Brajdes and Vizireanu 2012), and nutritional components and decreased levels of anti-nutrients compared with those in the non-germinated seeds. Anti-nutrients including protease inhibitors, lipase inhibitors, amylase inhibitors, phytic acid, oxalic acid, and oxalates are natural or synthetic compounds that interfere the absorption of amino acids and essential nutrients (
Akande et al. 2010). Sprouts are small seedlings before the development of true leaves and roots. The use of sprouts has spread globally owing to their high nutritional value (
Wiczkowski et al. 2014) and easy accessibility in grocery stores as a fresh vegetable (
Koyama et al. 2013). The most prevalent sprouts are those of soybean, mung bean, and buckwheat seeds (
Liu et al. 2008).
Buckwheat sprouts have excellent anti-oxidant activity and are rich in amino acids, minerals, and flavonoids (
Kim et al. 2004). They have soft and slightly crispy texture and attractive fragrance and are consumed as a fresh vegetable in several countries (
Yiming et al. 2015). Tartary buckwheat sprouts contain higher rutin content (0.8%-1.7%) than that of common buckwheat sprouts, with 0.4%-0.5% rutin content. However, tartary buckwheat is less famous than common buckwheat because of its bitter taste. Despite the high nutritional value of buckwheat, there has been only limited research on the association between morphological diversity and flavonoid content of sprouts. There is a need to explore the diversity in phenolic content in buckwheat sprouts. The present study was conducted to evaluate 87 accessions of common type and 13 tartary type buckwheat germplasms for sprout growth traits and rutin and quercetin content.
MATERIALS AND METHODS
Plant materials and growing conditions
One hundred buckwheat germplasms collected from the National Genetic Resource Center (the RDA gene bank), Rural Development Administration (RDA) Jeonju, the Republic of Korea, were used. The germplasm collection included 87 common buckwheat and 13 tartary buckwheat accessions (
Table 1). The germplasm was sown at 25℃ ± 1℃ in a greenhouse in the National Institute of Agricultural Sciences (NAS), RDA by placing 100 seeds of each accession between two layers of wet filter papers in Petri plates.
After 10 days of sowing, the buckwheat germplasms were evaluated based on sprout traits such as leaf length, leaf width, fresh shoot weight, and extract weight. For leaf length and width measurement, 10 plants were selected and data were recorded in triplicates. For fresh shoot weight and extract, all sprouts were harvested and used to record the data.
Estimation of rutin and quercetin content
The rutin and quercetin content in buckwheat sprouts was measured using an HPLC system according to the method described by
Kim et al. (2008). One hundred milligrams of dried plant sample was homogenized in 1 mL of methanol (MeOH) containing 10% phosphoric acid. The sample was vortexed for 5 minutes and incubated at 37℃ for 3 hours, followed by vigorous shaking after each hour. The mixture was centrifuged at 12,000 × g for 10 minutes and filtered through a 0.45-mm disposable syringe (Advantec, Tokyo, Japan). The filtered sample was used to determine the flavonoid content using an HPLC system (Agilent Technologies 1200 series, California, USA). The flavonoids were separated using the Capcell PAK C18 column 5 mm, 4.6 mm × 250 mm, Shiseido, Tokyo, Japan. The injection volume of the sample was 10 mL and the absorbance of the sample was examined at 340 nm. The column oven temperate was adjusted to 40℃. The mobile phase, which consisted of solvent A (MeOH:water:acetic acid [5:92.5:2.5, (v/v/v)]) and solvent B (MeOH:water: acetic acid [92.5:2.5:2.5, (v/v/v)]), was delivered at a rate of 1.0 mL/minutes. Initially, the mobile phase was started with 10% solvent B (0 minutes), followed by a linear gradient with 36% solvent B for 27 minutes, 60% solvent B for 32 minutes, 60% solvent B for 35 minutes, 10% solvent B for 35.1 minutes, and 10% solvent B for 40 minutes. Rutin and quercetin were quantified by comparing the peak area of the sample with that of the representative standard compounds. The content of rutin and quercetin was calculated using the following formula:
The values are expressed as milligrams per 100 gram dry weight (mg/100 g DW).
Statistical analysis
Data were analyzed using the
t-test at
P ≤ 0.05, with Statistix ver. 8.1 software (McGraw-
Hill 2008). Correlation analysis and principal component analysis (PCA) were performed with RStudio version 1.2.1335 (
RStudio Team 2019), using PerformaceAnalytics, FactoMineR, factoextra and, devtools, packages of R version 3.6.0 (
R Core Team 2019).
RESULTS
Evaluation of sprout growth traits
The morphological comparison of common buckwheat and tartary buckwheat is presented in
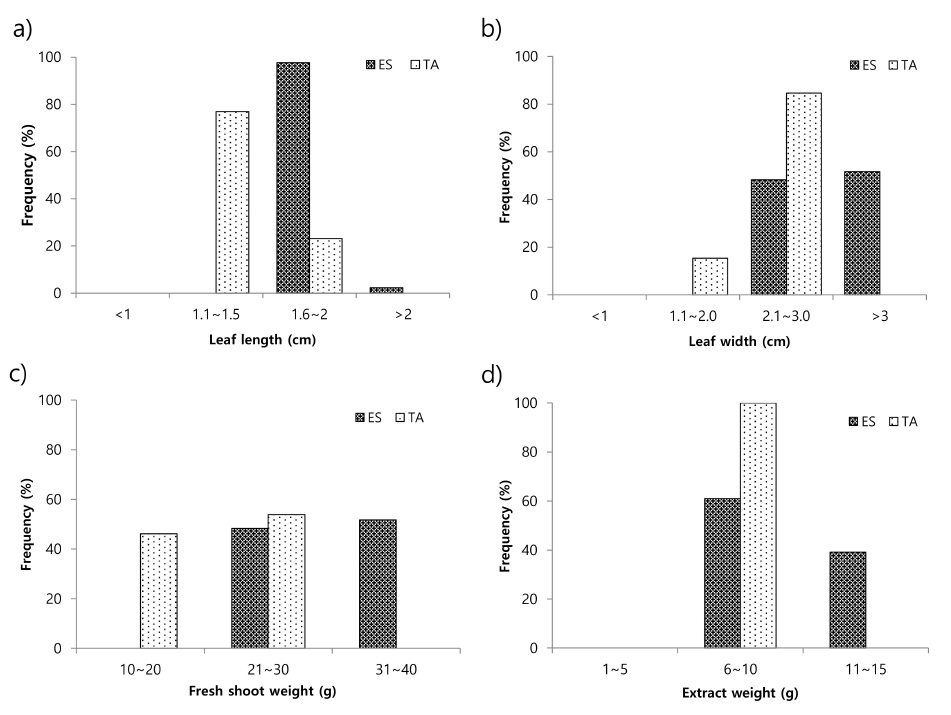
Table 2. The average leaf length (1.80 cm) of common buckwheat germplasm was significantly higher than that (1.51 cm) of tartary buckwheat germplasm. The leaf length of common buckwheat germplasm ranged from 1.6 to 2.1 cm, while that of tartary buckwheat germplasm ranged from 1.3 to 1.7 cm. Most of the common buckwheat germplasm had leaf length in the category of “1.6-2.0 cm”, whereas most of the tartary buckwheat germplasms in the category of “1.1-1.5 cm” (
Fig. 1a). Similarly, the average leaf width (3.08 cm) of common buckwheat germplasm was significantly higher than that (2.16 cm) of tartary buckwheat germplasm. The leaf width of common buckwheat germplasm ranged from 2.6 to 3.6 cm, whereas that of tartary buckwheat germplasm ranged from 1.9 to 2.3 cm. Moreover, we found that most of the common buckwheat germplasm had leaf width of > 3 cm whereas, most of the tartary buckwheat germplasm had leaf width in the category of “2.1-3.0 cm” (
Fig. 1b). Furthermore, the average fresh shoot weight (30.75 g) of common buckwheat germplasm was significantly higher than that (20.74 g) of tartary buckwheat germplasm. In common buckwheat germplasm, the fresh shoot weights ranged from 22 to 38 g, whereas that of tartary buckwheat ranged from 17 to 23 g. In common buckwheat, 43% of the accessions had fresh shoot weight in the category of “21-30 g” and 57% had fresh shoot weight in the category of “31-40 g”. Similarly, in tartary buckwheat germplasm, 44% of the germplasm had fresh shoot weight in the category of “10-20 g” and 56% germplasm had fresh shoot weight in the category of “21-30 g” (
Fig. 1c). The average extract weight (10.19 g) of common buckwheat germplasm was also significantly higher than that (7.12 g) of tartary buckwheat germplasm. In common buckwheat germplasm, the extract weight ranged from 6.3 to 12.3 g, whereas that of tartary buckwheat germplasm ranged from 6.4 to 8.8 g. Furthermore, 60% of the common buckwheat germplasms had extract weight in the category of “6-10 g” and 40% of the germplasm had extract weight in the category of “11-15 g”. However, in tartary buckwheat, all accessions had extract weight in the category of “6-10 g” (
Fig. 1d).
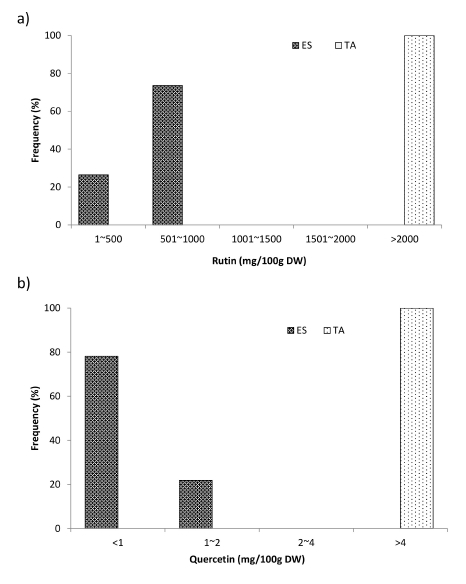
The results presented in
Table 2 revealed that the mean content of rutin (2729.9 mg/100 g DW) and quercetin (12.23 mg/100 g DW) was significantly higher in tartary buckwheat germplasm than in common buckwheat germplasm (553.5 mg/100 g DW and 0.78 mg/100 g DW, respectively). The content of rutin in common buckwheat sprouts ranged from 323.7 to 750.6 mg/100 g DW, whereas that in tartary buckwheat sprouts ranged from 2220.0 to 3185 mg/100 g DW. Similarly, the content of quercetin in common buckwheat sprouts ranged from 0.27 to 1.82 mg/ 100 g DW and that in tartary buckwheat sprouts ranged from 5.51 to 30.41 mg/100 g DW. In common buckwheat, 24% of the germplasm had rutin content in the category of “1-500 mg/100 g DW” and 76% of the germplasm had rutin content in the category of “501-1000 mg/100 g DW”. However, in tartary buckwheat, all accessions had rutin content in the category of “> 2000 mg/100 g DW” (
Fig. 2a). Similarly, 78% of the common buckwheat germplasm had quercetin content in the category of “< 1 mg/100 g DW” and 22% of the germplasm had quercetin content in the category of “1-2 mg/100 g DW”, whereas 100% of the tartary buckwheat germplasm had quercetin content in the category of “> 4 mg/100 g DW” (
Fig. 2b).
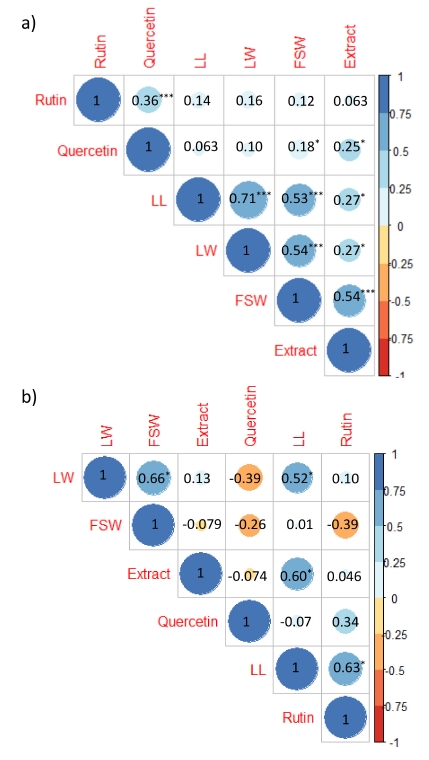
The correlation results of buckwheat sprouts revealed that there was a significant correlation between the leaf length, leaf width, fresh shoot weight, and extract weight. In addition, there was also a significant correlation between the content of rutin and quercetin. However, the leaf length, leaf width, fresh shoot weight, and extract weight showed a non-significant correlation with the rutin content. Similarly, the leaf length and leaf width also showed non-significant correlation with quercetin content; however, the fresh shoot weight and extract weight were significantly correlated with the quercetin content (
Fig. 3a). In tartary buckwheat sprouts, the leaf length showed a significant correlation with the leaf width, extract weight, and rutin content, and a non-significant correlation with the fresh shoot weight and quercetin content. Similarly, the leaf width showed a significant correlation with the fresh shoot weight and non-significant correlation with the extract weight, rutin content, and quercetin content. Similarly, the extract weight was non-significantly correlated with the rutin and quercetin content. In addition, there was no significant correlation between rutin and quercetin content (
Fig. 3b).
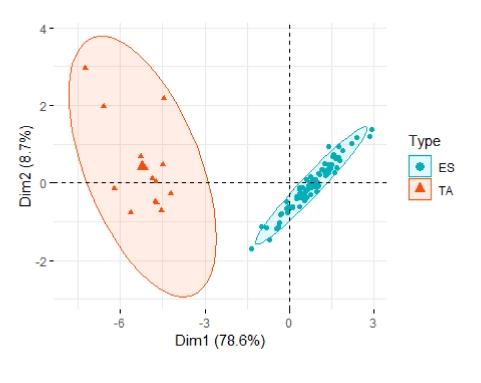
In the PCA, we observed that the maximum variation was explained by the first two principal components (PC1 and PC2), showing 78.61% and 8.71% of the total variation. The 3
rd, 4
th, 5
th and 6
th component of PCA exhibited 5.93%, 3.30%, 2.38%, and 1.05% of the total variability. The PCA biplot confirmed the high variability between species and within species, based on sprout growth traits and rutin and quercetin content (
Fig. 4). Moreover, all variables (loading > 0.385) contributed significantly to PC1. Analysis of loading (
Table 3) revealed that PC1 had the highest eigenvalue (4.72). The leaf width, fresh shoot weight, and rutin content were the main contributors of variability to PC1. The rutin and quercetin content was the major contributor and positive loading in PC2 and negative loading in PC1, and significantly correlated with each other.
Based on the content of rutin and quercetin, germplasms with high and low rutin and quercetin content have been identified from both species (
Table 4). In common buckwheat germplasm, five accessions (302328, 278145, 141448, 141447, and 100801) had the highest rutin content (> 700 mg/ 100 g DW) and three accessions (179846, 187867, and 288929) had the lowest rutin content (< 400 mg/100 g DW). Similarly, two accessions (302329 and 101282) had the highest quercetin content (> 1.5 mg/100 g DW) and six accessions (141458, 160602, 179843, 179846, 187869, and 288929) had the lowest content of quercetin (< 0.5 mg/ 100 g DW). Similarly, in tartary buckwheat germplasm, two accessions (226674 and 224676) had the highest rutin content (> 3000 mg/100 DW) and two accessions (301236 and 301238) had the lowest rutin content. Similarly, one accession (226673) had the highest content of quercetin (> 30 mg/100 DW) and five accessions (224676, 301235, 301236, 301240, and 301244) had the lowest content of quercetin (< 5 mg/100 g DW).
DISCUSSION
Buckwheat sprouts are excellent source of amino acids, minerals, and flavonoids; however, sprout growth and flavonoid content may vary among buckwheat germplasms. Regardless of the high nutritional value of buckwheat sprouts, there is limited research on the association between morphological diversity of sprouts and flavonoid content. In this study, we evaluated 87 accessions of common type and 13 tartary type buckwheat germplasms for sprout growth traits and rutin and quercetin content and their association. In our results, sprout growth traits of common buckwheat were significantly higher than those of tartary buckwheat. Our results are in good agreement with those of
Woo et al. (2018), who reported that the growth and developmental stages of common buckwheat and tartary buckwheat are significantly different. They further reported that the plant height, leaf size, dry weight, and fresh shoot weight of common buckwheat were significantly higher than those of tartary buckwheat.
Shin et al. (2010) also reported higher sprouts from common buckwheat seeds than that of sprouts from tartary buckwheat seeds. Common buckwheat contains larger seeds than that of tartary buckwheat. Seed size is positively correlated with seed vigor and larger seeds tend to do better in germination than small seeds do (
Ambika et al. 2014).
Moreover, in both species, the content of rutin and quercetin were in higher range than that of reported in buckwheat seeds. The content of flavonoids depends on the developmental stage, cultivation conditions, and cultivated species (
Park et al. 2000). Usually, buckwheat sprouts contain a significantly higher amounts of flavonoids including rutin, quercetin, and orientin, than those in buckwheat seeds (
Kim et al. 2004;
Sytar et al. 2016). Sprouting is a means of increasing polyphenolics as seeds undergo several physiological and morphological changes. The content of flavonoids increases gradually during the growth of seedling (
Kim et al. 2001); however, the number and concentration of phenolic components may differ according to the genotypes and growing conditions (
Park et al. 2000). A higher content of rutin and quercetin was found in the sprouts of common buckwheat grown under light conditions than that of sprouts grown under dark conditions (
Kim et al. 2006). It is thought that when the seeds are soaked, rutin in buckwheat hulls is transferred to the kernel (
Koyama et al. 2013), resulting in the increased content of flavonoids in germinating seeds. The content of flavonoids starts to increase with germination, and then decreases after the day 4 of germination (
Yiming et al. 2015). The content of rutin in sprouts significantly increased after 20 hours of germination, approximately 1.5 times higher than that in non-germinated buckwheat seeds (
Koyama et al. 2013). Moreover, environmental stresses also influence the content of flavonoids. Buckwheat sprouts subjected to environmental stresses such as drought, salt, and UV light are highly functional (
Yamanouchi et al. 2018). In our results, the average content of rutin and quercetin was 4.9 and 12.3 fold higher in tartary buckwheat sprouts than that of common buckwheat sprouts. Previously,
Kim et al. (2008) and
Liu et al. (2008) reported 4-5 fold higher rutin content in tartary buckwheat sprouts than that in common buckwheat.
Lee and Kim (2008) also reported that the rutin content (4.71 mg/100 g DW) in buckwheat grouts increased to 1524 mg/100 g DW in buckwheat sprouts.
Kim et al. (2005) reported 343.76 mg/100 g DW of rutin in buckwheat sprouts, which is 18 times higher than that in buckwheat seeds.
In order to investigate the relationship between sprout growth traits and rutin and quercetin content, we performed a correlation analysis. Correlation is a statistical method for investigating the strength of a relationship among quantitative traits, and a high correlation value between two traits indicates that the variables have a strong relationship with each other and a low value indicates that the variables have a weak relationship with each other (
Franzese and Luliano 2019). In our results we did not find any strong relationship between sprout growth traits of common buckwheat germplasm; however, leaf length and rutin content of tartary buckwheat showed a considerable relationship. In addition, the extract weight was non-significantly correlated with the rutin and quercetin content. This information could be helpful for plant breeders in selecting useful morphological traits for crop improvement.
Moreover, PCA has been suggested as a significant tool for plant breeders in order to group the diverse germplasms and evaluate the relationship among breeding materials, and to identify genetic diversity for crop improvement. We performed PCA to distinguish buckwheat germplasm based on type, sprout growth, and rutin and quercetin content. The first principal component, with the highest variability, showed an eigenvalue greater than 1. In general, the principal components whose eigenvalue is greater than 1 are considered significant. The components with higher eigen value have a greater variability than that of components whose eigenvalue is less than 1 (
Girden 2001). The leaf width, fresh shoot weight, and rutin content were the main contributors of variability to PC1. The variability in tartary buckwheat germplasm was higher than that in common buckwheat germplasm. This could be due to the wider range of rutin and quercetin content in tartary buckwheat germplasm. The genotypic difference is one of the key factors that influence the content of flavonoids. Earlier, serval scientists have reported variation among genotypes for particular traits by PCA in buckwheat and many other plants (
Yao et al. 2007;
Hussain et al. 2016;
Assefa et al. 2018;
Kapoor et al. 2018). These results could provide useful information for researcher to select variable traits for improving the nutritional value of high-yielding locally cultivated buckwheat cultivars.
Hence, we conclude that the difference in rutin and quercetin content in common and tartary buckwheat sprouts is not associated with morphological traits, but could be due to genetic difference and expression of flavonoid biosynthesis-related genes. In the future, comprehensive research is needed to evaluate diverse germplasms for diversity in the flavonoid content in sprouts. Moreover, extensive research is needed to compare the differential expression of flavonoid biosynthesis-related genes in both species. These selected accessions could be further used to investigate the genetic basis of rutin and quercetin content in common and tartary buckwheat using RNA-Seq analysis. Additionally, sprouts of the identified accessions with higher content of rutin and quercetin can be used as a dietary source of phenolic components to reduce the risk of various chronic diseases.
ACKNOWLEDGEMENTS
This study was supported by the “Research Program for Agricultural Science & Technology Development (Project No. PJ012478)” of the National Institute of Agricultural Sciences, Rural Development Administration.
Fig. 1Frequency distribution of a) leaf length, b) leaf width, c) fresh shoot weight (g), and d) extract weight in common and tartary buckwheat germplasm. ES: Fagopyrum esculentum, TA: Fagopyrum tataricum.
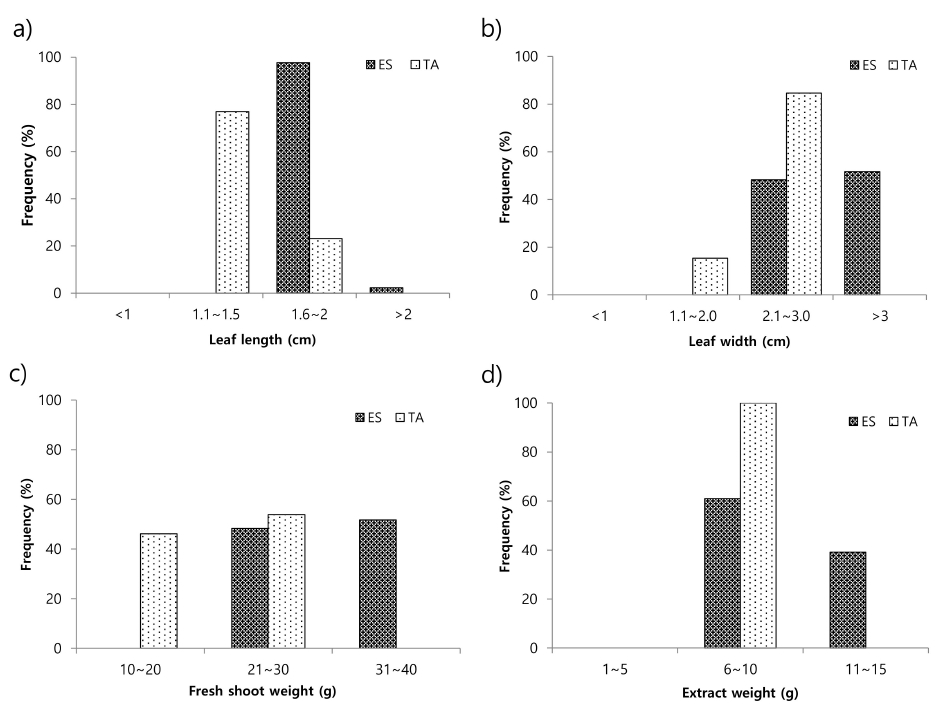
Fig. 2Frequency distribution of a) rutin and b) quercetin content in common and tartary buckwheat germplasm. ES: Fagopyrum esculentum, TA: Fagopyrum tataricum.
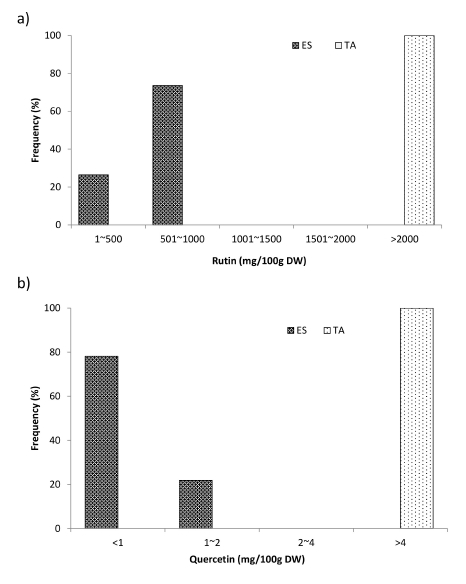
Fig. 3Correlation analysis between sprout growth traits and rutin and quercetin content in a) common buckwheat and b) tartary buckwheat germplasm. LL: leaf length, LW: leaf width, FSH: fresh shoot weight.
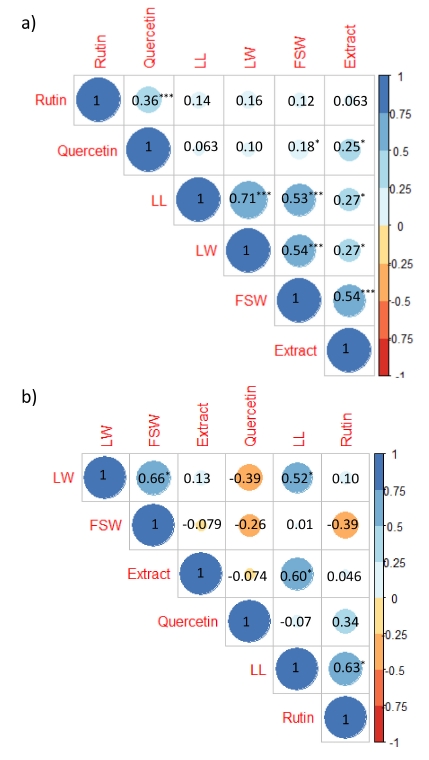
Fig. 4Principal component analysis of buckwheat germplasm for sprout growth traits and rutin and quercetin content. ES: Fagopyrum esculentum, TA: Fagopyrum tataricum.
Table 1List of common and tartary buckwheat accessions used to study sprout growth traits and rutin and quercetin content.
Table 1
|
S. No. |
Accessions IT |
Type |
S. No. |
Accessions IT |
Type |
S. No. |
Accessions IT |
Type |
S. No. |
Accessions IT |
Type |
|
1 |
28843 |
ESZ)
|
26 |
141444 |
ES |
51 |
189663 |
ES |
76 |
278145 |
ES |
|
2 |
28844 |
ES |
27 |
141447 |
ES |
52 |
194511 |
ES |
77 |
288928 |
ES |
|
3 |
100801 |
ES |
28 |
141448 |
ES |
53 |
199279 |
ES |
78 |
288929 |
ES |
|
4 |
101006 |
ES |
29 |
141450 |
ES |
54 |
199280 |
ES |
79 |
288931 |
ES |
|
5 |
101022 |
ES |
30 |
141453 |
ES |
55 |
199281 |
ES |
80 |
289118 |
ES |
|
6 |
101091 |
ES |
31 |
141458 |
ES |
56 |
199282 |
ES |
81 |
301213 |
ES |
|
7 |
101271 |
ES |
32 |
148424 |
ES |
57 |
199286 |
ES |
82 |
301231 |
ES |
|
8 |
103026 |
ES |
33 |
148427 |
ES |
58 |
200686 |
ES |
83 |
302264 |
ES |
|
9 |
103633 |
ES |
34 |
148429 |
ES |
59 |
208852 |
ES |
84 |
302327 |
ES |
|
10 |
104461 |
ES |
35 |
158263 |
ES |
60 |
210198 |
ES |
85 |
302328 |
ES |
|
11 |
105453 |
ES |
36 |
160602 |
ES |
61 |
220413 |
ES |
86 |
302329 |
ES |
|
12 |
105506 |
ES |
37 |
175920 |
ES |
62 |
220676 |
ES |
87 |
310514 |
ES |
|
13 |
105543 |
ES |
38 |
175975 |
ES |
63 |
225054 |
ES |
88 |
201753 |
TA |
|
14 |
105649 |
ES |
39 |
178415 |
ES |
64 |
225055 |
ES |
89 |
224676 |
TA |
|
15 |
105699 |
ES |
40 |
178417 |
ES |
65 |
250614 |
ES |
90 |
225088 |
TA |
|
16 |
105997 |
ES |
41 |
179841 |
ES |
66 |
250617 |
ES |
91 |
226673 |
TA |
|
17 |
108852 |
ES |
42 |
179843 |
ES |
67 |
250622 |
ES |
92 |
226674 |
TA |
|
18 |
108885 |
ES |
43 |
179844 |
ES |
68 |
250624 |
ES |
93 |
226675 |
TA |
|
19 |
108889 |
ES |
44 |
179846 |
ES |
69 |
250626 |
ES |
94 |
301235 |
TA |
|
20 |
108892 |
ES |
45 |
185685 |
ES |
70 |
101282 |
ES |
95 |
301236 |
TA |
|
21 |
108934 |
ES |
46 |
185686 |
ES |
71 |
261919 |
ES |
96 |
301238 |
TA |
|
22 |
109175 |
ES |
47 |
185704 |
ES |
72 |
261921 |
ES |
97 |
301240 |
TA |
|
23 |
121461 |
ES |
48 |
187867 |
ES |
73 |
261923 |
ES |
98 |
301241 |
TA |
|
24 |
134978 |
ES |
49 |
187869 |
ES |
74 |
261925 |
ES |
99 |
301242 |
TA |
|
25 |
138108 |
ES |
50 |
187871 |
ES |
75 |
278144 |
ES |
100 |
301244 |
TA |
Table 2Means and ranges of sprout growth traits and rutin and quercetin content in common and tartary buckwheat germplasm.
Table 2
|
Traits |
Common buckwheat |
Tartary buckwheat |
|
|
|
|
Mean |
Range |
Mean |
Range |
|
Leaf length (cm) |
1.80 Az) ± 0.09 |
1.6-2.1 |
1.51 B ± 0.09 |
1.3-1.7 |
|
Leaf width (cm) |
3.08 A ± 0.16 |
2.6-3.6 |
2.16 B ± 0.12 |
1.9-2.3 |
|
Fresh shoot weight (g) |
30.75 A ± 3.14 |
22.0-38.0 |
20.74 B ± 2.02 |
17-23 |
|
Extract weight (g) |
10.19 A ± 1.0 |
6.3-12.3 |
7.12 B ± 0.78 |
6.4-8.8 |
|
Rutin (mg/100 g DW) |
553.5 B ± 86.27 |
332.7-750.6 |
2729.9 A ± 274.14 |
2220-3185 |
|
Quercetin (mg/100 g DW) |
0.78 B ± 0.29 |
0.27-1.82 |
12.23 A ± 7.37 |
4.76-30.41 |
Table 3Loadings, eigenvalue and % variance of buckwheat germplasm for the diversity in sprout growth traits and the content of rutin and quercetin.
Table 3
|
|
PC 1 |
PC 2 |
PC 3 |
PC 4 |
PC 5 |
PC 6 |
|
Leaf length |
0.398 |
0.393 |
0.593 |
−0.382 |
0.253 |
−0.355 |
|
Leaf width |
0.441 |
0.010 |
0.298 |
0.002 |
−0.287 |
0.797 |
|
Fresh shoot weight |
0.411 |
0.347 |
−0.117 |
0.814 |
0.100 |
−0.154 |
|
Extract weight |
0.389 |
0.322 |
−0.737 |
−0.433 |
0.083 |
0.087 |
|
Rutin |
−0.422 |
0.391 |
0.045 |
0.056 |
0.676 |
0.455 |
|
Quercetin |
−0.385 |
0.685 |
0.036 |
−0.022 |
−0.616 |
−0.031 |
|
Eigenvalue |
4.72 |
0.52 |
0.36 |
0.20 |
0.14 |
0.06 |
|
% variance |
78.62 |
8.71 |
5.93 |
3.30 |
2.38 |
1.05 |
Table 4Selected germplasm with the highest and lowest content of rutin and quercetin.
Table 4
|
Species |
Flavonoids |
Description |
Accessions IT |
|
Common buckwheat |
Rutin |
Highest (> 700 mg/100 g DW) |
302328, 278145, 141448, 141447, 100801 |
|
Lowest (< 400 mg/100 g DW) |
179846, 187867, 288929 |
|
Quercetin |
Highest (> 1.5 mg/100 g DW) |
302329, 101282 |
|
Lowest (< 0.5 mg/100 g DW) |
179846, 288929, 141458, 187869, 160602, 179843 |
|
Tartary buckwheat |
Rutin |
Highest (> 3000 mg/100 g DW) |
226674, 224676 |
|
Lowest (< 2500 mg/100 g DW) |
301236, 301238 |
|
Quercetin |
Highest (> 30 mg/100 g DW) |
226673 |
|
Lowest (< 5 mg/100 g DW) |
301240, 301235, 224676, 301236, 301244 |
References
Reference
- Ahmed A, Khalid N, Ahmad A, Abbasi NA, Latif MSZ, Randhawa MA. 2013. Phytochemical and biofunctional properties of buckwheat: a review. J. Agric. Sci.. 152: 349-369.
- Akande KE, Doma UD, Agu HO, Adamu HM. 2010. Major antinutrients found in plant protein sources: their effect on nutrition. Pak. J. Nutr.. 9: 827-832.
- Ambika S, Manonmani V, Somasundaram G. 2014. Review on effect of seed size on seedling vigour and seed yield. Res. J. Seed Sci.. 7: 31-38.
- Assefa , AD , Jeong JI, Kim DJ, Jeon YA, Lee JR, Ko HC, et al. 2008. Assessing phenolic content and antioxidant potential diversity in Allium plants using multivariate data analysis. Hort. Environ. Biote.. 59: 759-773.
- Bonafaccia G, Fabjan N. 2003. Nutritional comparison of tartary buckwheat with common buckwheat and minor cereals. Zb. Bioteh. Fak. Univ. Ljublj. Kmet.. 81: 349-355.
- Brajdes C, Vizireanu C. 2012. Sprouted buckwheat an important vegetable source of antioxidants. Annals of the University Dunarea de Jos Galati Fascicli VI -Food Technology. 36: 53-60.
- Chao PL, Hsiu S, Hou Y. 2002. Flavonoids in herbs: biological fates and potential interactions with xenobiotics. J. Food Drug Anal.. 10: 219-228.
- Chauhan RS, Gupta N, Sharma SK, Rana JC, Sharma TR, Jana S. 2010. Genetic and genome resources in buckwheat -present status and future perspectives. Eur. J. Plant Sci. Biotechnol.. 4: 33-44.
- Chen QF, Huang XY, Li HY, Yang LJ, Cui YS. 2018. Recent progress in perennial buckwheat development. Sustainability. 10: 536
- Cook NC, Samman S. 1996. Flavonoids-Chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem.. 7: 66-76.
- Franzese M, Luliano A. Ranganathan S., Gribskov M., Nakai K., Schonbach C., 2019. Correlation analysis. editors. Encyclopedia of bioinformatics and computational biology. Academic Press. pp. 706-721.
- Girden ER. 2001. Evaluating research articles from start to finish. Sage Publications. Thousand Oaks, California, U.S.A..
- Hertog MGL, Kromhout D, Aravanis C, Blackburn H, Buzina R, Fidanza F, et al. 1995. Flavonoid intake and long-term risk of coronary heart disease and cancer in seven countries study. Arch. Intern. Med.. 155: 381-386.
- Huang XY, Zeller FJ, Hung KF, Shi TX, Chen QF. 2014. Variation of major minerals and trace elements in seeds of tartary buckwheat (Fagopyrum tataricum Gaertn.). Genet. Genet. Resour. Crop Evol.. 61: 567-577.
- Hussain I, Bano A, Faizanullah , Nosheen A. 2016. Multivariate analysis for elemental composition among indigenous common buckwheat genotypes of Baltistan. J. Anim. Plant Sci.. 26: 1725-1731.
- Jiang P, Burczynski F, Campbell C, Pierce G, Austria JA, Briggs CJ. 2007. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F. tataricum and F. homotropicum and their protective effects against lipid peroxidation. Food Res. Int.. 40: 356-364.
- Joshi BD, Rana RS. Williams JT., 1995. Buckwheat (Fogopyrum esculentum). editor. Cereals and Pseudocereals. Champman and Hall. New York, U.S.A.: pp. 85-127.
- Kapoor C, Avasthe RK, Chettri PK, Gopi R, Kalita H, Rana JC. 2018. Multivariate analysis to evaluate common and Tartary buckwheat germplasm. Indian J. Plant Genet. Resour.. 31: 134-141.
- Kim SL, Son YK, Hwang JJ. 2001. Development and utilization of buckwheat sprouts as a functional vegetables. Proc. of the 8th ISB. pp. 653-660.
- Kim SJ, Kim SK, Park CH. 2004. Introduction and nutritional evaluation of buckwheat sprouts as a new vegetable. Food Res. Int.. 37: 319-327.
- Kim YS, Kim JG, Lee YS, Kang IJ. 2005. Comparison of the chemical components of buckwheat seed and sprout. Korean J. Food Nutr.. 34: 81-86.
- Kim SJ, Kawaharada C, Suzuki T, Saito K, Hashimoto N, Takigawa S, et al. 2006. Effect of natural light periods on rutin, free amino acid and vitamin C contents in the sprouts of common (Fagopyrum esculentum Moench) and tartary (F. tataricum Gaertn.) Buckwheats. Food Sci. Technol. Res.. 12: 199-205.
- Kim SJ, Zaidul ISM, Suzuki T, Mukasa Y, Hashimoto N, Takigawa S, et al. 2008. Comparison of phenolic compositions between common and tartary buckwheat (Fagopyrum) sprouts. Food Chem.. 110: 814-820.
- Koyama M, Nakamura C, Nakamura K. 2013. Changes in phenols contents from buckwheat sprouts during growth stage. J. Food Sci. Technol.. 50: 86-93.
- Lee EH, Kim CJ. 2008. Nutritional changes of buckwheat during germination. J. Korean Soc. Food Cult.. 23: 121-129.
- Li J, Gong F, Li F. 2016. Hypoglycemic and hypolipidemic effects of flavonoids from tartary buckwheat in type 2 diabetic rats. Biomed. Res.. 27: 132-137.
- Li SQ, Zhang QH. 2001. Advances in the development of functional foods from Buckwheat. Crit. Rev. Food Sci. Nutr.. 41: 451-464.
- Liu CL, Chen YS, Yang JH, Chiang BH. 2008. Antioxidant activity of tartary (Fagopyrum tataricum (L.) Gaertn.) and common (Fagopyrum esculentum Moench) buckwheat sprouts. J. Agric. Food Chem.. 56: 173-178.
- McGraw-Hill C. 2008. Statistix 8.1 (Analytical Software, Tallahassee, Florida). Maurice/Thomas. Analytical Software. Tallahassee, U.S.A..
- Nam TG, Lee SM, Park JH, Kim DO, Baek NI, Eom SH. 2015. Flavonoid analysis of buckwheat sprouts. Food Chem.. 170: 97-101.
- Nonogaki H. 2006. Seed germination -The biochemical and molecular mechanisms. Breed. Sci.. 56: 93-105.
- Park CH, Kim YB, Choi YS, Heo K, Kim SL, Lee KC, et al. 2000. Rutin content in food products processed from groats, leaves, and flowers of buckwheat. Fagopyrum. 17: 63-66.
- R Core. 2019. R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria.
- Ratan P, Kothiyal P. 2011.
Fagopyrum esculentum Moench (common buckwheat) edible plant of Himalayas: A Review. Asian J. Pharm. Life Sci.. 1: 426-442.
- RStudio Team. 2019. RStudio: Integrated development for R. RStudio, Inc.. Boston, Massachusetts, U.S.A..
- Shin DH, Kamal AHM, Suzuki T, Yun YH, Lee MS, Chung KY, et al. 2010. Reference proteome map of buckwheat (Fagopyrum esculentum and Fagopyrum tataricum) leaf and stem cultured under light or dark. Aust. J. Crop Sci.. 4: 633-641.
- Sytar O, Bresticb M, Zivcak M, Tran LSP. 2016. The contribution of buckwheat genetic resources to health and dietary diversity. Curr. Genomics. 17: 193-206.
- Welbaum GE, Bradford KJ, Yim KO, Both DT, Oluoch MO. 1998. Biophysical, physiological and biochemical processes regulating seed germination. Seed Sci. Res.. 8: 161-172.
- Wiczkowski W, Szawara-nowak D, Debski H, Mitrus J, Horbowicz M. 2014. Comparison of flavonoids profile in sprouts of common buckwheat cultivars and wild tartary buckwheat. J. Food Sci. Technol.. 49: 1977-1984.
- Woo SH, Roy SK, Kwon SJ, Kamal AHM, Park SU, Chung KY, et al. M. Zhou, I. Kraft, G. Suvorova, Y. Tang, , 2018. Distribution of amino acid in buckwheat. editors. Buckwheat germplasm in the world. Academic press. pp. 295-302.
- Yamanouchi K, Sakamoto Y, Tsuiguchi T. 2018. Contemporary traditional vegetables in Japan: Physiological function of buckwheat sprouts. J. Nutr. Food Sci.. 8: 4
- Yao Y, Xuana Z, Hec Y, Lutts S, Korpelainen H, Li C. 2007. Principal component analysis of intraspecific responses of tartary buckwheat to UV-B radiation under field conditions. Environ. Exper. Bot.. 61: 237-245.
- Yiming Z, Hong W, Linlin C, Xialoi Z, Wen T, Xinli S. 2015. Evolution of nutrient ingredients in tartary buckwheat seeds during germination. Food Chem.. 186: 244-248.
- Zielińska D, Turemko M, Kwiatkowski J, Zieliński H. 2012. Evaluation of flavonoid contents and antioxidant capacity of the aerial parts of common and tartary buckwheat plants. Molecules. 17: 9668-9682.